
Abstract
Adipose-derived stromal cells (ASCs) are suggested to be potent candidates for cell therapy of ischemic conditions due to their ability to stimulate blood vessel growth. ASCs produce many angiogenic and anti-apoptotic growth factors, and their secretion is significantly enhanced by hypoxia. Utilizing a Matrigel implant model, we showed that hypoxia-treated ASCs stimulated angiogenesis as well as maturation of the newly formed blood vessels in vivo. To elucidate mechanisms of ASC angiogenic action, we used a co-culture model of ASCs with cells isolated from early postnatal hearts (cardiomyocyte fraction, CMF). CMF contained mature cardiomyocytes, endothelial cells, and progenitor cells. On the second day of culture CMF cells formed spontaneously beating colonies with CD31+ capillary-like structures outgrowing from those cell aggregates. However, these vessel-like structures were not stable, and disassembled within next 5 days. Co-culturing of CMF with ASCs resulted in the formation of stable and branched CD31+ vessel-like structures. Using immunomagnetic depletion of CMF from vascular cells as well as incubation of CMF with mitomycin C–treated ASCs, we showed that in co-culture ASCs enhance blood vessel growth not only by production of paracrine-acting factors but also by promoting the endothelial differentiation of cardiac progenitor cells. All these mechanisms of actions could be beneficial for the stimulation of angiogenesis in ischemic tissues by ASCs administration.
Introduction
Given the complexity of cellular signaling pathways in vivo, this environment is often difficult to mimic completely in vitro. Co-culture models have previously been used to successfully direct the differentiation of stem or precursor cells into neurons,6–8 hematopoietic cells,9,10 photoreceptor cells, 11 and hepatocytes, 12 and to promote cell survival and expansion in vitro.4,13,14 Co-culture models provide a method to study angiogenesis under controlled conditions, with the advantage of being able to replicate some tissue-derived signaling.
In the present work we show that ASCs stimulate vessel-like structure formation and stabilization in vivo in a Matrigel implant model. Using a co-culture model of ASCs with cells isolated from early postnatal heart cardiomyocyte fraction (CMF), we have clarified mechanisms of these stimulating effects of ASCs. Observed angiogenic effects are only partly mediated by paracrine angiogenic and anti-apoptotic factors produced by ASCs. We suppose that direct cell–cell contacts between ASCs and CMF cells appear to be important. Moreover, ASCs stimulate formation of capillary-like structures not only from preexisting endothelial cells in the CMF, but also by induction of progenitor cell differentiation into endothelial cells.
Materials and Methods
Animals
For Matrigel experiments ASCs were isolated from subcutaneous adipose tissue of Balb/c male mice 5–6 weeks old1,3 and maintained in Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA) containing penicillin 100 U/mL, streptomycin 100 U/mL, and 10% fetal bovine serum (DMEM/FBS) at 37°C and 5% CO2. For co-culture experiments ASCs were obtained from adult male Wistar rats. CMF of cells was harvested from hearts of early postnatal (2–5 days) Wistar rat pups. Animals were anesthetized by i.p. injection of 2.5% Avertin before manipulations.
Animal studies were conducted according to Institute of Experimental Cardiology–approved guidelines, as well as approved guidelines of the Institutional Animal Care and Use Committee of Cardiology Research Center.
Isolation and culture of ASCs
Cells from adipose tissue were isolated using the protocol described by Zuk et al., with minor modifications. 1 Briefly, subcutaneous fat tissue from inguinal region (regio inguinalis) was washed twice in sterile PBS and was minced into pieces of 2 mm in diameter in a Petri dish. Pieces were transferred to a conical tube, and collagenase I type (Worthington Biochemical, Lakewood, NJ), 220 U/mL, and dispase 40 U/mL (Invitrogen Corporation, Germany) were added (in equal volumes). Pieces were digested at 37°C for 30 min with constant mixing. Enzyme activity was neutralized with DMEM, containing 10% FBS (HyClone, Logan, UT), penicillin 100 U/mL, and streptomycin 100 U/mL, and the content was centrifuged at 400 g for 5 min to obtain a high-density stromal vascular fraction pellet. The cell pellet was treated with red cell lysis buffer (NH4Cl 154 mmol/L, KHCO3 10 mmol/L, and EDTA 0.1 mmol/L) for 10 min after centrifugation. Fresh medium was added, and SVF was collected by centrifugation, as detailed above, and filtered through a 40-μm nylon mesh (BD Biosciences, Franklin Lakes, NJ, Falcon Cell Stainer #352340) to remove cellular debris and remaining tissue. Cells were centrifuged and resuspended in MyeloCult media (Stem Cells, Vancouver, Canada, #M5300), penicillin 100 U/mL, and streptomycin 100 U/mL, and incubated at 37°C in 5% CO2 incubator. After 2 days the medium was changed and cells were cultivated in DMEM, containing 10% FBS, penicillin 100 U/mL, and streptomycin 100 U/mL at 37°C in 5% CO2 incubator. Cells normally reached confluent state in 4–5 days. Upon reaching the monolayer, cells were detached from the culture dishes using 0.25% trypsin solution, and passaged. For experiments, cells of the second passage were used.
Isolation and culture of cells from postnatal hearts
Hearts for co-culture were isolated from postnatal (2–4 days after birth) Wistar rat pups as described earlier15,16 with some modifications. Rat pups were anesthetized by 2.5% Avertin solution. Hearts were isolated into sterile Krebs-Ringer (Sigma, St. Louis, MO) solution, and 50 mM Ca2+ was added. Myocardial tissue was minced and then digested in collagenase A 0.2 mg/mL (Roche Molecular Biochemicals, Indianapolis, IN). Cells were harvested during five sequential steps of digestion and washing (10 min each step at 37°C), and fresh collagenase was added each time to the remaining heart tissue pieces. The isolated cells were preplated in preplating media (one volume of medium 199 and four volumes of DMEM, Horse serum 5%, FBS-5, penicillin 100 U/mL, and streptomycin 100 U/mL) for 1 h to remove fibroblasts and leukocytes. The obtained cardiomyocyte cell fraction (CMF) was cultured on 60 mm Petri dishes in DMEM containing FBS 5%, penicillin 100 U/mL, and streptomycin 100 U/mL at 37°C in 5% CO2 incubator. After that, CMF cells were used for immunomagnetic separation or were mixed with ASCs (with concentration 4:1, respectively) for co-culture experiments.
Immunofluorescence analysis
For immunofluorescence analysis cultured cells were washed with PBS and fixed in 4% formaldehyde for 4 min at room temperature. Cells were then washed and fixed with 70% ethanol for 10 min and washed with PBS. Nonspecific binding was blocked by incubation in 1% BSA with 10% serum of secondary antibodies donor for 1 h. Immunostaining was performed using the following antibodies (mouse or rabbit): connexin 43 (Zymed, South San Francisco, CA), NG2 (Chemicon, Billerica, MA), CD31 (PECAM-1) (Serotec, Raleigh, NC or Pharmingen), CD105 (endoglin) (R&D Systems, Minneapolis, MN), CD90 (Calbiochem, Gibbstown, NJ), GATA 4 (Santa Cruz, Santa Cruz, CA), Nkx2.5 (Santa Cruz), α-actinin (Sigma), cardiac myosin light chain-1 (Abcam, Fremont, CA), and troponin I (Chemicon). This was followed by incubation with secondary antibodies: Alexa donkey anti-rabbit 488 (Molecular Probes, Carlsbad, CA) or Alexa donkey anti-mouse 594 (Molecular Probes). Cells were counterstained with nuclear 4-,6-diamidino-2-phenylindole (Molecular Probes). For negative controls mouse or rabbit nonspecific IgGs were used in an appropriate concentration.
Blood vessels were visualized by immunofluorescent staining as described earlier.17,18 For visualization of endothelial cells, rat anti-mouse CD31 antibody (BD Pharmigen™, San Diego, CA) was used. Pericytes were revealed by the expression of high molecular weight melanoma antigen (NG2 antigen)19,20 using rabbit anti-NG2 (Chemicon) antibodies. Rabbit anti-mouse α-actin (Epitomics, Burlingame, CA) antibodies were used to visualize smooth muscle cells (SMCs). Alexa 488–conjugated anti-rat antibody (Molecular Probes) or Alexa 488–conjugated donkey anti-rabbit antibodies were used as secondary antibodies. Images were obtained using Zeiss Axiovert 200 M microscope equipped with CCD camera, AxioCam HRc, and Axiovision software (Zeiss), and further processed using Adobe Photoshop software (Adobe Systems).
Immunomagnetic sorting
CMF was depleted of endothelial cells and pericytes using immunomagnetic sorting. Briefly, magnetic beads were washed from sodium azide, and then two types of magnetic beads were prepared: pan-anti-mouse IgG–conjugated Dynabeads were incubated with mouse anti-rat monoclonal antibody against CD31, and sheep anti-rabbit IgG–conjugated Dynabeads were incubated with rabbit anti-rat monoclonal NG2 antibody for 40 min on ice. Then, both types of beads were incubated with CMF cells (for 1 h on ice), and CMF cell mixture was separated in a magnetic field. We aimed to separate two cell fractions: CD31+/NG2+ cells and CD31−/NG2− cells. The purity of the obtained populations was proven by flow cytometry analysis.
Co-culture experiments
For co-culture experiments we used ASCs (passage 2) and freshly isolated cells from CMF in ratio 1:4. We selected the following groups for analysis: CMF cultured with ASCs, CMF cultured in the presence of conditioned medium from ASCs, CMF cultured with ASCs exposed to mitomycin-C (mitC ASCs), ASCs cultured with CD31−/NG2− fraction of CMF (depl. CMF), and ASCs cultured with CD31+/NG2+ fraction of CMF (enrich. CMF). Control groups were ASCs and CMF cells cultured alone.
To prevent DNA synthesis and division of ASCs, we used mitomycin-C (Sigma) in several co-culture experiments. For that, 0.05 mg/mL of mitomycin-C was added to cultivation media for 24 h. ASCs were arrested in the cell cycle as confirmed by the immunofluorescent staining with antibody against PCNA (data not shown).
To evaluate the paracrine effect of ASCs and their secreted factors on the formation of CD31+ vessel-like structures from cells of CMF, we cultivated CMF cells in the presence of conditioned medium from ASCs. For these experiments the conditioned medium was centrifuged at 1000 g for 10 min and 100 times concentrated using Centrycon YM-10 (Millipore, Billerica, MA) filter.
On days 8–11, co-cultured cells were fixed and used for immunofluorescence analysis. To distinguish one cell type from another, cells were prelabeled by lipophylic fluorescent dye CM-DiI-Cell Tracker (Molecular Probes). According to manufacturer protocol, cells were washed with PBS and exposed to DiI (1.5 μL/mL) for 5 min at 37°C and then for 15 min at 4°C.
RNA extraction from ASCs, RT-PCR, and real-time polymerase chain reactions
Total cellular RNA was isolated from ASCs of the second passage and cultured for 2 days on standard cell culture plastic (1% O2 or standard conditions), using RNeasy Kit (Qiagen, Venlo, The Netherlands), followed by a reverse transcription reaction with the Fermentas Reverse Transcription kit (Fermentas, Burlington, Canada). Real-time polymerase chain reactions (PCRs) were performed using SYBR Green PCR Master Mix (Syntol, Russia) on the Rotor-Gene R6-3000 (Corbett Research, UK). PCRs were performed in a final volume of 25 μL reaction mix that contained 50 ng of cDNA product and 100–500 nmol/L of each primer. For all reactions, thermal cycling parameters were 10 min at 95°C followed by 40 cycles of 15 s at 95°C for denaturation, 20 s at empirically adjusted temperature for each primer for annealing, and 20 s at 72°C for extend. The oligonucleotides used as primers are summarized in Table 1. All data were duplicated for every sample. Level of expression was normalized to the expression of two house-keeping genes, GAPDH and L7.
Hypoxia model
After changing the medium to fresh DMEM containing penicillin 100 U/mL, streptomycin 100 U/mL, and 10% fetal bovine serum (DMEM/FBS), ASCs at second passage (70–80% of confluent) were cultivated in hypoxic incubator at 37°C, 5% CO2, and 1% O2 (hypASC), or in standard conditions (normASC) for 48 h. Then, cells were detached by 0.25% trypsin and counted.
Matrigel implantation into syngenic mice
About 400 μL of ice-cold growth factor–reduced Matrigel matrix was premixed with 7 × 105 ASCs (after cultivating in either hypoxia or normoxia conditions) in 100 μL of complete DMEM. Eight- to 9-week-old syngenic Balb/c mice were anesthetized with by i.p. injection of 2.5% Avertin. About 500 μL of cell suspension in Matrigel was injected subcutaneously into the right and left sides of the mouse back (spatula region) using 23G needle. As a negative control we used Matrigel containing 100 μL of complete DMEM and, as a positive control, Matrigel with the inclusion of 50 ng/mL of bFGF (R&D Systems) and heparin. Ten days after injection, mice were sacrificed and the entire Matrigel plugs were isolated. Quantification of blood vessels was achieved using immunofluorescent visualization of blood vessels on frozen Matrigel sections.17,18
Quantification of vessel density
Vessel size and density were evaluated using MetaMorph 7.1 (Molecular Devices) and Adobe Photoshop software (Adobe Systems). vessel density was determined as described earlier. 20 All vessels were counted separately in three groups: capillaries (CD31+ vessels without lumen and with length less than 20 μm), medium vessels (CD31+ vessels with length or diameter 20–50 μm), and large vessels (with diameter >50 μm).
Flow cytometry
The expression of c-kit, as well as CD31 and NG2 antigens, on ASCs or cells from CMF was analyzed using flow cytometry. Cells were detached from culture dish by EDTA solution, suspended, filtered through 40 μm cell strainer, and incubated with primary antibodies or control IgGs for 1 h at room temperature, followed by the incubation with secondary antibodies, conjugated with Alexa488. Analysis was performed on MoFlo instrument (DakoCytomation).
Tube assay
Human umbilical vein endothelial cells (HUVECs; a kind gift from Dr. O. Antonova Cardiology Research Complex, Moscow, Russia) were maintained and propagated in DMEM containing 10% FBS (HyClone), 200 μg/mL endothelial cell growth factor (RELIA Tech GmbH, Germany), heparin 5 U/mL, sodium pyruvate 1 mM (Gibco BRL, Carlsbad, CA), penicillin 100 μg/mL, streptomycin 100 μg/mL (Gibco BRL), and HEPES 20 mM (Helicon, Moscow, Russia). Experiments were performed on cell cultures of the 2–4 passages. The formation of vascular-like structures by HUVECs on growth-factor-reduced Matrigel (BD Biosciences) was performed as previously described.18,21 Twenty-four-well culture plates were coated with Matrigel according to the manufacturer's instructions. About 5 × 104 of HUVECs were seeded on prepared Matrigel. Conditioned medium was collected from ASCs, cultured in either hypoxic or standard conditions, and concentrated 100 times using 10 kDa Centricon (Millipore). HUVECs were incubated at 37°C for 24 h in growth medium containing either 1/100 of concentrated conditioned medium, or 0% or 20% of FBS. Then, every culture well was photographed using Axiovert 200 M (Zeiss, Göttingen, Germany). The total length of formed capillary-like structures was quantified in five randomly chosen fields using MetaMorph software (Molecular Devices).
Statistical analysis
All experiments were repeated at least three times. Data are expressed as mean ± SEM. Data were analyzed using Mann-Whitney U-criteria. Differences with p < 0.05 was considered as statistically significant.
Results
Hypoxia-treated ASCs enhance all stages of angiogenesis in Matrigel implants
In this study we evaluated the stimulating effect of hypoxia-treated ASCs on blood vessel growth in Matrigel implants in vivo. Cells responded to the 48 h incubation in hypoxic conditions by upregulation of angiogenic growth factor mRNA. Thus, mRNA of vascular endothelial growth factor (VEGF), hepatocyte growth factor (HGF), its receptor c-met, basic fibroblast growth factor (bFGF), platelet-derived growth factor B (PDGFB), and transforming growth factor beta (TGFβ) was upregulated as was analyzed by real-time PCR. The elevation of mRNA levels in ASCs treated by hypoxia was statistically significant for the VEGF (3.5 ± 0.8–fold, p < 0.05), HGF (4.1 ± 0.9–fold, p < 0.05), c-met (3.1 ± 0.7, p < 0.05), and PDGFB (2.7 ± 0.6, p < 0.05) genes. However, for bFGF (1.7 ± 0.3, p = 0.068) and TGFβ (1.8 ± 0.4, p = 0.075) only a trend toward an increase was noted. Observed elevation of angiogenic growth factor mRNA was accompanied by significant decline of anti-angiogenic factors, including thrombospondin-1 and endostatin; for plasminogen activator inhibitor-1 (PAI-1), the decrease in expression was not significant (Table 2).
Gene expression was assessed by real-time polymerase chain reactions (PCR) using SYBR GREEN for amplicon's detection. Level of expression was normalized to the expression of housekeeping genes GAPDH and L7.
To prove that hypoxia treatment has indeed stimulated the production of angiogenic growth factors by ASCs, we tested the ability of conditioned medium collected from those cells to upregulate the formation of capillary-like structures by HUVECs on Matrigel in vitro. Conditioned medium collected from hypoxia-treated ASCs has stimulated the formation of capillary-like structures up to 1.7 times, whereas conditioned medium from control cells, only up to 1.5 times (Fig. 1). These data confirm that hypoxia induces angiogenic factor production by ASCs, which stimulate formation of capillary-like structures by endothelial cells in vitro.
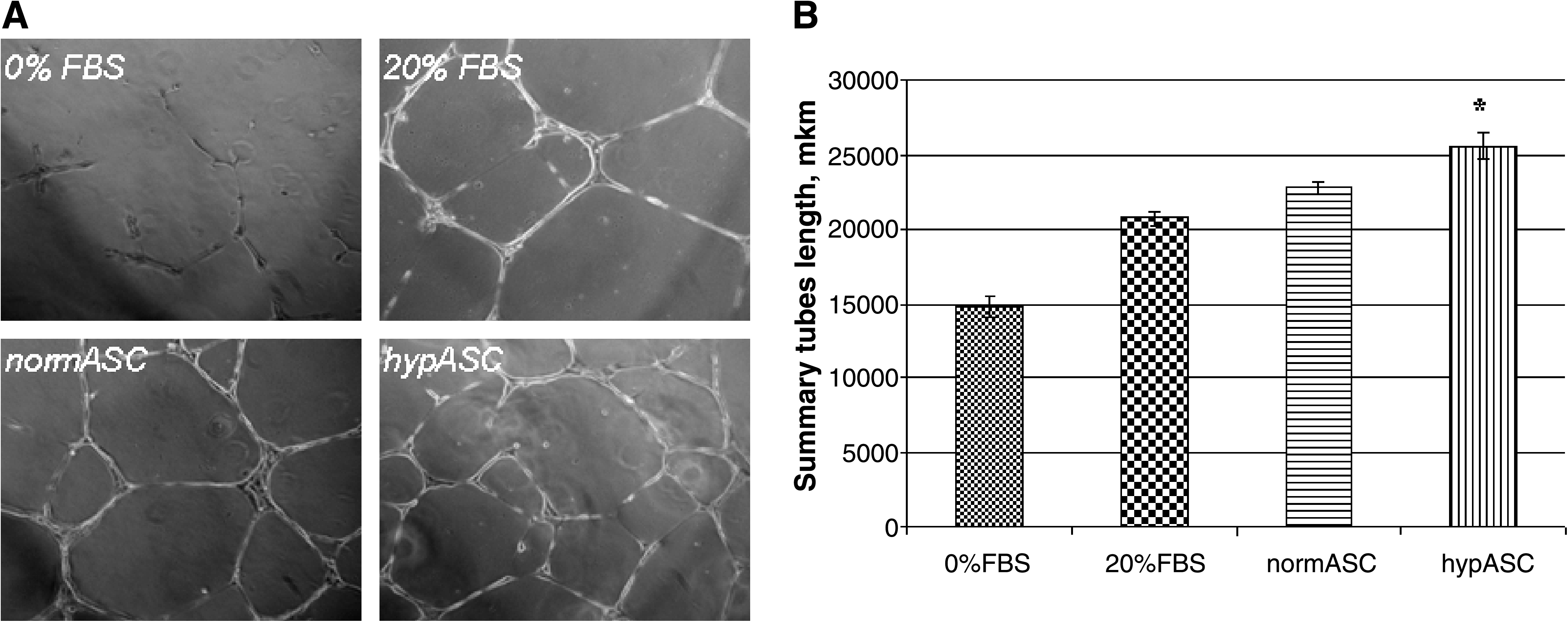
Formation of capillary-like structures in vitro. (
To assess the angiogenic effect of ASCs cultured in hypoxic conditions, we utilized Matrigel implant model as described earlier. 18 We have subcutaneously injected Matrigel containing ASCs treated by hypoxia (hypASC) or cells incubated in standard normoxic (normASC) conditions in the mouse back (spatula region). As a negative control we used Matrigel containing 100 μL of complete DMEM, and as a positive control Matrigel with the inclusion of 50 ng/mL of bFGF (R&D Systems) and heparin. After 10 days Matrigel plugs were isolated and their vascularization with vessels outgrowing from arteria subclavia was assessed. At day 10, Matrigel plugs with hypASC and normASC were weighed, and the hemoglobin content was evaluated. Matrigel plugs with hypASC were similar in weight with plugs contained normASC (0.174 ± 0.07 vs. 0.166 ± 0.03 for normASC, p >0.05). The hemoglobin content normalized to the weight of the analyzed fragment of Matrigel was a bit higher in plugs contained hypASC (3.47 ± 0.44 vs. 2.96 ± 0.28 for normASC, p > 0.05), but the difference was not statistically significant (data not shown).
Size and type/quality of blood vessels as well as their density in Matrigel implants were assessed using immunofluorescent staining with subsequent image analysis. Matrigel sections were taken from various parts of the plug and stained with antibody against endothelial marker CD31 (Fig. 2A–C). The vascular density was significantly higher in the plugs containing hypASCs versus plugs containing normASC (153.1 ± 21.4 vs. 80.3 ± 6.4; p < 0.05). The presence of hypASCs enhanced the number of all types of vessels, including large ones with lumen diameter > 50 μm (2.0 ± 0.3 vs. 0.8 ± 0.2; p <0.05), medium vessels with length or diameter 20–50 μm (5.6 ±0.5 vs. 3.1 ± 0.2; p < 0.05), and capillaries (145.5 ± 21.1 vs. 76.3 ± 6.5; p < 0.05) (Fig. 2G–I). Further, as revealed by double immunostaining with antibodies against endothelial marker CD31 and a marker of pericytes NG2,19,20 all vessels determined by size as medium and large vessels were mature18–20 because they were surrounded by cells expressing NG2 (Fig. 2D–F) and SMCs expressing α-actin (data not shown).
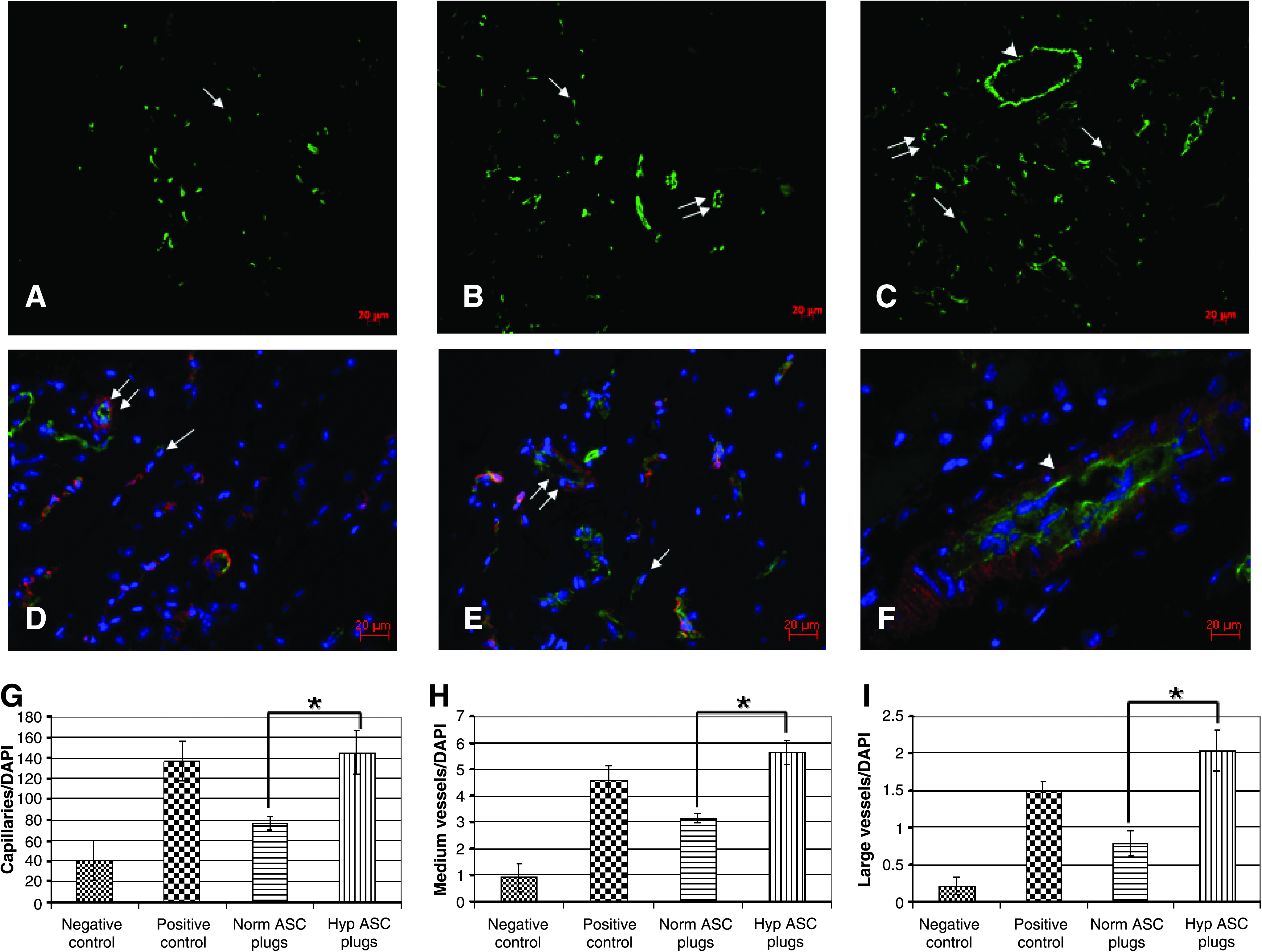
Vascularization of Matrigel plugs. (
The obtained data indicate that injection of hypoxia-treated ASCs in the Matrigel plugs stimulates all stages of blood vessel growth: the initial formation of capillaries as well as the maturation of the newly formed vessels.
ASCs promote vessel outgrowth in CMF
To clarify mechanisms of ASC angiogenic action, we utilized a co-culture model of ASCs with cells isolated from early postnatal hearts. Although freshly isolated ASCs contain a substantial amount of CD31/CD144+ endothelial cells, they are mostly eliminated in selective culture conditions during 1–2 passages. 22 Our cultures of ASCs contained only 0.09% of CD31+ endothelial cell (Fig. 3) and did not form vessel-like structures on the second passage. However, these ASC cultures contained up to 45.55% of NG2-expressing pericytes as shown by flow cytometry and immunofluorescent staining (Fig. 3). Further, these cells also expressed connexin-43 (Fig. 3), a protein of gap junctions between endothelial cells and pericytes. 23
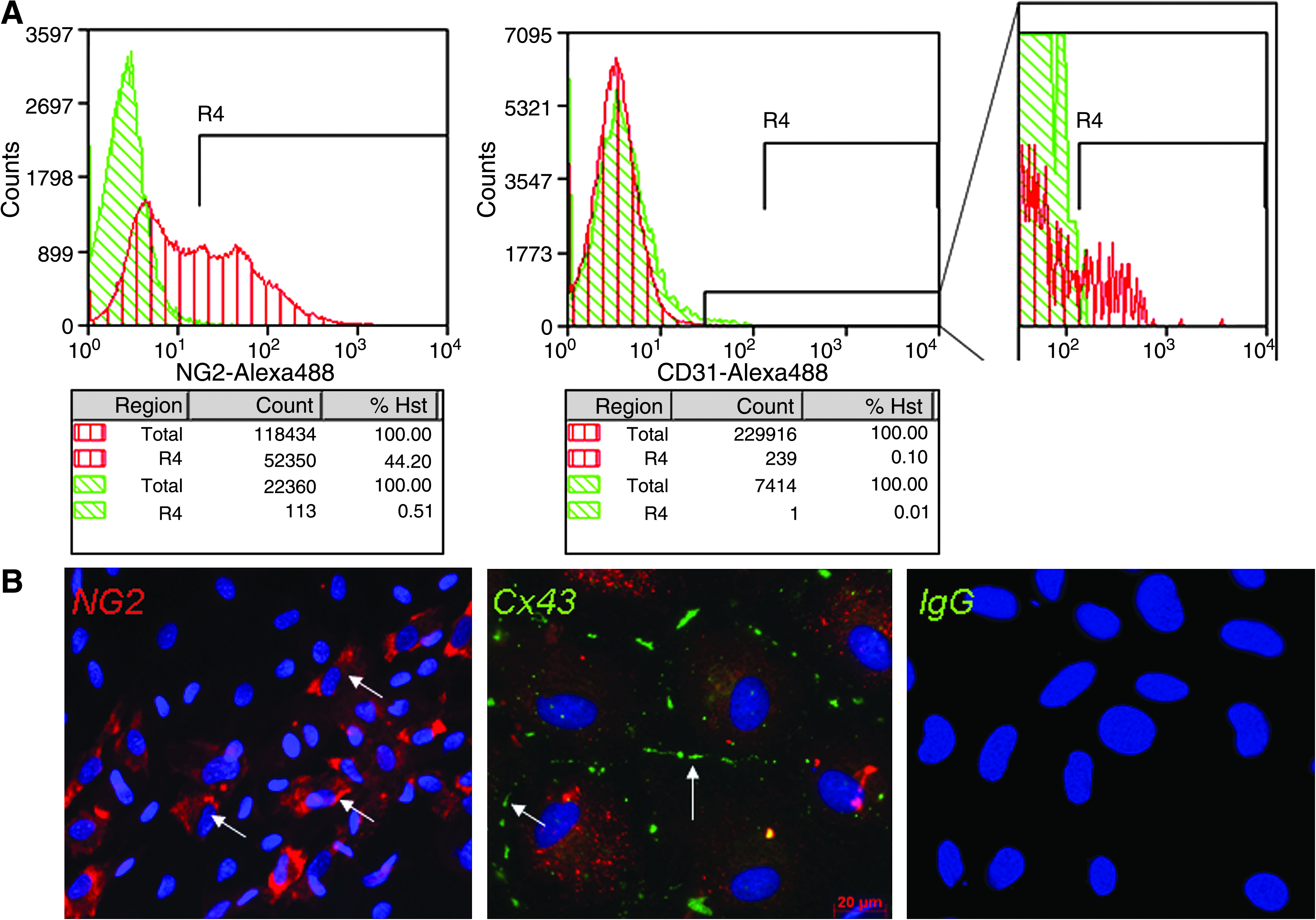
Characterization of the ASCs of the second passage. (
Cell fraction isolated from early postnatal rat hearts consisted of mature cardiomyocytes expressing α-actinin, myosin light chain-1, and connexin-43 (Fig. 4B), as well as fibroblasts and SMCs expressing CD90 and α-actin (data not shown). This fraction also contained minor populations of endothelial cells expressing CD31 (PECAM-1) (0.13%) and c-kit+ early progenitor cells (3.44%) (Fig. 4A). Consistent with the observation of c-kit+ population, we identified in CMF cells expressing markers of early cardiogenesis such as Nkx2.5 and GATA-4, 24 as well as CD105 (endoglin)—a marker of angiogenic endothelial cells 25 (Fig. 4B). On the second day of culture, cells from the CMF formed spontaneously beating colonies with CD31+ capillary-like structures outgrowing from those cell aggregates (Fig. 5). However, those CD31+ vessel-like structures were not stable and disassembled after 5 days of culture.

Characterization of the cell culture isolated from rat postnatal hearts (CMF). (

Colonies with outgrowing vessel-like structures in CMF culture. (
Addition of ASCs to cells of CMF increased the formation of CD31+ vessel-like structures up to sixfold. In contrast to culture of cells from CMF alone, these structures were thick, branched, and stable up to 21 days (Fig. 6). To assess if angiogenic factors secreted by ASCs contribute to this effect, we cultured cells of CMF in the presence of conditioned medium from ASCs for 8 days. Consistent with the stimulatory effect observed in tube assay, addition of conditioned medium collected from ASCs upregulated the formation of CD31+ vessel-like structures. However, these capillary-like structures were less branched than vascular structures formed in CMF co-culture with ASCs. Further, capillary-like structures formed in the presence of conditioned medium were less stable and disassemble after 15 days of culture. This evidence suggests that the angiogenic factors secreted by ASCs are necessary but might not be sufficient for the formation of stable vessel-like structures of cultures of CMF cells. Presumably, the contact interaction between cells of CMF and ASCs is required for the stabilization of the newly formed vessel-like structures.

Vascular structures formed in CMF co-cultured with ASCs. (
ASCs' contact with vessels improves their stability
To elucidate if ASCs contact vessels, outgrowing from CMF, we co-cultured CMF cells with ASCs labeled with lipophylic red fluorescent dye CM-DiI-Cell Tracker (Fig. 6A). After 8 days of co-culture we stained outgrowing vessels with antibody against CD31 (green fluorescence in Fig. 6A). In most cases CD31+ tubules did not contain DiI tracker, suggesting their CMF origin, while DiI-labeled ASCs located adjacently to those CD31+ structures. These data indicate that ASCs stabilize vessel-like structures that originate from CMF.
To confirm that the capillary-like structures indeed originated from CMF, we co-cultured CMF cells with mitC ASCs. Mitomycin-C blocks DNA synthesis and cell proliferation, but does not interfere with protein synthesis as shown by Tomasz and colleagues 26 and confirmed by our immunofluorescent data with antibody against PCNA (data not shown). After 8 days the number of vessel-like structures observed in co-culture of CMF with mitC ASCs was about 3.5 times higher than in CMF cultured alone. Consistent with the above observations, the number of vessel-like structures in co-culture of CMF with mitC ASCs was also significantly higher than in CMF cultured in the presence of conditioned medium from ASCs. Given that ASCs express pericyte marker NG2, as well as connexin 43 (Fig. 3), which is known to be an important component of gap junctions mediating interactions between endothelial cells and pericytes, 23 we suppose that pericytes from ASCs form direct cell–cell contacts with endothelial cells from CMF, thus stabilizing the newly formed vascular structures.
ASCs enhance endothelial differentiation of progenitor cells from CMF
To determine whether the vessel-like structures are formed from the preexisting endothelial cells or they result from endothelial differentiation of progenitor cells present in CMF, as well as to prove that perivascular cells within observed vascular structures do not originate from CMF, we depleted the heterogeneous CM fraction of mature endothelial cells and pericytes using immunomagnetic sorting (Fig. 6). CD31−/NG2− population of CMF was co-cultured with ASCs for 8 days. Surprisingly, the number of CD31+ vessel-like structures formed in this co-culture was similar to the co-culture of ASCs with total CMF population (Fig. 6). CD31−/NG2 −CMF fraction contains progenitor cells as shown in Figure 4 and by others,27,28 and our present data suggest that ASCs might stimulate endothelial lineage differentiation of these progenitor cells. It also indicates that perivascular cells within the formed vascular structures originate from ASCs (Fig. 6).
As expected, we observed two times increase in the amount of vessel-like structures in co-cultures of ASCs with CD31+/NG2 +-enriched population of CMF compared with co-culture of ASCs with CMF. Immunofluorescent staining with antibody against CD31 (Fig. 6C, red fluorescence) revealed that vessel-like structures in these co-culture were thicker and more branched than CD31−/NG2− structures.
Discussion
The concept of regenerative medicine using autologous stem cells for cell therapy and tissue engineering has emerged over the past years. 29 Clinical trials suggest that stem cells may contribute to myocardial regeneration after infarction.30–32 Most of the clinical trials used bone marrow–derived or blood circulating progenitor cells that are limited in number or difficult to harvest. Recently, ASCs were shown to contain multipotent mesenchymal stem cells that could be easily isolated and expanded in vitro.1,4,33 The plasticity of ASCs and their ability to differentiate into osteoblasts, myoblasts, chondrocytes, and adipocytes are well documented. 1 It was also demonstrated using animal models that administration of ASCs locally or systemically stimulates regeneration of the ischemic muscles and their neovascularization.3,5,34–36 The potential of ASCs to exert paracrine effects on endothelial cells in vitro resulting in enhanced capillary-tube formation suggests that these cells could be a novel source of cells for cell therapy of ischemic tissues.3,4 In the present study, using co-culture system, we demonstrate that ASCs stimulate formation and stabilization of CD31+ vessel-like structures from a mixed population of cells isolated from postnatal hearts (CMF). Our data indicate that angiogenic potential of ASCs is not limited to paracrine effects, as it was suggested in recent work. 22 Results obtained in experiments on co-culturing ASCs with CD31+/NG2+ or CD31−/NG2− cells of the CMF suggest that not only do ASCs stimulate formation of CD31+ vessel-like structures from preexisting endothelial cells, but also ASCs possibly induce differentiation of progenitor cells present in the CMF. We show that cell–cell contacts appear to be important for the formation of numerous stable vessel-like structures in vitro. Because expression of connexin 43 was shown to be crucial for vessel stabilization due to establishing endothelium cell interaction with pericytes and SMCs,23,37 we suppose that cell–cell contacts between CMF and ASCs, mediated by gap junctions expressing connexin 43, could be involved in the formation of stable vessel-like structures.
The present data confirm previous findings that ASCs produce angiogenic and anti-apoptotic factors3,4,38–40 such as VEGF, HGF, its receptor c-met, bFGF, PDGFB, and TGFβ. According to the results of Potier et al., the mesenchymal stromal cells' viability is not affected by the short-term (48 h) hypoxia. 41 Using this model of hypoxia, we investigated the difference in gene expression profile in ASCs under hypoxia, which mimics the ischemic environment in vivo. We show that gene expression of all of these factors is increased in response to hypoxia 1.7–4.1-fold, which is of great importance because introduction of ASCs into ischemic muscle could potentially mediate better survival of the injured tissues and their neovascularization. Obtained results are consistent with some recent reports, in which it was shown that many of progenitor cell populations, including cultivated mesenchymal stromal cells, respond to hypoxic stimuli by upregulating angiogenic and antiapoptotic factors' gene expression of VEGF, HGF, bFGF, and others,3,38,40,42 mostly through stabilization of the hypoxia-inducible factor (HIF)-1α protein. 39 Activation of the HIF-1–mediated signaling events by hypoxia is complex and regulated by a cascade of molecular events that are reviewed in detail in a recent paper. 43 Under hypoxic conditions, HIF-1α accumulates, dimerizes with HIF-1β, recruits the coactivators CBP/p300, and activates the transcription of genes encoding angiogenic cytokines, including VEGF, angiopoietins, PDGFB, and TGFβ. 44 The protein products of these genes are secreted and bind to their cognate receptors, which are located on the plasma membrane of vascular endothelial cells, pericytes, and circulating angiogenic cells.
Angiogenesis is characterized by activation of endothelial cells, their migration, proliferation, and capillary sprouting. In vivo migrating endothelial sprouts are frequently accompanied by migrating pericytes or SMCs. Vessel wall maturation is completed by recruitment of perivascular cells resulting in stabilization of the newly formed vessels.45,46 It was previously shown by Traktuev that a population of ASCs contains pericytes. In the present work we also demonstrate that ASCs contain a population of NG2+ cells, corresponding to pericytes. 22 Further, cells originating from ASCs align along the newly formed CD31+ vessel-like structures and stabilize them in vitro, that is, function as pericytes. We also show using Matrigel implantation model of angiogenesis in vivo that ASCs cultured in hypoxic conditions have an ability not only to stimulate blood vessels growth into Matrigel plugs but also to promote the remodeling and stabilization of the newly formed vessels better than ASCs cultured in standard conditions.
Role of ischemia and hypoxia-inducible expression of growth factor genes, particularly in mesenchymal stromal cells, for arteriogenesis and collateral enlargement was discussed in numerous studies.38,47 The processes of arterialization and collateral enlargement rely on endothelial cell activation, vascular cell proliferation, and rearrangement of the extracellular matrix, and can be triggered by different stimuli, including some growth factors, inflammation cytokines, and hemodynamic stress. Growth factors and receptors such as PDGFB, PDGF-β receptor, bFGF, and angiopoietin-1 are important in the perivascular cell recruitment process, which is extremely necessary for proper vessel maintenance and microvascular network architectures. 48 Interstitial progenitor cells migrate to the PDGFB gradient, invest the capillary channels, and differentiate into smooth muscle phenotype. In response to TGFβ, SMCs contribute to arterialization, presumably through proliferation, differentiation, and migration to capillaries. 49 TGFβ is also required to coordinate the upregulation of the SMC differentiation program, including smooth muscle myosin heavy chain expression. 50 All the mechanisms of actions, including secretion of factors stimulating angiogenesis and vessel maturation as well as formation of cell–cell contacts, could be beneficial for the stimulation of vascularization in ischemic tissues by ASC administration.
Conclusions
ASCs are suggested to be potential candidates for cell therapy of ischemic tissues due to their ability to stimulate blood vessel growth. Studying the mechanisms of ASCs' angiogenic action, we show that ASCs produce angiogenic and anti-apoptotic growth factors, and their secretion is significantly enhanced by hypoxia. However, ASCs enhance blood vessel growth not only by production of paracrine-acting factors but also by promoting the endothelial differentiation of progenitor cells. Moreover, ASCs contain a subpopulation of NG2-expressing pericytes, which stabilize the newly formed vessel-like structures in vitro. All these characteristics of ASCs could be beneficial for the stimulation of angiogenesis in ischemic conditions.
Footnotes
Acknowledgments
This work was supported by CRDF Grant 07–04-91141 and by a grant (No. 02.435.11.3022) from Russian Government.
Disclosure Statement
The authors state that no competing financial interests exist.