
Abstract
Pheochromocytomas are widely believed to induce cardiomyopathy via hypersecretion of catecholamines, including norepinephrine (NE). NE can have direct cardiomyocyte toxicity and/or can stimulate myocardial remodeling secondary to the induction of hypertension. Yet, the development of cardiomyopathy is not entirely related to catecholamine dose or the extent of hypertension. To explore these effects, we engineered a polymeric encapsulation system to control PC12 cell kinetics and NE release in vitro and in vivo. Primary neonatal rat cardiomyocytes incubated with pheochromocytoma-conditioned media exhibited greater cytoskeletal changes than myocytes cultured with identical doses of NE alone, including more profound dose-dependent decreases in desmin, β-tubulin, and vinculin and upregulation of dystrophin. Cardiomyocyte contractility was 29 ± 6% greater at given levels of NE release. Agarose-encapsulated PC12 cells retain cell viability and structural integrity in vivo. These implants induce a 30% greater degree of cardiac enlargement as compared to pumps releasing equivalent doses of NE. Protein level alterations observed in vitro were mirrored in vivo after implantation of encapsulated cells or NE pumps for 28 days. Together, these data suggest that pheochromocytoma-induced cardiomyopathy is not solely a catecholamine-mediated event; rather, the pathogenesis of this dilated cardiomyopathy appears to be dependent upon secondary factors unexamined to date.
Introduction
Clinical experience also questions this assumption: up to 36% of patients with pheochromocytomas exhibit no hypertension or abnormal catecholamine levels, but develop PICM.1,12 This clinical paradox, together with data suggesting the potency of noncatecholamine endocrine factors in the pathogenesis of cardiomyopathy, led us to hypothesize that the cardiotoxic effects of pheochromocytoma are not solely catecholamine mediated, and that rather a complex of tumor-secreted factors modulates cardiomyocyte remodeling even at noncardiotoxic levels of catecholamines.
Previous attempts at a direct comparison between the effects of pheochromocytomas and NE alone in vivo have been hindered by a lack of control over catecholamine secretion rate, resulting in supraphysiologic levels of circulating NE. 12 These models not only invariably induce cardiomyopathy but may also mask any secondary factors at work. While these models yield insight into the role of catecholamines in PICM, they fail to explain the progression of the disease in patients with undetectable or low degrees of catecholamine elevation, 12 and the reversibility of PICM seen in early stage pheochromocytoma patients.7,13–17
To explore the effects of NE alone versus pheochromocytoma-secreted factors, neonatal rat cardiomyocytes were exposed to NE or conditioned media of pheochromocytoma (PC12) cells with contractility and cytoskeletal remodeling tracked. An in vivo model using controlled release of pheochromocytoma factors or NE was also developed. Tumor secretions increased the contractility of and induced greater remodeling effects on isolated cardiomyocytes than equivalent doses of NE alone, and had similar effects on hearts in situ when cells and NE pumps were implanted in vivo. The effects of tumor secretions were observed to outweigh those of NE alone in cardiomyocyte and cardiac function and remodeling. Such findings validate the synergistic effects of pheochromocytoma-secreted products with catecholamines in general and speak to the more general etiology of cardiomyopathy and tumor pathology.
Materials and Methods
Cell culture and characterization
Rat pheochromocytoma cells (PC12; ATCC, Manassas, VA) were cultured in F12K media (ATCC) with 10% horse serum (Hyclone, Logan, UT), 5% fetal bovine serum (Hyclone), and 100 units/mL penicillin/streptomycin (Invitrogen, Carlsbad, CA). 18 Mouse pheochromocytoma cells (MPC 4/30/PRR, kindly provided by Dr. Tischler; New England Medical Center, Boston, MA) were maintained in RPMI 1640 media (Invitrogen) with 10% horse serum (Hyclone), 5% fetal bovine serum (Hyclone), and 100 units/mL penicillin/streptomycin (Invitrogen). 19 Both cell lines were grown at 37°C and 5% CO2, and passaged every 2 days.
Cardiomyocyte isolation and contractility
Cardiomyocytes were obtained from the ventricles of 2-day-old neonatal rats using established methods 20 under an institute-approved protocol. Freshly isolated cells were preplated for 60 min to enrich for cardiomyocytes, and then plated 3 × 105 cells/cm2 in laminin-coated 60-mm Petri dishes. Culture media was conditioned by secretions of PC12 cells using beads of agarose-encapsulated PC12 and determining equivalent NE concentrations, based on drug delivery studies (Fig. 1). Identical controls were cultured in media supplemented with NE. On culture day 4, contractility was assessed on autocontracting cardiomyocytes using an optically transparent, environmentally controlled test chamber, an inverted microscope (Zeiss Axiovert 200M; Zeiss, Oberkochen, Germany), and a digital video camera (Sony XCD-X710, Tokyo, Japan). Machine visual screening software (Visible Discovery; Reify, Cambridge, MA) calculated the aggregate change index (ACI), a measure of contractility-induced change in the spatial organization of light intensity (i.e., a measure of contractile amplitude). 21 The contractile frequency of cardiomyocytes was quantified as the mean value of the peaks of the ACI for each concentration. The ACI function does not require image segmentation and therefore is applicable to high-density neonatal cardiomyocyte cultures in which cell edges cannot be readily localized.
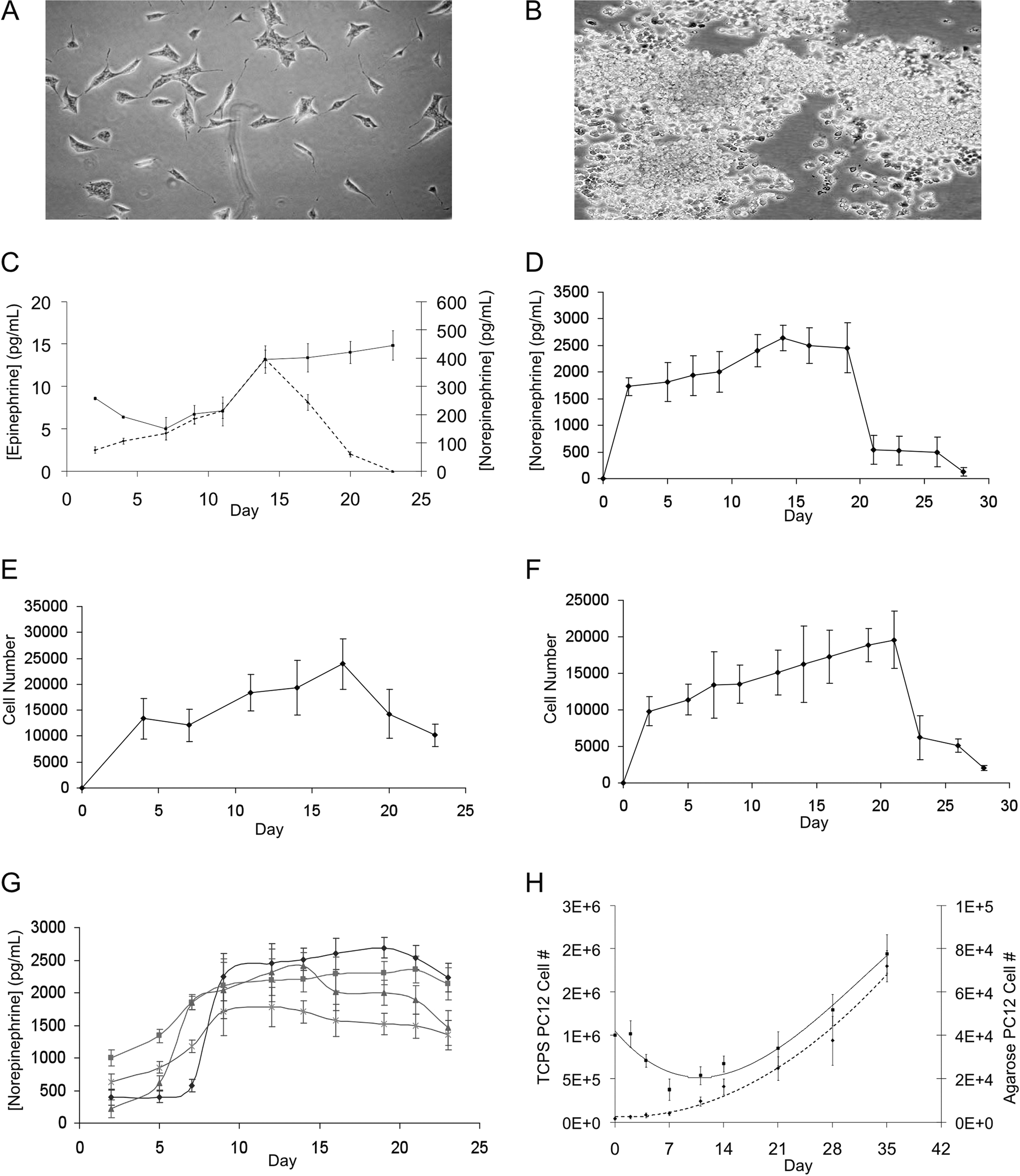
Encapsulation of MPC and PC12 cells does not affect their growth or secretory ability. Representative images of MPC (
Cardiomyocyte protein analysis
Confluent cardiomyocytes were scraped off tissue culture polystyrene plates (TCPS), counted, aliquoted (1 × 106 cells/vial), and incubated with lysis solution (20 mM Tris, 150 mM sodium chloride [NaCl], 1% Triton X-100, 0.1% sodium dodecyl sulfate, 2 mM sodium orthovanadate, 2 mM phenylmethyl-sulfonyl fluoride (PMSF), 50 mM NaF, and protease inhibitor cocktail; Roche, Basel, Switzerland). Samples were boiled for 5 min with Laemmli sample buffer and separated on Nupage gels (Invitrogen). Cardiomyocytes incubated in growth media without PC12-conditioned media or NE served as controls. Proteins were transferred onto membranes using iBlot stacks (Invitrogen), blocked, and incubated with dystrophin polyclonal (Abcam, Cambridge, MA), desmin polyclonal (Abcam), β-tubulin monoclonal (Abcam), or RPL32 polyclonal (Aviva Systems Biology, San Diego, CA) antibody. Membranes were incubated with a goat anti-rabbit or goat anti-mouse monoclonal antibody (Santa Cruz Biotechnology, Santa Cruz, CA) at a 1:1000 dilution for 2 h at room temperature. The membranes were then washed in phosphate-buffered saline (PBS)-T (PBS with 0.05% Tween 20) and incubated with the Western Supersignal Femto kit (Pierce, Rockford, IL). The membranes were exposed on a FluorChem SP (Alpha Innotech, San Leandro, CA).
Encapsulation and survival of MPC and PC12 cells in agarose beads
Agarose beads were developed as previously described. 22 PC12 cells were scraped, centrifuged, and resuspended in a preheated solution of 2.5% (w/w) agarose (Type VII; Sigma-Aldrich, St. Louis, MO) in 0.9% NaCl. The mixture was drawn into a pipette, and 10 μL aliquots sheared into a mineral oil bath (∼600 mL). The beads were separated using a 1000 μm pore size nylon mesh (Small Parts, Miramar, FL) and washed with PBS. Cell proliferation within agarose bead was assayed with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (Sigma-Aldrich, St. Louis, MO) per manufacturer's instructions. The optical density was read on a microplate photometer (BioTek Instruments, Winooski, VT) and compared to a standard curve with known numbers of PC12 cells. Epinephrine, NE, and dopamine secretion rate was assayed daily from media containing agarose-encapsulated cells by ELISA (Rocky Mountain Diagnostics, Colorado Springs, CO). Ascorbic acid (50 μg/mL) was added to the media to minimize oxidation of catecholamines.
Animal experiments
Female Sprague-Dawley rats (8 weeks old, 250 g) were obtained from Charles River (Wilmington, MA). A total of 20 PC12-encapsulated agarose beads were implanted in the retroperitoneal cavity adjacent to the kidneys, to mimic the spatial release of pheochromocytoma cells, for an overall secretion rate of 9.2 ng/day. Osmotic pumps (Model 2004; Alzet, Cupertino, CA) delivering 0.25 μL/h were loaded with 9.06 μM solution of NE in acidic saline (0.1 mg/mL ascorbic acid in saline). Control animals received osmotic pumps loaded with the ascorbic acid/saline solution. No statistical difference in cardiac function was found in rats implanted with empty agarose beads and nonsurgical, nonimplanted rats. All animal studies were performed in accordance with protocols approved by the IACUC at the Massachusetts Institute of Technology. Catecholamine levels present in rats before were tracked through weekly blood draws via the retroorbital plexus. Plasma catecholamines were quantified by ELISA (Rocky Mountain Diagnostics).
Cardiomyocyte protein extraction
Hearts were excised, washed in phosphate buffered saline (PBS), and the left ventricle (LV) separated and frozen in liquid nitrogen. Sections (2 × 2 × 2 mm) were weighed and sectioned in a microtome, and lysed in 1 mL solution (20 mM Tris, 150 mM NaCl, NaOH pH 8.0, pH with NaOH, 1% Triton X-100, 0.1% sodium dodecyl sulfate, 2 mM sodium orthovanadate, 2 mM PMSF, 50 mM NaF, and protease inhibitor cocktail; Roche). Proteins were isolated as described above. Membranes were incubated with dystrophin polyclonal antibody (Abcam), desmin polyclonal antibody (Abcam), β-tubulin monoclonal antibody (Abcam), or RPL32 polyclonal antibody (Aviva Systems Biology). The secondary antibodies used were goat anti-rabbit or goat anti-mouse monoclonal antibody (Santa Cruz Biotechnology) at a 1:1000 dilution.
Histology
Rat hearts were excised, rinsed in PBS, blotted dry, and weighed. The hearts were then pressure perfused with PBS for 5 min followed by perfusion with 10% neutral-buffered formalin until interpreted as visibly firm and pale. The hearts were placed in 10% neutral-buffered formalin for 18 h, subsequently processed and fixed in paraffin (Polysciences, Warrington, PA), and serial coronal sections were cut and stained with Masson's Trichrome (American HistoLabs, Gaithersburg, MD).
RNA preparation and semiquantitative RT-PCR
Isolated hearts were excised and perfused with PBS (Invitrogen) for 5 min. A biopsy punch was used to section out an 8 × 8 mm section of the left ventricle. Total RNA was isolated with the use of Qiashredder and RNeasy spin columns, including chromosomal DNase digestion (Qiagen, Germantown, MD). Twenty micrograms of total RNA was reverse transcribed into cDNA with the use of oligo(dT)24 primers containing a T7 RNA polymerase promoter. First-strand cDNA was synthesized using Taqman RT-PCR kit (Applied Biosystems, Foster City, CA). Specific primers were designed using Primer3. 23 Real-time PCR analysis was performed with an Opticon Real-Time PCR Machine (Biorad, Hercules, CA) using SYBR Green PCR Master Mix (Applied Biosystems). The resulting data were analyzed with the complementary Opticon computer software (Biorad). All samples (n = 5 per group) were measured in triplicate, and the expression level was normalized to GAPDH expression.
Results
Agarose encapsulation of MPC and PC12 cells
MPC (Fig. 1A) and rat pheochromocytoma cells (PC12, Fig. 1B) were encapsulated in agarose beads. Both cells continued to grow after encapsulation and released quantifiable levels of catecholamines. Whereas encapsulated PC12 cells retained a narrow range of cell number and NE release (Fig. 1D, F), MPC were less viable and the NE release highly variable in vivo (Fig. 1C, E). Dopamine levels in PC12 cells were undetectable, 24 as were epinephrine levels, given the low or nonexistent expression of phenylethanolamine-N-methyltransferase, the enzyme responsible for converting NE to epinephrine. 25 The lack of epinephrine or dopamine secretion makes PC12 cells then an ideal side-by-side comparison to the NE-secreting pumps. Varying formulations of agarose beads were evaluated, including the addition of collagen and use of cell aggregates, to optimize cell growth and secretory function. Addition of collagen and use of cell aggregates induced minimal changes in their secretory ability (Fig. 1G). Agarose-encapsulated PC12 cells at a density of 4 × 106 cells/mL maintained similar growth kinetics and metabolism to PC12 cells grown on polystyrene dishes (TCPS) (Fig. 1H). NE secretion from encapsulated cells (29.24 ± 1.09 ng) was statistically indistinguishable from cells cultured on TCPS (30.29 ± 0.73 ng, p > 0.05).
Cardiomyocyte cytoskeletal contractility and remodeling
Primary neonatal rat cardiomyocytes were incubated with either PC12-conditioned or NE-containing media. NE induced a dose-dependent increase in cardiomyocyte contraction force and frequency (Fig. 2A, B). Encapsulated PC12-conditioned media exhibited similar increases in contractile effect to equivalent NE doses below 0.1 nM, greater increases at doses of 0.1 to 1 nM, and significantly greater increases at 5 and 10 nM (Fig. 2). PC12-conditioned media induced an almost twofold greater contractile effect at 5 nM NE (p < 0.01). The altered chronotropic effects of PC12-conditioned media versus identical doses of NE were dose dependent. Expression of cytoskeletal proteins followed suit (Fig. 3). The effects of dose-dependent effects of NE were enhanced by PC12-conditioned media; desmin (−35 ± 8% vs. −22 ± 7%), β-tubulin (−34 ± 9% vs. 8 ± 12%), and vinculin (34 ± 5% vs. 1 ± 3%) were all reduced and dystrophin increased to a greater extent at the lower levels of NE release by the PC12 media.
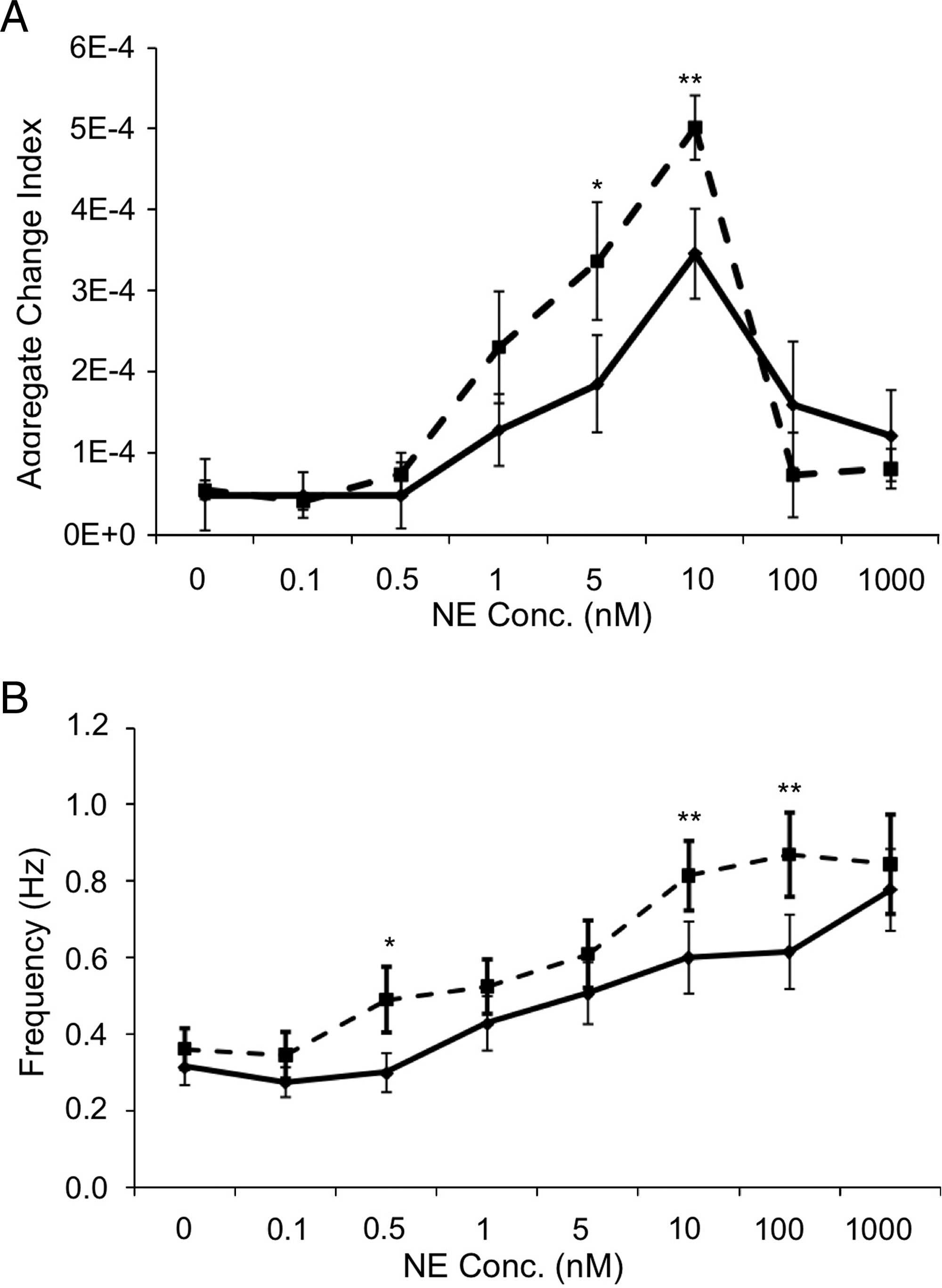
PC12-conditioned media induces a greater contractile effect on cardiomyocytes than identical doses of NE alone. (
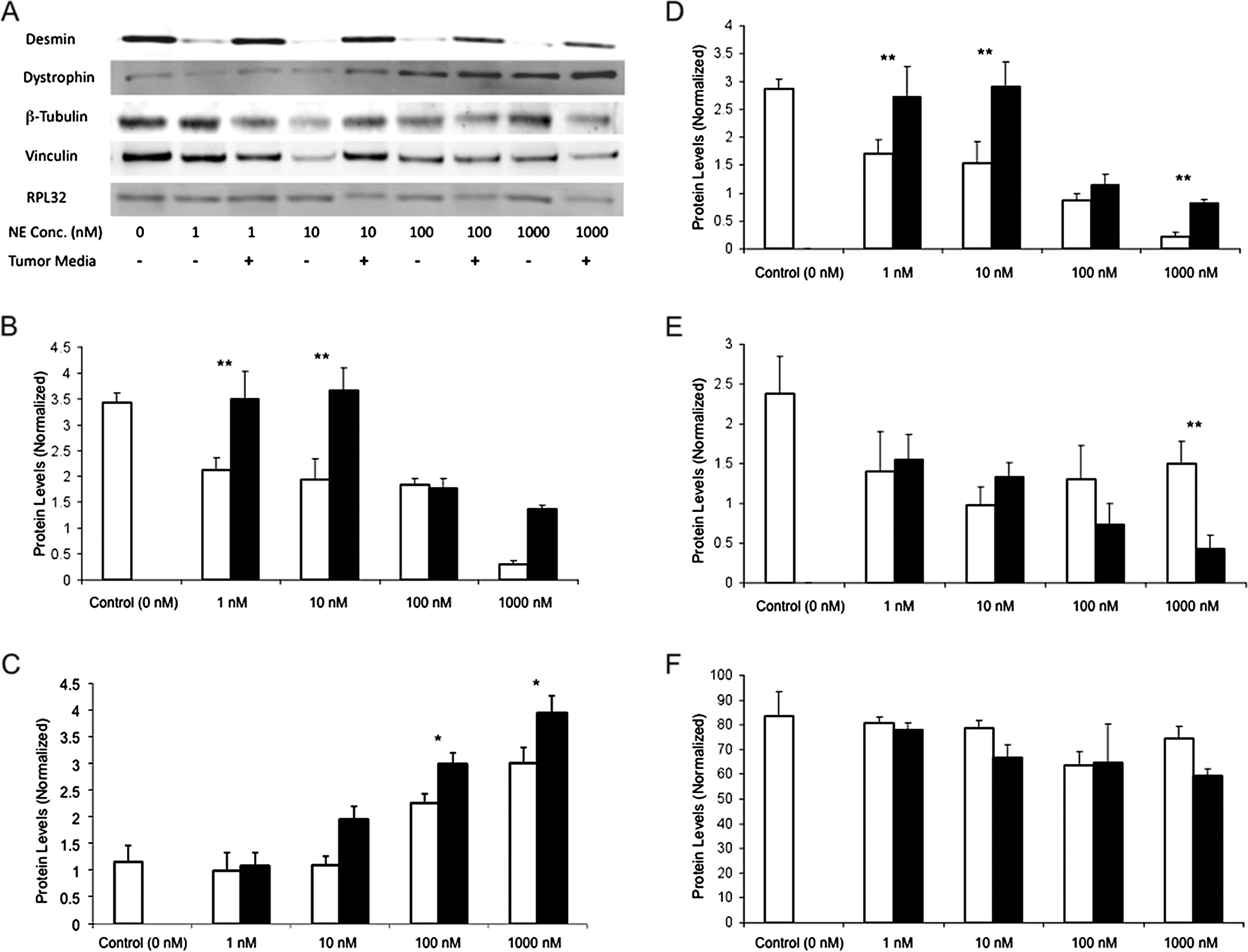
Cardiomyocytes incubated with PC12-conditioned media undergo differential cardiomyocyte remodeling from those incubated with identical doses of NE alone. (
In vivo model and cardiac morphology
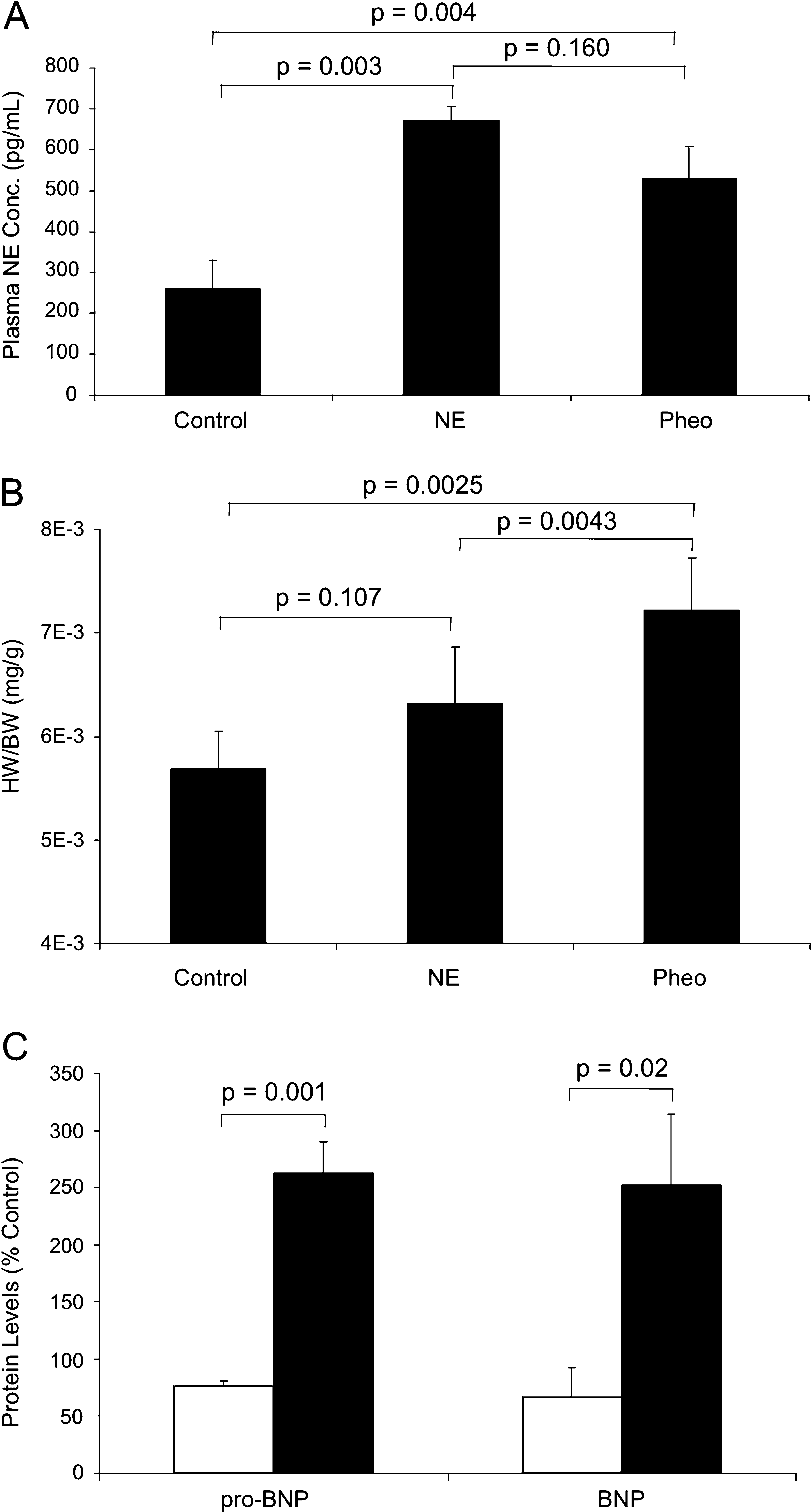
Preliminary data showed that encapsulated PC12 cells survived longer and secreted greater levels of NE than encapsulated MPC for the time period of interest; encapsulated PC12 cells were therefore used in subsequent animal studies. Implantation of agarose-encapsulated PC12 cells in the retroperitoneal cavity of rats demonstrated the ability of PC12 cells to secrete catecholamines in vivo without eliciting an immune reaction. Histological analysis revealed the formation of a fibrous capsule (120 μm) at the bead surface, which did not inhibit their secretory ability, based on circulating plasma NE levels (Fig. 5E, F). NE-secreting osmotic pumps were implanted to match the NE secretory rate of implanted encapsulated PC12 constructs. As validation of our delivery methods, the implantation of osmotic pumps secreting 9.2 ng NE per day induced an increase in plasma NE concentration by 310 ± 62 pg/mL in rats. Similarly, agarose-encapsulated PC12 cells secreting 9.2 ng NE per day increased plasma NE concentration by 250 ± 49 ng/mL in rats (Fig. 4A).
Agarose-encapsulated PC12 cells retain cellular integrity and secretory ability, and induce cardiac pathology. (
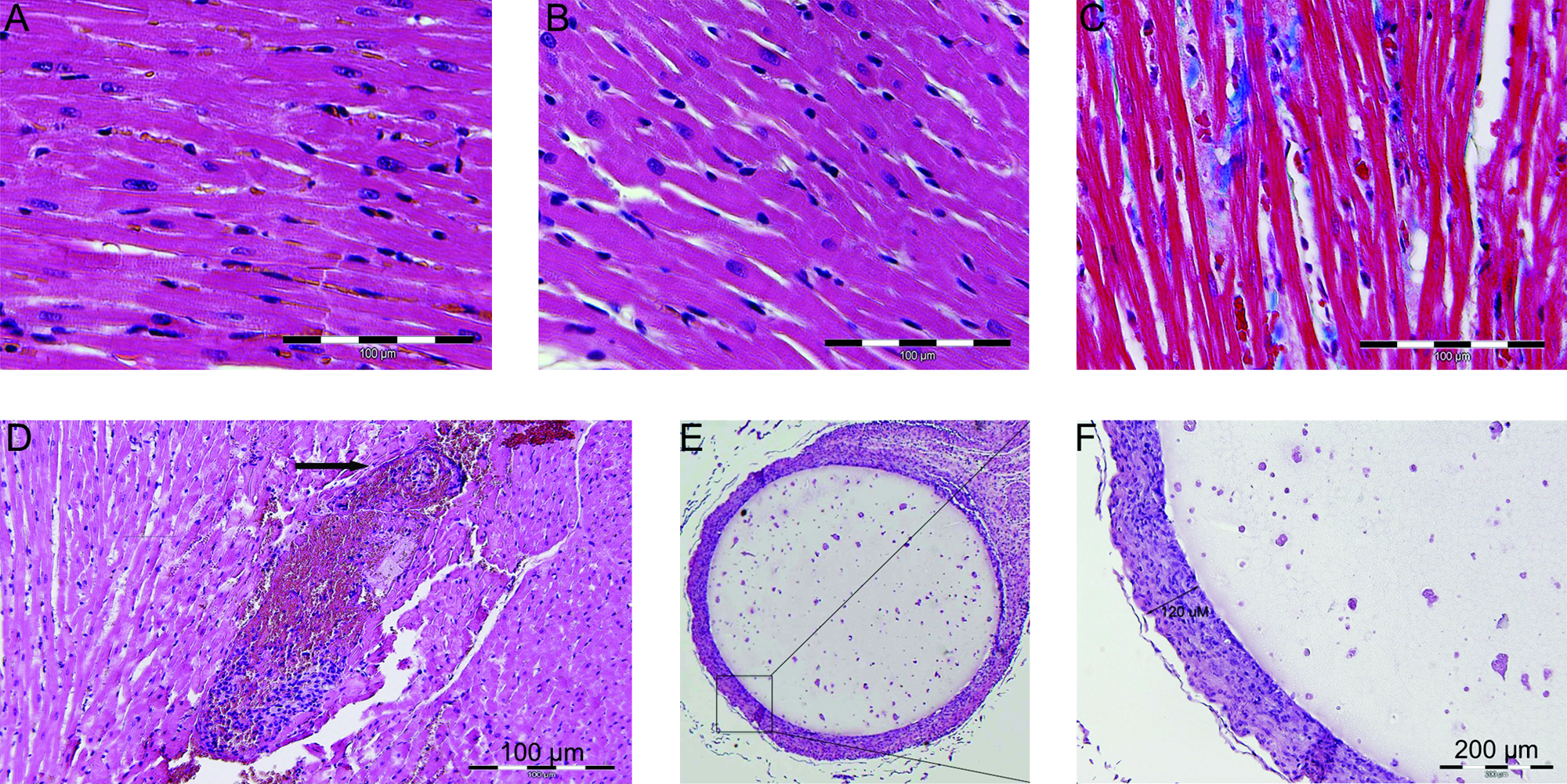
Implanted pheochromocytoma cells induce cardiac pathology suggestive of dilated cardiomyopathy. Representative left ventricular light microscopy image of control (
Heart weight normalized to body weight was used as a measure of cardiac remodeling after 28-day implantation. The hearts of Pheo animals were 31% larger than controls (p < 0.01) and 20% greater than NE rats (p < 0.01), the latter statistically indistinguishable from controls (Fig. 4B). Brain natriuretic peptide (BNP) and its precursor, proBNP, are released from the cardiac ventricles in response to pressure overload, particularly in the context of heart failure.26,27 proBNP and BNP mRNA levels were elevated 250 ± 35% and 280 ± 20%, respectively, above control rats 28 days after implantation. Rats implanted with NE pumps secreting identical doses of NE exhibited mRNA levels indistinguishable from controls (78 ± 12% and 68 ± 11% vs. controls). These statistically significant changes highlight the ability of pheochromocytomas to induce cardiac pathology with accelerated kinetics versus catecholamines alone. Histological analyses of cardiac tissue showed greater fibrosis, vessel hemorrhage, and myofiber necrosis in Pheo versus NE and control rats (Fig. 5A–D).
In vivo cell and pump release kinetics and effects on cardiac function and cytoskeleton
Agarose-encapsulated PC12 cells or osmotic pumps secreting 9.2 ng NE per day were implanted retroperitoneally in rats for 28 days. Cytoskeletal protein expression changed in vivo as it had in culture, with greatest reductions in all proteins in cell-implanted rats and to a greater extent than equivalent amounts of NE alone. PC12-implanted rats exhibited greatest changes in these proteins; desmin (30 ± 7% decrease vs. control; 31 ± 9% decrease vs. NE), β-tubulin (80 ± 15% decrease vs. control; 77 ± 9% decrease vs. NE), dystrophin (23 ± 9% decrease vs. control; 9 ± 2% decrease vs. NE), and vinculin (26 ± 11% decrease vs. control; 19 ± 9% decrease vs. NE; Fig. 6).

Implanted pheochromocytoma cells induce cardiomyocyte remodeling similar to in vitro observations. Cardiac protein levels after implantation of agarose-encapsulated PC12 cells and NE-secreting pumps for 28 days. Each western blot is followed by a densitometry analysis normalized to RPL32 levels. Rats implanted with NE-secreting pumps exhibited desmin levels similar to controls (1 ± 2% increase vs. control), while β-tubulin, vinculin, and dystrophin levels decreased (10 ± 3% vs. control, 9 ± 2% vs. control, and 16 ± 4% average, respectively). Rats with PC12 implants exhibited decreases in all four proteins relative to controls (desmin 30 ± 7%, dystrophin 23 ± 9%, β-tubulin 80 ± 15%, and vinculin 26 ± 11% vs. RPL32-normalized controls).
Discussion
To the best of our knowledge, this study is the first to analyze the direct impact of NE versus the complete complex of factors secreted by pheochromocytomas on primary cardiomyocyte physiology. Traditional models of PICM have relied upon the implantation of free PC12 cells resulting in uncontrolled cell growth and release of catecholamines.12,28 Although these studies provided important information on the cardiotoxic effects of catecholamines and the ability of large pheochromocytomas to induce cardiomyopathy, the magnitude of NE released may have masked the effects of secondary factors secreted by these tumors. Moreover, the rapid growth of injected free PC12 cells renders them unsuitable in the study of nascent tumors, which often induce cardiomyopathy in the context of low levels of catecholamines.
Pheochromocytomas have long been postulated to induce cardiomyopathy via hypersecretion of catecholamines. Excessive adrenergic stimulation has been established as cardiotoxic in multiple species. Mann et al. demonstrated the dose-dependent effects of NE resulting in increased cell death, decreased protein synthetic ability, and altered cardiomyocyte morphology suggestive of decreased contractility. 29 But the pheochromocytoma secretome also includes other known cardiotoxic factors, including TGF-β and IL-6. Limited data show that TGF-β induces a hypertrophic response in cultured ventricular cardiomyocytes and stimulates endogenous TGF-β release. 30 IL-6 and IL-6–like cytokines induce hypertrophic and antiapoptotic pathways in cardiomyocytes through Akt activation. 31
Freshly isolated neonatal cardiomyocytes were used to determine whether PC12-conditioned media induced a greater effect at the single-cell level than cardiomyocytes treated with identical doses of NE alone. Our data strengthen the hypothesis that there exist secondary factors released by pheochromocytoma cells that alter the pathogenesis of cardiomyopathy beyond the response seen with catecholamines alone, at low levels increasing function of and strain on cardiomyocytes, and at higher levels hastening loss of function. While an overload of catecholamines may be sufficient for the pathogenesis of PICM, we suggest that these are not wholly responsible, and may even be unnecessary for the development of cardiomyopathy in patients; these findings may also begin to explain the development of PICM in clinically silent and normo-adrenergic pheochromocytoma patients. 8 We also validated a new approach for studying the complete effects of pheochromocytoma on cardiomyocyte and cardiac physiology.
Although the role of the cytoskeleton as a stabilizing factor of cellular structure and functional integrity is well established, the structural basis of cardiomyopathy and heart failure is still not completely understood. 24 In recent years, mutations in at least five genes have been identified as causes of dilated cardiomyopathy and its clinical manifestations, including the contractile apparatus proteins dystrophin, 25 desmin,32,33 and vinculin. 34 Studies have also shown a strong association between elevated expression of microtubule proteins and altered cardiac contractility, 35 in which increased β-tubulin levels increase cardiomyocyte viscosity and stiffness. 36 Importantly, these contractile apparatus proteins are all also important biomarkers for structural cardiovascular disease: well-established pathways of cardiomyocyte remodeling in the context of heart failure involve compensatory cytoskeletal remodeling via the upregulation of β-tubulin, vinculin, dystrophin, and desmin. Our data suggest that in vitro studies done to date using NE alone to approximate the entire pheochromocytoma secretome may mask its true effects on cardiomyocyte remodeling, as these conditions elicit dramatically different patterns of damage and response. Indeed, whereas at high doses NE-induced protein changes approximate heart failure, complete PC12 media induced reductions (uncharacteristic of heart failure) in β-tubulin, vinculin, and desmin. Further, our data indicate that PC12 cell secretions induce a greater frequency of contraction than equivalent doses of NE alone and that encapsulation of PC12 cells does not affect their ability to secrete bioactive factors. More importantly, our observations demonstrate a different pattern of remodeling in response to the entire spectrum of pheochromocytoma-secreted factors. Here we show high degrees of correlation between all four proteins with each of the others examined when incubated with pheochromocytoma-conditioned media (Table 1). In marked contrast, only one set of proteins, desmin–dystrophin, was correlated in the context of NE alone. These data reveal a coordinated remodeling response, masked in simplified NE-only models, that begins to offer mechanistic insight into the pathogenesis of PICM.
PC12-
All six combinations of the four proteins examined displayed highly correlated alterations of expression when incubated in PC12-conditioned media relative to baseline. With NE treatment alone, only desmin and dystrophin were closely correlated to each other.
The protein cytoskeletal changes exhibited to a greater degree and in different patterns after PC12-conditioned media exposure than equivalent doses of NE alone corroborate the belief that pheochromocytomas secrete secondary factors that alter cardiac function in addition to and possibly independent of secreted catecholamines. Our cytoskeletal data do not indicate a mechanism of damage, but clearly demonstrate dose-dependent cardiomyocyte remodeling via mechanisms involved in cardiomyopathy and other cardiotoxic events. Further, the diverging cytoskeletal remodeling seen in dilated cardiomyopathy (DCM) versus our model of PICM might begin to explain the diverging reversibility of DCM and PICM.
We hypothesized that just as NE and complete pheochromocytoma secretions have differential effects on cardiomyocyte physiology in vitro, so too do these factors elicit different responses at the myocardial level in vivo. Traditional pheochromocytoma models have been hindered by the use of free pheochromocytoma cells whose unregulated growth and catecholamine secretion induce accelerated development of cardiomyopathy in the context of NE levels (40–75× basal levels) not always seen clinically.28,37 These models, while yielding insight into the role of catecholamines in PICM, fail to explain the progression of the disease in patients with no observed catecholamine elevation, in those with lower levels of catecholamines, 12 and the reversibility of PICM seen in early stage pheochromocytoma patients.7,15–17,38
We employed agarose encapsulation of PC12 cells to eliminate the unpredictable cell growth and NE release kinetics observed in previous systems. This approach has been used to success in studying other cell types, including Chinese hamster ovary cells. 22 In this study, we determined the optimal cell density per agarose bead to maximize nutrient and oxygen transport to embedded cells, while restricting cell growth (data not shown). This strategy resulted in predictable and reproducible cell growth and NE release kinetics mirroring that of PC12 cells grown in two-dimensional culture systems, allowing for direct comparison to osmotic pump-delivered NE alone over the period of study. Further, implantation of agarose-encapsulated PC12 cells demonstrated that the cells continued to grow and thrive in vivo for a period of 28 days while releasing predictable and quantifiable levels of NE.
The differential impact of pheochromocytoma-secreted factors versus NE alone was demonstrated in the ability of implanted PC12 cells to induce a greater degree of cardiomyopathy than equivalent doses of NE alone in vivo. Pheochromocytomas not only induced a greater increase in heart weight, but also did so in the presence of lower plasma NE concentrations than rats implanted with NE pumps. The increased effects of PC12 cells were further observed at the single-cell level as changes in cytoskeletal protein levels, similar in degree and direction of alteration to our in vitro observations. Pheochromocytoma cells induced significant decreases in desmin, β-tubulin, and vinculin expression, each to a greater degree than by NE alone. In rats implanted with pumps secreting identical doses of NE alone, expression of these proteins was reduced to a lesser extent, except in the case of desmin, which was indistinguishable from controls. Moreover, alterations in these cytoskeletal protein levels exhibited high degrees of correlation with each other, indicating coordinated myocardial remodeling responses, similar to cardiomyocytes in vitro. Together, these data demonstrate that pheochromocytoma secretions induce dissimilar cardiac changes at both the single-cell and whole-heart level in comparison to equivalent doses of NE alone.
In our novel models, PC12-conditioned media induced greater chronotropic effects and altered patterns of cardiomyocyte remodeling than equivalent doses of NE alone, suggesting net-additive effects of PC12 paracrine and endocrine factors on contractility and toxicity. This, together with the observation that pheochromocytomas induce dose-dependent cytoskeletal derangements different from those seen in both NE and organic and compensatory DCM, suggests a different pathologic process at play than seen in both the traditional model of clinical cardiomyopathy and the preferred basic approach of studying PICM in vitro. Our findings may yield a more physiologic model of PICM that may begin to explain disparities between clinical and bench models of this disease.
Footnotes
Acknowledgments
This work was supported by an NIH Pre-Doctoral Fellowship (to H.R.M.), an NIH Postdoctoral Fellowship F32-HL-84968 (to G.C.E.), a Sarnoff Cardiovascular Research Foundation Fellowship (to N.M.), NASA Grant NNJ04HC72G (to L.E.F.), and an NIH Grant R01 49039 (to E.R.E.). The authors wish to acknowledge Tayyba Anwar for her technical assistance, and A.M. Garakani and T. Miller of Reify Corporation for the generous loan of software (Visible Discovery) and hardware (Sony XCD-X700 camera) that enabled collection of the contractility data.
Disclosure Statement
No competing financial interests exist.