
Abstract
Ligament tissue engineering based on cell-seeded biomechanically functional constructs is a commonly studied strategy toward native anterior cruciate ligament replacement. Little is known about the survival and differentiation of the seeded cells after the transplantation. We applied retroviral genetic marking to trace implanted cells and studied their differentiation by species-specific immunolabeling of the extracellular matrix produced. Goat bone marrow stromal cells were transduced with a MoMuLV-based vector encoding the ΔLNGFR gene. Transduced cells were seeded onto poly(lactic-co-glycolic acid) (PLGA) fibers and implanted subcutaneously into nude mice and left for various periods up to 6 weeks. Immunohistochemistry for LNGFR expression showed survival of the seeded cells after transplantation for up to 6 weeks. Immunohistochemistry for collagen type I and III showed the production of fibrous tissue inside the scaffolds. Moreover, using a goat-specific anti-collagen type III, donor-derived matrix could be demonstrated. We conclude that bone marrow stromal cells survived in vivo and at least partially differentiated after implantation.
Introduction
Although many in vitro and in vivo studies have focused on tissue engineering of ligaments, little is known about the fate of the seeded cells after the transplantation. By use of physical separation from donor and host tissue and selective destruction of native cells, it was shown that fibroblasts in a reconstructed patellar tendon autograft for ACL reconstruction in a rabbit model died after reconstruction in the knee joint and the graft was rapidly repopulated by host cells 4 weeks after transplantation.17,18 Rabbit skin and ACL fibroblasts labeled with PKH26-GL and seeded on collagen scaffolds were found to be present in the intraarticular environment after 6 weeks. 19 However, as is a common problem with fluorescent membrane labels, label transfer and signal dilution hamper proper data interpretation.
A means to unequivocally study the presence of donor cells in an implanted construct is retroviral transduction with nonnative marker genes. LacZ (β-galactosidase) labeling has been applied to trace allografted and autografted cells in two rabbit models of ligament repair and autografting,20,21 but till now, no tissue-engineered ligament constructs have been evaluated for the presence of genetically labeled donor cells after implantation.
Another issue in addition to the actual presence of viable donor cells in vivo is their functionality. It is not known whether implanted cells actively take part in tissue formation, let alone whether successful tissue engineering actually requires the implanted cells to produce the desired matrix in vivo. To demonstrate that transplanted cells do not only survive but also produce extracellular matrix, the origin of the deposited tissue should be identified. One option to address this problem is to use species-specific recognition of proteins produced by the donor cells.
The objective of current study was to trace BMSC cells seeded onto braided PLGA scaffolds and to demonstrate their viability and functionality in terms of matrix production in vivo. Therefore, as a first step to demonstrate the applicability of this method, goat BMSCs were transduced with the ΔLNGFR gene, seeded on braided PLGA scaffolds, and implanted in an ectopic nude mice model. The origin of the produced matrix inside the scaffolds was qualitatively studied by species-specific immunostaining of collagen.
Materials and Methods
Cell culture
Goat BMSCs from bone marrow aspirates of a female Dutch milk goat (weight 60 kg, 19–24 months old) were harvested and expanded as described earlier. 22 The cells were cryopreserved at passage 1. Within 6 months, the goat BMSCs (passage 2) were thawed and replated in standard culture medium consisting of αMEM supplemented with 30% fetal bovine serum (FBS; Gibco, Paisley, Scotland, lot# 3030960S), L-glutamine (2 mM; Invitrogen, San Diego, CA), ascorbic acid (0.2 mM; Sigma, St Louis, MO), and penicillin/streptomycin (each 100 U/mL; Gibco). When confluent, the cells were detached and replated at 5000 cells/cm2 in culture medium containing 15% FBS.
The Phoenix-Ampho retrovirus packaging cell line was cultured in Dulbecco's modified Eagle's medium (Gibco) supplemented with 10% FBS, penicillin (100 U/mL), streptomycin (100 μg/mL), and 2 mM L-glutamine. Cultures were passaged twice a week and selected for gag, pol, env expression every 8 weeks using hygromycin B (300 μg/mL; Roche diagnostics, Mannheim, Germany) and diphtheria toxin A (1 μg/mL; Sigma-Aldrich, Zwijndrecht, The Netherlands). The construction of the retroviral vector pLZRS-TK has been described previously. 23 Twenty micrograms of DNA of a retroviral plasmid construct was transfected into 70% confluent Phoenix-Ampho packaging cells by calcium phosphate precipitation. 24 Twenty-four hours after transfection, medium was replaced with fresh culture medium. The following day, retroviral supernatant was collected, filtered through a 0.45 μm filter, and stored at −80°C. For additional harvest of retroviral supernatant, transfected Phoenix-Ampho cells were cultured for 3 days in the presence of puromycin (1 μg/mL; Sigma) followed by 2 days in culture medium without puromycin.
Retroviral transduction of goat BMSCs
Transduction was performed by replacing the standard cultured medium of the BMSCs (P3) by a threefold dilution of the retroviral supernatant in culture medium supplemented with 6 μg/mL polybrene (Sigma). Culture medium supplemented with polybrene alone was used for mock-transduction (control). The BMSCs were cultured for another 24 h, after which the medium was renewed. Two days later the cells were harvested and analyzed.
Transduction efficiency was determined by flow cytometry analysis. The BMSCs were labeled with primary mouse anti-LNGFR monoclonal antibody (20.4 culture supernatant, 1:20) for 20 min at 4°C, washed, and then incubated with goat anti-mouse phycoerythrine-conjugated IgG1 (Southern Biotechnologies, Birmingham, AL). Cells were washed in PBS–1% FBS and resuspended in standard culture medium, immediately before analysis of 10,000 events on a flow cytometer (FACS Calibur; Becton and Dickinson, San Jose, CA).23,25 In vitro culture was conducted to investigate long-term label expression. Two separately produced batches of transduced cells were subcultured further in 25 cm2 flasks for 6 weeks and analyzed by FACS at each passage. The remainder of the labeled BMSCs were cultured until confluence and subsequently seeded.
Transduction efficiency and long-term stability of cells
In a separate study in vitro culture was conducted to investigate long-term label expression. 25 Two batches of transduced cells were cultured in 25 cm2 flasks for 6 weeks and analyzed by FACS at each passage. To assess proliferative capacity, transduced and mock-transduced cells were cocultured in a 50%/50% mixture in a separate study. 25
Scaffolds
The degradable scaffolds were made of Panacryl (size 2/0; Ethicon, Somerville, NJ), which is composed of poly(L-lactide/glycolide) 11 μm bundles, containing approximately 40 monofilaments. 16 Nine bundles were organized in parallel, and both ends of the structure were sutured together (1 cm in length). The scaffolds were sterilized by repeated cycles of incubation in ethanol 70%, and after air-drying washed with sterile PBS. Before seeding, the scaffolds were preincubated overnight in standard culture medium.
Cell seeding
After trypsinization, both transduced and control BMSCs (P4) were resuspended at 2.5 × 106 cells per mL in standard culture medium. The scaffolds were seeded with 250,000 cells/scaffold and allowed to attach for 2 h, after which the scaffolds were submerged in culture medium consisting of αMEM supplemented with 15% FBS, L-glutamine (2 mM), ascorbic acid (0.2 mM), and penicillin/streptomycin (each 100 U/mL). All scaffolds were cultured for 7 days to allow for proliferation of seeded cells in 25-well bacterial culture plates (Sterilin, Stone, UK) before implantation.
Surgical procedure
All animal experiments were conducted with approval of the ethics committee of the University of Utrecht. Ten female NMRI nude mice (age, 4–6 weeks; weight, 20–25 g; Harlan, Horst, The Netherlands) were anaesthetized with an intraperitoneal ketamine (100 μg/mL), xylazine (20 μg/mL), and atropine (0.5 mg/mL) solution. After disinfection of the back with 70% alcohol, separate subcutaneous pockets were created for implantation of the scaffolds on the back of the mice. Implantations included two mice per evaluation period and two scaffolds with transduced cells and one scaffold with mock-transduced cells in each mouse (in addition to three tissue-engineered bone constructs that were implanted in the course of a separate study). The mice were sacrificed after time periods of 2, 4, and 10 days, and after 4 and 6 weeks. The implants were retrieved and processed for histological evaluation.
Histology
Samples were fixed in 10% buffered formalin. Dehydration was performed through a graded alcohol series before paraffin embedding. Four-micrometer-thick longitudinal sections of the scaffolds were deparaffinized, rehydrated, stained with hematoxylin and eosin, and analyzed with light microscopy (E600 Nikon Eclipse, Nikon, Japan). In addition, parts of the sections were stained for 60 min in Picrosirius Red (Direct Red 80; Aldrich, Milwaukee, WI). The presence of a collagenous matrix was viewed using polarization microscopy (E600 Nikon Eclipse).26–28 Collagen type I appears as thick, bright (strongly birefringent) yellow or red filamentous structures shining against a dark background, whereas collagen type III highlights as thin filamentous structures with a weak birefringence of a greenish color. 28
Immunohistochemistry
Paraffin-embedded scaffolds were deparaffinized and rehydrated, and endogenous peroxidase activity was blocked with 1.5% H2O2 in phosphate-citrate buffer for all samples. The samples were stained for LNGFR-positive cells and the presence of collagen type I and III to asses survival and functionality of the transplanted cells.
For the LNGFR immunolocalization, the Dako Ark™ kit was applied according to the manufacturers' recommendations (Dako Corporation, Carpinteria, CA) to prevent nonspecific staining due to reactivity of the secondary anti-mouse IgG antibodies with the surrounding murine tissue. For the primary antibody a mouse anti-LNGFR (2.56 μg/mL; Dako Corporation) was used.
For collagen immunodetection the sections were first treated with pepsin for 15 min at 37°C. Subsequently, the samples were incubated with rabbit anti-human collagen type I (26.7 μg/mL) or collagen type III (1:200; both Biogenesis, Dusseldorf, Germany) for 1 h at room temperature (RT). As secondary antibody, ready-to-use PowerVision (poly-HRP-anti-rabbit IgG; Immunologic, Duiven, The Netherlands) was applied according to the manufacturers' recommendations. Diaminobenzidine (Liquid DAB+ Substrate; Dako Corporation) was used to obtain a signal on the sections. To verify whether the primary antibodies for collagen type I and III could be used for species-specific immunolabeling, control sections containing goat muscle and skin and mouse skin and tendon tissue were labeled with each antibody.
An additional type III immunolocalization with an antibody known to detect both goat and mouse tissue was performed to demonstrate the presence of collagen type III in mouse tissue. Sections of mouse tendon were blocked with 1.5% H2O2 in phosphate-citrate buffer and subsequently treated with pepsin for 15 min at 37°C. Subsequently, the samples were incubated with rabbit anti-rat collagen type III (1:80; Division of Morwell Diagnostics, Zürich, Switzerland) for 1 h at RT. As secondary antibody, a biotinylated goat anti-rabbit (3 μg/mL; Vector laboratories) was applied for 1 h at RT. Subsequently, streptavidin horse radish peroxidase (SA-HRP; Beckman Coulter, Fullerton, CA) was applied for 1 h at RT. Finally, diaminobenzidine was used to obtain a signal in matrix.
All samples were counterstained with hematoxylin and analyzed by light microscopy (E600 Nikon Eclipse).
Results
Transduction efficiency and stability of labeled cells
The genetic labeling procedure of goat BMSCs resulted in a 40–60% transduction efficiency, measured 3 days after transduction. In vitro long-term culture showed a slight decrease of the percentage of labeled cells in the first week, but it stabilized at 30–40% positive cells for up to 6 weeks. In the coculture experiment the actual percentage of labeled cells showed to be initially lower during the first week, indicating a relative decreased proliferation compared to the unlabeled cell fraction. However, during the subsequent weeks, the percentage transduced cells in the coculture reached the expected percentage, indicating a minimal influence on cell proliferation and viability at the long term. 25 Viability immediately before seeding was in the range for normal BMSC P3 cells. Bone formation of the transduced BMSCs was described in a separate study, 25 showing their multipotency.
Histology
All mice survived the implantation periods and all samples were retrieved without signs of infection. As demonstrated by light microscopy of the paraffin-embedded scaffolds, cells and extracellular matrix were present throughout the whole scaffold (Fig. 1). At day 10 the cells and tissue in and around the fibers of the scaffold showed alignment along the fibers. At this time also small blood vessels were seen in the center of the scaffold. With time, the tissue inside the scaffold appeared much denser as demonstrated by the hematoxylin and eosin staining. Although no quantitative analysis was done, the cell density in this formed tissue appeared comparable to the earlier time points. Picrosirius Red staining likewise demonstrated a comparable increase in collagen matrix inside the scaffolds in time.
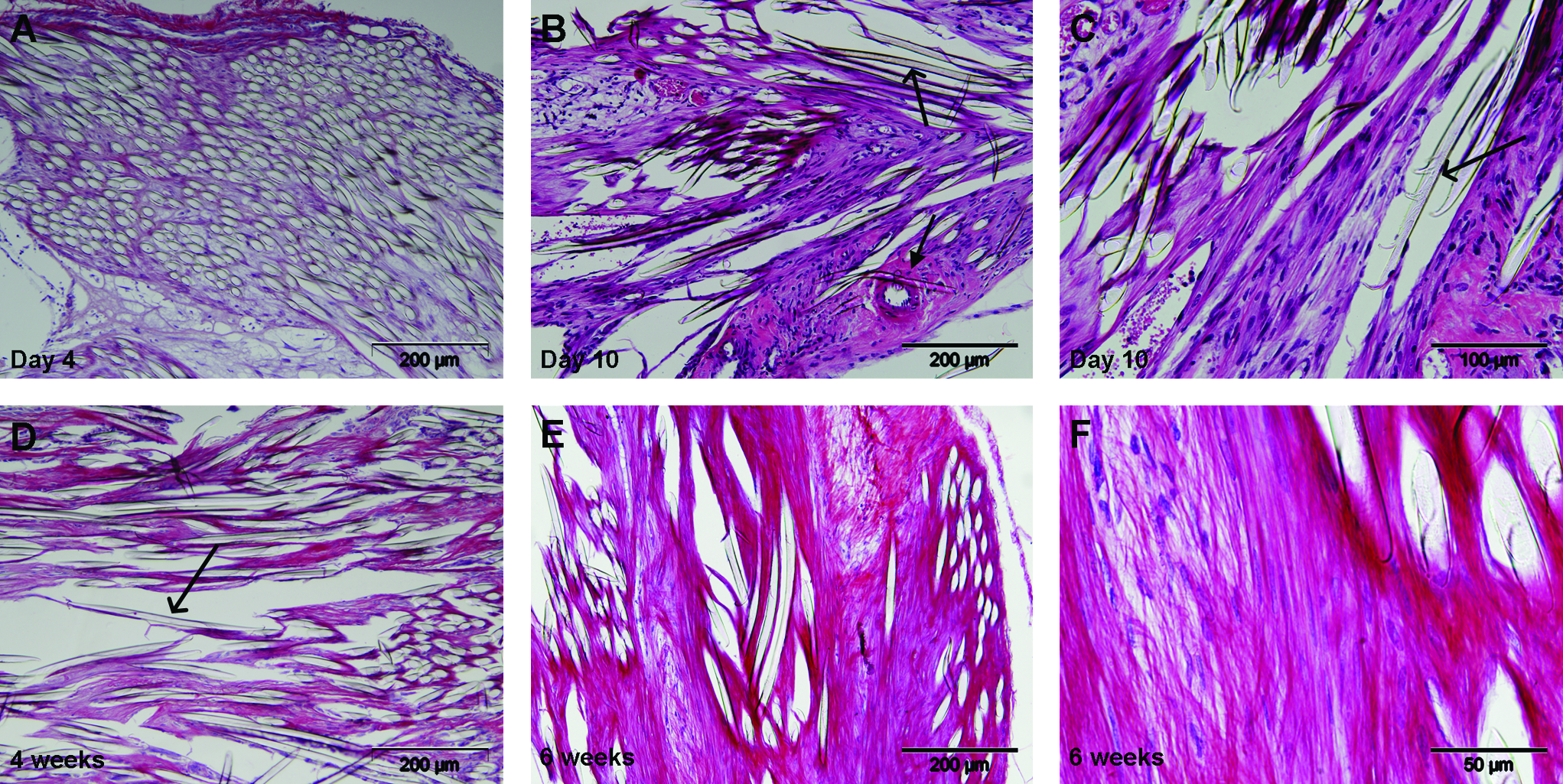
Histology of scaffolds stained with hematoxylin and eosin (
Polarization analysis indicated that both collagen type I and III could be seen inside the scaffolds, with collagen type III also seen adjacent to the scaffolds (Fig. 2).

Polarization microscopy of scaffolds cultured for 10 days (
Immunohistochemistry for detection of LNGFR expression
Labeled cells could be traced unequivocally from day 2 up to 6 weeks after implantation. The labeled cells were spread homogenously throughout the scaffold, without a specific preference for localization (Fig. 3). Due to the small sample size, no statements can be made about differences in cell density in time. Control scaffolds with mock-transduced cells and sections incubated with secondary antibody only were always negative (data not shown).
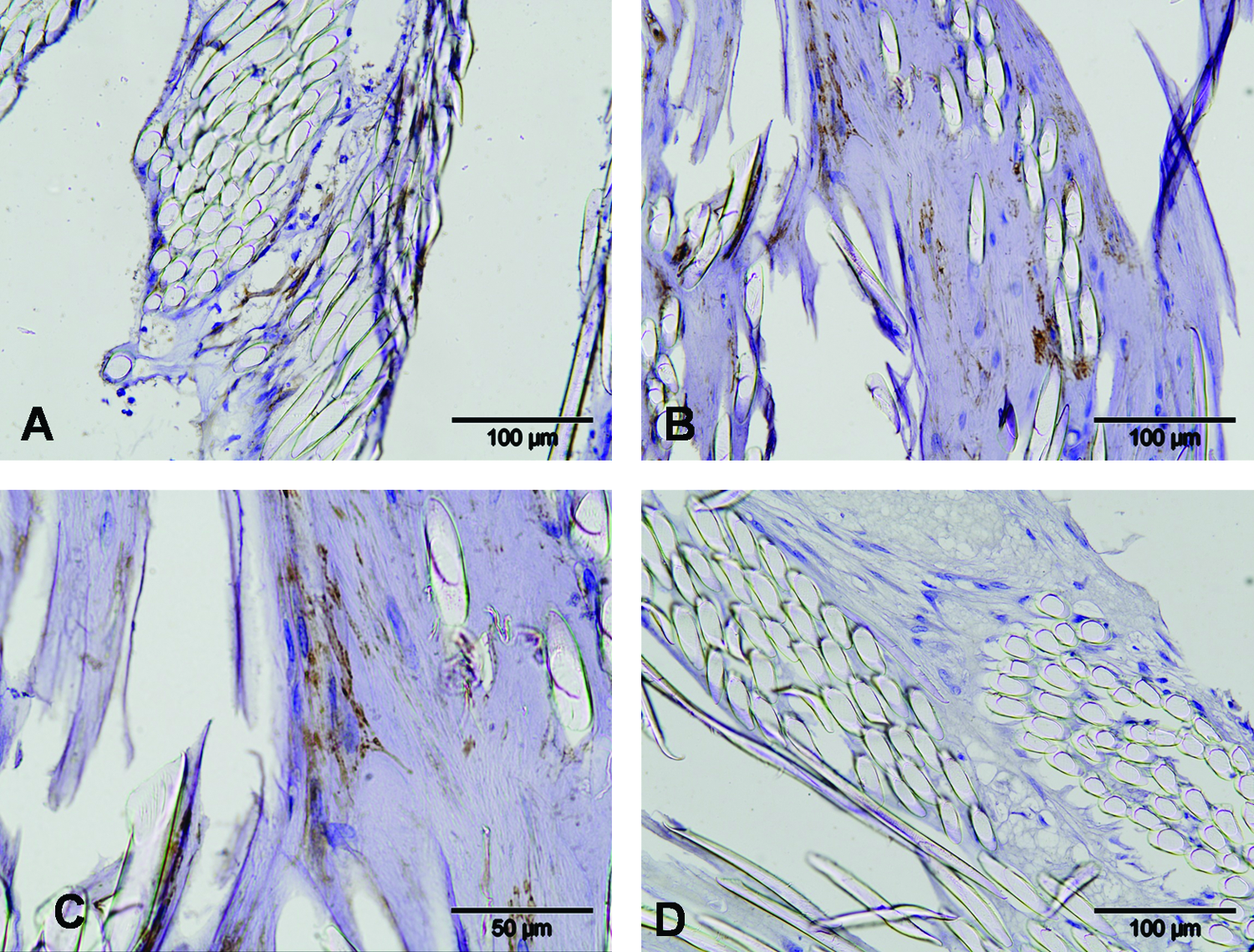
Immunohisto-chemistry for LNGFR-labeled cells at day 2 (
Collagen I labeling
Both mouse tendon and skin and goat skin tissue showed immunoreactivity for collagen type I. Inside the scaffolds there was a very dark positive staining that increased from day 10 to day 42. Around the scaffolds the tissue stained lightly (Fig. 4).
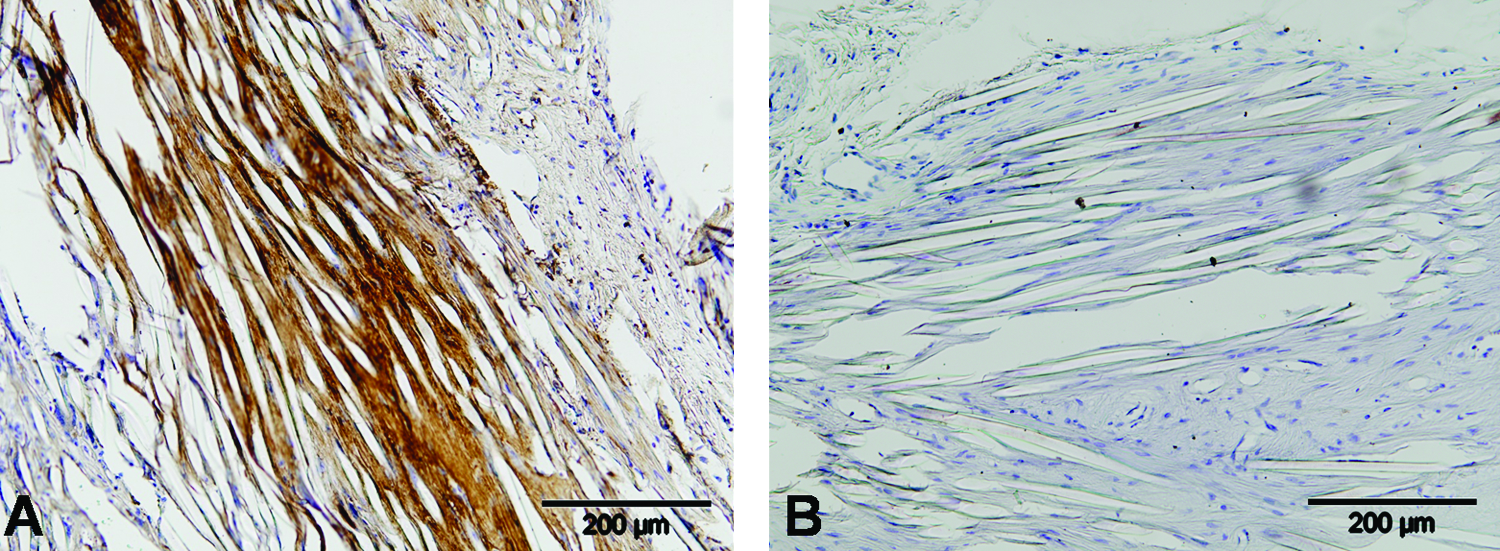
Immunohistochemistry for collagen type I inside the scaffolds at 6 weeks (
Collagen III labeling
Immunohistochemistry for collagen type III using the rabbit anti-human antibody demonstrated a goat tissue-specific staining in goat muscle and skin sections (Fig. 5A), with mouse skin and tendon tissue sections consistently negative (Fig. 5B). Using the rabbit anti-rat antibody, collagen type III could be demonstrated in both the above-mentioned mouse tissues, showing collagen type III is indeed present in the mouse skin and tendon (Fig. 5C). Staining with the goat-specific rabbit anti-human antibody collagen type III could be specifically identified inside the scaffold (Fig. 5D–G). Collagen type III could be identified from day 10 up to 6 weeks after implantation. The control tissue (PBS, mouse tissue) was always negative.

Immunohistochemistry for collagen type III for goat skin (
Discussion
In the current study we demonstrate the capacity of BMSCs to survive in vivo in a tissue-engineered construct for the replacement of ligamentous structures. Moreover, to our knowledge, this is the first study showing that this contribution represents more than just stimulating invading host cells by the release of factors, known to be produced by BMSCs, but that the cells actually also produce matrix. After seeding on PLGA scaffolds, goat BMSCs survived for up to 6 weeks when transplanted subcutaneously in nude mice. At least part of the extracellular matrix was indeed produced by the transplanted goat BMSCs as shown by species-specific labeling of type III collagen, thus indicating some form of differentiation. Conventional and Picrosirius Red histological staining suggested an increase in tissue density within the scaffold in time.
The first aim of current study was to follow the fate of seeded ΔLNGFR-transduced goat BMSCs implanted in an ectopic nude mice model. LNGFR has been applied successfully for tracing many cell types, by both flow cytometry and immunohistochemistry.23,25,29–31 Transplantation of genetically modified autologous cells may be more challenging in more clinically relevant models of immunocompetent, larger animal models such as in a goat. However, several in vivo studies have already been conducted introducing foreign genes, such as luciferase and GFP in vivo without any serious immunological reaction being reported.32,33 Previous research aiming at tracing transplanted and transduced cells showed variable survival periods for such cells. The density of GFP-transduced allogeneic chondrocytes embedded in alginate and transplanted into osteochondral defects in rabbits decreased to 15% within 4 weeks. 33 EGFP-labeled allogeneic chondrocytes transplanted into a similar rabbit model could not be detected after 6 weeks anymore. 34 It was unclear whether a decrease in density of labeled cells was due to cell death, through rejection by the host cells, or due to loss of expression. Also, the process of transduction itself could have had an effect on the behavior of transplanted cells. Both studies showed a down-regulation of cartilage gene expression of the transfected cells in vitro, but neither study investigated to what extent this down-regulation changed the actual functionality of the cells after transplantation. In the current study, the effect of transduction on in vitro cell viability and proliferative capacity in current study was minimal, as shown previously with coculture experiments on proliferation of labeled and unlabeled cells. 25 Still it cannot be excluded that transduction had some effects on in vivo differentiation. The method used to isolate the BMSCs has been shown to generate BMSCs capable of differentiation into the chondrogenic, osteogenic, and adipogenic cell line in vitro.22,35,36 Also, the multipotency of the same transduced population used in this study has been shown by their induction of bone formation in vivo. 25
Despite unequivocal demonstration of donor cells in a given in vivo situation, nothing can be concluded concerning the actual functionality of the implanted cells. The presence of labeled cells inside osteocyte lacunae may be regarded as proof of the bone-forming capacity of these cells25,37; however, in other tissues such as ligament, it is more difficult to pinpoint the origin of the produced matrix. The above-mentioned study on the fate of transgenic transplanted chondrocytes into articular defects did not show integration of these cells into the repair tissue, and therefore the authors concluded that the cells did not form the repair tissue. 33 In the current study, in addition to showing the presence of the transplanted cells, the second aim was to prove their differentiation by species-specific immunostaining of collagen type III produced after xenotransplantation. This is an important step toward demonstrating the functionality and thus the need for implanted cells in orthotopic locations. As to the application of the biomaterials used, PLGA-based constructs have been evaluated extensively before. They were found to allow for efficient attachment and proliferation of and matrix production by seeded goat BMSCs in vitro.16,38 Maximum loads for braided constructs were shown to be around 60 (PLLA + PLGA) 39 to 100 N (PLGA or PLLA), 40 with ultimate tensile strength decreasing to 50% after 16 weeks in vitro. However, the maximal tensile load of PLA/PLGA constructs after 20 weeks in vivo had decreased to about 10% of native ligaments, indicating biomechanical functionality at later time-points is only partial and stressing the need for efficient stimulation of matrix deposition after implantation. 41
In addition to collagen type III, production of collagen type I inside the scaffolds was clearly demonstrated, although we were not able to show that this was formed by the transplanted cells. Collagen type I and III are found in bone and in soft connective tissues like skin and are major extracellular matrix components of tendon and ligaments. 42 Wound healing in general is associated with relatively high contents of type III collagen, which has also been shown for regenerating tendon tissue, 43 and therefore the tissue formed by the labeled cells may partly reflect scar tissue. As the major goal of this study was to investigate the in vivo behavior of BMSCs implanted on biomaterial scaffolds for ligament reconstruction, rather than the production of ligament tissue, no efforts were undertaken to further specify the nature of the tissue. Putative markers further identifying the tissue as ligament-like would have been fibronectin, tenascin-C, scleraxis, and tenomodulin, but in the end none of these are truly specific, as they are also found in other tissues and cells such as cartilage, dermis, testis, and sclerae.44–50 For future in vivo studies on the functionality of transplanted cells, a combination of the markers mentioned above, as well as the ratio of collagen type I versus III, may prove some insight into the nature of the tissue formed. Also, the unique orientation of the collagen fibers in ligaments and tendons resulting in its high mechanical properties are important determinants to investigate. However, as has been shown extensively that under unstrained conditions extracellular matrix is deposited without any particular orientation,4,13 the latter parameter is only meaningful when analyzed on constructs that actually have been biomechanically loaded in vivo, in contrast to the current study.
Still the lack of a subset of ligament markers in any given construct at an ectopic location may not preclude functionality at orthotopic locations. Exposure to the proper biomechanical cues in vivo may be sufficient to direct differentiation into the tenogenic/ligament lineage, as has been shown for fibroblasts differentiating into functional tendon tissue in vivo.51,52
In conclusion, we demonstrate that BMSCs survive in vivo transplantation on PLGA scaffolds and are able to produce collagenous matrix inside these scaffolds designed for tissue engineering of the ACL. Future studies are required to show whether this also occurs in seeded constructs implanted at orthotopic locations and will need to address the final biomechanical functionality of these constructs.
Footnotes
Acknowledgments
The authors acknowledge Mirjam Stijns for her help with the cell labeling and The Netherlands Technology Foundation (STW; grant UGN.4966) for financial support. Dr. Saris is supported by the Netherlands Organisation for Health Research and Development (NWO) and Dr. Creemers by the Dutch Arthritis Association.
Disclosure Statement
No competing financial interests exist.