
Abstract
To regenerate the complex tissue such as bone–cartilage construct using tissue engineering approach, controllable differentiation of bone marrow stromal cells (BMSCs) into chondrogenic and osteogenic lineages is crucially important. This study proposes to test a minimum common osteochondrocytic differentiation medium (MCDM) formulated by including common soluble supplements (dexamethasone and ascorbic acid) used to induce chondrogenic and osteogenic differentiation. The MCDM coupled with supplemented growth factors was tested for its ability to differentiate BMSCs into osteogenic and chondrogenic lineages in both two-dimensional and three-dimensional culture systems. When transforming growth factor β3 was added to MCDM, BMSCs differentiated to chondrocyte-like cells, evidenced by the expression of glycosaminoglycans and type II collagen, whereas osteogenic differentiation was induced by supplementing osteogenic protein-1, resulting in detectable expression of osteopontin and osteocalcin. These chondrogenic and osteogenic differentiation markers were significantly enhanced in the three-dimensional cultures compared to the two-dimensional monolayer cultures. The results achieved in this study lay a foundation for future development of osteochondral graft, which could be engineered from bilayered scaffold with spatially loaded growth factors to control BMSC differentiation.
Introduction
Most studies involving bone marrow stromal cell (BMSC) cultures have focused on proliferation of autologous unspecialized BMSCs from bone marrow in a basal culture medium and their further differentiation into mesenchymal lineages, by employing lineage-specific induction media. In response to appropriate culture conditions, BMSCs can selectively form each of the mesodermal lineages—that is, adipocytes, osteoblasts, and chondrocytes.2,3 Optimal chondrogenic differentiation medium consists of high-glucose–Dulbecco's modified Eagle's medium (HG-DMEM) supplemented with transforming growth factor β, insulin, transferrin, selenium and linoleic acid premix (ITS), dexamethasone (DEX), ascorbate 2-phosphate (AA), sodium pyruvate (Na pyruvate), and antibiotics. 4 Promotion of osteogenic differentiation is achieved by supplementing HG-DMEM with osteogenic supplements consisting of AA, DEX, and β-glycerophosphate (β-GP). 2 Differentiated osteoblasts or chondrocytes have shown the potential to repair specific in vivo partial-thickness bone or cartilage defects, respectively, when these cells, in a mono-culture, were seeded into a tissue construct. 5 However, such constructs have the limitation of addressing diseases involving complex integrated tissue defects such as osteochondral defects.
Development of osteochondral constructs will ideally require integrated constructs supporting the growth and function of two different tissues—namely, cartilage and bone. 5 Although a well-defined culture medium exists for in vitro chondrogenesis and osteogenesis of BMSCs, there is no common osteochondrocytic differentiation medium that can support the multilineage differentiation of BMSCs in a biphasic scaffold for the development of osteochondral composite constructs.
The goal of this study was to evaluate a minimal common and essential culture medium supplemented with growth factors to direct BMSCs toward osteogenic and chondrogenic differentiation in two-dimensional (2D) and 3D cultures. Minimum common osteochondrocytic differentiation medium (MCDM) would be essential in developing a co-culture system for osteoblasts and chondrocytes to fabricate biphasic osteochondral constructs employing bioreactors. Cell permeable molecules such as DEX and ascorbic acid have both proven to be essential in inducing the differentiation of BMSCs to osteogenic and chondrogenic lineages. 6 Therefore, we designed a cell culture medium consisting of the basal medium (HG-DMEM + 10% FBS) supplemented with the common soluble supplements, DEX and ascorbic acid, for osteochondrocytic differentiation. Efficacy of the MCDM to promote chondrogenesis and osteogenesis was achieved by supplementing growth factors, that is, TGF-β3 for chondrogenesis7,8 and osteogenic protein-1 (OP-1) for osteogenesis.9–12
Materials and Methods
Procurement of samples and cell isolation
The bone marrow samples (n = 5) for isolation of mesenchymal stem cells (MSCs) were obtained from osteoarthritis patients (mean age, 65 years) undergoing elective knee and hip replacement surgery after informed consent. Ethics clearance for this project was approved by the Human Ethics Committee at Queensland University of Technology and Prince Charles Hospital. Bone marrow sample (10 mL) was mixed with 2 mL of heparin to prevent coagulation, and diluted with 20 mL of phosphate buffered saline (PBS). Cells were fractionated in a density gradient solution (Lymphoprep; Axis-Shield PoC AS, Oslo, Norway) by centrifuging at 800 g for 10 min. The resulting mononuclear cells appearing as a buffy interface were isolated and cultured in low-glucose–DMEM (LG-DMEM; Gibco–Invitrogen, Mt. Waverley, Australia) supplemented with 10% v/v fetal bovine serum (FBS; Thermo Scientific, Melbourne, Australia) and 1% v/v penicillin and streptomycin (P/S; Gibco-Invitrogen, Mt Waverley, Australia) at 37°C in a humidified atmosphere with 5% CO2. Initial medium change was performed on the fifth day to remove nonadherent cells, after which medium was replaced twice a week. The cells were passaged at approximately 80% confluence. After expanding through two passages, the cells were suspended in cryopreservation medium containing 10% dimethylsulfoxide (DMSO; Sigma-Aldrich, Castle Hill, Australia) and stored in liquid nitrogen for future use.
Two-dimensional and three-dimensional chondrogenic differentiation
Two-dimensional chondrogenic differentiation
BMSCs at passage 3 were recovered and plated as monolayers at a density of 2.5 × 104 cells/well in 24-well plates. At approximately 90% confluence the cells were grown in four types of media described in Table 1 (control medium [Ctrl], MCDM, chondrogenic medium, and MCDM supplemented with TGF-β3 [MCDM+TGF-β3]) for a period of 3 weeks. The control medium, also referred to as basal medium, comprised of HG-DMEM (Gibco-Invitrogen) supplemented with 10% FBS and 1% P/S; chondrogenic medium comprised of basal medium supplemented with 5 μg/mL ITS (Sigma-Aldrich, Castle Hill, NSW, Australia), 100 nM DEX (Sigma-Aldrich), 50 nM AA (Sigma-Aldrich), 1 mM Na pyruvate (Sigma-Aldrich), 0.35 mM proline (Sigma-Aldrich), and 10 ng/mL TGF-β3 (R&D Research, Bio Scientific, Gymea, Australia) 4 ; MCDM was formulated by supplementing 100 nM DEX and 50 nM AA to the basal medium. For MCDM+TGF-β3 treatment group, 10 ng/mL TGF-β3 was supplemented to MCDM.
LG-DMEM, low-glucose–Dulbecco's modified Eagle's medium; HG-DMEM, high-glucose–Dulbecco's modified Eagle's medium; DEX, dexamethasone; P/S, penicillin and streptomycin; ITS, insulin, transferrin, selenium and linoleic acid premix.
Three-dimensional chondrogenic differentiation of BMSCs in fibrinogen
BMSCs at passage 3 were resuspended in fibrinogen to form a mixture containing 2 × 105 cells/mL. Fibrin gels were prepared using a Tisseel Duo 500® kit (Baxter Healthcare, Sydney, Australia) according to the manufacturer's instructions. Each fibrin disk was polymerized by combining 0.25 mL calcium-activated bovine thrombin with 0.25 mL of the fibrinogen–cell suspension. The resulting 3D fibrin disk contained 100 U of thrombin, 18 mg of fibrinogen, and 1 × 105 cells in a total culture volume of 0.5 mL, the disk thickness being 4 mm. Disks containing cells were then subjected to the four types of culture media described in Table 1 for a period of 3 weeks.
Two-dimensional and three-dimensional osteogenic differentiation
Two-dimensional osteogenic differentiation
BMSCs at passage 3 were recovered and plated as monolayers at a density of 2.5 × 104 cells/well in 24-well plates. At full confluence the cells were treated for 3 weeks with four types of media described in Table 2 (Ctrl, MCDM, osteogenic medium, and MCDM supplemented with OP-1 [MCDM+OP-1]) to induce osteogenic differentiation.
OP-1, Osteogenic protein-1.
The composition of the control medium (Ctrl) and the MCDM was the same for both chondrogenic and osteogenic treatments. Osteogenic differentiation medium comprised of basal medium supplemented with 100 nM DEX, 50 nM AA, and 10 mM β-GP. For OP-1 treatment group, 40 ng/mL of OP-1 (R&D Research) was supplemented to the MCDM.
Three-dimensional osteogenic differentiation of BMSCs in HA/TCP scaffolds
Pieces of rectangular hydroxyapatite/tricalcium phosphate (HA/TCP) scaffolds (average pore size, 500 μm; porosity, 87%) were prepared by a polyurethane foam replica method. 13 Briefly, the ceramic slurry was prepared by mixing 160 g of HA with 40 g of β-TCP using a ball mill under wet conditions for 2 h. The resulting paste was then dried at 100°C for 24 h, followed by heat treatment at 900°C for 2.5 h with cooling and heating rates set at 5°C/min. The HA/TCP ceramics had an overall porosity of approximately 85–89%, the macropores typically 600 μm in diameter. The scaffold size used in this study was 5 × 5 × 3 mm (75 mm3). The scaffolds were placed into individual wells of a 24-well plate, and each scaffold was seeded with 2.4 × 105 cells/mL. Specifically, trypsinized BMSCs were resuspended in expansion medium at a final concentration of 2.4 × 106 cells/mL, and 100 μL of the cell suspension was applied to the scaffold (cell viability >95%, as determined by trypan blue staining). After 1 h of sedimentation each well was filled with the expansion medium and incubated at 37°C. After 24 h the scaffolds were transferred to wells containing the four different types of media (Table 2) and maintained for 3 weeks.
Histological analysis
Fibrin gel samples were harvested after 3 weeks of culture and fixed in 4% para-formaldehyde solution, dehydrated with 100% ethanol, and then embedded in paraffin. For hematoxylin and eosin staining, tissue sections were de-paraffinized with xylene and ethanol before Mayer's hematoxylin and eosin staining was applied. Chondrogenic matrix deposition was observed by staining the sections with 0.5% alcian blue (pH 1.0) for 3 min to visualize the sulfated glycosaminoglycans (GAGs).
Immunohistochemical analysis
For immunohistochemical staining, tissue sections were de-paraffinized and treated with proteinase K for 10 min, and then 3% hydrogen peroxide was added to suppress endogenous peroxidase activity. For chondrogenesis samples polyclonal antibody to type II collagen (1:100 dilution; Lab Vision, Fremont, CA) or aggrecan (1:100 dilution; Lab Vision) was added to the slices and incubated at the room temperature for 1 h. Sections were then incubated with a biotinylated swine-anti-mouse, rabbit, goat antibody (Dako Multilink, Carpinteria, CA) for 15 min, and then incubated with horseradish perioxidase–conjugated avidin–biotin complex for 15 min. Antibody complexes were visualized by the addition of a buffered diaminobenzidine substrate for 4 min. The reaction was stopped by immersion and rinsing of sections in PBS. Sections were lightly counterstained with Mayer's hematoxylin for 40 s, rinsed with running tap water for 3 min, dehydrated with ascending concentrations of ethanol solutions, cleared with xylene, and mounted with a coverslip using DePeX mounting medium (BDH Laboratory Supplies, Poole, England).
Scanning electron microscopy (SEM)
After 3 weeks' incubation in differentiation media, the scaffolds were rinsed three times with PBS followed by fixation in 3% glutaraldehyde solution overnight. The scaffolds were then washed in 0.1 M cacodylate buffer, postfixed in 1% aqueous osmium tetroxide for 1 h, dehydrated through a series of ascending ethanol solutions (50%, 70%, 90%, and 100%), and followed by two changes of 100% amyl acetate for 15 min each. After critical-point drying with CO2, the samples were sputter coated with gold and examined with a Quanta 200 Scanning Electron Microscope (FEI, Hillsboro, OR).
Reverse transcription–polymerase chain reaction (RT-PCR) analysis
Total RNA from 2D and 3D cultures was isolated using Trizol (Sigma-Aldrich) following the manufacturer's instructions. Samples were treated with DNase to remove any genomic DNA contamination. Total RNA was quantitated by measuring the absorbance ratios at the optical density of 260 and 280 nm. cDNA was synthesized with Superscript III (Invitrogen, Mt. Waverley, Australia) from 1 μg of total RNA, and PCR was performed to determine the expression of genes using primers (Table 3) for the chondrogenic differentiation (type II collagen [COL2]) and osteogenic differentiation (osteopontin [OPN] and osteocalcin [OCN]). Glyceraldehyde phosphate dehydrogenase was used as a house keeping gene for internal control.
Statistical analysis
Data were analyzed using one-way ANOVA for RNA expression and the comparison of calcium concentration. All analyses were performed with SPSS 13.0 (SPSS, Chicago, IL). Difference of p < 0.05 was considered statistically significant.
Results
Chondrogenic induction in 2D and 3D cultures
Alcian blue staining revealed the deposition of sulfated GAGs to assess chondrogenesis in the monolayer cultures (Fig. 1A–D) and in 3D fibrin gel cultures (Fig. 1 a–d). In chondrogenic medium (Fig. 1D, d) and the MCDM+TGF-β3 (Fig. 1C, c) positive stain was identified in both 2D and 3D cultures. The cells cultured in control medium (Fig. 1A, a) and MCDM alone (Fig. 1B, b) did not show any obvious staining. The monolayers induced in the MCDM+TGF-β3 (Fig. 1C) demonstrated more intense alcian blue staining than the cells cultured in chondrogenic medium (Fig. 1D). However, it was noted that BMSCs induced in the MCDM+TGF-β3, as well as chondrogenic medium in 2D cultures, exhibited spindle-shaped cells spreading over the culture plate, whereas the BMSCs cultured in 3D fibrin gel in the same media showed cell aggregation in a round and flattened morphology (Fig. 1C, D).

The differentiation potential of BMSCs toward chondrogenesis in 2D (
The histological sections of 3D fibrin cultures revealed the expression of GAGs in the cultures of MCDM+TGF-β3 (Fig. 2C) and chondrogenic medium (Fig. 2D), whereas no expression was detected in either the control medium (Fig. 2A) or MCDM alone (Fig. 2B). Immunohistochemical staining showed peri-cellular expression of aggrecan (Fig. 2E) and type II collagen (Fig. 2F) in the culture of MCDM with TGF-β3 similar to the chondrogenic culture. In cultures of control medium or MCDM alone, no staining was observed (result not shown).

The histological sections of the 3D fibrin cultures were stained for the deposition of glycosaminoglycans (GAGs), COL2, and aggrecan. The expression of GAGs was noted in the cultures of MCDM+TGF-β3 (
The chondrogenic differentiation was further validated by performing RT-PCR for the expression of COL2 mRNA in all the four treatment groups in the 2D and 3D cultures. COL2 expression was not detectable in the control media of 2D culture, whereas in the 3D control media or MCDM cultures some COL2 expression was detectable. The most prominent COL2 expression was detected in 2D and 3D cultures in MCDM+TGF-β3, as well as in chondrogenic medium (Fig. 3). Three-dimensional cultures showed positive expression for COL2 in all the treated groups, and compared with 2D cultures, COL2 mRNA expression was significantly upregulated (p = 0.012), especially in the MCDM+TGF-β3 and the chondrogenic medium group (Fig. 3).
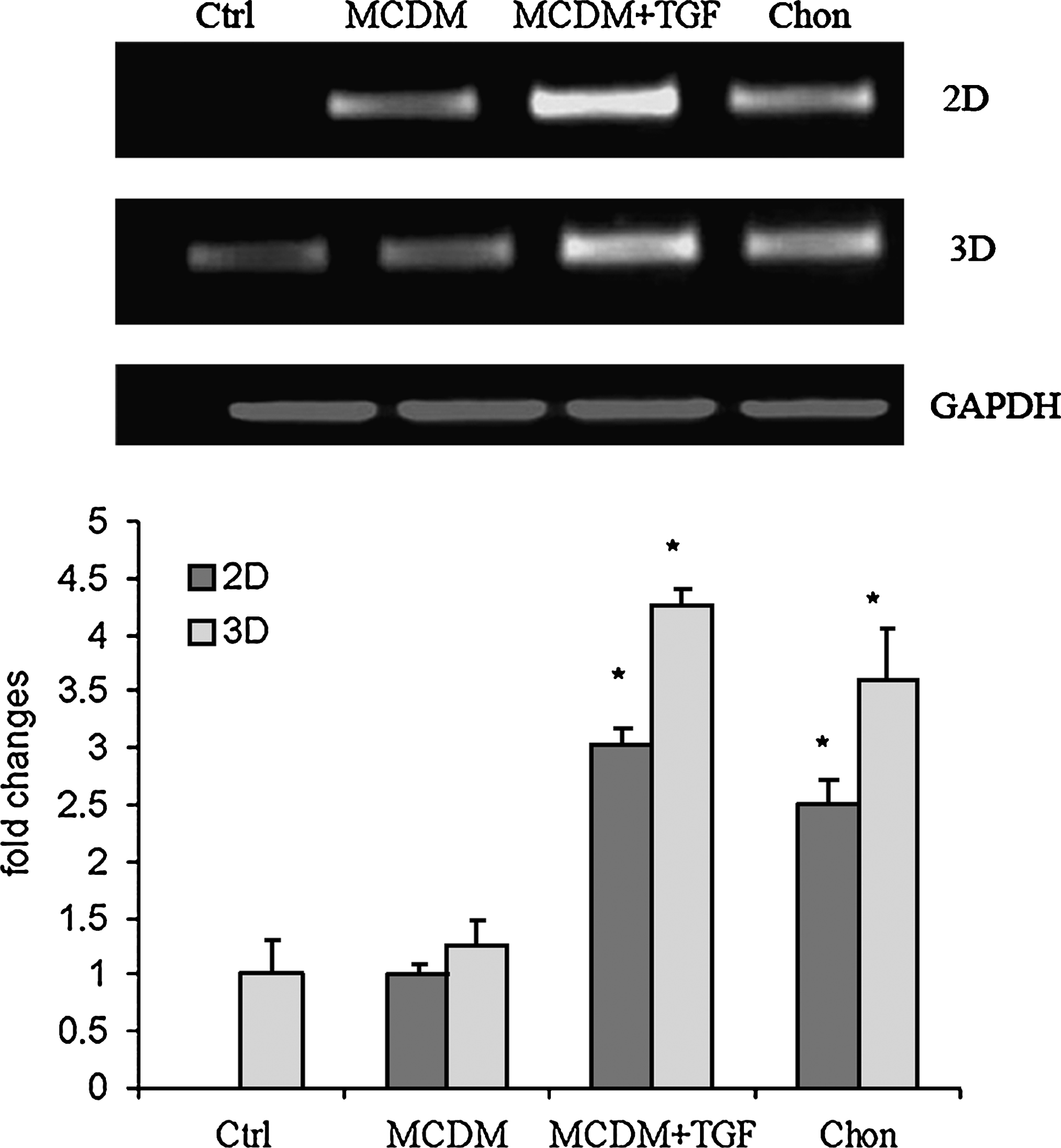
RT-PCR analysis for the expression of chondrogenic marker COL2 was performed on the 2D and 3D cultures. The cells were induced toward chondrogenesis by treating them with four types of media—control medium (Ctrl), minimum common osteochondrocytic differentiation medium (MCDM), MCDM supplemented with TGF-β3 (MCDM+TGFβ3), and chondrogenic medium (Chon)—for a period of 3 weeks. *, p < 0.05 in comparison with DCM group and 3D culture respectively.
Osteogenic induction in 2D and 3D cultures
Mineralization and nodule formation characteristic of osteogenesis was determined by von Kossa staining for the deposition of calcium phosphate. Cell cultures treated with osteogenic medium resulted in a greater extent of the nodule formation (Fig. 4D). In the other groups, no nodule formation was observed, although there was some mineral deposition in the monolayer cultures treated with MCDM + OP1 (Fig. 4C). The osteogenic differentiation results were further supported by calcium assay, which detected the calcium concentration in the monolayer cultures; calcium concentration in the matrix was found to be highly significant in the cells treated in osteogenic medium (p ≤ 0.0001) (Fig. 5), whereas little or no calcium was detected in the other treatment groups.

The differentiation potential of BMSCs toward osteogenesis in monolayers was assessed by inducing them in four types of media as shown in Table 2 for a period of 3 weeks. The monolayers were stained for the deposition of calcium phosphate crystals using the von Kossa staining. The figure shows relatively greater deposition of calcium phosphate crystals in the cells treated with osteogenic medium (
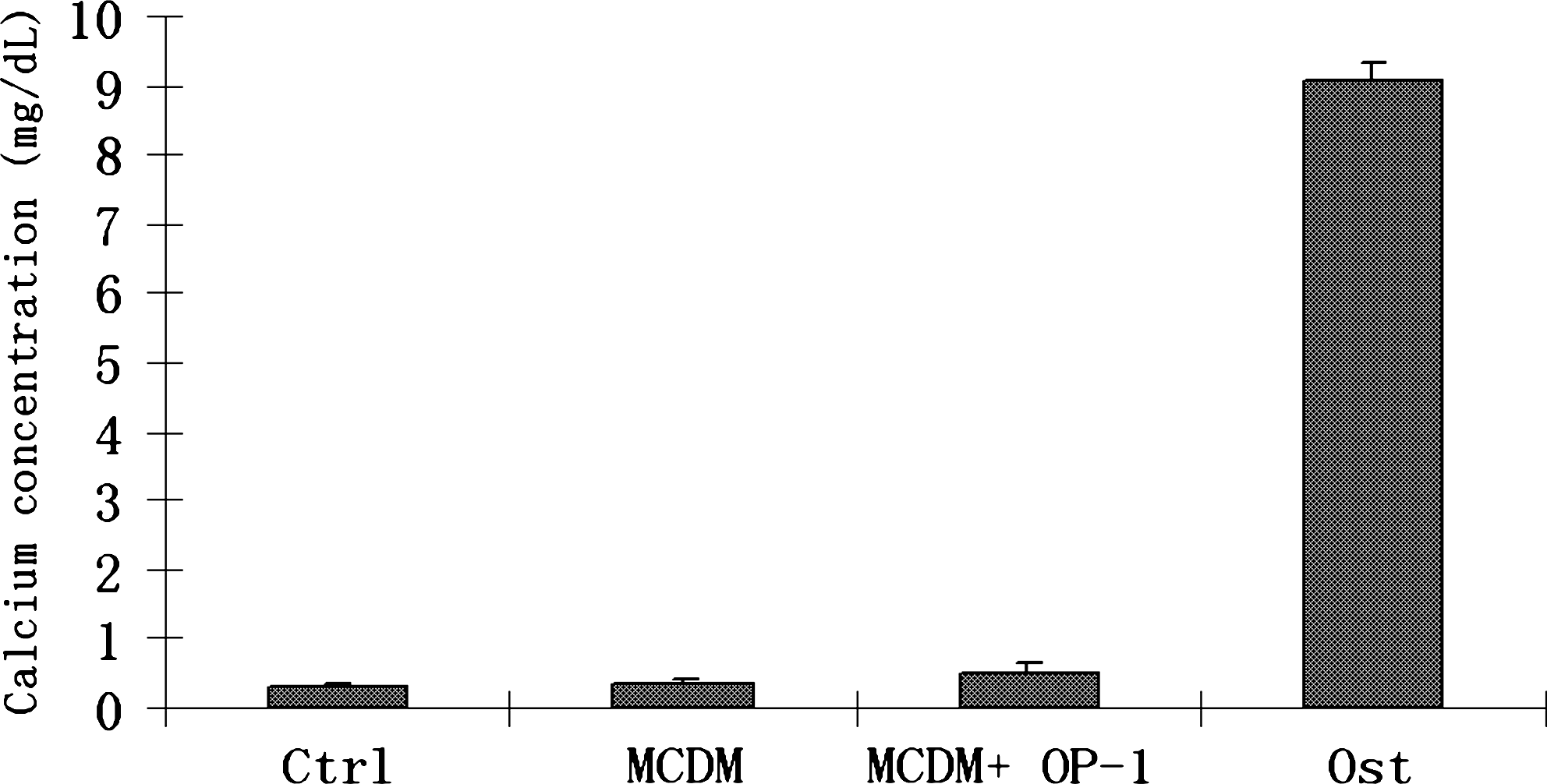
The differentiation results were further confirmed by calcium deposition assay. Calcium concentration in the matrix was found to be highly significant (p ≤ 0.0001) in the cells treated with the osteogenic medium and little detectable calcium levels in the other treatment groups.
In 3D cultures, BMSCs were seeded onto the HA/TCP scaffolds and cultured in different culture medium. SEM results revealed that BMSCs grew well in the HA/TCP scaffolds in Ctrl (Fig. 6A), MCDM (Fig. 6B), MCDM+OP-1 (Fig. 6C), and osteogenic media (Fig. 6D). The BMSCs spread out and formed web-like structures, and in the MCDM+OP-1 treatment group, many cells grew and bridged across the holes (Fig. 6C). RT-PCR showed that the mRNA expression of the osteogenic markers OPN and OCN was detectable in MCDM, MCDM+OP-1, and osteogenic medium, but not in the control medium. The relative density of the bands was measured using NIH Image V1.62, which revealed that the expression of OPN and OCN was more pronounced in the MCDM+OP-1 and osteogenic medium compared to the MCDM alone (Fig. 7).

BMSCs were seeded onto the HA/TCP scaffolds and cultured in different culture medium. SEM results revealed that BMSCs grew well in the HA/TCP scaffolds in Ctrl (

The mRNA expression of the osteogenic markers OPN and OCN in 2D (
Discussion
The various approaches proposed to date, for the fabrication of osteochondral composite constructs, are based on the progress made in material science, cell biology, and bioreactor technology. 1 Many strategies have been put forward for the engineering of osteochondral constructs, which have resulted from the use of one14,15 or more cell types cultured into either single-layer 16 or composite scaffolds17,18 covering a broad spectrum of compositions and biomechanical properties. 1
Selective differentiation of uncommitted cells into desirable lineages is achieved by manipulating the culture environment and incorporating selective growth factors within the scaffold. Differentiation of adult stem cells into particular cell lineages is an area of interest for researchers seeking to use stem cells for therapeutic applications.19,20 Cultured MSCs provide a useful model for evaluating the multiple factors responsible for the progression of cells from undifferentiated precursors to committed phenotypes. 2 In this study the effects of soluble mediators and medium supplements commonly used to induce chondrogenic and osteogenic differentiation were assessed in a minimal essential medium with supplemented growth factors for osteochondrocytic differentiation. A broader perspective of utilizing the MCDM is to fabricate and maintain the integrated osteochondral composite construct, where growth factor release can be controlled spatially and temporally from the biphasic scaffold itself or via spatial separation of double chamber bioreactors to prevent the diffusion of growth factors for osteogenic and chondrogenic differentiation. More importantly, the common ingredients in MCDM for osteogenesis and chondrogenesis will potentially provide an environment for the maintenance of tissue integrity of osteochondral construct, as well as the formation of the transitional zone of bone and cartilage in vitro.
In this study MCDM was formulated from the common soluble supplements used to induce chondrogenic and osteogenic differentiation. The medium was tested for its ability to differentiate 2D and 3D cultures of BMSCs into osteogenic and chondrogenic lineages based on the supplementation of the respective growth factors OP-1 and TGF-β3. Our findings showed that the MCDM supplemented with TGF-β3 or OP-1 was able to induce the proliferation and differentiation of BMSCs, respectively, into chondrocytes and osteocytes. Interestingly, the deposition of matrix proteins in the 2D culture induced in MCDM+TGF-β3 group was more pronounced than the monolayers induced in the conventional chondrogenic medium. This was supported by the results of RT-PCR analysis for the expression of the chondrocyte marker type II collagen, which was more intensely expressed in the cells induced with MCDM+TGF-β3 (Fig. 3). Consistent with 2D cultures, the mRNA expression of the 3D cultures for collagen II expression was found to be greater in the MCDM+TGF-β3 group than the mRNA expression of the 3D cultures induced in chondrogenic medium (Fig. 3). Although the amount of TGF-β3 in the MCDM+TGF-β3 and the chondrogenic medium was same, the difference in the expression of type II collagen may be attributed to the culture systems between 2D and 3D and the supplementation of 10% FBS instead of ITS premix to the common medium. Conventional medium used for chondrogenic differentiation consists of ITS premix supplement, which is a commercially available supplement for the culture of mammalian cells under serum-free conditions. Comparative studies involving chondrocyte growth and metabolism in ITS-supplemented serum-free medium to serum-supplemented medium revealed no significant difference in cell proliferation, or in protein and proteoglycan synthesis rate. 8
In contrast to the chondrogenesis results, the mineralization and nodule formation for osteogenesis in 2D culture induced in the conventional osteogenic medium was relatively greater when compared to the monolayers induced in the MCDM+OP-1 as revealed by von Kossa staining and RT-PCR results (Figs. 4 and 7). This may be attributed to the lack of β-GP in the OCDM+OP-1. β-GP is known to be the most important factor to accelerate in vitro calcification and induce extensive calcium deposition. 21 This was evident from the results of calcium assays that revealed that calcium concentration was significantly higher in the cultures treated with osteogenic medium when compared to the rest of the media types. A comparison between the mRNA expression of the bone-relevant markers, OPN and OCN, in 3D cultures, grown in the osteogenic medium and the OCDM+OP-1, showed insignificant differences. Chondrogenic and osteogenic 3D cultures grown in the control media and the MCDM alone groups faintly expressed the respective markers for chondrogenesis (COL2) and osteogenesis (OPN and OCN), despite the absence of the respective growth factors. This observation suggests a possible role of 3D cultures themselves for inducing lineage commitment of cell differentiation. Similar studies involving 2D and 3D culture models have demonstrated enhanced matrix deposition and bone/cartilage formation in the constructs generated by 3D culture of bone marrow cells when compared to scaffolds loaded with 2D expanded cells.22–24 It has been proposed that 3D cultures and the scaffold surface can also exert significant influence on the differentiation of the seeded stem cells. Mechanosensitivity and modification of the scaffold surface has recently been extended to the differentiation of MSCs.25,26 For example, when cultured on agarose gels with increasing cross linking densities, human MSCs differentiated into neuronal, muscle, or bone lineages according to the stiffness of the matrix that approximated that of brain, muscle, and bone tissue, respectively. 27
Previous attempts to engineer biphasic cell–scaffold constructs using hMSCs relied on seeding the 3D scaffolds with predifferentiated cells that were expanded in 2D monolayers28,29 as there is no common culture medium to support both chondrogenesis and osteogenesis. It has been suggested from earlier studies that 2D expansion may lead to changes in phenotype and loss of differentiation potential with serial passaging. Further, predifferentiated cells have decreased proliferation capacity and may not actively integrate well in construct.30,31 Future attempts to co-culture osteoblasts and chondrocytes for fabricating an integrated construct have been proposed employing innovative double-chamber bioreactors.32,33 Such technology potentially circumvents the need for predifferentiation of cells. Double-chamber bioreactors have been designed to support co-culture of different kinds of cells as they have two independent medium–circulation systems. 32 Hence, it can be used for generating composite osteochondral graft by applying the concept of MCDM. The essential regulatory molecules (TGFβ-3 and OP-1) can be spatially separated by the two independently circulation system, and MCDM will provide a common transitional environment in initiating events between differentiating chondrocytes and osteocytes for interface regeneration. Indeed, chondrocytes and osteoblasts have been reported to modulate each other in a co-culture system under physiological conditions.34,35
In conclusion, this study has demonstrated that culture-expanded BMSCs could be differentiated in vitro into chondrogenic and osteogenic lineages using an MCDM combined with growth factors, TGF-β3, and OP-1, respectively. The results suggest a potential for future fabrication of osteochondral composite graft in MCDM with a biphasic scaffold with spatially loaded growth factors and/or double-chamber culture system to control BMSC differentiation.
Footnotes
Acknowledgments
We would like to thank Ms. Wei Shi for her technical help in histology and Mr. Thor Friis for his proof reading of the whole manuscript. This research was supported by the Seeding Grant (2007) from the Institute of Health and Biomedical Innovation, Queensland University of Technology, and a discovery grant from the Australia Research Council (DP0881045).
Disclosure Statement
To the best of our knowledge, there are no competing financial interests in this study.