
Abstract
Polyelectrolyte multilayer coatings have emerged as substrates to control cellular behavior, but interactions with human multipotent mesenchymal stromal cells (MSCs) have not been studied. We looked at layer-by-layer coatings of cationic poly-L-lysine (PLL) and anionic hyaluronic acid (HA) as substrates for MSCs of placenta and adipose tissue. This system allows for modulation of thickness (number of deposition cycles), stiffness (chemical cross-linking of bulk layer), and adhesiveness (fibronectin (FN) interface). Native, as-built PLL/HA multilayer coatings were poorly adhesive for MSCs despite spectroscopy-confirmed high surface density of pre-adsorbed FN. Stratification of cross-linked PLL/HA multilayers of different stiffnesses revealed that multilayers modified with a high cross-linking regimen became efficient substrates for MSC adhesion and proliferation. MSCs on cross-linked multilayers grew to confluence. Using comparative confocal microscopy analysis of PLL/HA multilayers with physically adsorbed versus chemically coupled FN, we demonstrated that cross-linking strongly influenced FN surface distribution, leading to denser presentation of adhesion sites for cells. The covalent affixation of FN promoted focal adhesion formation and was critical to maintaining densely grown MSC cultures over weeks for their differentiation. Multilayer-bound MSCs were capable of differentiating into osteocytes and chondrocytes upon culture with induction factors. Together, cross-linked, FN-terminated PLL/HA multilayers provide a versatile platform for studies of human MSCs for biotechnological or therapeutic applications.
Introduction
Several studies have recently investigated cell interactions with PEMs. The biophysically and biologically best-studied systems are arguably multilayer coatings of anionic HA and cationic PLL.3,10,11 Even micrometer-thick films of (PLL/HA) i have been achieved, where i is the number of layer pairs. 12 HA is a natural, linear polysaccharide with repeating disaccharide units of D-glucuronic acid and N-acetyl-D-glucosamine with β(1Π4) interglycosidic linkage and is found in the extracellular matrix of connective tissue, where it is present as a hydrated network with collagen fibers. Its average molar mass ranges from approximately 105 to 107 Da. HA provides excellent biocompatibility, and native and modified HA have been used for ophthalmic surgery, arthritis treatment, drug delivery, and tissue engineering. 13 The counter polycation PLL is widely used in cell culture to promote cell adhesion to solid substrates and can be easily conjugated with bioactive molecules. Mechanistic studies by Picart et al. indicated that PLL/HA multilayer buildup begins with isolated islands that grow by addition of new polyelectrolytes on their top and by mutual coalescence and that continuous films are obtained after the eighth layer pair deposition. 3 Native, as-built PEM coatings are soft, elastic substrates that did not support cell adhesion well. Increasing stiffness of PLL/HA multilayers was most influential for cell adhesion. 9 Using chemical cross-linking of PLL/HA multilayers by water-soluble carbodiimide in combination with N-hydroxysulfosuccinimide coatings were obtained with an effective Young's modulus that was more than 10 times as large as in a native, non-cross-linked multilayer substrate. 11 Tests of cross-linked PLL/HA multilayer substrates with primary chondrocytes and smooth muscle cells showed greater cell anchoring and spread on the cross-linked layers.9,14
Mesenchymal stromal cells (MSCs) represent a particularly interesting cell type for research and therapy because of their ability to differentiate into mesodermal lineage cells such as osteocytes, chondrocytes, cardiac muscle, and endothelial cells. 15 In addition, they secrete large amounts of pro-angiogenic and anti-apoptotic cytokines 16 and possess remarkable immunosuppressive properties. 17 MSCs have been derived from many different organs and tissues, but much of the literature is on MSCs from bone marrow or adipose tissue. 15 Dominici et al. 18 have recently developed a general, minimal working definition of MSC isolates that rests on three criteria: plastic adherence in in vitro culture; molecular surface profile (expression of CD105, CD73, and CD90 but lack of CD45, CD34, CD14 or CD11b, CD79alpha or CD19, and human leukocyte antigen DR); and in vitro differentiation into osteoblasts, chondroblasts, and adipocytes. These criteria apply to the human placenta–derived MSCs (Fig. 1) and human adipose tissue–derived MSCs of the present study. Interactions between MSCs and PEM substrates have not been studied before. Here we describe PLL/HA multilayer substrates that we engineered with physical and biological properties that permit adhesion, proliferation, and differentiation of human MSCs.
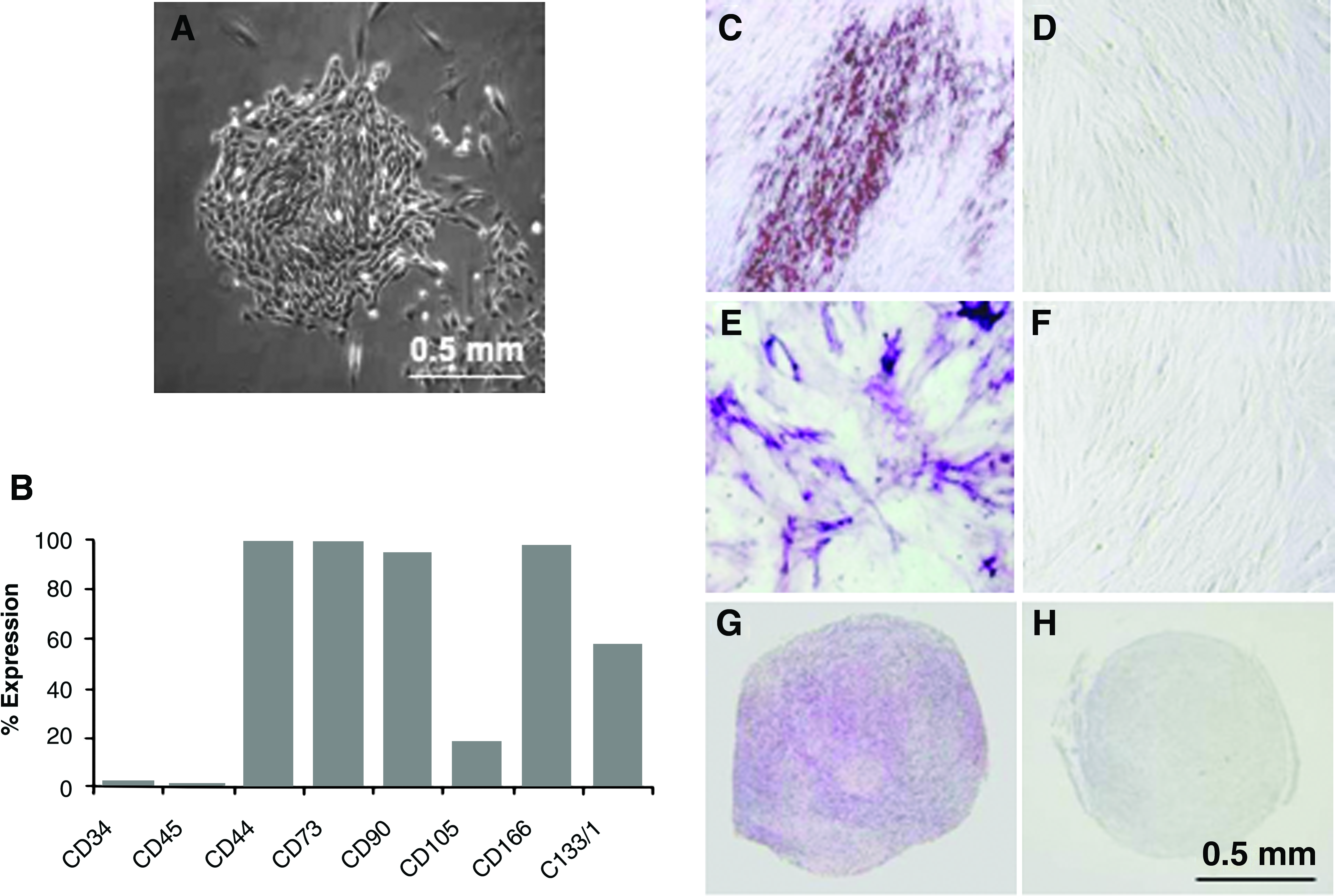
Characterization of placenta-derived mesenchymal stromal cells (PD-MSCs) from stroma of term human placenta.
18
(
Materials and Methods
Reagents
If not otherwise indicated, the reagents were purchased from Sigma-Aldrich Chemie GmbH (Buchs, Switzerland).
Cells
Placenta-derived (PD) MSCs were isolated according to a previously published protocol 19 from term human placentas that were collected after elective Caesarean section. All tissue samples were obtained after informed consent of the mothers. The ethical committee of the District of Zurich, which operates under federal legislation, approved the experiments. Human adipose tissue–derived (AT) MSCs were kindly provided by Dr. G. Wouters (Cryo-Save Labs, Mechelen, Belgium). The mouse myoblast cell line C2C12 was kindly provided by A. Franco-Obregon, Swiss Federal Institute of Technology (Zurich, Switzerland). All cell types were cultured in non-hematopoietic stem cell expansion medium (Miltenyi Biotec GmbH, Bergisch-Gladbach, Germany). Cultures were grown to near confluence in a humidified incubator at 37°C and 5% carbon dioxide. Cell passage was performed using trypsin–ethylenediaminetetraacetic acid (Gibco, Invitrogen AG, Basel, Switzerland).
Flow cytometry
Cells were collected using trypsinization. Reaction was performed on live cells, using 105 cells per antibody reaction. Cells were incubated for 45 min at 4°C. All primary antibodies were directly labelled as indicated with fluorescein thiocyanate (FITC), phycoerythrin (PE), or allophycocyanin (APC). The following antibodies were used: anti-CD44-FITC, anti-CD73-PE, anti-CD90-FITC, anti-CD105-FITC, anti-CD166-PE (BD Pharmingen, Erembodegem, Belgium); anti-CD133/1-APC, anti-CD34-PE, anti-CD45-FITC (Miltenyi Biotec). Cells were fixed in 4% buffered formalin and analyzed using a flow cytometer (FACSCalibur, Becton Dickinson, Franklin Lakes, NJ). A minimum of 104 gated events were acquired for each sample.
Substrate cleaning
Silicate glass coverslips (for cell culture experiments) or silicate waveguides slides (for optical measurements) was pretreated as previously described. 20
Polyelectrolyte and fibronectin solutions for multilayer buildup
Solutions of poly(L-lysine) hydrobromide (PLL, Mw 15-30 kDa) and hyaluronic acid sodium salt from bovine vitreous humor (HA, Mw 300 kDa) were prepared at 1 mg/mL in 10 mM 4-(2-hydroxyethyl)piperazine-1-ethanesulfonic acid, pH 7.4, 0.15 M sodium chloride (HEPES-2 buffer). 9 Human recombinant fibronectin (FN) (Biomedical Technologies Inc., Stoughton, MA) was used at 50 μg/mL in HEPES-2 buffer. All solutions were sterile filtered through 0.22-μm filters before use. Fluorescent labeling of FN was performed with Alexa Fluor 488 protein labeling kit (Invitrogen, Basel, Switzerland) following the manufacturer's protocol.
Preparation of native and cross-linked PLL/HA multilayer coatings
(PPL/HA) i multilayer coatings, where i is the number of layer pairs, were prepared on glass coverslips 15 mm in diameter that were placed on the bottom of 12-well tissue culture plates. Sequential layering of the polyelectrolytes and rinsing were done using manual pipetting. PLL was first adsorbed by 5 min immersion of the glass slide in PLL solution. Then the PLL solution was removed, collected, and rinsed with HEPES-2 buffer for 2 min. Then HA was adsorbed and subsequently rinsed following the same procedure. Adsorption of PLL and HA was performed alternately until the desired multilayer was built. In some experiments, an additional layer of FN was adsorbed for 45 minutes. The multilayers were further processed for cross-linking (see paragraph below) or subjected to three final rinsing steps (2 × 1 h, then 3 h) with 0.9% sodium chloride (NaCl) in deionized water.
Cross-linking of multilayers was performed with N-(3-dimethylaminopropyl)-N′-ethylcarbodiimide hydrochloride (EDC) in combination with N-hydroxysulfosuccinimide sodium salt (NHS), similar to the process described by Richert et al. 9 The coupling chemistry is based on the reaction between activated carboxylate groups of HA and the ammonium groups of PLL and results in the formation of amide bonds. EDC and NHS were prepared in 0.9% NaCl in deionized water and mixed together immediately before use. The various EDC concentrations tested were 0, 8, 40, 80, and 800 mM; the NHS concentration was 200 mM in all reactions. The pH of mixed cross-linker solution was 5.5. The multilayers were incubated in EDC/NHS for 12 h at 4°C. Then the EDC/NHS solution was removed, and the multilayers were rinsed three times with 0.9% NaCl in deionized water (2 × 1 h, 1 × 3 h) to hydrolyze unreacted cross-linkers.
Validation of polyelectrolyte multilayer buildup using optical waveguide light mode spectroscopy
Deposition of polycations, polyanions, and FN on waveguides was measured in situ using optical waveguide light mode spectroscopy (OWLS) as described previously.4,20 The instrument and the waveguides were obtained from MicroVacuum Ltd. (Budapest, Hungary). The waveguide was mounted into a flow-through cell with a volume of 15 μL and left in HEPES-2 buffer overnight. After stabilization of the buffer baseline, polyelectrolyte solution was injected and allowed to adsorb for 5 min for PLL and HA, with 2-min rinsing steps in between. Adsorption of PLL and HA layers was performed alternately until the desired number of layers was built. In some experiments, an outer layer of FN was injected and allowed to adsorb for 120 min.
Cell attachment experiments
Cell attachment experiments were performed on PEM-coated, 15-mm glass coverslips placed in 12-well plates. AT-MSCs, PD-MSCs, and myoblasts were seeded at various cell densities as indicated in the text of the manuscript and cultured in non-hematopoietic stem cell culture medium (Miltenyi Biotec). Control cultures were grown on tissue culture plastic. For analysis of cell spreading, cytoskeletons were stained with rhodamine-labelled phalloidin according to the manufacturer's instructions (Invitrogen). Cell nuclei were stained with 4′6-diamidino-2-phenylindole dihydrochloride (DAPI; Molecular Probes, Eugene, OR), and images were acquired using a DMIL fluorescence microscope (Leica, Heerbrugg, Switzerland) equipped with a DML digital camera (Leica) at 10 ×, 20 ×, or 40 × magnification. Cell densities were quantified colorimetrically using crystal violet staining. For that, slides were fixed in methanol for 20 min and then transferred into new 12-well plates and incubated with 0.5% crystal violet (AppliChem GmbH, Darmstadt, Germany) in 20% methanol for 1 h at room temperature and then rinsed with water and air-dried. Destaining was performed using overnight incubation in 2% sodium dodecyl sulphate. Optical density of the extracts was determined at 600 nm. In some experiments, cell counts were obtained from fluorescence micrographs of DAPI-stained cells using automated image analysis software ImageJ 1.34s (National Institutes of Health, Bethesda, MD).
Differentiation assays
Differentiation of PD-MSCs and AT-MSCs was induced by their culture in commercial differentiation media (AdipoDiff, ChondroDiff, and OsteoDiff induction media; Miltenyi Biotec). Control cultures were grown in non-hematopoietic stem cell expansion medium. After differentiation, cells were examined microscopically with a Zeiss Axiovert 200M (Carl Zeiss, Feldbach, Switzerland) equipped with a digital camera (AxioCam MRc, Carl Zeiss) and analyzed with AxioVs40 V 4.5.0.0 imaging software (Carl Zeiss).
For osteogenic differentiation, MSCs were maintained in OsteoDiff induction medium for 2 weeks, with fresh medium added every third day. Alkaline phosphatase (ALP) expression was demonstrated using cytochemical staining with 5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazolium (FAST BCIP/NBT) tablets (Sigma-Aldrich). For that, cells were fixed in pre-cooled methanol for 5 min, rinsed with deionized water, and then incubated with FAST BCIP/NBT substrate for 10 min at room temperature. Finally, stained cells were washed with water and examined microscopically.
For adipogenic differentiation, MSCs were cultured in AdipoDiff medium for 3 weeks, with fresh medium added twice weekly. Oil Red O was used to visualize fat droplets following the manufacturer's instructions. Briefly, cells were washed twice with PBS, fixed in pre-cooled methanol for 5 min, rinsed with deionized water, and then incubated with Oil Red O (Sigma-Aldrich) staining reagent for 20 min at room temperature. Finally, stained cells were washed with water and analyzed microscopically.
Chondrogenic differentiation was performed under micromass conditions in ChondroDiff medium according to the manufacturer's instructions. Briefly, 5 × 106 MSCs were pelleted using 5 min of centrifugation at 290 g in a cell culture centrifuge. The micromass was kept in ChondroDiff medium for 24 days, with fresh medium added every 48 h. The pellets were fixed in 10% formalin and then embedded in paraffin. Staining for proteoglycans was performed on deparaffinized 5-μm sections using 40 s of incubation with 0.3% Toluidine Blue O in water.
MSC differentiation on PEM substrates
PD-MSCs, passage 6, were plated onto FN-terminated, cross-linked (PLL/HA)9-PLL multilayers at 12.5 × 103 cells/cm2. The cells were grown to confluence before differentiation was initiated in 14 day culture in ChondroDiff or OsteoDiff induction medium. Then cells were fixed in 4% paraformaldehyde and stained for ALP expression to demonstrate osteogenic differentiation. Chondrogenic differentiation was demonstrated by staining with polyclonal goat anti-human collagen II antibodies (Santa Cruz, Ltd., Santa Cruz, CA) and secondary FITC-conjugated anti-goat antibodies (DAKO Cytomation, Glostrup, Denmark).
Confocal microscopy and image analysis
Confocal images were acquired using a Leica TCS-SP 2 confocal microscope. The data were further processed using Imaris v. 2.6 software (Bitplane Scientific Solutions AG, Zurich, Switzerland).
Immunocytochemistry
For immunofluorescent staining, PD-MSCs or C2C12 cells were plated at a density of 1.5 × 103 cells/cm2 on native or cross-linked FN-terminated (PLL/HA)9 layers and then cultured for 24 h. Cells were then permeabilized with 0.1% Triton X-100 and stained for vinculin using FITC-conjugated anti-vinculin antibodies (1:100; Sigma-Aldrich); cell nuclei were stained using DAPI.
Statistical analysis
Data are shown as means ± standard deviations. Two-tailed unpaired t-tests were performed using GraphPad Prism version 4.00 for Windows (GraphPad Software, San Diego, CA). Significance was set at p < 0.05.
Results
Characterization of MSCs from human term placenta
We performed our experiments with MSCs from human placental and adipose tissue because these sources are readily accessible for biological and biotechnological applications. Plating of collagenase/dispase digests from 25 g of placental tissue, typically on tissue culture plastic, followed by culture in non-hematopoietic stem cell culture medium initiated approximately 20 to 40 colonies of fibroblast-like cells after 1 to 2 weeks (Fig. 1A). The cells exhibited fast proliferation up to passage 15, which was as far as we tested. The phenotypic and differentiation characteristics of our PD-MSC preparations conformed to the criteria defining multipotent mesenchymal stromal cells. 18 Figure 1B–H shows the data for PD-MSCs, passage 6. The cells were positive for mesenchymal stromal markers CD44, CD73, CD90, CD105, CD166, and CD133, a marker for multipotentiality, and negative for hematopoietic markers CD34 and CD45. The placenta-derived MSCs displayed the potential to differentiate toward the adipogenic, osteogenic, and chondrogenic pathways, as demonstrated by the respective histochemical stains (Fig. 1C, E, G). Without the addition of differentiation factors, PD-MSCs did not differentiate (Fig. 1D, F, H). Human AT-MSCs, passage 2, demonstrated similar mesodermal differentiation capacity as PD-MSCs (data not shown).
Interaction between MSCs and native PEM substrates
To promote interactions with MSCs, the following parameters of PLL/HA multilayer buildup were controlled. (1) Thickness was varied according to the number of PLL/HA layer pairs. (2) Cell adhesiveness was approached through deposition of cell adhesion glycoprotein FN as the outermost layer. (3) Stiffness of the multilayer was increased by introducing covalent cross-links and amide bonds between HA and PLL. (4) FN was covalently affixed or physically adsorbed onto multilayer. Coatings were prepared with three-, six-, or nine-layer pairs of PLL/HA, followed by an end layer of positively charged PLL and the final (optional) adsorption of FN. The resulting constructs were designated (PLL/HA) i -PLL-FN, where i is the number of layer pairs. Figure 2A depicts the stepwise adsorption of PLL, HA, and FN as monitored in situ by OWLS as the example of a (PLL/HA)9 coating (see also Suppl. Fig. 2 for additional adsorption curves). OWLS showed successful adsorption of FN, which assumed surface densities between 350 and 700 ng/cm2 on three-, six, or nine-layer-pair constructs (Table 1). These adsorbed FN surface densities are biologically significant. Saturation levels of 350 to 400 ng/cm2 were previously reported to represent approximately the amount of FN necessary to produce a monolayer coating based on the dimensions of the molecule.21,22 Such densities are known to support strong cell adhesion on solid synthetic surfaces as well as on thin multilayer films. 23
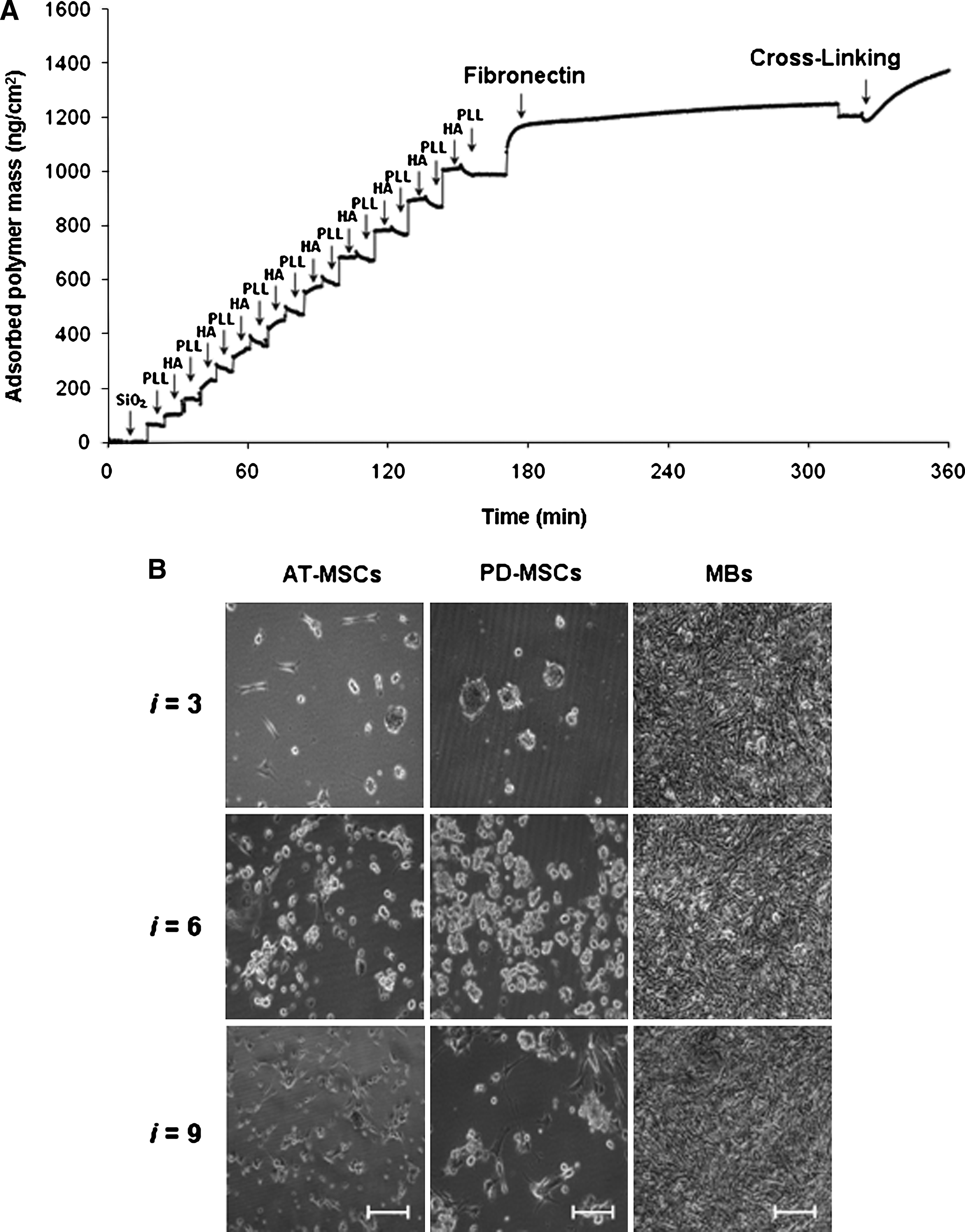
MSCs do not adhere to native cationic poly-L-lysine and anionic hyaluronic acid (PLL/HA) multilayer coatings. (
PLL-HA, poly-L-lysine-hyaluronic acid; FN, fibronectin; PD, placenta-derived; MSC, mesenchymal stromal cells; AT, adipose tissue-derived.
None of the FN-terminated PLL/HA multilayer substrates, when used as built, functioned as adhesive substrate for PD-MSCs or AT-MSCs (Fig. 2B). Both types of MSCs, when seeded at 3.5 × 104 cells/cm2, attached poorly. This was not from toxicity of the PEM compounds, because cells could adhere and spread at the bottom of the same well on the tissue culture plastic near the coated glass slide (not shown). It was also not from loss of function of FN from its adsorption to PEM substrate. We tested the mouse myoblast cell line C2C12, which was previously used to examine effects of substrate-dependent changes in FN conformation for integrin ligation and cell adhesion on synthetic surfaces. 22 All native (PLL/HA)3,6,9-PLL-FN multilayers were adhesive for C2C12 cells, and under the high seeding density conditions of this experiment, complete cell monolayers formed within 24 h (Fig. 2B). We concluded that native, as-built (PLL/HA) i -PLL-FN layers were inadequate surfaces for MSC adhesion.
Interaction of MSCs with cross-linked PEM substrates
Recent studies of PLL/HA multilayer surfaces have revealed that stiffening of bulk multilayers enhanced adhesion of smooth muscle cells, chondrosarcomas, or osteoblasts.9,10,24 Protocols have been described for controlled stiffening of PLL/HA bulk multilayers by converting the purely electrostatic cross-linking of the native multilayers (ionic interactions) to covalent cross-linking between ammonium groups of PLL with carboxylate groups of HA (amide bonds) using EDC in combination with sulfo-NHS.11,25 We tested (PLL/HA)9-PLL-FN multilayers cross-linked to different degrees by varying concentration of EDC from 0 to 800 mM in the EDC/NHS cross-linking mix for their ability to mediate PD-MSC adhesion (Fig. 3). Phase microscopy 72 h after cell seeding showed that a gradual increase of multilayer stiffness correlated with a gradual increase of well-spread, elongated PD-MSCs with well-organized actin fibers. MSCs on native (EDC 0) or low-cross-linked PLL/HA multilayers (EDC 8) remained roundish and loosely attached to the multilayers; they lacked actin fibers or displayed cortical actin organization. Visual inspection indicated that multilayer coatings formed under the highest cross-linking regime, EDC 800, were optimal for MSC adhesion. Quantitative colorimetric analysis of MSC densities confirmed that multilayer substrates formed with the EDC 800 regime performed significantly better (p < 0.05) than less-cross-linked substrates (Fig. 3B). Under such a high cross-link regimen, the Young's modulus of PLL/HA multilayers was previously shown to reach a plateau value of 400 kPa. 25 Phase microscopic follow-up of low-density MSC cultures on cross-linked multilayers showed that MSCs maintained their ability to proliferate (Fig. 4A). Cell counts obtained from DAPI-stained cultures revealed that PD-MSCs and AT-MSCs exhibited similar proliferation rates (Fig. 4B). The stiffening of the bulk layer produced a strong positive effect for cell adhesion and outgrowth for multilayers of all thickness (i.e. (PLL/HA)-PLL-FN constructs with three-, six-, and nine-layer pairs) (Fig. 5). According to gross visual analysis, the positive effect of cross-linking for MSC adhesion appeared comparable between multilayers of different thicknesses. MSCs seeded at high density (3.5 × 104 cells/cm2) formed dense PD-MSC and AT-MSC layers 48 h after seeding (Fig. 5A). Quantitative analysis revealed that PD-MSC densities on multilayers from six and nine PLL/HA pairs were significantly higher (p < 0.05) than those on three PLL/HA pairs (Fig. 5B). Multilayer thicknesses did not affect AT-MSC densities. Without cross-linking, cell densities were low (Fig. 5B). Table 1 summarizes the results.
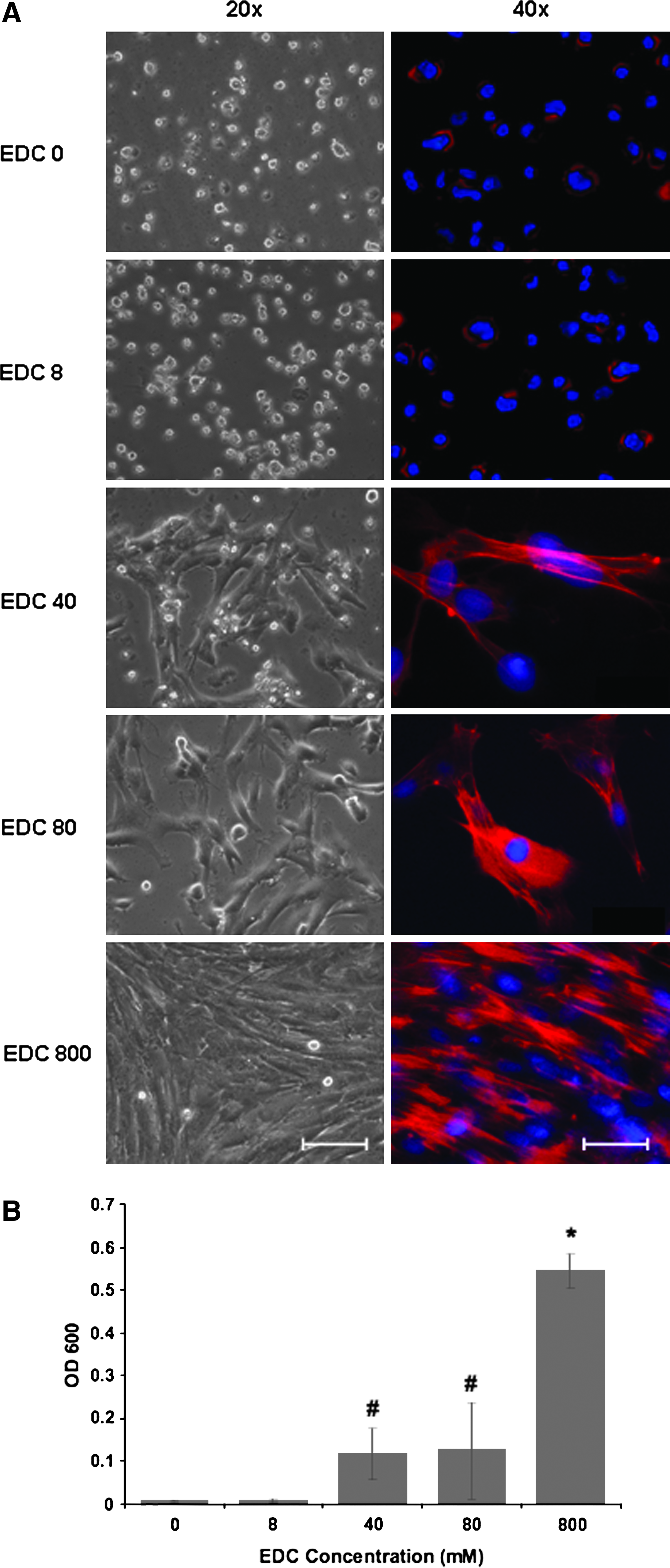
Stratification of stiffened multilayers for promotion of MSC adhesion. (
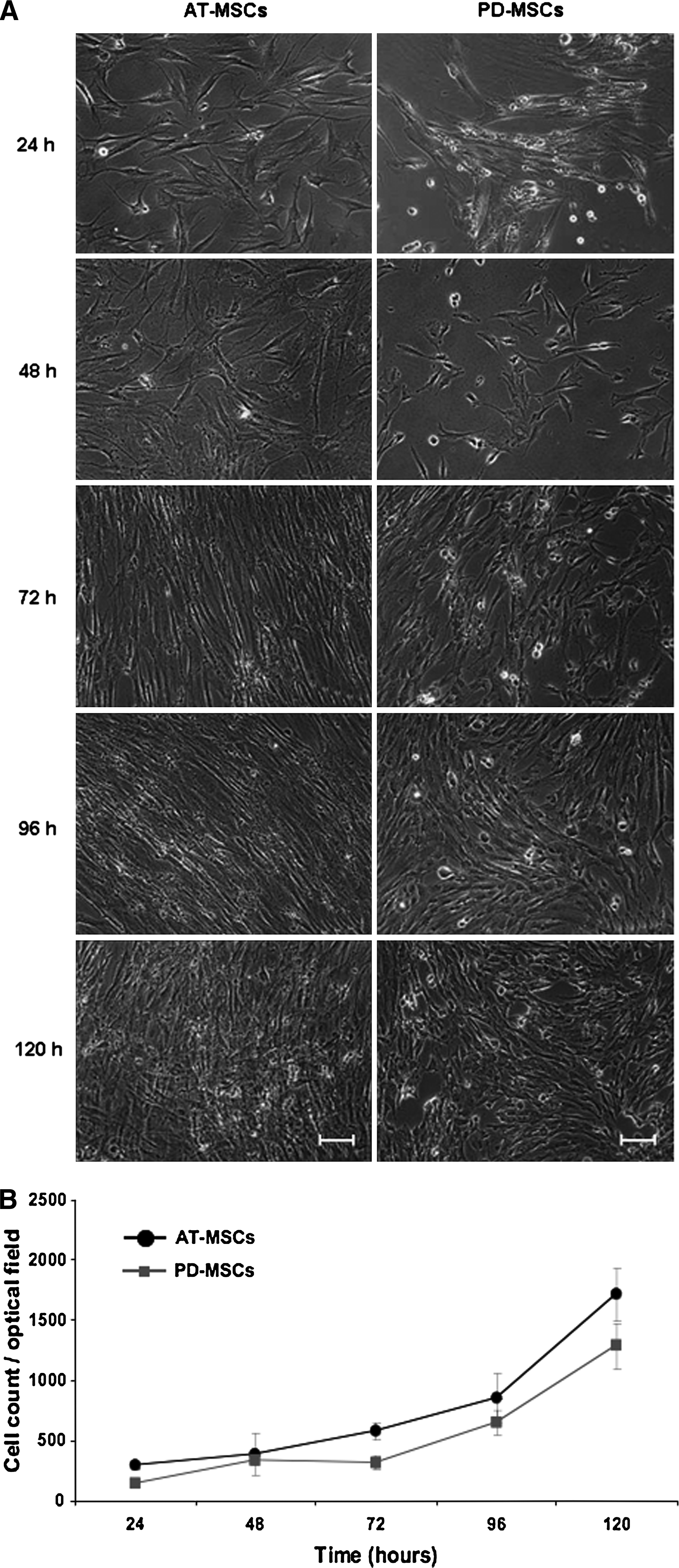
Proliferation of MSCs on cross-linked multilayers. (
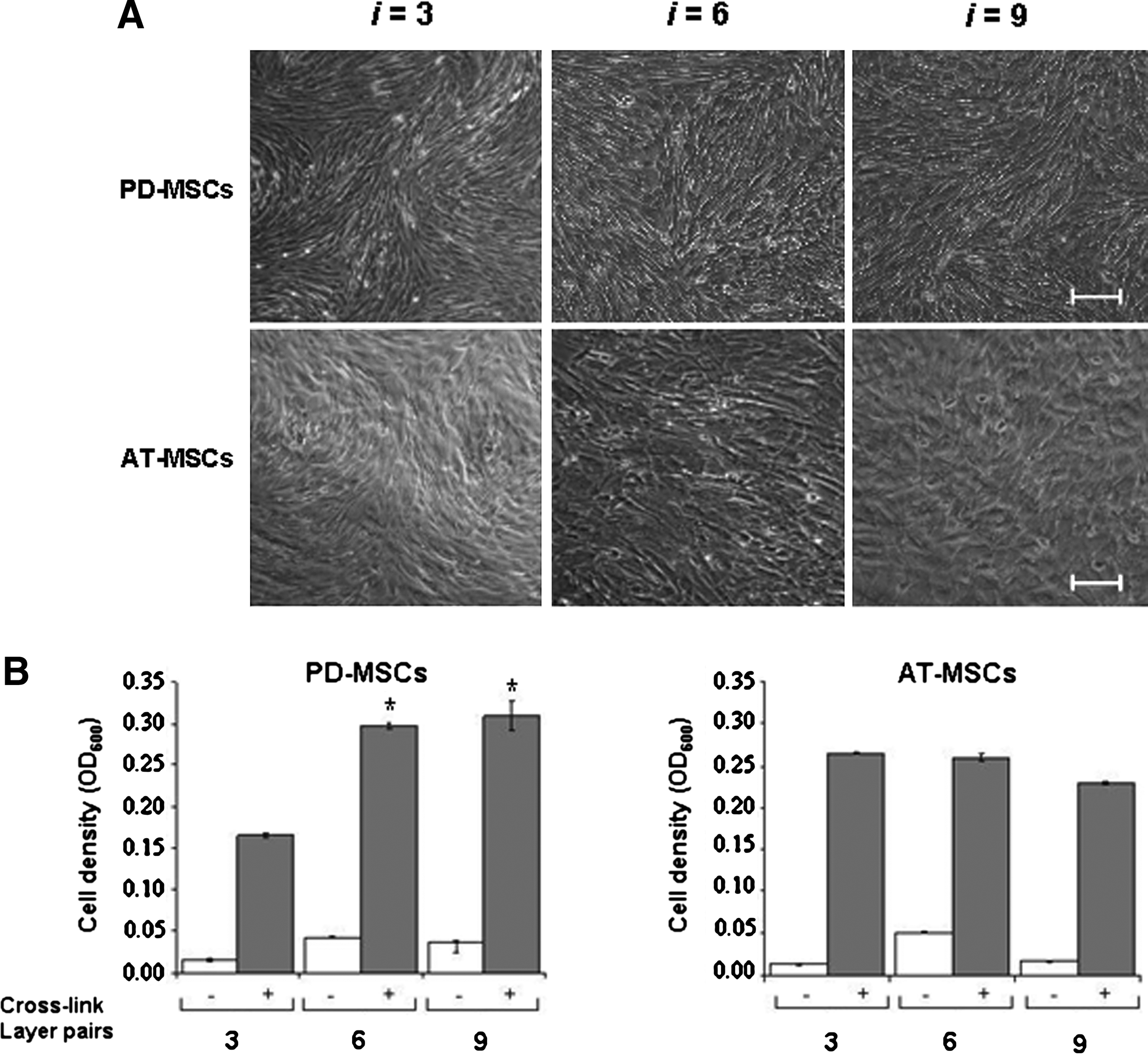
Stratification of multilayer thickness for MSC adhesion. (
The effect of cross-linking for the FN interface
We next investigated the effect of cross-linking for the FN outer layer. For that, we compared the distribution of FN on cross-linked and native (PLL/HA)9 layers, using fluorescently (Alexa Fluor 488, Invitrogen AG, Basel, Switzerland) labelled FN, confocal microscopy, and data processing with Imaris imaging software. Three layer preparations with FN were compared (Fig. 6): native bulk layer with physically pre-adsorbed FN, cross-linked bulk layer followed by physically pre-adsorbed FN, and cross-linked bulk layer with cross-linked FN. This analysis revealed different FN surface patterns. Top- and side-view images of Figure 6 illustrate the different FN interfaces. Native and cross-linked PEM layers with physically adsorbed exhibited a diffuse, spotty, and spiky surface distribution of FN (Fig. 6A, B); by contrast, chemically coupled FN appeared condensed in a narrow zone (Fig. 6D–F). At the resolution of our confocal microscope, the chemically coupled FN appeared as fine regular mesh forming from coalescing droplets. The condensed distribution of chemically coupled FN suggests greater FN surface density at the interface with cells. The stiffening of PEM layers by cross-linking along with concomitantly greater FN ligand density at the cell-PLL/HA coating interface had a profound effect on focal adhesion formation of MSCs, as demonstrated by immunofluorescence staining for vinculin, a component in the focal adhesion protein complex (Fig. 7). MSCs grown on cross-linked PEM layers, but not those on native PEM layers, showed punctate expression of vinculin at the tips of elongated cell structures (Fig. 7A, B). These results indicate that the functionality of FN for binding to the FN adhesion receptor, integrin α5β1, on cells was not affected. MSCs did not attach and spread well on cross-linked PLL/HA layers with physically adsorbed FN (data not shown). Parallel vinculin staining of the murine C2C12 myoblasts also showed focal adhesions on native layers and little effect of cross-linking the bulk layer and FN (Fig. 7C, D). However, we noted that cell areas of C2C12 cells on native layers often appeared larger than on cross-linked layers, and cells exhibited many filopodia. In conclusion, cross-linked PLL/HA multilayer substrates formed with cross-linked FN possess better adhesion properties for MSCs.

Cross-linking of PLL/HA layers affects FN surface distribution. Confocal microscopy of native or cross-linked (PLL/HA)9 multilayers formed with a physically adsorbed or chemically coupled FN outer layer. The green color represents the fluorescently labeled FN. (
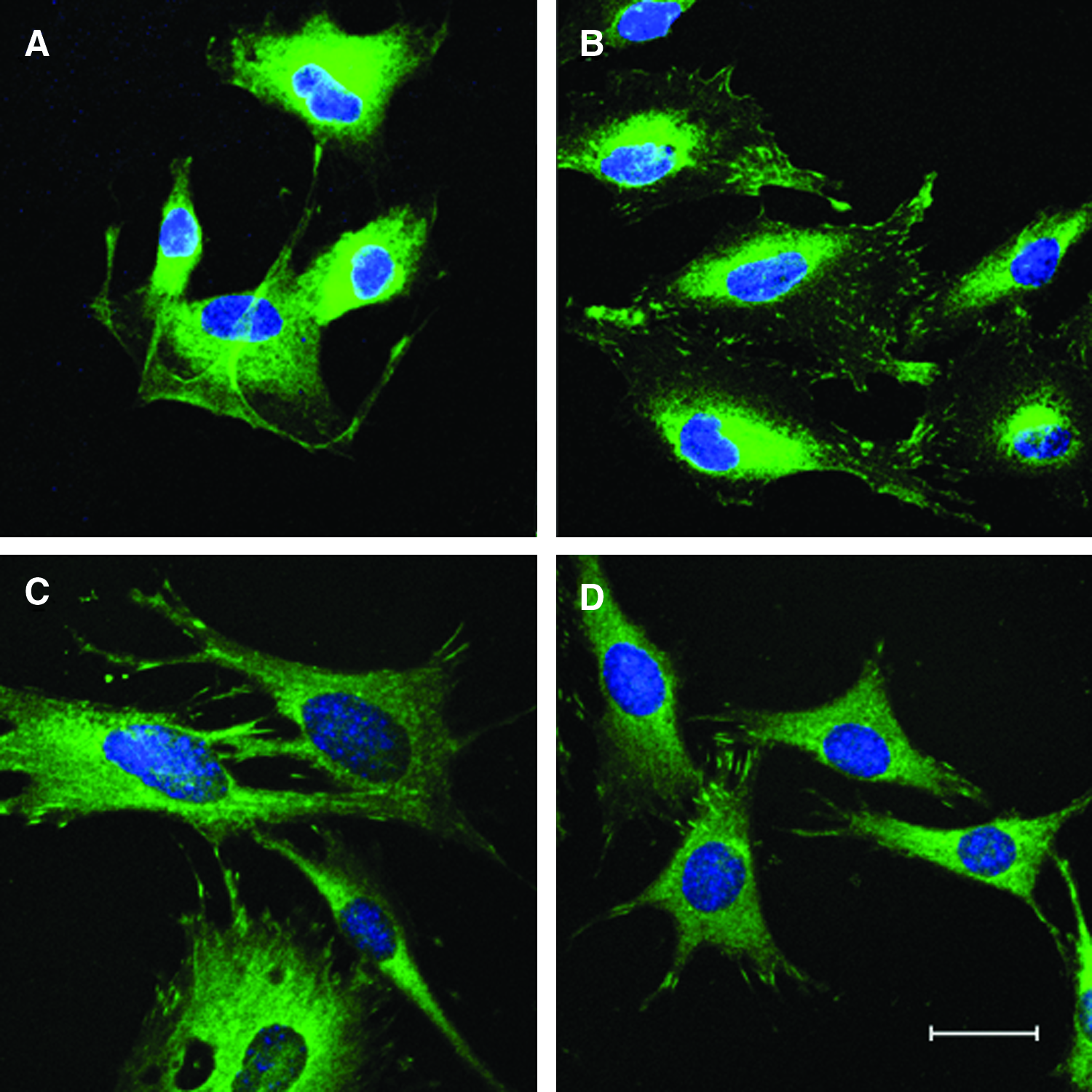
Immunofluorescent staining for focal adhesion formation on native and cross-linked FN-terminated (PLL/HA)9 layers. Cells were stained for focal adhesion protein vinculin (green) and cell nuclei counterstained with DAPI (blue). Confocal images of PD-MSCs on (
The role of FN for stability of MSC cultures on PLL/HA multilayer substrates
Our next steps of investigation concerned long-term stability of densely grown MSC cultures and their differentiability on PLL/HA multilayers. We examined the role of the outer layer of FN for stability of dense MSC cultures, comparing side-by-side physically adsorbed FN with covalently affixed FN. Covalent binding of FN to multilayers was critical for maintenance of dense MSC cultures, whereas purely physical FN adsorption performed insufficiently (Fig. 8B, C). Only MSC cultures on covalently cross-linked FN were complete after 1 week (Fig. 8B). Beyond this period, the cultures gradually began to detach focally (holes in the cell layer), especially under the conditions of differentiation culture in osteogenic or chondrogenic medium; spontaneous lift of MSC layer as sheets rolling up from the edges was occasionally observed (Fig. 8C). PD-MSC and AT-MSC (data not shown) cultures on cross-linked (PLL/HA)9-PLL-FN remained capable of differentiating into mesodermal lineages, as demonstrated by biochemical stainings for chondrogenic and osteogenic differentiation markers, respectively (Fig. 9).
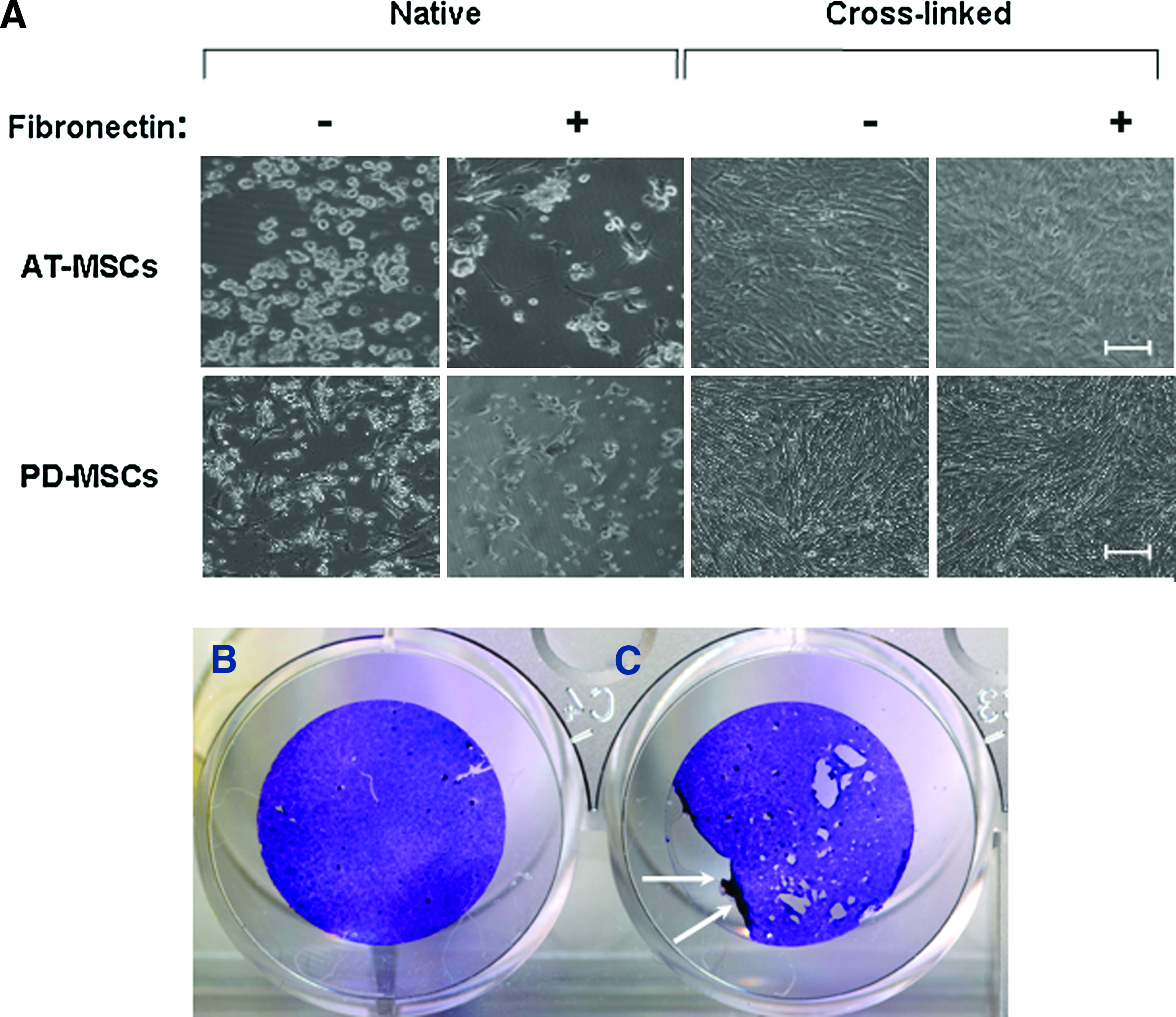
The role of terminal multilayer-bound FN for initial MSC monolayer formation. (
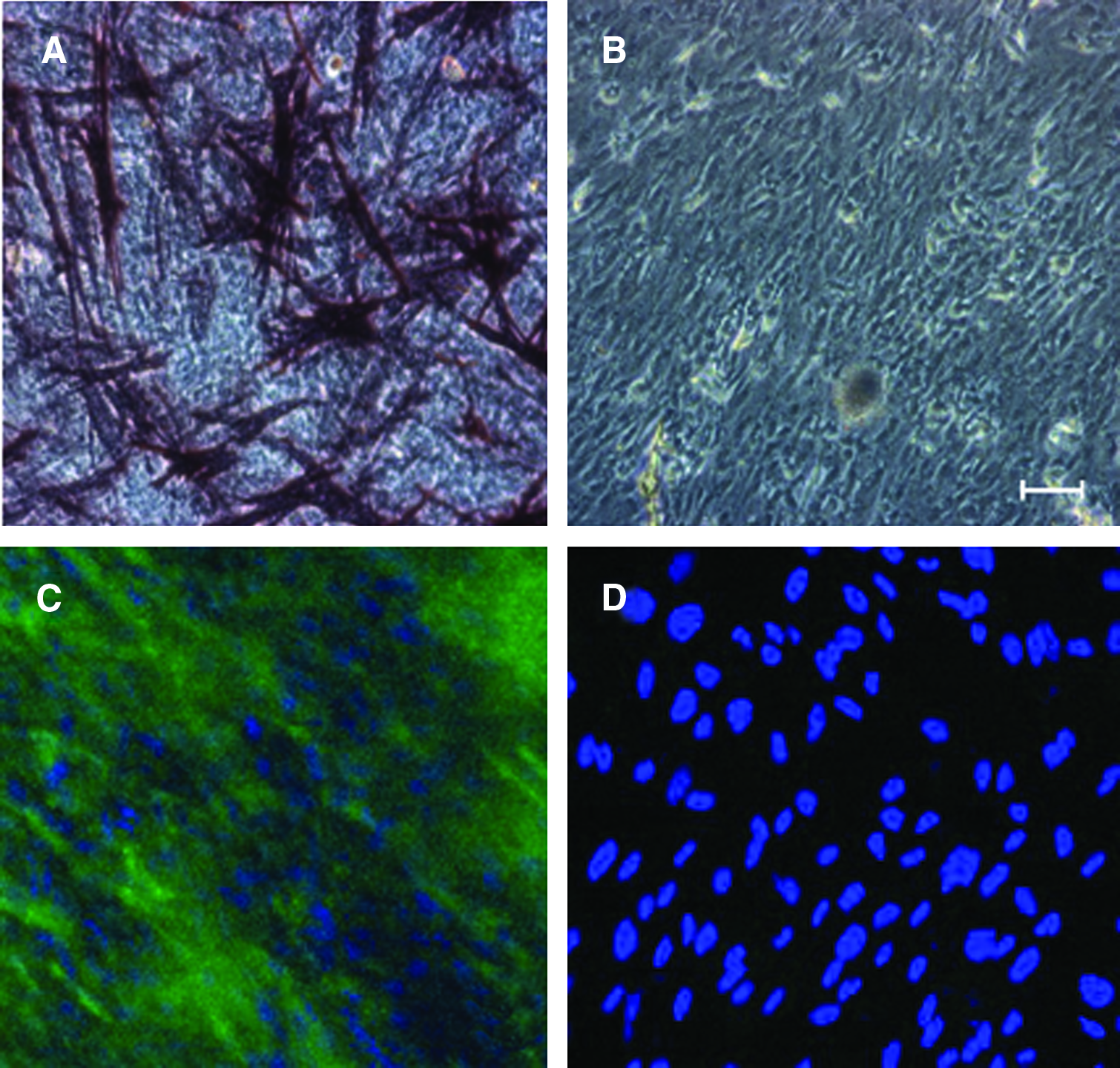
Multilayer-bound MSCs remain capable of mesodermal lineage differentiation upon culture with soluble induction factors. Images show PD-MSC cultures on (PLL/HA)9-PLL multilayers with covalently affixed FN. (
Discussion
Considerable knowledge about PLL/HA coatings has been developed over recent years, including for coatings from PLL and HA. In this report, we examined the utility of PEM coatings as a platform for interaction with human MSCs. Their physical and biological properties to promote adhesive interactions with human MSCs were successfully designed and demonstrated. We demonstrated that MSCs interacting with covalently cross-linked, FN-terminated PLL/HA multilayers continue to proliferate and remain differentiable. These are prerequisites for the use of PEM platforms for study of MSCs in biotechnological or even therapeutic applications.
For full PEM-driven control of cellular response, continuous PEM coating was sought to minimize influences from the underlying glass substrate on which the PEM coatings were built up. Deposition of eight-layer pairs of PLL/HA was previously found to be sufficient to produce continuous PEM coating with a smooth surface, 3 which was confirmed under our deposition conditions. This can be readily appreciated from the image given in Suppl Fig. 1, showing a (PLL/HA)9 coating with an experimental scratch to prove the coating. PLL/HA layer pair numbers did not influence MSC adhesion in the native or cross-linked condition (Fig. 3), which suggests that the influence of the glass substrate on MSC adhesion response was minimal.
The mechanical properties, not the thickness, of PLL/HA multilayers were critical for MSC adhesion. As shown in earlier studies, post-formation chemical cross-linking of PEM layers with EDC/NHS limits polyelectrolyte diffusion inside the bulk PEM structure and increases its stiffness without significant change of PEM thickness. 9 FN-terminated PLL/HA multilayers processed with a high cross-linking regime promoted PD-MSC and AT-MSC adhesion instantly and efficiently, whereas native multilayers, which are soft, 25 were almost adhesion-resistant to MSCs. Our findings add to the broad notion that adhesive spreading of cells correlates with the effective stiffness of PLL/HA, as well as other PEM coatings, and more generally with the stiffness of synthetic material substrates. How MSCs probe and adapt to the stiffer PLL/HA substrate is beginning to be understood. As noted in, 26 focal adhesions provide MSCs with the necessary force transmission pathways to “feel” their gel matrix microenvironment through actin-myosin contractions. Stiffer substrates produce stiffer and increasingly tense cells. 26 Studies with murine NIH3T3 fibroblasts plated on FN-coated polyacrylamide gels showed that cells change their internal stiffness, governed by cytoskeletal assembly and production of internal stresses, to roughly match the stiffness of the substrate. 27 Some cell types seem capable of anchoring even to soft multilayer substrates. We found that mouse C2C12 myoblasts, unlike the MSCs, adhered and spread on soft, native PLL/HA multilayers (Fig. 2B) and formed vinculin-positive focal adhesions (Fig. 7C, D). Reasons for different adhesion responses of MSCs and C2C12 cells on native multilayers with surface-adsorbed FN remain speculative. It may be the result of different “inside out” signaling pathways in C2C12 cells and MSCs that lead to different activation stages of the responsible FN adhesion receptor, α5β1. A two-stage activation mechanism for α5β1 binding to surface adsorbed FN was described. 28 Another possibility is that adsorption of FN to a PLL/HA multilayer substrate produces a conformational change in FN and that this altered FN is bound by α5β1 on C2C12 cells and MSCs at different levels. Studies by Garcia et al. have shown that adsorption of FN to substrates changes FN conformation and subsequently alters its binding to α5β1. 22 Finally, C2C12 myoblasts may possess an inherent natural capacity to comply with soft substrate environments. In support of the latter, emerging knowledge suggests that cellular responses to substrate stiffness (e.g., for maximal spreading or differentiation) are cell-type dependent in that the effective range of substrate stiffness depends on the tissue type from which the cells were derived. 29 The reported value for elasticity of striated muscle tissue is approximately 8 to 17 kPa, 30 which is within the range of elasticity reported for native PLL/HA coatings. 31 Hence, it is conceivable that C2C12 myoblasts by nature are better prepared to attach and spread on soft PEM substrates.
Recent studies have shown that tissue-level matching substrate stiffness strongly affects lineage specification and morphology of human MSCs. 26 A simple alteration of substrate compliance evoked specific MSC differentiation (e.g., soft polyacrylamide gel substrates that mimic brain were neurogenic, whereas stiff substrates that mimic collagenous bone were osteogenic). 30 In light of this emerging knowledge, PLL/HA multilayer substrates may offer great opportunities for compliance-specific differentiation assays of MSCs, because the Young's modulus of PLL/HA multilayer coatings can be readily modulated over a range from 0 to 400 kPa, depending on their cross-linking degree. 25 Along these lines, Ren et al. reported that modulation of stiffness of PLL/HA multilayer substrates in turn modulated C2C12 myoblast cell differentiation and their organization into myotubes. 32
We investigated the role of FN outer layers for MSC culture initiation, focal adhesion formation, and maintenance of densely grown MSC cultures. PLL-terminated multilayers were chosen for study, because other studies suggested that positive-terminal PEM layers favorably influence the distribution of FN molecules by inhibiting their aggregation in the adsorption layer, unlike negative-terminal layers. 33 With regard to adsorbed FN mass, our OWLS measurements revealed that the negatively charged FN adsorbed to terminal layers of HA at a similar level, despite the negative charge and hydrated nature of the latter (Suppl Fig. 2). Our confocal microscopic analysis of the FN interface revealed that cross-linking of FN-terminated PLL/HA layers changed the FN distribution from diffuse to condensed, resulting in greater presentation of FN adhesion sites to cells. Our results showed that the details for preparation of the FN interface on PLL/HA coatings matter greatly, at least for MSC culture. Only bulk layers with cross-linked FN were effective substrates for MSCs, which indicates that cross-linking does not affect the function of FN. It is likely that the stiffening by cross-linking plus the greater FN ligand density at the cell–PEM layer interface resulting from cross-linking synergistically accounts for the enhanced MSC spreading and focal adhesion formation (Figs. 6, 7). Physically adsorbed and chemically coupled FN were found distributed only at the top of the PLL/HA coating. This confirms studies of FN adsorption by Wittmer et al. 23 that have indicated that FN rapidly complexes with PLL with little or no FN penetration into the PEM bulk. Covalent fixation of FN to PLL/HA coatings proved necessary to maintain densely grown MSC cultures during differentiation culture over weeks (Fig. 8B). The amounts of FN adsorbed to PLL/HA multilayers, as determined according to OWLS, were 350 to 700 ng/cm2 (Table 1). These amounts favorably compared the (high) plateau densities of adsorbed FN measured on bacterial or tissue culture polystyrene. 22 FN is known as flexible glycoprotein with substrate-sensitive conformations, which in turn can modulate cell proliferation and differentiation. 22 Our data indicate that EDC/NHS-mediated cross-linking of multilayer-bound FN does not impair its functionality. Similar observations were made for collagen coatings cross-linked with EDC/NHS, which led to greater endothelial cell adhesion and proliferation on vascular grafts than with physically adsorbed collagen. 34
Innovative use of charge-responsive polyelectrolyte coatings 20 could provide a new means for non-enzymatic harvest of MSCs and their transplantation as intact, cohesive cell sheets into diseased tissues, a concept known as “cell sheet engineering.” 35 Data from the laboratory of one of us (J.V.) have demonstrated that monolayered polycationic copolymer poly(l-lysine)-grafted-poly(ethylene glycol) that were formed on conducting metal oxide substrate desorbed from their metal oxide substrate after the application of a positive potential, lifting complete sheets of osteoblasts cultured on the polyelectrolyte monolayer. 20 Although the cross-linked, FN-terminated PLL/HA multilayer coatings under present study permitted formation of complete MSC layers, they do not dissolve upon electrochemical polarization (unpublished observations). Further modifications of cross-linked PLL/HA multilayers might allow for electrochemically controlled lift of MSC sheets for transplantation, which awaits addressing in future experiments.
Footnotes
Acknowledgments
This work was funded by a grant of the Competence Centre for Materials Science and Technology (CCMX) of the ETH-Board, Switzerland, and the European Commission FP6-grant ‘Crystal.’ We thank Dr. Tomaso Zambelli and Orane Guillaume-Gentil for assistance with OWLS, Drs. Fouzia Boulmedais and Pierre Schaaf for helpful suggestions, and Esther Kleiner for technical assistance with histology.
Disclosure Statement
No competing financial interests exist.
Part of this paper was presented at the Tissue Engineering and Regenerative Medicine International Society Meeting (TERMIS)-EU Annual Meeting 2008, June 22–26, Porto, Portugal. The published meeting abstract is: Semenov OV, Malek, A, Voros J, Zisch AH. Stem cell sheet engineering on polyelectrolyte multilayer films. Tissue Engineering Part A 14:779 (2008).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.