
Abstract
In vitro vascularization is an upcoming strategy to solve the problem of insufficient blood supply after implantation. Although recent publications show promising results, these studies were generally performed with clinically irrelevant endothelial cell model systems. We tested the use of endothelial progenitor cells (EPC) obtained from umbilical cord blood and human mesenchymal stem cells (hMSC) from the bone marrow for their use in a prevascularized bone tissue engineering setting. MSC were differentiated toward endothelial cells. They formed capillary-like structures containing lumen, stained positive for CD31, attained the ability to take up acetylated low-density lipoproteins, and formed perfused vessels in vivo. However, in a three-dimensional coculture setting with undifferentiated hMSC, the cells stopped expressing CD31 and did not form prevascular structures. EPC from the cord blood were able to form prevascular structures in the same coculture setting, but only when the state of endothelial differentiation was mature. The amount of prevascular structures formed when using EPC was less than when human umbilical vein endothelial cells or human dermal microvascular endothelial cells were used. The degree of organization, however, was higher. We conclude that EPC can be used for complex tissue engineering applications, but the differentiation stage of these cells is important.
Introduction
In the field of bone tissue engineering, bone implant materials like ceramics are combined with osteoprogenitor cells. Human mesenchymal stem cells (hMSC) are commonly used for this purpose. These pluripotent cells can be isolated from the bone marrow and have the ability to differentiate into adipogenic, chondrogenic, and osteogenic lineages. 4 Differentiation toward osteoprogenitor cells can be achieved by stimulation with for instance BMP-2 or dexamethasone.5–7 Numerous groups, including our own, have shown that the combination of artificial scaffolds and osteoprogenitor cells can lead to the formation of new bone in both ectopic and orthotopic sites.8–12 However, the effect of the use of this technique in clinical bone defects is still unpredictable. 13 The limited clinical success may be explained by a lack of vascularization. Vascularization is vital for the survival of the implanted cells on the carrier material after implantation. Apart from that, vascularization is a critical process during bone growth and repair. Studies have shown that fracture healing and ectopic new bone formation can be blocked by the administration of angiogenesis inhibitors,14,15 while others have shown that new bone formation in porous scaffolds was significantly increased by the insertion of a vascular pedicle in the scaffold.16,17
Previous studies have shown that endothelial cells can form vascular structures in vitro18–20 that can anastomose to the vasculature of the host after implantation,21,22 Moreover, prevascularization of a tissue can enhance the implant perfusion and survival. 22 We previously reported on the coculture of hMSC and human umbilical vein endothelial cells (HUVEC) in a spheroid coculture system. 23 This resulted in the formation of osteogenic tissue containing a prevascular network, which can be used for modular bone tissue engineering. The aforementioned studies, however, were performed, in general, with nonclinically applicable endothelial cell sources like HUVEC. If one wants to implement in vitro prevascularization in clinical applications, one should be able to use an endothelial cell source that can be readily isolated from adult patients in sufficient numbers in an acceptable timeframe. Here, we tested the potential use of endothelial progenitor cells (EPC) isolated from blood and differentiated from MSC in comparison with both microvascular and macrovascular mature endothelial cells.
The goal of the current study was twofold: (1) to differentiate hMSC toward E(P)C and (2) to investigate the potential role of these cells and EPC from the blood for prevascularized bone tissue engineering. We investigate whether these cells are capable of forming or inducing three-dimensional (3D) prevascular networks in a coculture with undifferentiated hMSC. If EPC in coculture with hMSC do indeed form a prevascular network, they would be an easily accessible and practical cell source for prevascularized bone tissue engineering.
Materials and Methods
Culture of HUVEC and human dermal microvascular endothelial cells
HUVEC and human dermal microvascular endothelial cells (HMVEC) were purchased from Lonza (Basel, Switzerland). Cells were grown at 37°C in a humid atmosphere with 5% carbon dioxide (CO2) in endothelial growth medium-2 (EGM-2; Lonza). Cells were routinely split at a 1:5 ratio and cultured <5 passages. Cells from only passage 3 or 4 were used to seed the coculture experiments.
Isolation and culture of hMSC
Bone marrow aspirates were obtained from three donors, aged 27, 43, and 65, with written informed consent. Cells were isolated as published before. 24 In short, aspirates were resuspended using a 20-G needle and plated at a density of 5 × 105 mononucleated cells/cm2. Cells were grown in MSC proliferation medium (minimal essential medium [α-MEM; Invitrogen, Carlsbad, CA] supplemented with 10% fetal bovine serum [FBS; Cambrex, East Rutherford, NJ], 0.2 mM ascorbic acid [AsAP; Sigma, St. Louis, MO], 2 mM L-glutamine [Invitrogen], 100 U/mL penicillin [Invitrogen], 10 μg/mL streptomycin [Invitrogen], and 1 ng/mL basic fibroblast growth factor [bFGF; Instruchemie, Delfzijl, The Netherlands]) at 37°C in a humid atmosphere with 5% CO2. Cells were characterized by flow cytometry and tested >90% positive for CD29, CD44, CD105, and CD166. Cells tested negative for CD31. Cells were routinely split at a 1:5 ratio and cultured <5 passages. MSC from passage 3 or 4 were used to seed the coculture experiments. The results in this paper were similar for all three donors.
Endothelial induction of hMSC
MSC from passage 4 were used for the endothelial induction assays. Cells were seeded on tissue culture plastic and cultured in MSC proliferation medium without bFGF supplemented with 50 ng/mL vascular endothelial growth factor (hrVEGF165) (Sigma) for 10 days. Cells were split at a 1:6 ratio at subconfluence to assess the effect of growth factor stimulation on cell differentiation. For induction on Matrigel, the wells of a six-well plate were coated with 0.9 mL growth factor–reduced Matrigel (BD Biosciences, Erembodegem, Belgium) per well. Cells were seeded at a density of 2 × 104 cells per well. Cells were grown in EGM-2 at 37°C in a humid atmosphere with 5% CO2. The formation of capillary-like structures was followed over time using an inverted microscope (Nikon Eclipse TE300; Tokyo, Japan). At day 10, pieces of Matrigel were frozen in Cryomatrix (Thermo Shandon, Waltham, MA) at −60°C. Cross sections (6 μm) were cut with a cryotome and stained with hematoxylin (Sigma). For immunohistochemical analysis, hMSC were seeded on Matrigel-coated cover slips instead of six-well plates. MSC induced on Matrigel for 10 days were used for coculture experiments (endothelial-induced hMSC [EC-MSC])
Acetylated low-density lipoproteins uptake assay
MSC that were cultured on Matrigel in EGM-2 for 9 days were removed by incubating in a 1:1 mixture of 0.25% trypsin and dispase for 15 min to prevent background labeling of the gel. Recovered cells were seeded on cover slips and cultured in EGM-2 for 1 more day. Subsequently, cells were incubated in EGM-2 supplemented with 10 μg/mL DiI-labeled acetylated low-density lipoproteins (DiI-acLDL; Invitrogen) for 4 h at 37°C in a humid atmosphere with 5% CO2. Finally, cells were washed with phosphate-buffered saline (PBS) three times for 5 min. The labeling of cells was subsequently analyzed using a Nikon Eclipse E600 fluorescent microscope.
In vivo assay
MSC were cultured on Matrigel in EGM-2 for 14 days to allow them to organize into capillary-like structures. Four pieces of Matrigel (approximately 4 × 4 × 2 mm) containing these structures were then implanted subcutaneously in the dorsal region of two male nude mice. The mice were anesthetized with 2.5% isoflurane, after which the samples were implanted in separate pockets. Two weeks after implantation, the animals were euthanized and the implants were retrieved. Samples were snap-frozen in Cryomatrix.
Isolation and culture of human EPC
After obtaining signed informed consent from the mother, umbilical cord blood was collected from the clamped umbilical cord section attached to the placenta normally discarded after birth. Only blood from healthy newborns after uncomplicated deliveries was used, all at the Oudenrijn Hospital in Utrecht, the Netherlands. For the experiments described in this study, between 30 and 90 mL per donor had been obtained and used for culture. Mononuclear cells (MNC) were isolated from umbilical cord blood by Ficoll density gradient separation (Histopaque 1077; Sigma). MNC were plated on gelatin (Sigma)–coated six-well plates at a density of 107 cells per well in induction medium (M199 medium [Invitrogen] containing 20% fetal calf serum [Invitrogen], 0.05 mg/mL bovine pituitary extract [Invitrogen], 10 U/mL heparin [Leo Pharma, Breda, The Netherlands], and antibiotics [penicillin 100 U/mL and streptomycin 100 μg/mL; Invitrogen]). Medium was changed after 4 days, washing nonadherent cells away. After 7 days cells had adopted a spindle-shape morphology and proliferate minimally low-proliferative spindle-shaped EPC (LP-SS-EPC).
Subsequently, medium was changed to differentiation medium (EGM-2 medium, supplemented with 20% fetal calf serum [Invitrogen] and antibiotics [penicillin 100 U/mL and streptomycin 100 μg/mL; Invitrogen]), facilitating differentiation toward high-proliferative cobblestone-shaped EPC (HP-CS-EPC). Twice weekly, half of the medium was removed and replaced with fresh medium. Upon reaching confluence, HP-CS-EPC were passaged onto fibronectin (Becton Dickinson, Franklin Lakes, NJ)–coated wells in regular EGM-2 medium, which was used for further culture. The total culture period for obtaining HP-CS-EPC was 4 weeks on average. LP-SS-EPC cultured for a similar period of time without switching to differentiation medium retained the spindle-shape morphology. These cells, late outgrowth LP-SS-EPC, were also used for coculture experiments.
EPC characterization
EPC were evaluated for the ability to bind FITC-labeled Ulex Europeus Lectin-1 (UEA-1; Vector Laboratories, Burlingame, CA) and to take up DiI-acLDL (Invitrogen). For this, EPC cultured on fibronectin-coated coverslips were washed in PBS and placed in EGM-2 with 1:100 diluted FITC-labeled UEA-1 for 1 h at 37°C, followed by a 2-h incubation in EBM-2 with 1:400 diluted DiI-acLDL at 37°C. Finally, cell nuclei were stained with DAPI, and cover slips were mounted on glass slides using Vectashield (Vector Laboratories).
The presence of CD31 and von Willebrand factor (vWF) was identified in EPC cultured on coverslips, fixed in 4% paraformaldehyde, and permeabilized (only for vWF staining) using 0.1% saponin in PBS. Coverslips were incubated with anti-CD31 or anti-vWF antibody (Dako, Glostrup, Denmark), followed by an FITC-labeled goat-anti-mouse secondary antibody (BD Pharmingen, Erembodegem, Belgium) and DAPI. Isotype-stained sections served as controls. Staining was evaluated by fluorescence microscopy using a Leica DMR microscope (Leica, Wetzlar, Germany).
Labeling of cells with 5(6)-carboxyfluorescein diacetate N-succinimidyl ester
Cells were detached using trypsin-EDTA and resuspended in PBS to a final concentration of 1 × 106 cells/mL with 10 μM 5(6)-carboxyfluorescein diacetate N-succinimidyl ester (CFSE) (Sigma). After 10-min incubation at 37°C, cells were washed in PBS, resuspended in 5 mL of medium, and again incubated for 5 min at 37°C to allow excessive CFSE to wash out. Cells were washed with PBS twice before being used for further experiments.
Generation and culture of coculture spheroids
Coculture spheroids were prepared as described before. 23 In brief, different percentages of CFSE-prelabeled endothelial cells (being in different samples HUVEC, HMVEC, EC-MSC, early outgrowth LP-SS-EPC, late outgrowth LP-SS-EPC, or HP-CS-EPC) and undifferentiated hMSC were pooled to a total of 5 × 105 cells in a round-bottom 10 mL tube (Greiner, Frickenhausen, Germany). The cells were re-suspended in 4.5 mL osteogenic differentiation medium (α-MEM supplemented with 10% FBS, 0.2 mM AsAP, 2 mM L-glutamine, 100 U/mL penicillin, 10 μg/mL streptomycin, 10−8 M dexamethasone [Sigma], and 0.01 M β-glycerophosphate [Sigma]) and consequently centrifuged at 1600 rpm for 2 min. The tubes with the cell pellets were incubated at 37°C in a humid atmosphere with 5% CO2, which allowed for the spontaneous formation of coculture spheroids. The spheroids were cultured for 10 days with a medium change at days 5 and 8.
Immunohistochemical analysis of spheroids
After harvesting, spheroids were frozen in Cryomatrix (Thermo Shandon) at −60°C. Sections (6 μm) were cut with a cryotome. Sections were fixed in cold acetone for 5 min and air-dried. Sections were rehydrated for 10 min, after which they were incubated for 30 min with 10% FBS in PBS to block nonspecific background staining. Sections were incubated with mouse-anti-human CD31 (does not cross react with mouse tissue) primary antibody (Dako) for 1 h. Sections were washed in PBS and subsequently incubated with the secondary antibody (horseradish peroxidase–conjugated goat-anti-mouse immunoglobulin antibody; Dako) for 45 min. Slides were developed with diaminobenzidine (DAB; Dako) as substrate, and weakly counterstained with hematoxylin (Sigma). For fluorescent staining, Alexa Fluor 594–conjugated goat-anti-mouse immunoglobulin antibody (Invitrogen) was used as the secondary antibody. These samples were neither developed nor counterstained. For in vivo samples, the Dako ARK kit was used to reduce background staining. To check the viability of cells in cocultures of hMSC and EPC, sections were stained using the DeadEnd colorimetric TUNEL system (Promega, Madison, WI) according to the supplier's protocol. Staining was evaluated using a Nikon Eclipse E600 microscope.
Flow cytometry
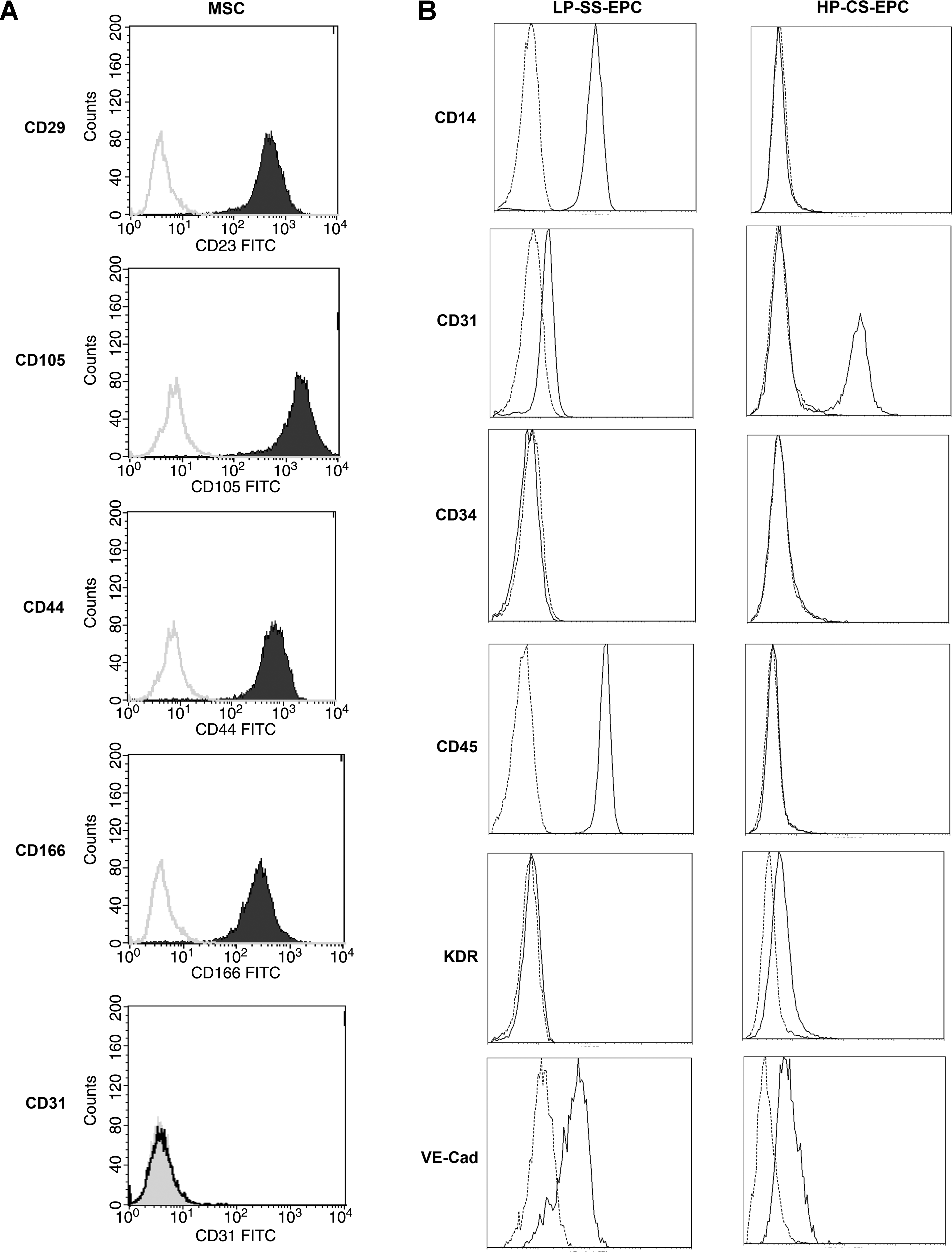
To evaluate the hMSC used in this study, the cells were analyzed using flow cytometry before endothelial differentiation. First, cells were incubated in block buffer containing control mouse antihuman IgG2a (Dako), anti-CD29 monoclonal antibody (Dako), anti-CD44 (Pharming, Leiden, The Netherlands) antibody, anti-CD105 (Dako) antibody, anti-CD166 (RDI, Concord, MA) monoclonal antibody, or anti-human CD31 (Dako) antibody. The cells were incubated on ice for 45 min and washed three times with PBS containing 1% BSA and 0.05% NaN3 (wash buffer). Incubation with the secondary antibody, goat anti-mouse-IgG–fluorescein isothiocyanate (Dako), was performed for 45 min on ice. After the hMSC were washed three times with wash buffer, the cells were incubated for 10 min with Viaprobe (Becton Dickinson) for live–dead staining. The cell suspension was analyzed using a flow cytometer (Becton Dickinson). The data were analyzed via Cell Quest software (Becton Dickinson).
Apart from that, both LP-SS-EPC and HP-CS-EPC were analyzed using flow cytometry. Cells were stained with FITC-conjugated CD31 (Serotec, Oxford, UK), FITC-conjugated CD34 (BD Biosciences), PE-conjugated KDR (R&D Systems, Minneapolis, MN), PE-Cy5–conjugated CD14 (Serotec), PE-Cy7–conjugated CD45 (BD Biosciences), or appropriate isotype controls. Flow cytometry was performed on a Beckman Coulter FC500 flow cytometer (Beckman Coulter, Fullerton, CA) and analyzed using the manufacturer's software.
Image analysis
Cross sections were made from the middle of the spheroids. Images of the entire surface of cross sections were taken at a magnification of 100×, unless otherwise stated (Nikon Eclipse E600). Images were combined to make a single image of the entire cross section of each sample. Subsequently, CD31-positive areas were pseudo colored and Bioquant image analysis software (Bioquant Image Analysis, Nashville, TN) was used to determine the percentage of the cross section that stained positive for CD31. For each group, cross sections of four separate spheroids were analyzed. Statistical analysis was performed using the Student's t-test.
Results
Endothelial induction of hMSC
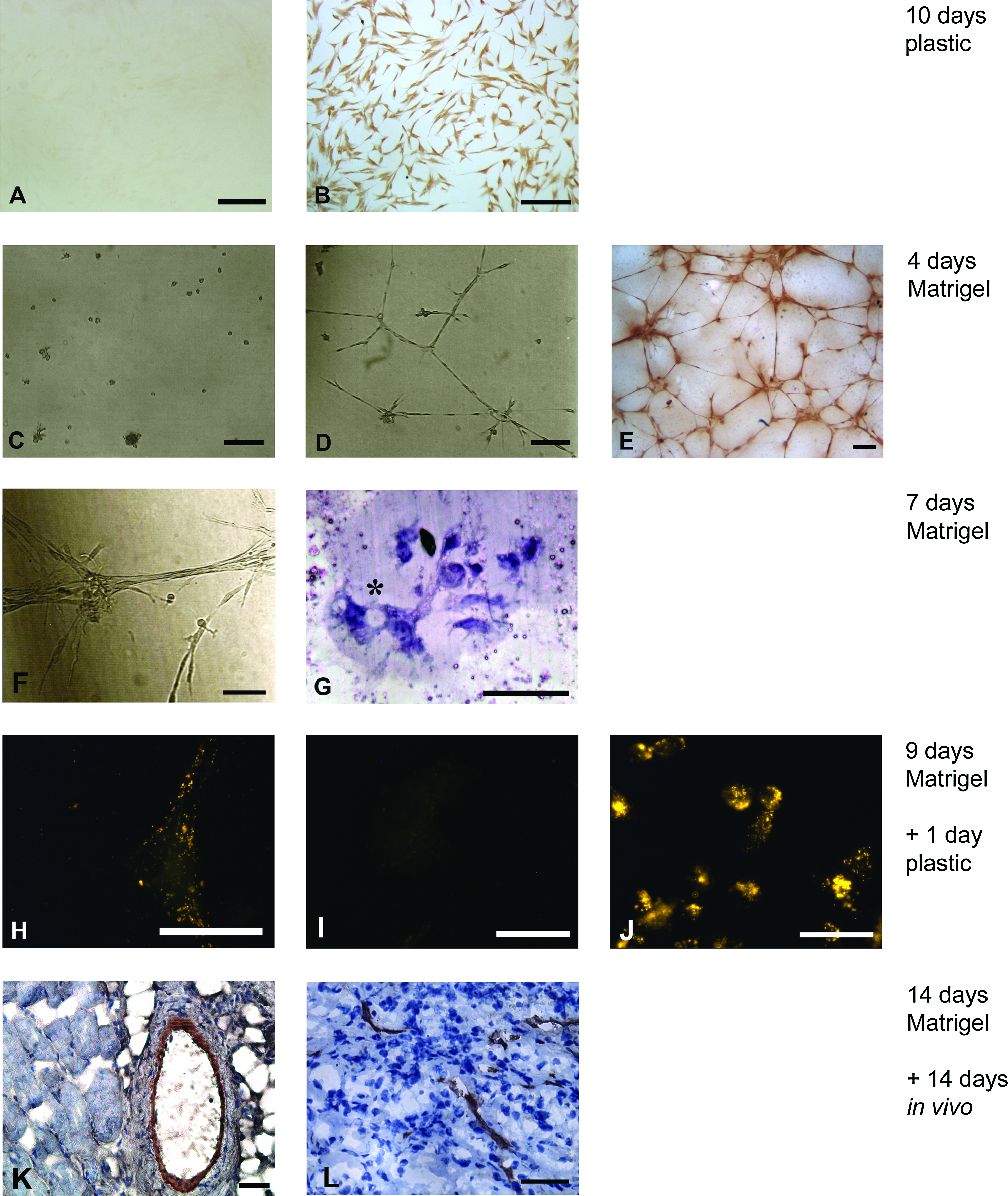
We investigated if hMSC from the bone marrow could be differentiated toward endothelial cells, for the use in prevascularized tissue engineering. Before differentiation, hMSC stained positive for the surface markers CD29, CD44, CD105, and CD166, and negative for CD31 (Fig. 1A). The addition of 50 ng/mL VEGF to hMSC resulted in the expression of the endothelial marker CD31 in approximately all the cells after 10 days as assessed by immunostaining (Fig. 2B). For improved endothelial differentiation, hMSC were cultured on Matrigel in EGM-2. This resulted in the formation of capillary-like structures within 4 days, whereas cells that were cultured on Matrigel in MSC proliferation medium retained a round morphology and did not organize or proliferate (Fig. 2C, D), or express the endothelial markers CD31 and vWF (data not shown). Visual observation revealed that >90% of the cells present were incorporated in these structures. Immunohistochemical analysis showed that the structures that were formed expressed the endothelial marker CD31 after 4 days of culture (Fig. 2E). After 7 days, the capillary-like structures had organized further into tubes containing multiple cells (Fig. 2F). Cross sections showed that lumina were present (Fig. 2G). Cells that were cultured on Matrigel in EGM-2 for 9 days were removed from the gel and reseeded on tissue culture plastic. Even though their morphology returned to a fibroblast-like morphology, they were able to take up acLDL (Fig. 2H), although uptake was limited when compared to HUVEC. Four samples of Matrigel containing capillary-like structures were implanted subcutaneously in nude mice. Because of resorption of the Matrigel, only two pieces of samples could be retrieved together with surrounding tissue after 14 days of implantation. Cross sections stained for human-specific CD31 revealed vessels of varying sizes, consisting of implanted human cells (Fig. 2K, L). A small fraction of these vessels was perfused, as was shown by the presence of erythrocytes (Fig. 2K).
Flow cytometric characterization of hMSC and EPC. (
Endothelial induction of hMSC. (
Endothelial progenitor cells
EPC are another promising cell source for prevascularized tissue engineering, due to their accessibility. Within hours after initial plating, MNC in part clustered and attached to the well surface; from the adherent clusters spindle-shaped cells grew within 4 to 7 days (early outgrowth LP-SS-EPC; Fig. 3A). Depending on culture conditions, LP-SS-EPC either maintained their spindle-shape morphology, or were induced to undergo a phenotypical change and change in proliferation rate while adopting a cobblestone morphology (HP-CS-EPC; Fig. 3D). Importantly, HP-CS-EPC did not appear to originate from distinct foci of proliferation in the well, but rather from a generalized transition in phenotype of the LP-SS-EPC (Fig. 3B). Late outgrowth LP-SS-EPC migrated out of the initial clusters to become dispersed throughout the well, but proliferated minimally and never reached confluence (Fig. 3C).
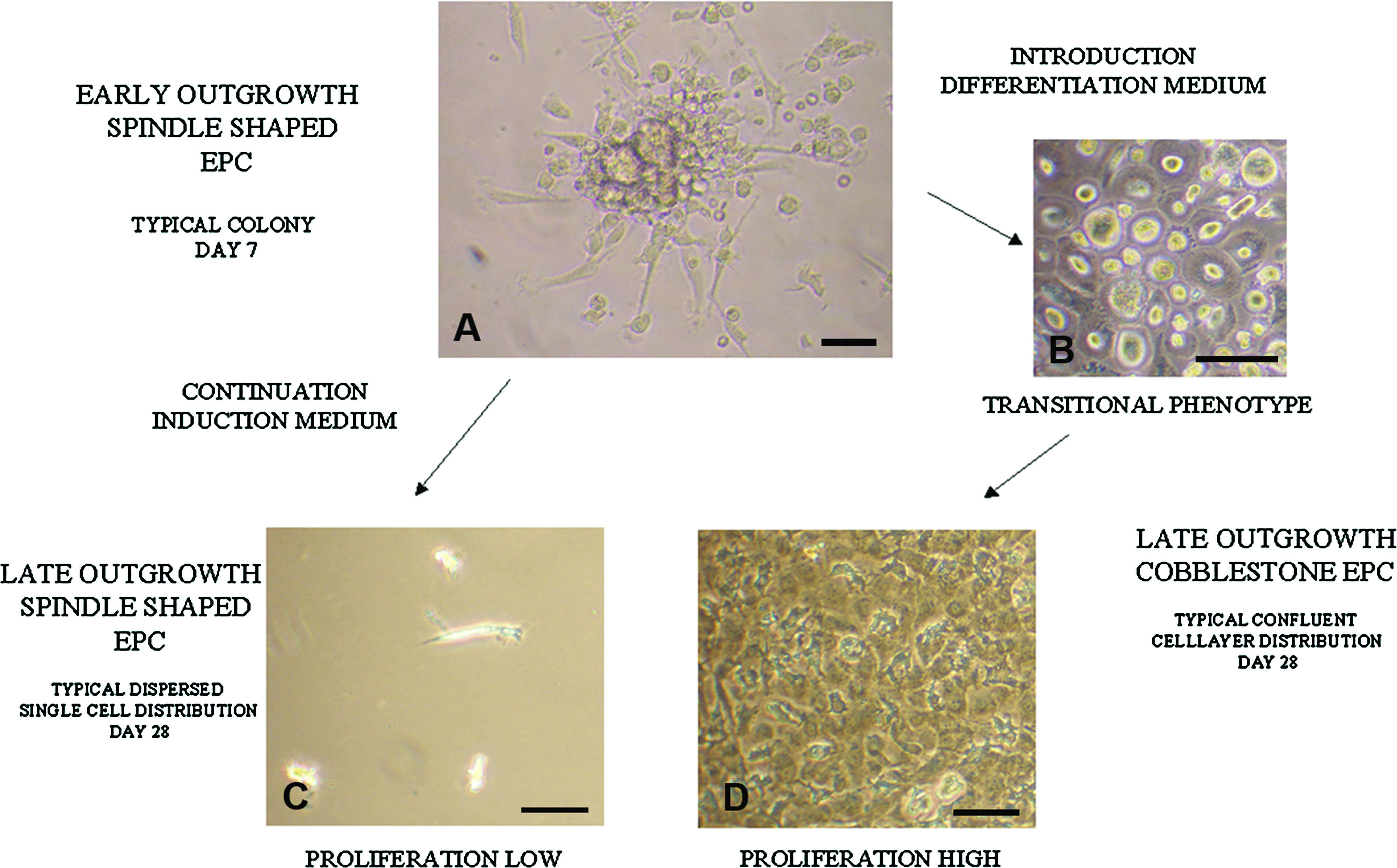
EPC morphology by light microscopy. At day 7 after plating blood MNC in EPC induction medium, typical colonies of early outgrowth spindle-shaped EPC were observed with the spindle-shaped cells predominantly located at the colony edges (
Both spindle- and cobblestone-shaped EPC bound UEA-1, stained positive for CD31, and took up acLDL (Fig. 4), which are considered typical EPC characteristics. Interestingly, vWF expression, which is a highly specialized characteristic of endothelial cells, was not observed in early or late outgrowth LP-SS-EPC. In contrast, HP-CS-EPC did show vWF expression, although expression was not as high as in HUVEC (Fig. 4). These observations are consistent with an immature endothelial phenotype in early and late outgrowth LP-SS-EPC versus a more mature endothelial phenotype in HP-CS-EPC.
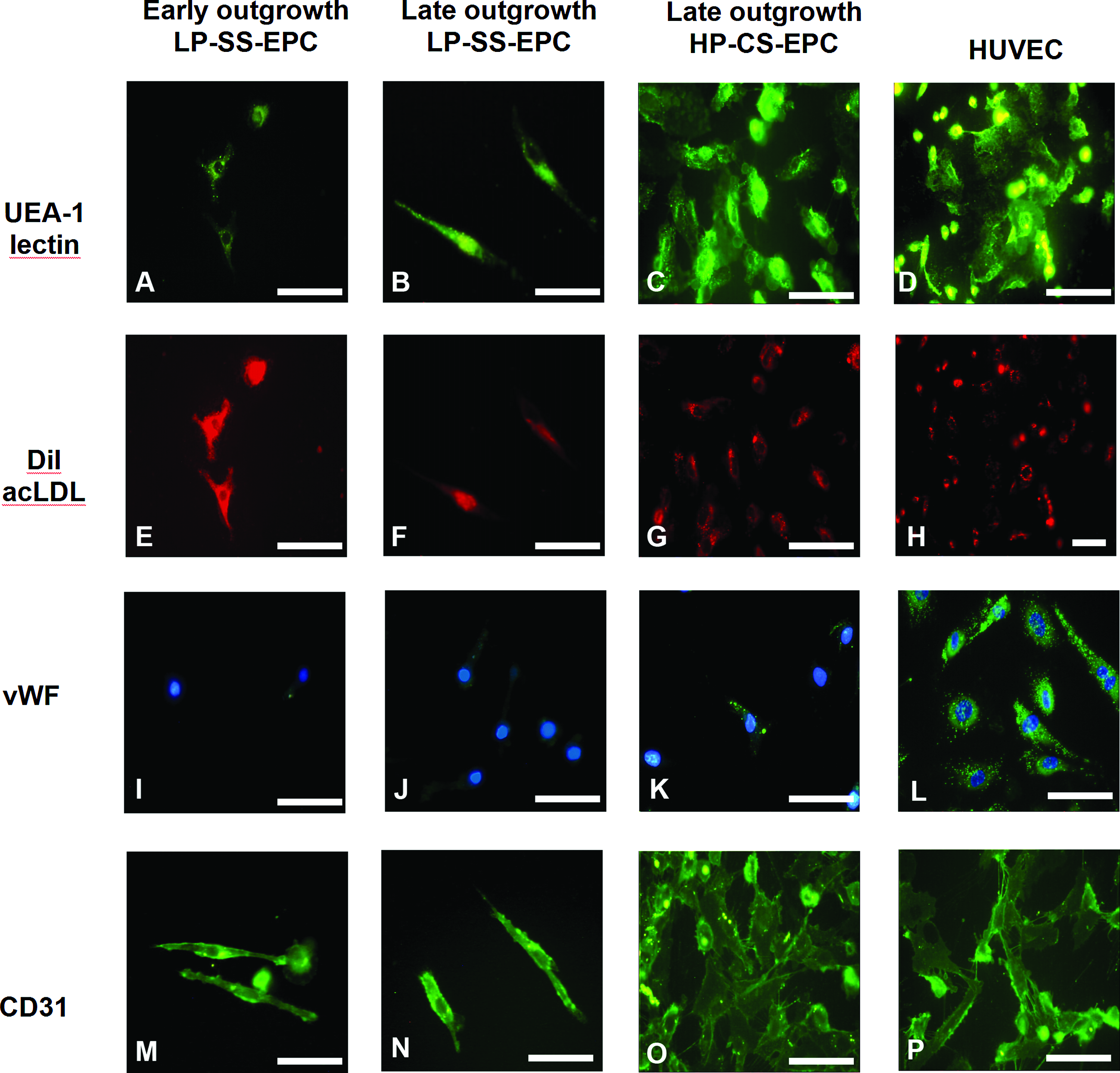
EPC characterization. Early and late outgrowth LP-SS-EPC and late outgrowth CS-HP-EPC bound FITC-labeled UEA-1 lectin, (
Coculture spheroids
The formation of coculture spheroids was similar as reported before. 23 Solid spheroids with a diameter of approximately 1 mm were formed spontaneously between days 3 and 5 with the method described. These spheroids remained intact for the remainder of the 10-day culture period. This current study revealed that the different E(P)C used did not alter the formation of coculture spheroids.
Prevascular network formation
The in vitro formation of a prevascular network in a tissue-engineered construct has the potential to improve vascularization after implantation, and thus survival of the implant. Different E(P)C were used to study their potential for the formation of a prevascular network in a bone tissue engineering setting. Immunostaining for the endothelial marker CD31 revealed that samples that were seeded with EC-MSC, early outgrowth LP-SS-EPC, or late outgrowth LP-SS-EPC did not contain CD31-positive cells after 10 days of coculture (Fig. 5J). However, round, unorganized CFSE-labeled EPC were still present (Fig. 5K). Moreover, TUNEL staining revealed that these cells were not apoptotic (Fig. 6). Together, this indicates that these cells had lost their endothelial potential during the course of coculture.
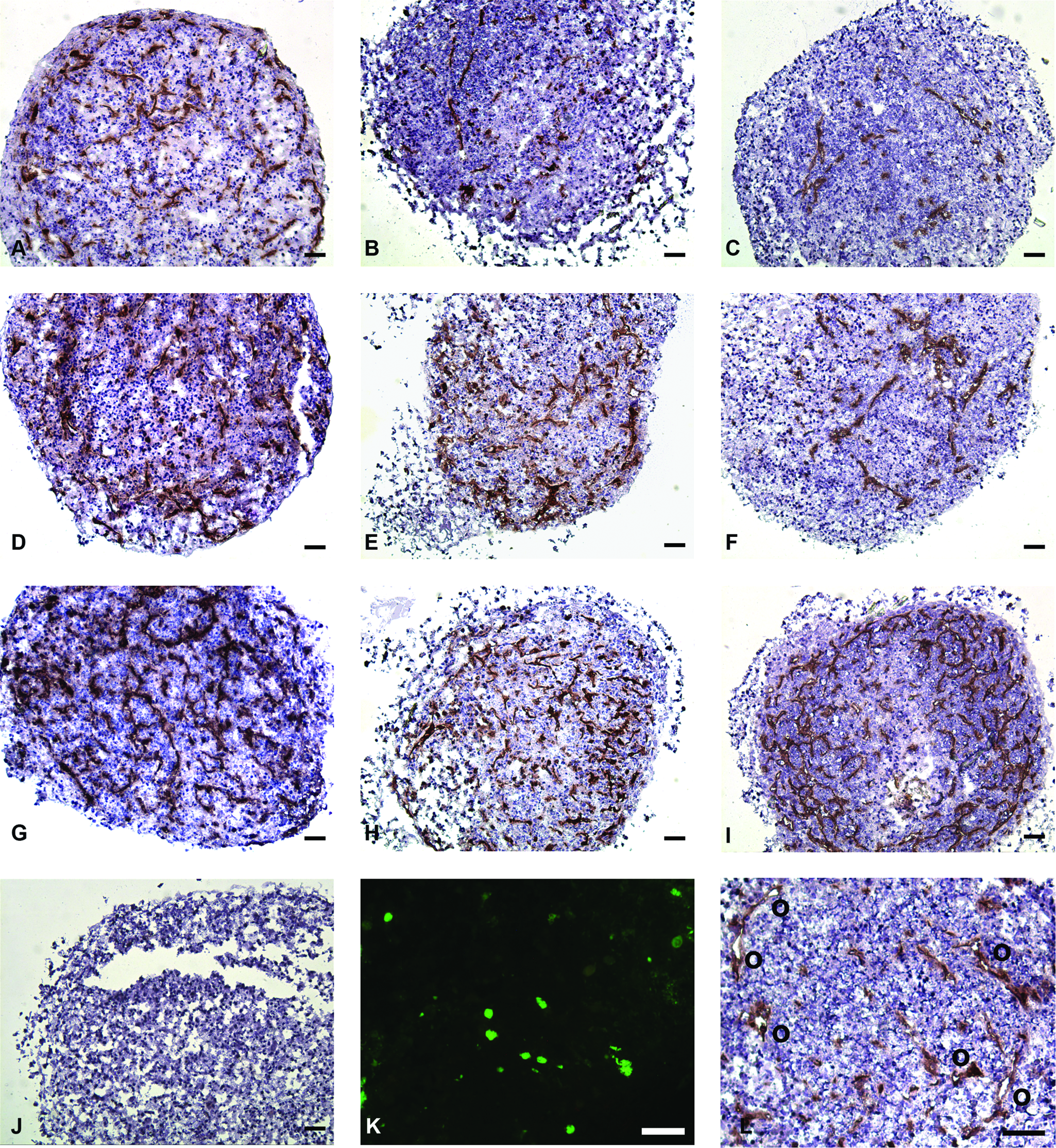
CD31 staining (brown) of coculture spheroids cultured in osteogenic differentiation medium for 10 days. Counterstained with hematoxylin. Scale bars = 100 μm. (
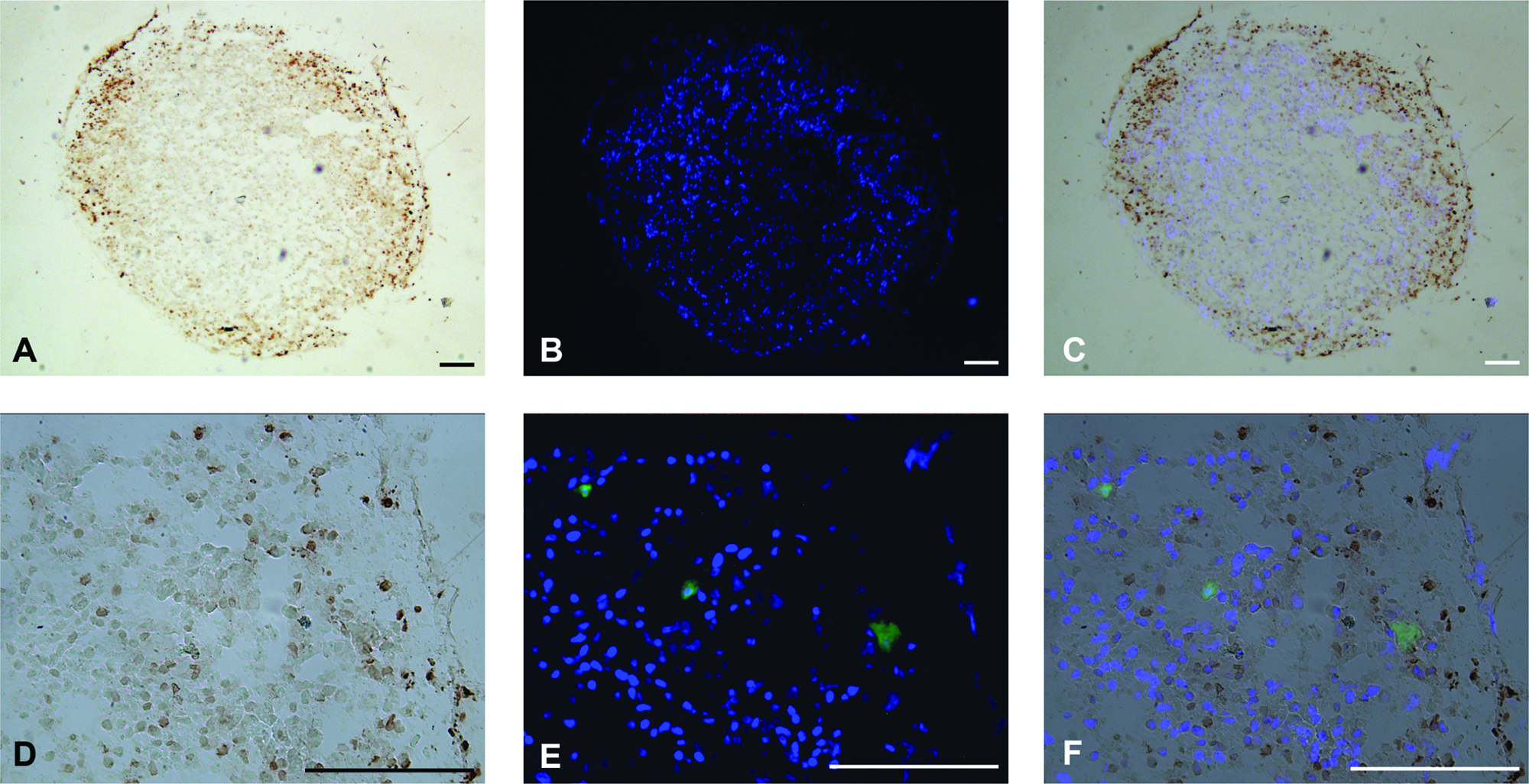
TUNEL assay of coculture spheroids of hMSC and CFSE prelabeled LP-SS-EPC. (
Samples seeded with HP-CS-EPC, HMVEC, or HUVEC did contain CD31-positive prevascular structures after 10 days (Fig. 5). The amount of CD31-positive structures varied when different cells were used. The amount of CD31-positive structures was quantified as the percentage of a cross section of the spheroid that stains positive for CD31. Quantification showed that seeding low percentages of HUVEC has a positive effect on the survival/proliferation of these cells, as reported before 23 (Fig. 7A). When 5%, 10%, or 15% HUVEC was seeded, the area that stained positive after 10 days of coculture was less than 5%, 10%, or 15%, respectively. When 1% or 2% of HUVEC was seeded, however, 5.8 ± 0.6% and 6.9 ± 2.4% of the area stained positive for CD31, respectively. When HMVEC was used for the coculture, results were similar to HUVEC for 5% or 10% endothelial cells seeded. Contrary to HUVEC, however, seeding low percentages of HMVEC hardly had a positive effect on the survival/proliferation of these cells. When 1% or 2% of HMVEC was seeded, 1.7 ± 1.0% and 1.7 ± 0.8% of the area stained positive for CD31, respectively. HP-CS-EPC performed comparable to HUVEC and HMVEC for 5%, 10%, and 15% endothelial cells seeded. When lower percentages of EPC were seeded, however, there was no positive effect on the survival/proliferation of these cells. Seeding 1% or 2% EPC resulted in 0.22 ± 0.26% and 0.71 ± 0.56% of the area staining positive for CD31, respectively.
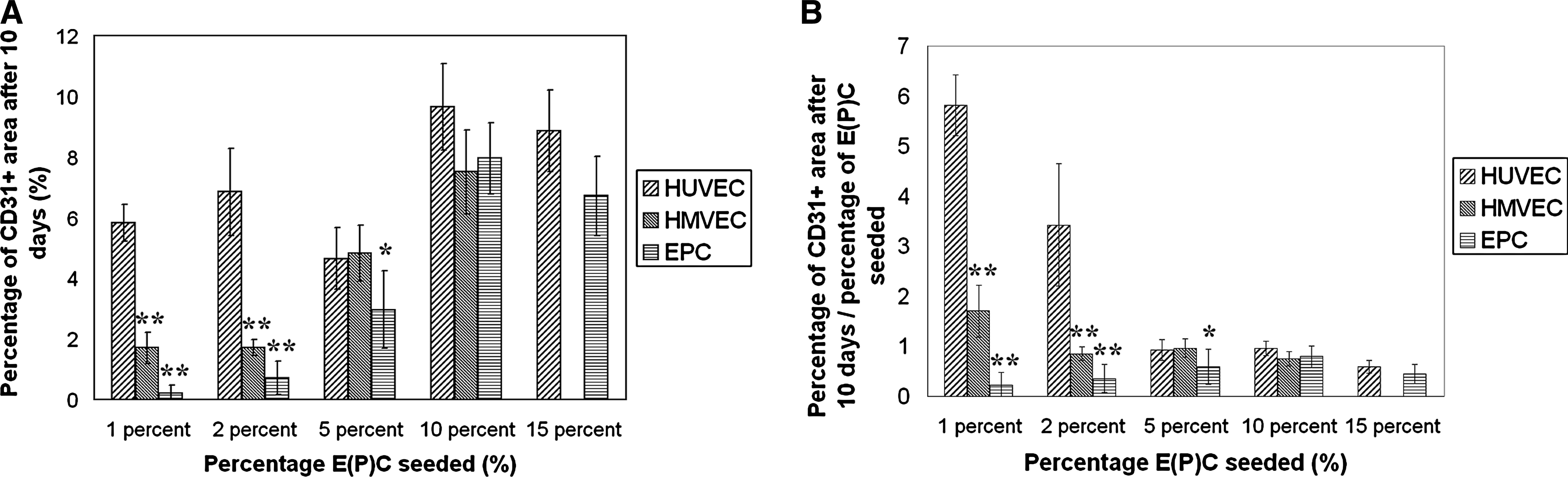
Quantification of the prevascular structures. Spheroids were seeded with different percentages of HUVEC, HMVEC, and HP-CS-EPC and cultured in vitro for 10 days. Results shown are mean values ± standard deviation (n = 4). (
As for the morphology of the CD31+ structures, a difference was observed when HUVEC, HMVEC, or HP-CS-EPC were used for the cocultures. Whereas HUVEC tended to organize more in cell clumps in stead of prevascular structures at higher seeding densities, HP-CS-EPC and to a lesser extent HMVEC organized into more elongated prevascular structures. Apart from that, the number of lumen was higher when HP-CS-EPC were used (Figs. 5L and 7B). The number of lumen when using HMVEC also seemed higher compared to HUVEC. However, a statistical difference was only evident when 10% of endothelial cells were seeded.
Discussion
Vascularization is recognized as an important factor in tissue engineering. It has been shown that the addition of endothelial cells to tissue cultures in vitro can result in the formation of prevascular structures.21,22,25 Moreover, we recently showed that prevascularizing a tissue in vitro can enhance tissue vascularization and survival in vivo. 22 We also developed a spheroid coculture system consisting of HUVEC and hMSC that resulted in the formation of prevascular networks. 23 Moreover, the addition of HUVEC resulted in an increased expression of the osteogenic marker alkaline phosphatase, suggesting that the osteogenic differentiation of MSC is not negatively influenced by the addition of endothelial cells. These spheroids can be used as modular systems for bone tissue engineering. However, most of these studies were performed with endothelial cell model systems that are not directly clinically applicable. For clinical use, an endothelial cell source should be found that can be easily isolated from the patient and can be expanded to sufficient numbers.
There are studies indicating that hMSC can differentiate toward endothelial cells in vitro.26–28 Because hMSC are already being used for bone tissue engineering, endothelial cells derived from this cell source would be ideal for prevascularized bone tissue engineering. If MSC from the bone marrow could be used for this purpose, it would mean that one could prepare a prevascularized bone tissue engineering construct from a single, easily accessible, bone marrow biopsy. This exciting concept has recently been demonstrated for rat MSC. 29
This study confirms that hMSC from the bone marrow can be differentiated toward endothelial cells in vitro. The bone marrow–derived hMSC used in this study were isolated and cultured according to a standardized procedure. 24 Even though there is no single specific marker for hMSC, a combination of several markers has been suggested for the identification of these cells. These include the surface-expressed antigen endoglin receptors CD105 and CD44, the activated leukocyte cell adhesion molecule CD166, and β-integrin CD29. 30 The cells used in this study were >90% positive for CD29, CD44, CD105, and CD166 and negative for CD31 (see Fig. 1), which makes them consistent with MSC, yet distinct from multipotent adult progenitor cells. 31 The addition of 50 ng/mL VEGF to MSC proliferation medium without bFGF resulted in the expression of CD31 in approximately all the cells after 10 days as assessed by immunostaining. However, the expression was minimal when assessed with flow cytometry analysis (data not shown), which is consistent with another report in literature. 28 The reason that CD31 was detectable by immunostaining, but only minimally by flow cytometry, can be explained by the fact that developing slides with DAB can result in amplification of a weak signal. Apart from that, the morphology of the cells did not change drastically, indicating that differentiation toward endothelial cells was limited in this setting. Culturing the cells on Matrigel in EGM-2 resulted in enhanced endothelial differentiation. Visual observation revealed that >90% of the cells present were incorporated in CD31-positive capillary-like structures, indicating that this behavior was not caused by the differentiation of a small subset of cells. Subcutaneous implantation of the capillary-like structures showed that human vessels were present after 2 weeks. Some vessels containing human cells were perfused with blood as indicated by the presence of erythrocytes. Although a quantitative analysis of this in vivo study could not be performed due to resorption of the Matrigel, these results do show that hMSC have the possibility to differentiate toward endothelial cells in vitro and can be incorporated in functional perfused vessels after implantation.
Another possible endothelial cell source for prevascularized bone tissue engineering is EPC. EPC originate from the bone marrow and can be mobilized upon secretion of angiogenic factors like SDF-1α and VEGF by ischemic tissue. 32 EPC then circulate in the blood and can differentiate into functional endothelium. 33 EPC can be derived from hematopoietic stem cells or from angiogenic monocytes.32,34 EPC can be isolated and differentiated from peripheral blood using ex vivo culture. 33 These EPC display endothelial phenotypical characteristics and can enhance neovascularization by incorporation and differentiation, and by the secretion of angiogenic factors affecting resident endothelium.35–37 EPC seeding of tissue-engineered small-diameter vascular grafts resulted in formation of an endothelial layer capable of fully covering the luminal surface in vitro.38,39 Importantly, clinical application of ex vivo cultured EPC has proven to be safe and feasible, as autologous EPC cultured from blood have been applied in an experimental setting to treat patients with acute myocardial infarction. 40 With promising results in vitro and in vivo and easy accessibility, EPC are an attractive candidate for prevascularized tissue engineering strategies.
Several isolation protocols for EPC have been established. In this study, EPC were isolated without prior sorting. MNC from the cord blood were directly seeded on gelatin and cultured under different conditions. This yielded three distinct groups of EPC in different stages of endothelial differentiation that allowed us to investigate the effect of the stage of endothelial differentiation for use in a complex tissue engineering application. At present, human cord blood–derived EPC are difficult to use for therapeutic vasculogenesis because cord blood transplantation is generally allogenic. However, cryo-preserved autologous cord blood may become available in the future. 41 Apart from that, there are striking similarities between EPC isolated from the cord and peripheral blood. 42 Future studies will investigate if the results in this paper can be replicated with EPC from the peripheral blood.
The direct in vitro differentiation of hMSC has been reported before.26–28 The state of differentiation as reported in these papers is comparable to the state achieved with the method described in this paper. Although these papers mention the promising possibilities of endothelial cells from hMSC for tissue engineering, they do not investigate the use of these cells for this purpose. To our knowledge, this is the first report of the use of endothelial progenitors from hMSC in a (complex) tissue engineering application. Apart from that, EPC from the blood have successfully been tested for their use in tissue engineering applications.43–45 However, the complexity of most models was limited. In general, the EPC were used as the sole cell source, or only combined with a single mature cell type. 46 This means that optimal culture conditions and media could be used to sustain the EPC, or that the secretion of disturbing growth factors by other cell types was limited. Tissue engineering, however, is focusing more and more on complex culture systems using multiple (progenitor) cell types to mimic the in vivo situation.47,48 It is likely that these situations will be more challenging for the cells, because they require the use of suboptimal culture conditions and intermediate medium systems to allow all cell types to perform simultaneously. This means for instance that the bulk addition of angiogenic growth factors like VEGF is not possible, because this may negatively influence the development of the base tissue that is to be vascularized. Apart from that, the use of multiple progenitor cells is likely to give rise to cellular interactions that could influence the differentiation of the progenitor cells used.
When labeled EC-MSC, early outgrowth LP-SS-EPC, or late outgrowth LP-SS-EPC were cocultured with undifferentiated hMSC, no positive staining for CD31 could be seen after 10 days. However, labeled cells were still present in the spheroid cross sections. The cells had not organized and were present as round, single cells. A TUNEL staining revealed that the cells were not apoptotic. The lack of CD31 staining is an indication that the EPC lost their endothelial potential during the course of coculture. HP-CS-EPC, as well as HUVEC and HMVEC, on the other hand, were capable of forming well-organized CD31-positive prevascular structures during 10 days of coculture with hMSC. Because only HP-CS-EPC, the EPC used with the most mature endothelial phenotype, and the mature HUVEC and HMVEC are capable of forming prevascular structures, these results clearly demonstrate the difference between the use of immature and mature EPC in this complex tissue engineering setting. With this, we have identified a degree of EPC predifferentiation to be a prerequisite for successful application in similar tissue engineering strategies. vWF expression might be a potential marker for an adequate state of differentiation.
Seeding low percentages of HUVEC had a positive effect on the survival/proliferation of these cells in coculture spheroids. The same cannot be said about HMVEC or HP-CS-EPC. As discussed in our previous paper, 23 the reason for this phenomenon remains unclear. Most likely, the communication between the different cell types is more optimal when low percentages of HUVEC are seeded, allowing for a better proliferation and organization of these cells. However, the possibility that a subfraction of hMSC differentiates toward endothelial cells and gets incorporated into the prevascular structures cannot yet be ruled out.
When comparing the morphology of the prevascular structures when using HP-CS-EPC, HUVEC, or HMVEC, distinct differences can be seen. HMVEC and especially HP-CS-EPC form more elongated structures, containing more lumen as compared to HUVEC. Similar results were obtained in a study by Sieminski et al. They compared the formation of microvascular networks in collagen gels by EPC from the blood and several vessel-derived endothelial cells. The amount of structures and lumen was highest when EPC were used. 49 It is interesting to note that in this system, the rate of organization is the opposite from the amount of structure formation when the different E(P)C are compared. Whether there is a direct relation between the amount of prevascular structures and the organization of these structures cannot be concluded from this data. However, it is known that differentiation of cells is often accompanied by a decrease in proliferation. 50 Why HP-CS-EPC, the cells with the lowest degree of maturation, produce the best organized prevascular structures remains unknown. It could be that a lower degree of maturation is accompanied by a higher degree of plasticity, enabling the cells to adopt better to new environments.
In summary, we have demonstrated the differentiation of hMSC toward endothelial cells in vitro. Although the differentiation was such that the cells formed capillary-like structures, stained positive for the endothelial marker CD31, were able to incorporate acLDL, and were incorporated in perfused vessels in vivo, the cells were not able to retain their endothelial characteristics when they were put in a 3D coculture environment with undifferentiated hMSC. The same was seen when two immature EPC types isolated from cord blood were put in this coculture environment. Although the cells were still present after 10 days, the cells did not organize and stopped expressing the endothelial surface marker CD31. On the other hand, a more mature EPC isolated from the cord blood was able to form prevascular structures in the 3D coculture with hMSC. Although the amount of structures was lower as compared to HUVEC and HMVEC, the degree of organization was higher. This study shows that EPC have potential for use in complex tissue engineering applications like prevascularized bone tissue engineering. However, the endothelial phenotype should be mature enough to prevent the loss of endothelial potential of the cells.
Footnotes
Acknowledgments
The authors thank Sanne Both for her assistance with the in vivo experiment. The authors thank Joost O. Fledderus for his assistance in EPC culture. This work was financially supported by IsoTis OrthoBiologics. The research of Jan de Boer was financially supported by a Senter/Novem grant.
Disclosure Statement
No competing financial interests exist.