
Abstract
Recently, bone marrow–derived mesenchymal stem cells (MSCs) have been paid more attention for cartilage regeneration. This study evaluated the potential of using MSCs seeded in plasmid transforming growth factor β1 (pTGF-β1)–activated three-dimensional chitosan/gelatin scaffolds for improving cartilage repair in vivo. Significant cell proliferation and transforming growth factor β1 protein expression were observed in vitro in pTGF-β1-activated scaffolds. Transforming growth factor β1–activated scaffolds showed high collagen type II and aggrecan expression and low collagen type I expression during in vitro cultivation. MSC-based pTGF-β1-activated scaffolds also exhibited cartilage histology with high secretion of collagen type II in vitro under the stimulation of pTGF-β1. In rabbits with full-thickness cartilage defects, the implantation of MSC-based pTGF-β1-activated scaffolds not only significantly promoted chondrogenic differentiation of MSCs and hyalin-like cartilage matrix synthesis, but also remarkably improved the overall repair of rabbit cartilage defects and exhibited favorable tissue integrity at 10 weeks postsurgery. These results suggest that MSC-based localized pTGF-β1-activated scaffolds have potential applications for in vivo cartilage repair.
Introduction
Cell-based cartilage tissue engineering is a promising future alternative to current surgical practices. As a cell source for cartilage tissue engineering, autologous chondrocytes have limited doubling potential, easily lose their phenotype after multiple passages on culture dishes, and cease to produce cartilage extracellular matrix (ECM) so that degradation of repaired tissue can be seen, especially in a long-time clinical observation.8–10 Bone marrow–derived mesenchymal stem cells (MSCs) are adult multipotent cells with the capacity to differentiate into cells of osteogenic, chondrogenic, adipogenic, and myogenic lineages, and recently are considered as a promising cell pool for cartilage repair by virtue of their easy isolation, extensive expansion, and high chondrogenic potential.11–13
Although regeneration of cartilage using MSCs is impressive, sufficient hyaline repair of cartilage is not always achieved. 14 Many lines of evidence show that growth factors, such as transforming growth factor β1 (TGF-β1), TGF-β3, fibroblast growth factor, and insulin-like growth factor 1 (IGF-1), are needed and used as triggers of chondrogenesis of MSCs.15–18 However, the biggest issue with formulations intended for long-term controlled release is the instability of the growth factor proteins themselves; their short half-lives in vivo require them injected repeatedly. 19
The gene delivery system is an alternative to the delivery of growth factors because DNA is more stable and flexible than protein, which is able to provide the capability to achieve sustained and localized expression of growth factors to cartilage defects. 20 Hosseinkhani et al. showed that delivery of plasmid BMP-2 into MSCs grown in three-dimensional PGA/collagen scaffolds could significantly promote target gene expression and enhance osteogenesis in vitro. 21 Capito et al. found that plasmid IGF-1-transfected chondrocytes could present enhanced biosynthesis and chondrogenesis in modified collagen scaffolds in vitro. 22 In some studies, cationic liposomes were used as gene delivery carriers, but they were not ideal local gene delivery systems because of their toxicity, poor controllability, and operability in complicated microenvironments in vivo.23,24 Thus, till now, there has been few report regarding the use of liposomes to deliver therapeutic genes in vivo in cartilage tissue engineering.
Our previous studies have showed that plasmid transforming growth factor β1 (pTGF-β1)–activated scaffold can promote chondrocytes proliferation in vitro, 25 as well as help cartilage reparation in vivo. 26 The scaffolds consist of chitosan and gelatin, which are chosen to highly simulate cartilage ECM, glycosaminoglycan, and collagen, and provide a favorable environment for chondrogenesis. Most importantly, cationic chitosan can serve as a gene carrier that could facilitate DNA into MSCs. Accordingly, DNA release can be controlled depending on scaffold composition. 27 In this study, we evaluated the chondrogenic potential of MSC-based pTGF-β1-activated chitosan/gelatin scaffold in cartilage tissue engineering. Such a scaffold acted as both a therapeutic gene carrier without any toxicity and a tissue-engineered matrix that could maintain the sustained localized expression of TGF-β1 protein to induce chondrogenic differentiation of MSCs. Our results in vitro showed that MSCs in pTGF-β1-activated scaffold achieved robust expression of chondrogenic phenotype. In adult rabbit full-thickness cartilage defect model, MSC-based pTGF-β1-activated scaffold could significantly promote chondrogenic differentiation of MSCs and hyalin-like cartilage matrix synthesis, remarkably improve the overall repair of rabbit cartilage defects, and exhibit favorable tissue integrity after 10 weeks. Altogether, our results suggested that MSC-based localized pTGF-β1-activated scaffolds have potential applications for cartilage tissue engineering.
Materials and Methods
Animals
Twenty-one 4-month-old New Zealand white rabbits 28 (Jinling Hospital, Nanjing, China) were fed a standard diet and acclimated in a quiet quarantine room for 1 week before the experiments. All rabbits received humane care in accordance with the Guidelines for the Care and Use of Laboratory Animals published by the National Institutes of Health (NIH publication No. 85–23, revised 1985).
MSC isolation and culture
MSCs were obtained from New Zealand rabbits according to the technique reported. 29 Briefly, the heparinized bone marrow aspirates obtained from the upper tibial metaphysis of rabbits were centrifuged at 1200 rpm for 20 min, and the cell pellets were plated overnight in T-50 cm2 cell-culture flasks (Greiner, Frickenhausen, Germany) in a basal medium composed of low-glucose Dulbecco's modified Eagle's medium (Life Technologies, Grand Island, NY) supplemented with 10% fetal bovine serum (Life Technologies) and antibiotics (penicillin 100 units/mL and streptomycin 100 μg/mL; Life Technologies), and maintained in an incubator of 5% CO2 at 37°C. Unattached cells were removed after 24 h incubation by washing twice with phosphate-buffered saline (PBS, pH 7.4) and subsequent medium changes. After that, medium was replaced every 3 days until the cells reached 80% confluence. Then, the cells were washed twice with PBS, detached by treatment with 0.25% trypsin–ethylenediamine tetraacetic acid solution (Life Technologies), washed twice with expansion medium followed by centrifugation (1200 rpm for 5 min), and subcultured at 1:3 in triple flasks under the same conditions. The cells at passages 2–4 were used. For cell proliferation, for in vitro differentiation, and after rabbit cartilage repair, MSCs at passage 2 were used. For in vitro gene transfection, cells at passages 3–4 were used.
Fabrication of pTGF-β1-activated chitosan/gelatin scaffolds
The pTGF-β1-activated chitosan/gelatin scaffolds were fabricated as described previously. 25 Chitosan (derived from crab shell with minimum deacetylation degree of 85%, MW 4 × 105; Sigma-Aldrich, St. Louis, MO) was mixed with gelatin B (from bovine skin; Sigma-Aldrich) in 1% acetic acid solution (1% chitosan and 1% gelatin). The solution was stirred for 2 h at 50°C, poured into a stainless steel mold, frozen at −20°C for 1 h and −80°C for 2 h, and finally lyophilized for 48 h to produce a columned scaffold of approximately 2 mm diameter and 2 mm thickness. The scaffolds were immersed into a crosslinking solution of 1-ethyl-(dimethylaminopropyl) carbodiimide/N-hydroxy-succinimide/2-morpholinoethane sulfonic (15 mM/15 mM/50 mM, pH 5.4; Amersco, Solon, OH) for 4 h. After thorough washes with distilled water and desiccation, the scaffolds were sterilized by ethanol, equilibrated with 0.1 M sterilized Na2HPO4 for 10 min, washed with distilled water to a neutral pH, and lyophilized again.
Eighty microliters of sterilized plasmid TGF-β1 (0.5 μg/μL, pCMV-TGF-β1; Fulengen, Guangzhou, China) was dropped into a 20 mg scaffold and kept at 4°C overnight to allow DNA incorporation with chitosan/gelatin complex completely. Then, the plasmid TGF-β1-activated scaffolds were frozen into liquid nitrogen for 60 s and lyophilized. The same plasmid vector, containing coding sequence of EGFP instead of TGF-β1 sequence, was used as an empty plasmid control (0.5 μg/μL, pCMV-EGFP; Fulengena).
MSC seeding and proliferation in scaffolds
The scaffolds without plasmids above, with pTGF-β1 or pEGFP, were presoaked in fibronectin (100 μg/mL; Sigma-Aldrich) for 10 s to support MSC adherence. About 20 μL of MSC suspension was seeded drop by drop at a density of 2 × 105 cells/scaffold onto the surface of scaffold and placed into a 24-well tissue-culture plate. After 2 h incubation at 37°C to allow MSCs to diffuse into and adhere to the scaffolds, 1 mL of medium was added slowly to the plate wells, which precoated with 2% agarose (Sigma-Aldrich) to prevent cells from adhering to the plate. After 1, 2, 3, 5, 7, 10, and 14 days, using 0.2% trypsin and 0.2% collagenase II, gelatin and chondrocyte-specific ECM was degraded, and three-dimensional scaffolds were broken down. Cells in the scaffolds were digested, and the numbers were estimated by a hemacytometer. Scaffolds incubated for 2 weeks were used for in vivo cartilage repair in a rabbit model.
As reported previously,29,30 recombinant human TGF-β1 growth factor (rh-TGF-β1) could really stimulate chondrogenesis of rabbit MSCs. So another five MSC-seeded DNA-free scaffolds were incubated in chondrogenic medium containing rh-TGF-β1 1 ng/mL (PeproTech, Rocky Hill, NJ).
Scanning electron microscopy examination
MSC-seeded scaffolds were examined using scanning electron microscopy (SEM; Hitachi X-650, Tokyo, Japan) to observe their morphologies. Samples incubated for 3 days in vitro were washed twice with sterilized PBS, fixed in 10% formalin for 12 h, followed by dehydration through a graded series of ethanol for 30 s at each ethanol concentration, and lyophilized for 16 h. The samples were finally sputter coated with gold–palladium film (ratio 60:40) for SEM observation.
GFP and TGF-β1 expression in gene-activated scaffold in vitro
To validate that plasmid DNA really incorporates into cells in gene-activated scaffold, MSC-seeded pEGFP-activated scaffolds were washed by 1× PBS, frozen sectioned at 5 μm, and then observed under a fluorescence microscope after a 3-day culture (TE2000; Nikon, Nikon, Japan).
Commercially available ELISA assay kit (eBioscience, San Diego, CA) was used for TGF-β1 expression analysis in vitro. MSC-seeded scaffolds were incubated in 1 mL medium at 37°C. Then, the medium was collected and changed with fresh medium every 3–4 days over a period of 2 weeks. Four samples collected at each time point were for ELISA analysis. MSC-seeded scaffolds without plasmid and MSC-seeded pEGFP-activated scaffolds were used as control groups.
Real-time PCR assay
Engineered constructs incubated after 7 and 14 days were extracted for total cellular RNA using Trizol Reagent (Life Technologies) according to the manufacturer's protocol. RNA concentration was estimated spectrometrically on the basis of A260. RNA samples were reverse transcribed using Oligo dT primers and AMV transcriptase (Life Technologies). Gene expression analysis was detected using Eva Green in ABI Biomed 7300 Real-Time PCR System as described in the manufacturer's instruction. Gene-specific oligonucleotide primers included collagen type I (5-CAATCACG CCTCTCAGAACA-3, 5-TCGGCAACAAGTTCAACATC-3), collagen type II (5-CAACAACCAGATCGAGAGCA-3, 5-CCAGTAGTCACCGCT CTTCC-3), collagen type X (5-ATAAAGA GTAAAGGTATACC-3, 5-CTGTTGTCC AGGTTTTCCTG-3), aggrecan (5-TCACAATCTTCCAGGAGCGA-3, 5-CACAATG CCGAAGTGGTCG T-3), and glyceraldehyde-3-phosphate dehydrogenase (5-TCACCATCTTC CAGGA GCGA-3, 5-CACAATGCCGAAGTGGTCGT-3) as internal controls for mRNA loading. Relative gene expression levels were normalized to the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase.
Cartilage repair studies in a rabbit model
Four-month-old New Zealand white rabbits with full-thickness cartilage defects were prepared as models for cartilage repair study as described previously. 26 A consistent cylindrical defect (3 mm diameter × 4 mm depth) was created in the middle of each patellar groove with a cylindrical drill (2 mm diameter). Then, the defect in one knee was filled with MSC-seeded scaffold with pTGF-β1 as engineered cartilage repair group (n = 15), and MSC-seeded scaffold without pTGF-β1 filled in the contralateral defect was used as a control (n = 15). Additional six rabbits (n = 12) were used as defect groups without any treatment on both knees. Subsequently, patella was repositioned, and trochlea, muscle, and skin were closed with sutures. Cartilage repair results were observed and evaluated by the modified O’ Driscoll score system after 3, 7, and 10 weeks.
Histological analysis
MSC-seeded scaffolds incubated for 2 weeks in vitro were washed in PBS, fixed in 4% phosphate-buffered paraformaldehyde at 4°C for 1 h, dehydrated through a graded series of ethanol, embedded in paraffin, and sectioned at 10-μm thickness.
The rabbits were sacrificed at 3, 7, and 10 weeks after transplantation; all knees were retrieved, washed in PBS, fixed in 10% phosphate-buffered formalin (pH 7.4) for 24 h, decalcified in 20% EDTA for 2 weeks, dehydrated through a graded series of ethanol, embedded in paraffin, and sectioned at 10 μm on the slides.
For histological analysis, slides were stained with hematoxylin and eosin (H&E). Immunohistochemical staining was used to detect collagen II secretion. Slides were blocked with 5% bovine serum albumin (Boster, Wuhan, China) at 25°C for 20 min, incubated with 10 μg/mL mouse anti-rabbit primary antibody of collagen II (Calbiochem, Darmstadt, Germany) at 4°C overnight, washed with PBS, incubated with biotinylated goat anti-mouse secondary antibody (Boster) at 37°C for 20 min, washed with PBS, and reacted with Streptavidin–Biotin Complex (SABC; Boster) at 37°C for 20 min. Finally, the revelation was performed using Diaminobenzidine (DAB; Boster) as the chromogen.
As a His-tag was added to the TGF-β1 sequence, cellular immunochemical analysis was used to discriminate endogenous and exogenous TGF-β1. After a 3-day culture, pTGF-β1-activated scaffolds were sectioned at 5 μm, and then detected by mouse anti-His-tag monoclonal antibody (Santa Cruz Biotechnology, Heidelberg, Germany).
Statistical analysis
Each experiment in vitro was performed in triplicate for every time point. Data were presented as mean ± standard deviation (SD) and analyzed statistically using two-way analysis of variance (ANOVA) with significance set at p-value <0.05.
Results
Characterization of MSC-based scaffolds
Figure 1 shows the SEM pictures of different chitosan/gelatin scaffolds. As shown in Figure 1A, the scaffold appeared porous, and its pore size was about 50–100 μm, which would benefit for DNA incorporation and MSC culture. Figure 1B clearly shows the inner structure of the pore in the porous scaffold. MSCs cultured on scaffold looked round as shown in Figure 1C (× 300) and in Figure 1D (× 900).

Cross-sectional SEM photographs of scaffolds. (
EGFP expression and exogenous TGF-β1 detection
As shown in Figure 2A, the gene-activated scaffold could really deliver plasmid EGFP into MSCs and facilitate GFP expression.
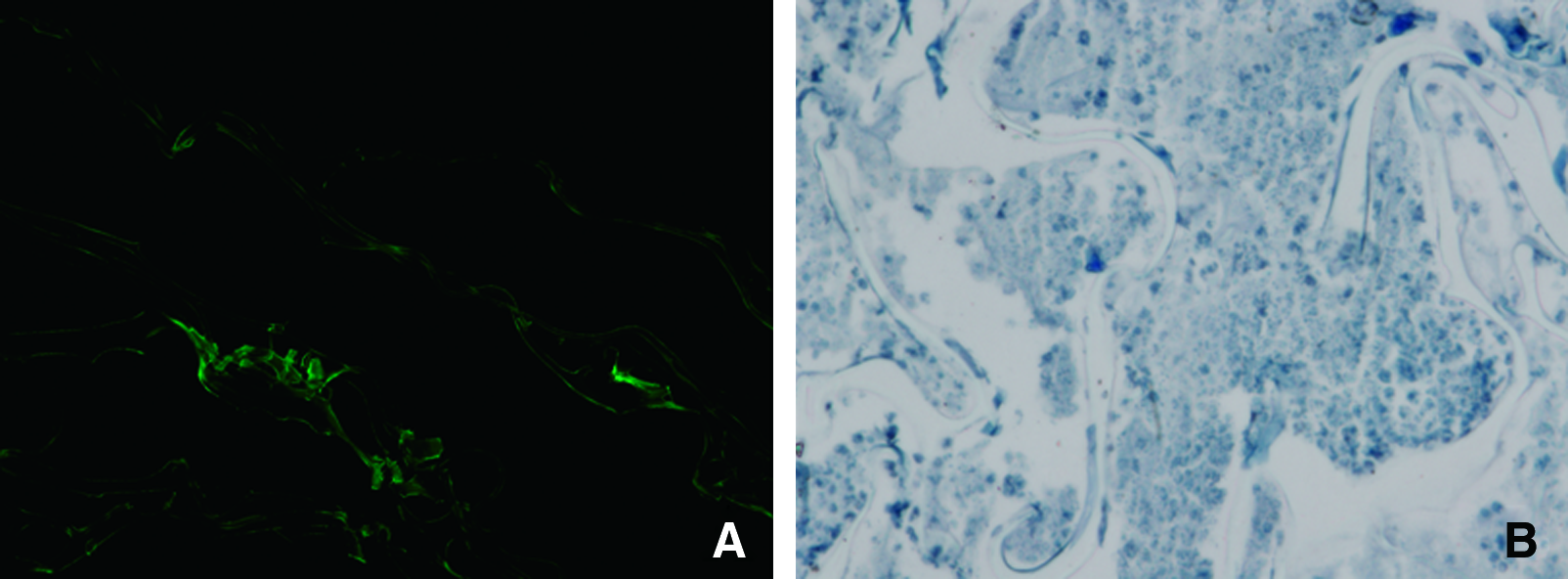
EGFP expression and exogenous TGF-β1 detection in gene-activated scaffold in vitro. (
His-tag was detected by cellular immunochemical assay, which demonstrated that secreted TGF-β1 (blue staining in Fig. 2B) from gene-activated scaffolds was not endogenous.
Cell proliferation assay
As shown in Figure 3, the cell number of all the three groups gradually increased with time. It is demonstrated that all three kinds of scaffolds were advantageous for cell viability. However, there are still obvious differences among them. After cultured from 5 to 14 days, in DNA-free scaffold group, cell number increased from 2 × 106 to 8 × 106, while in control group using empty plasmid DNA (pEGFP-activated scaffold group), it increased from 3 × 106 to 1 × 107, and in pTGF-β1-activated scaffold group, cell number increased from 4 × 106 to 1.2 × 107.

MSC proliferation analysis in chitosan/gelatin scaffold in vitro. Data are presented as mean ± SD (n = 3).
Effect of in situ TGF-β1 gene expression
To evaluate the effect of plasmid TGF-β1-activated scaffold as a localized gene delivery system in vitro, TGF-β1 gene release and expression on MSC-based scaffolds was carried out. TGF-β1 gene release as a function of time was a controlled process, which was the same as our previous result (data not shown). As shown in Figure 4, the TGF-β1 expression level in DNA-free and pEGFP-activated scaffolds were nearly equivalent during 2-week cultivation. However, in pTGF-β1-activated scaffolds, the protein secretion showed a significant increase compared to controls. The TGF-β1 protein reached the maximum value at the seventh day and was about twofold higher than controls after 2 weeks.

TGF-β1 expression analysis by ELISA assay in MSC-based scaffolds. Data are presented as mean ± SD (n = 4).
Hyaline cartilage-specific genes expression
Quantitative gene expression of cartilage-specific genes in all three kinds of scaffolds was analyzed by real-time PCR. As shown in Figure 5, pTGF-β1-activated scaffold showed a significant change in cartilage-specific genes expression. After 14 days, gene expression of collagen II (Fig. 5A) and aggrecan (Fig. 5B) in the pTGF-β1-activated group was much higher than that of two control groups. Almost a 13-fold increase in collagen II gene expression and about a 5-fold increase in aggrecan gene expression could be observed in the pTGF-β1-activated group compared with other two groups. However, collagen I gene expression was lower in the pTGF-β1-activated group at days 7 and 14 than those in control groups that indicated a low trend of fibrogenesis with the presence of pTGF-β1. Collagen X, which is a hypertrophic chondrocyte-specific factor, could not be detected in all three experimental groups.

Cartilage matrix gene expression in MSC-based scaffolds analyzed by real-time PCR after 7 and 14 days, respectively. Gene expression was analyzed by real-time PCR for (
Immunohistochemical staining of MSC-based scaffolds in vitro
As the major component of cartilage-specific ECM, collagen type II is the most important marker of chondrogenesis, and immunohistochemical staining assay of collagen type II was carried out as shown in Figure 6. In pTGF-β1-activated scaffold after a 2-week culture, increased staining of collagen II was observed (Fig. 6C), consistent with the accumulation of cartilaginous ECM, while the staining of the DNA-free scaffold was weak as shown in Figure 6B. Altogether, it is demonstrated that plasmid TGF-β1 in scaffold was very important for chondrogenic differentiation of MSCs in vitro. As a positive control, rh-TGF-β1 also had an effect on rabbit MSC chondrogenesis and stimulated more collagen II in vitro (Fig. 6D).
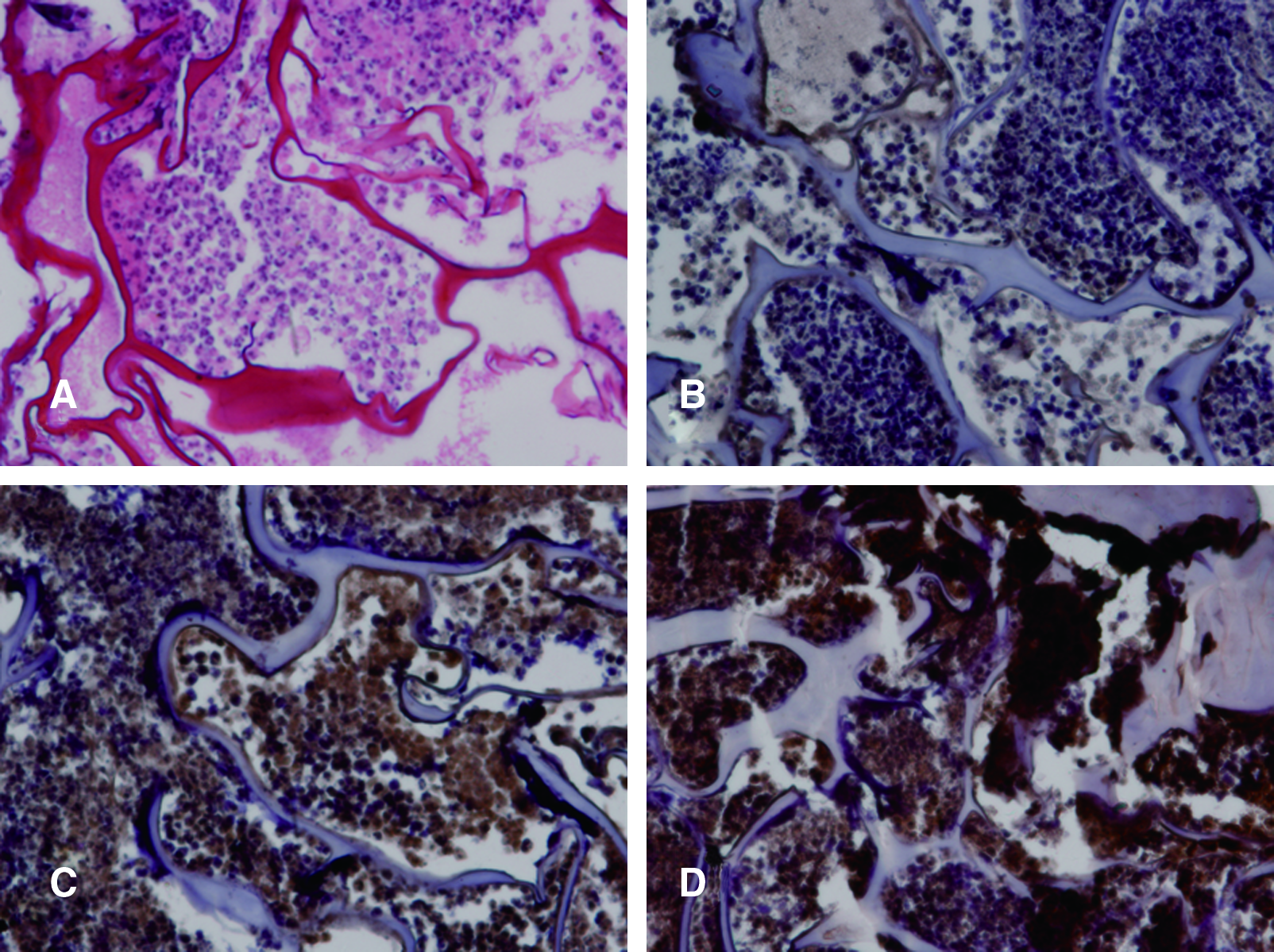
Cartilage matrix characterization by H&E and immunohistochemical staining of collagen type II after incubated for 2 weeks in vitro. (
Gross assessment of cartilage repair in vivo
Photographs of all specimens are shown in Figure 7. The defect in control group was almost empty after 3 weeks postsurgery, and it filled with a few reddish granular tissues at the seventh week (Fig. 7A and D, respectively). After 10 weeks, the flat-like surface and obvious gap around the defect could be seen (Fig. 7G). In contrast, after surgery for 3 weeks, the defect in the pTGF-β1-activated group was filled with translucent, newly formed cartilage-like tissue with recognized margin (Fig. 7C), whereas the defect in DNA-free group (Fig. 7B) was mostly depressed and not as smooth as that in the pTGF-β1-activated group. For the defect in the pTGF-β1-activated group, most of the margin around the defect was closed with new tissue after 7 weeks (Fig. 7F). In the DNA-free group, the new tissue in the defects was partly integrated with adjacent tissue, and it was somewhat loose to palpation (Fig. 7E). After 10 weeks, the newly formed cartilage in the pTGF-β1-activated group appeared tenacious and well integrated with the margin, and its gloss of surface was nearly similar to that of native cartilage (Fig. 7I), while the new tissue in DNA-free group seemed rough on the surface (Fig. 7H).

Macrophotographs of the articular cartilage repair in vivo in three experimental groups taken at the indicated times. No-treatment control group (
Histological analysis of cartilage repair in vivo
Figure 8 shows the histological sections stained by H&E, and collagen type II immunohistochemical staining during cartilage repair in vivo. Limited ECM could be observed by H&E staining in the control group after surgery for 3, 7, and 10 weeks (Fig. 8A, D, and G, respectively). The same result was also seen in Figure 8J, M, and P using collagen type II immunohistochemical staining, indicating that the intrinsic repair ability of cartilage was limited.

Histological (
In the pTGF-β1-activated group, stainings of ECM (Fig. 8C) and collagen type-II (Fig. 8L) were more hyaline-like than those in both no-treatment control and DNA-free group after 3 weeks. Meanwhile, cells in the pTGF-β1-activated group presented more rounded shape and showed cellular distribution resembling the distribution of chondrocytes in native cartilage (Fig. 8C), whereas some fibroblast-like cells with elongated nuclei could be observed in DNA-free group (Fig. 8B). After 7 weeks, thicker repaired cartilaginous layer (Fig. 8F) was showed in the pTGF-β1-activated group, as well as denser signal of collagen type-II (Fig. 8O); however, this characteristic was only observed in three of five defects in DNA-free group (Fig. 8E and N). At 10 weeks postsurgery, although the denser staining of ECM and collagen II could be detected in DNA-free group, there was formation of fibrous capsule in repaired tissues (Fig. 8H and Q). For comparison, extremely abundant collagen type II expression and complete chondrocyte-like cells surrounded by dense ECM was observed in the pTGF-β1-activated group (Fig. 8I and R), suggesting the efficient cartilage repair using the combination of MSCs and pTGF-β1-activated scaffold.
Histological scores
Representative sections through the center of defects were scored in triplicate using a modification of scoring system developed by O’ Driscoll et al. as shown in Table 1. The average histological scores of three groups were presented in Table 2. At each time point, compared with control and DNA-free group, higher average scores were presented in the pTGF-β1-activated group. After 10 weeks postsurgery, histological assessment of the pTGF-β1-activated group exhibited a good score of 23.2 ± 1.2, indicating that the improved cartilage repair was achieved using MSC-based tissue engineering.
Average score is statistically significant (p ≤ 0.05).
Discussion
Bone marrow–derived MSCs can overcome many of the problems associated with differentiation into chondrocytes and remodeling of new ECM.31,32 Here, we evaluated the potential of rabbit bone marrow–derived, MSC-based, pTGF-β1-activated chitosan/gelatin scaffolds, for improving cartilage repair in vivo, which appeared much better than autologous chondrocyte-based cartilage repair in our previous report. 26
Biocompatible chitosan and gelatin can be manipulated to achieve the porous scaffolds as seen in the SEM photographs; their morphological similarity to the ECM of the natural environment provides a structural support for MSC adhesion and a guidance for their growth and differentiation.33–35
Chondrogenesis is the earliest step that could affect a great many aspects of skeletogenesis. MSCs can differentiate into chondrocytes that express specific molecular markers such as aggrecan and collagen II. Differentiated MSCs can be distinguished from undifferentiated ones, which remain at the periphery of the skeletal element called perichondrium, 36 and then the innermost chondrocytes can further differentiate into hypertrophic chondrocytes that express collagen X, the only known hypertrophic chondrocyte-specific molecular marker.37,38 The other major problem in cartilage regeneration is the dedifferentiation of chondrocytes with fibroblast-like morphology, which usually occurs during in vitro expansion.39–41 This process is accompanied by a down-regulation of collagen type II expression and an significant up-regulation of collagen type I expression.42,43
Various bioactive factors such as TGF-β superfamily and IGF-1 have been shown to be critical for the induction of chondrogenic differentiation of MSCs, with some studies showing that synergistic use of TGF-β1 and IGF-1 did not result in widespread improvement of MSC chondrogenesis, both in vitro 44 and in vivo, 45 compared with single growth factor. In our previous study, cationic chitosan in the scaffold could simultaneously serve as a gene carrier to express and maintain TGF-β1 protein, which was necessary for chondrocyte proliferation. 25 Our results reported here further supported that chitosan/gelatin scaffold as TGF-β1 gene delivery system could maintain a chondrocyte-like phenotype when using MSCs for cartilage reparation, in terms of enhanced aggrecan and collagen over time. In addition, in pTGF-β1-activated scaffolds, mRNA levels of collagen type II and aggrecan were remarkably up-regulated, while mRNA expression of collagen type I, usually suggested as an osteogenetic marker during the limb development, was consistently low with progress toward a cartilaginous phenotype. It seems reasonable to speculate that gene-activated scaffolds are convenient and efficient for the maintenance of a chondrocyte-like phenotype of MSCs.
It is well known that DNA itself (especially DNA containing CpG sequences) can activate cells, including MSCs. Therefore, a control group using empty plasmid DNA is necessary. We used the same plasmid vector containing EGFP sequence as the control plasmid group in cell proliferation, ELISA, and Q-PCR assays. The results showed that the MSC proliferation was really activated by empty plasmid, about 25% higher than DNA-free group, as well as a little increase in RNA and protein expression. But it is still lower than that in the pTGF-β1-activated group.
In the gross assessment of cartilage repair, it appears that the similar repair results between the DNA-free group and the pTGF-β1-activated group after 10 weeks postsurgery, which raised the controversial focus whether MSCs were enough to repair the defects or TGF-β1 plasmid was needed for cartilage repair in vivo. In fact, the interior composites of defects were significantly different between the two groups. The real repaired degrees were evaluated by cartilage-specific ECM in defect area, which could only be detected by histological and collagen II immunohistochemical staining—the most popular and classic methods to evaluate the quality of repaired cartilages at present time.30,46–48 Although there are some other techniques for evaluating the repairing cartilage, such as micro-NMR and mechanical test, the well-recognized method is still histological and immunohistochemical assessment.49,50 In this study, in the pTGF-β1-activated group, the neo-surfaces of repaired tissues were firm to touch and stained with much denser collagen II, while those in DNA-free group were rather softer even though they seemed to be apparently filled, and relatively weak collagen II staining was detected. It is to be noted that although collagen II staining in Figure 8Q is also strong, it is obvious that in pTGF-β1 group cartilage-specific matrix is more homogenous, which means higher stability of repaired tissue. Accordingly, compared to DNA-free group, higher scores were observed in the pTGF-β1-activated group using modified O’ Driscoll grading score system. It is noteworthy that, after 10 weeks, newly formed cartilage tissues performed a quite exciting integration with adjacent host cartilage in the pTGF-β1-activated group. This is very important because any of micro-motions and frictions between repaired and host tissues may induce further cartilage degeneration.
Conclusion
In summary, our results demonstrate that MSC-based pTGF-β1-gene-activated scaffolds are beneficial for in vitro and in vivo cartilage tissue engineering and have potential for further clinical application of cartilage regeneration.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (50673041 and 30771036), Jiangsu Provincial Natural Science Foundation (BK2007144), the National Basic Research Foundation of China (973 Program 2006CB503909 and 2006CB503908), and the 111 project from the Chinese Ministry of Education.
Disclosure Statement
No competing financial interests exist.