
Abstract
Aim:
The aim of this study was to analyze and compare the deposition of cartilage-specific extracellular matrix components and cellular organization in scaffold-free neocartilage produced in microgravity and simulated microgravity.
Methods:
Porcine chondrocytes were seeded (100 × 106/mL) into cylindrical culture chambers (n = 8) and cultured in the following environments: (i) microgravity during the Flight 7S (Cervantes mission) on the International Space Station (ISS), (ii) simulated microgravity in a random positioning machine (RPM), and (iii) normal gravity (1 g, control). After 16 days, each neocartilage tissue was processed for histology, immunohistochemistry, quantitative real-time reverse transcriptase–polymerase chain reaction, and histomorphometric analysis.
Results:
Weaker extracellular matrix staining of ISS neocartilage tissue was noted compared with both Earth-cultivated tissues. Higher collagen II/I expression ratios were observed in ISS samples compared with control tissue. Conversely, higher aggrecan/versican gene expression profiles were seen in control 1 g samples compared with microgravity samples. Cell density produced in microgravity was significantly reduced compared with the normal gravity neocartilage tissues.
Conclusion:
Tissue cultivated on the RPM showed intermediate characteristics compared with ISS and 1 g conditions. These data indicate that the RPM system does not sustain microgravity. Although microgravity impacts the development of in vitro generated cartilage, simulated microgravity using the RPM may be a useful tool to produce cartilaginous tissue grafts with fewer cells.
Introduction
The RWV has been shown to create a microgravity environment with low shear stress and a number of studies have shown it suitable for the production of cartilaginous tissues from isolated chondrocytes.13–17 More recently, Ohyabu et al. 18 used rabbit bone marrow stromal cells in an RWV to produce cartilage tissue. Conza et al. 19 showed that neocartilage cultivated on an RPM exhibits a much more regular distribution of cells within a well-developed cartilage extracellular matrix (ECM) than the layered structures produced under normal conditions without a supporting scaffold. Despite these positive reports, microgravity can inhibit the early stages of cartilage formation and does not induce the formation of neocartilage tissue comparable to tissues produced in normal gravity.
Initiation of chondrogenesis during limb bud formation and in in vitro culture systems rely on cell-to-cell and cell-ECM interactions, which are mediated through a number of surface molecules and their corresponding transduction pathways.20–22 Chondrogenesis is retarded or reduced when cells are placed in microgravity environments before cell-to-cell contacts are established.23,24 However, if condensation is allowed to occur prior to microgravity exposure, the subsequent extent of chondrogenesis is not compromised, although the amount of matrix produced is reduced 5 compared with 1 g controls.
Despite the negative aspects related to chondrogenesis in microgravity, the increased cell spacing described by Conza et al. 19 has direct implications for the production of cartilage grafts with cell distributions closer to mature tissue and with potentially fewer starting cells. The latter feature is a common conundrum in modern cell-based therapies in which cell expansion in culture is a prerequisite to obtain sufficient cell numbers for therapeutic purposes. More importantly, chondrocyte expansion in monolayer leads to changes in cell differentiation status25,26 resulting in reduced chondrogenic capacity of dedifferentiated cells and hence production of hyaline-like neocartilaginous tissue.27,28
The aim of this study was to compare the neotissue cultured in normal gravity (1 g) with tissue cultured in (i) simulated microgravity as induced by the RPM and in (ii) an actual microgravity environment. For the latter environment, bioreactor hardware was designed suitable to be transported on the International Space Station (ISS) during a 10-day flight, which functioned as an “in-flight” incubator. To the best of our knowledge this was the first time that a biological space flight experiment was run in parallel to an RPM reference experiment from the same tissue source. The direct comparison is important to further validate whether cells exposed to simulated microgravity respond in a similar way to real microgravity conditions. The importance of such a comparison is necessary to validate whether Earth-based simulated microgravity systems are truly comparable to microgravity in space; the latter represents an expensive option.
Materials and Methods
Cell isolation and culture chambers
Full-thickness porcine articular cartilage was harvested from the hip joint of a slaughter aged male pig (165 days old and 75 kg weight) and the cells were released via enzymatic digestion. The minced cartilage pieces were exposed to pronase for 2 h (1 mg/mL, 10 mL/g of tissue; Roche, Basel, Switzerland), washed in phosphate-buffered saline (PBS), and subsequently incubated overnight in collagenase (0.25 mg/mL, 10 mL/g of tissue; Roche). The cells were counted, directly seeded into nine bioreactor chambers, as previously described, 29 at a density of 100 × 106 cells/mL per chamber, and placed into culture chambers (CCs) containing medium (Dulbecco's modified Eagle's medium/Nutrient Mix F12 (Life Technologies, Carlsbad, CA) with the additives: 1.125 g/L NaHCO3, 10% fetal calf serum, 1 μg/mL insulin, 50 μg/mL vitamin C, and 100 μg/mL penicillin/streptomycin (Gibco BRL (former), now Invitrogen, Basel, Switzerland). Briefly, the bioreactor chambers consist of a tube with an upper and lower membrane to enclose the seeded cells inside the CC and to allow nutrient exchange (Fig. 1).

Experimental design. Porcine articular cartilage was collected and the chondrocytes were released via enzymatic digestion. The isolated cells were seeded into nine CCs. Phase I: All nine CCs were put on the RPM at 25°C for 36 h. Phase II: Three were randomly selected and transported to the launch site in Baikonur, Kazakhstan. The other six CCs were kept in an incubator in Zurich at 25°C and randomly shaken to simulate transport. Phase III: At 4.5 days after seeding, two out of the three CCs taken to Baikonur were placed into microgravity (
Normal and microgravity environments
Porcine chondrocytes within the CCs were exposed to three different gravity environments (Fig. 1):
ISS, microgravity RPM, simulated microgravity Normal gravity (1 g)
Following chamber seeding, all nine CCs were placed on the RPM for 36 h at 25°C. The RPM simulates microgravity conditions and was developed by T. Hoson in Japan and manufactured by Dutch Space (former Fokker Space). The machine is composed of an inner and an outer frame with a platform in the middle to accommodate the samples (Fig. 1B). The frames rotate independently at a randomly varying speed (0.1–2 radians/s). The whole device was maintained inside a room (at the specified temperatures throughout the experiment) in normal atmospheric CO2 conditions and without humidification to simulate the conditions on the ISS. The nine CCs were subsequently removed from the RPM and three were randomly chosen and transferred to a transportation container (portable incubator; Fig. 1C) for transport from Zurich, Switzerland, to the launch site at the Baikonur Cosmodrome, Kazakhstan (3 days before launch), and kept at 25°C during transportation. Meanwhile, during transport, the other six CCs were maintained in Zurich in an incubator at 25°C. The remaining six CCs are considered to be the reference CCs. To partially simulate transport disturbances the reference CCs were turned every 8 h and lightly shaken at irregular intervals. Upon arrival in Baikonur the medium was changed. The medium in the reference CCs was changed at the same time. Two of the three Baikonur CCs were randomly selected (the third CC was disposed) and integrated into the specially designed flight incubator (Fig. 1C) with a regulated temperature of 36°C. In parallel, the six reference CCs were also placed in an incubator at the same temperature in Zurich. At L-18 (18 h before the launch) the experiment was handed over to Russian technicians who installed the flight incubator in the Soyuz capsule TMA-3. At the time of launch, three of the reference CCs were mounted on the RPM and the remaining three were kept in the incubator at 36°C. After 48 h in orbit the Soyuz spacecraft docked to the ISS and 3 h later the experiment was transferred from the Soyuz to the ISS, where the battery pack was replaced. After 8 days in orbit on the ISS the experiment was disconnected from its power supply and returned to Earth in the Soyuz capsule TMA-2 after 6 h. Twelve hours after landing, the neocartilage tissue was transferred to a transport vessel filled with PBS and the samples were transported back to Zurich at ambient temperature (25°C). (At the same time neocartilage tissue from the three RPM CCs and the three 1 g CCs was transferred into vessels filled with PBS and kept at 25°C). Twenty-four hours later, all eight samples (three 1 g, three RPM, and two microgravity samples) were transported to Bern, Switzerland, for analysis. We suggested that the cells cultured on the ISS should have been fixed for histology and/or frozen for RNA isolation. In our case this was not possible either on ISS or on ground because of European Space Agency (ESA) handling and transport restrictions.
Hardware specifications
The experiment hardware was designed to match the specifications and safety standards of both ISS and Soyuz manned missions. Additionally, the following physical specifications had to be fulfilled: maximum size: 125 mm × 110 mm × 110 mm, maximum weight: 600 g. This led to the development of the cell cultivation system as shown in Figure 1C. The cell cultivation system consisted of three layers of containment, the two cell CCs, containment 2, containment 3, the outer aluminium cylinder, and a temperature regulation system. The temperature regulation system consisted of a heating foil surrounding the two CCs, and a thermal controller and a thermostat as a safety measure to cut the power as soon as the surface temperature would reach 40°C. As additional safety measure a heat fuse set to 70°C was attached to the heat foil. The CC, containment 2, and containment 3 were each tested to withstand inside overpressure of 1.1, 1.65, and 3 bar, respectively.
Evaluation of neocartilage
Histology and immunohistochemistry
All samples were cut in half. One set from each treatment was fixed for 24 h with 4% paraformaldehyde, embedded in paraffin, and cut into 4 μm thick sections. Histological staining included hematoxylin eosin for an overview of tissue structure and cell distributions, Alcian blue and safranin-O–Fast Green for glycosaminoglycan deposition, and Masson's trichrome for collagen produced. Immunohistochemistry was performed to assess deposition of collagen types I and II. Sections were pretreated with 1 mg/mL chondroitinase and then incubated with monoclonal primary antibodies for collagen type I (Quartett, Berlin, Germany) or collagen type II (II-II6B3; Hybridoma Bank, University of Iowa, Iowa City, IA) for 1 h. The slides were subsequently incubated with a biotinylated goat anti-mouse secondary antibody for 35 min, followed by StrABC/AP for 45 min (Dako, Glostrup, Denmark). Bound antibodies were revealed via incubation with New Fuchsin/Naphthol AS-BI substrate (Sigma Chemical, Buchs, Switzerland). Fetal growth plates were used as positive controls.
Quantitative real-time reverse transcriptase–polymerase chain reaction
The second half of all samples was used to determine the expression levels of cartilage specific genes. Total RNA was isolated from each sample and from freshly isolated porcine chondrocytes for reference. The RNA obtained with Qiagen RNeasy kit (Qiagen, Basel, Switzerland) was converted to cDNA using random hexamers (Promega, Dübendorf, Switzerland) and Omniscript reverse transcriptase (Qiagen). Gene expression levels of collagen type I, collagen type II, aggrecan, and versican were determined with quantitative real-time reverse transcriptase–polymerase chain reaction (PCR) using the ABI Prism 7700 Sequence Detection System (Perkin Elmer/Applied Biosystems, Rotkreuz, Switzerland). The PCR master mix (Perkin Elmer/Applied Biosystems) was based on AmpliTaq Gold DNA polymerase (Perkin Elmer/Applied Biosystems). Each cDNA sample (2.5 μL in a total volume of 25 μL per reaction) was analyzed in duplicate. The gene of interest and the reference gene (18S ribosomal RNA) were labeled with 6-carboxyfluorescein (Microsynth, Balgach, Switzerland). Sequences and final concentrations used for human 18S, collagen types I, II, aggrecan, and versican primers and probes were as previously published.30,31
For each cDNA sample, the threshold cycle (Ct) value of each target sequence was subtracted from the Ct value of the reference gene to obtain the ΔCt. The level of expression of each target gene was then calculated as 2ΔCt. The efficiencies of primers/probe amplification for the chosen genes have been previously established.25,30,32,33
Histomorphometric analysis
Histomorphometric analysis was conducted to assess the effect of different gravity conditions on cell distribution within each neotissue. To this end, custom Matlab software was used to determine the number of cells per standard field area of six hematoxylin and eosin-stained slide images (40 × magnification) per condition.
Statistical analysis
To compare cell densities between each treatment condition, a T-test was employed (Microsoft Excel). p-Values lower than 0.05 were considered significant.
Results
All neocartilage samples were soft, especially the tissue produced on the ISS (it could not be handled with forceps). Neocartilage formed in normal gravity was continuous in shape and form that corresponded to the shape of the cylindrical bioreactor, compared to ISS and RPM tissues, which consisted of irregular cell aggregates. The discontinuous matrix was particularly pronounced in ISS cultures (Fig. 2). Deposition of proteoglycans, as illustrated by safranin-O–Fast Green (Fig. 2A–C) and Alcian blue (Fig. 2D–F) staining of neocartilage cultivated on the ISS was weaker compared to both RMP and normal 1 g cultivated tissues. Very weak staining intensity of Masson's trichrome was observed in all samples (data not shown).

Histological evaluation of immature neocartilage produced on the ISS, on Earth in a RPM, or in normal gravity conditions (1 g control). Alcian blue staining (
Collagen type II immunostaining (Fig. 3A) revealed substantial collagen deposition in all conditions, albeit visually weaker in ISS cultivated tissue, confirming the other histological observations. Immunostaining for collagen type I was extremely weak to negative in all constructs examined (Fig. 3A).
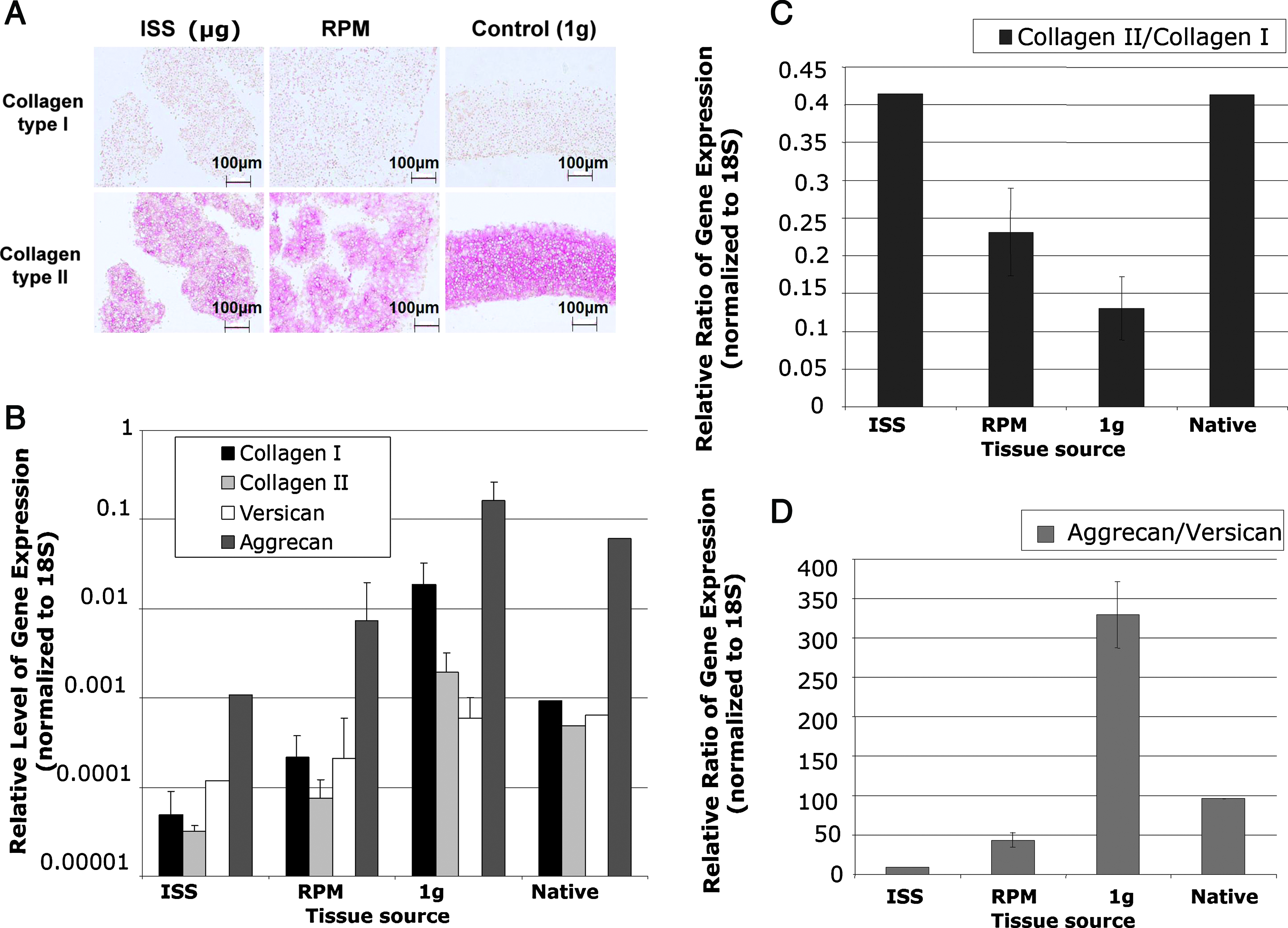
Detection of cartilage-specific products at the protein and gene expression levels. Collagen type I and type II deposition in neocartilaginous tissues cultured under different gravity conditions was assessed using immunohistochemistry (scale length: 100 μm). (
Quantitative real-time reverse transcriptase–PCR of individual gene expression levels (normalized to 18S ribosomal RNA) shows a clear effect of microgravity for all genes examined (Fig. 3B). Gene expression level ratios indicate that ISS cultivated neotissue had substantially higher collagen type II/type I compared to the other treatments and comparable to normal cartilage (Fig. 3C). An inverse expression profile of aggrecan/versican ratio is evident in all neocartilage (Fig. 3D) compared to that depicted for collagen. The highest aggrecan/versican expression ratios were observed under normal gravity conditions (1 g) and the lowest in the ISS neotissues. RPM tissue showed intermediate aggrecan/versican expression ratios compared to the other treatment groups.
The microgravity conditions created either on the RPM or on the ISS significantly decreased (p < 0.05) the cellular density and correspondingly increased cell spacing compared to control 1 g tissues (Fig. 4), with the most striking reduction in cell density evident in RPM tissues. Average cell densities for the ISS sections were calculated at 5434.6 ± 62.7 cells/cm2 and 4707.3 ± 106.8 cells/cm2 in the RPM sections. Control (1 g) sections contained 5911.0 ± 100.3 cells/cm2.
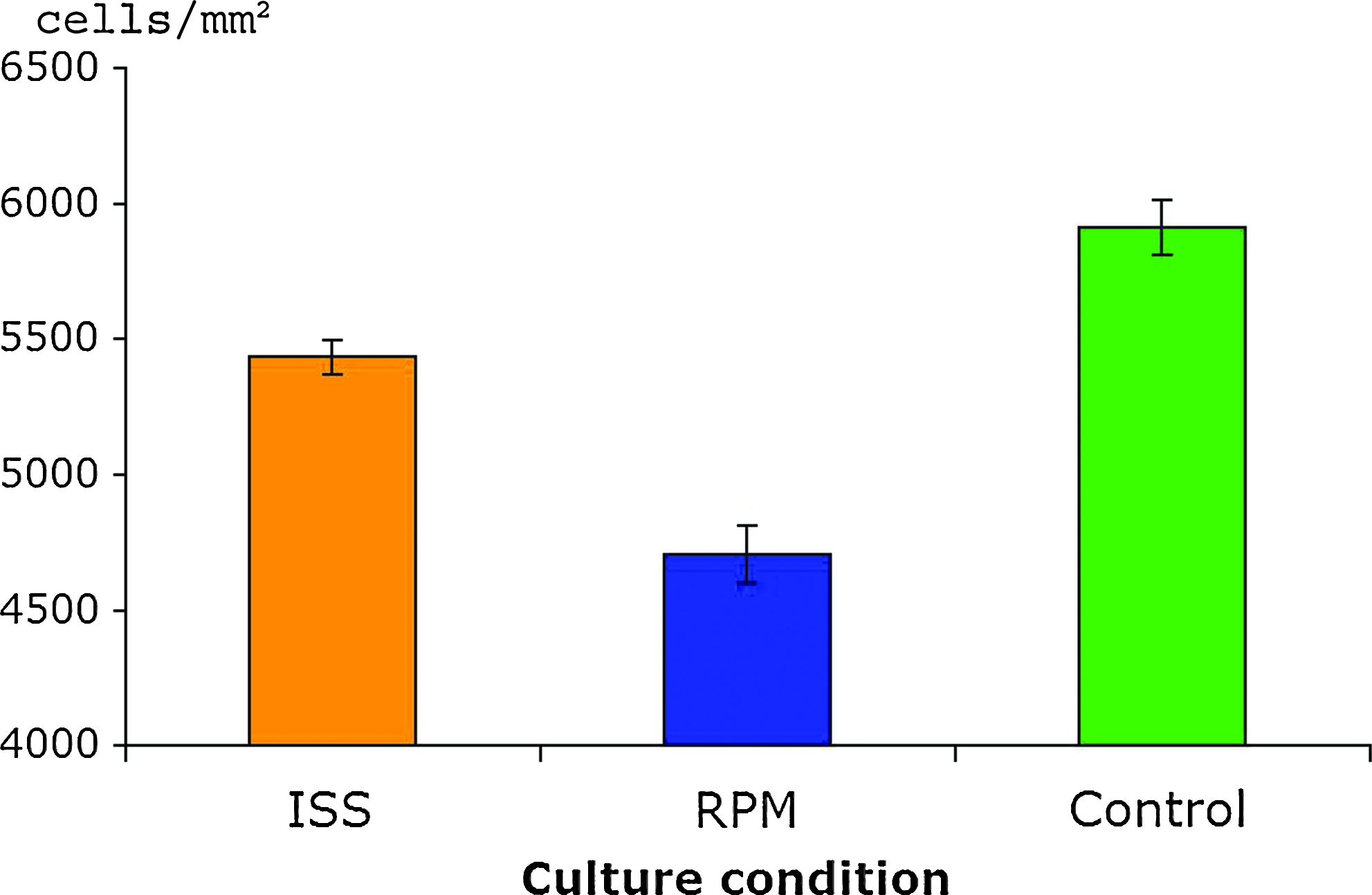
Histomorphometry and cell distribution. Cell density was assessed in neocartilaginous tissues produced under different gravity conditions employing custom Matlab software and expressed as mean number of cells per mm2. p-Values lower than 0.05 were considered statistically significant. Color images available online at www.liebertonline.com/ten.
Discussion
This study further substantiates that microgravity impacts the development of de novo cartilage as previously reported.5,13,23,34 Importantly, we have been able to directly compare neocartilage formation in 1 g, microgravity, and simulated microgravity from the same cell source. Cultivation of porcine chondrocytes on the ISS led to less ECM deposition compared to neocartilage tissue produced in either the RPM or in normal gravity, as judged by matrix staining intensity and by the higher aggrecan to versican ratios. Tissue cultivated on the RPM generally showed intermediate characteristics compared with ISS and 1 g conditions. These data indicate that the RPM system is not equivalent to real microgravity. Nevertheless, RPM and ISS morphological features and cell spacing were similar.
Mechanical pressure loading studies on chondrocytes and mesenchymal stem cells have shown increased gene expression and protein production of ECM, including aggrecan and collagen type II.35–37 ECM gene expression is likely regulated by gravitational cues translated through mechanotransduction pathways. 38 In the current study the normal 1 g neotissue represents a loaded condition when compared with both the microgravity and the simulated microgravity environments. The reduced aggrecan/versican individual expression and their ratios in microgravity constructs are consistent with previous observations.39,40 Yasuoka et al. 40 showed reduced aggrecan expression in the nucleus pulposus and annulus fibrosus of rats subjected to tail suspension.
Strikingly, ISS tissue displayed higher collagen type II to type I expression ratios compared with normal gravity control tissue, despite not translating to higher protein levels as observed in the immunohistochemistry images. The latter observation may be explained by the reduction in individual gene expression levels of collagen type I and type II in microgravity and RPM compared with normal gravity. Saito et al. 41 provided evidence that gravitational load may affect the posttranslational modification of collagen, particularly the collagen maturation pathway through altered activities of enzymes involved in crosslink formation. Beekman et al. 42 has shown that impaired crosslinking of collagen interferes with collagen deposition. Thus, the overexpression of collagen RNA in ISS samples, seen in this study, perhaps did not translate to protein or could be related to similar posttranslational modifications. It is also important to consider that the cells used in this study were freshly digested and seeded directly into the bioreactors (CCs). Therefore, from the start of chamber culture, these cells were not in a dedifferentiated state. A markedly higher expression ratio of collagen type II to type I was also seen in the RPM samples compared with the 1 g controls. One other explanation for the differences in collagen transcription could be the response of the ISS cultivated tissue upon return to 1 g conditions. Using microarrays on rats either subjected to normal or microgravity, Taylor et al. 43 noted changes in gene expression profiles that appeared to be due to a “readaptation” to the normal gravity environment. Montufar-Solis et al. 44 also reported alterations in rat growth plate characteristics upon reloading after microgravity exposure. Harvesting of RNA after having fixed or frozen the test samples while still in zero gravity would resolve this issue. 44 In our case this was not possible, neither on the ISS nor on ground due to ESA and National Aeronautics and Space Administration (NASA) handling and transport restrictions. An important message of this paper is for others to consider these variables when conducting these types of experiments. Additionally the use of 1 g centrifuge controls in microgravity would be desirable, 34 and in this study, this was not possible because of strict weight and size limitations.
The extent of chondrogenesis is reduced in microgravity environments, especially if cell-to-cell contacts necessary to initial condensation cannot take place. 23 However, if condensation is allowed to occur prior to microgravity exposure, the subsequent extent of chondrogenesis is not compromised, although the amount of matrix produced is lower. This notion is largely consistent with our present observations. In all treatments in this study, the cells seeded into chambers were permitted cell-to-cell contact during 3 days (transport phase), which is likely sufficient for condensation to occur. It has been hypothesized that cells in the clinostats are subjected to constant deformation that prevents normal response, which does not provide the correct cues for continued cell differentiation and further ECM production. 5
RPM-cultivated chondrocytes created tissue with matrix composition characteristics in between the control 1 g and the ISS-produced neocartilages, but produced neocartilage with more evenly dispersed and further spaced apart cells. This characteristic may be a useful means of spacing chondrocytes closer to native tissue without the requirement of a scaffold, and moreover, to reduce the need for a large number of cells. The number of cells that can be obtained from a clinically relevant sized biopsy is not sufficient for cell-based therapies such as autologous cartilage implantation or to produce a functional neocartilage implant. Thus, the classic expansion phase in monolayer is needed to increase cell numbers, adding to the probability of reduced chondrogenic capacities. 28 This point is especially a factor in scaffold-free methods that rely on the volume of cells to act as their own scaffold support and tissue volume. 29 The RPM therefore offers both opportunities: a reduction in the amount of cells and the prefabrication of implants without the complications associated with scaffolds. The biomechanical features of RPM tissues are questionable, yet use of mechanical loading after RPM culture may be a suitable strategy to compensate this deficiency. A possible system whereby engineered cartilage may be produced utilizing the microgravity conditions created in the RPM coupled with a mechanical loading regime to produce engineered grafts with less cells is outlined in Figure 5.
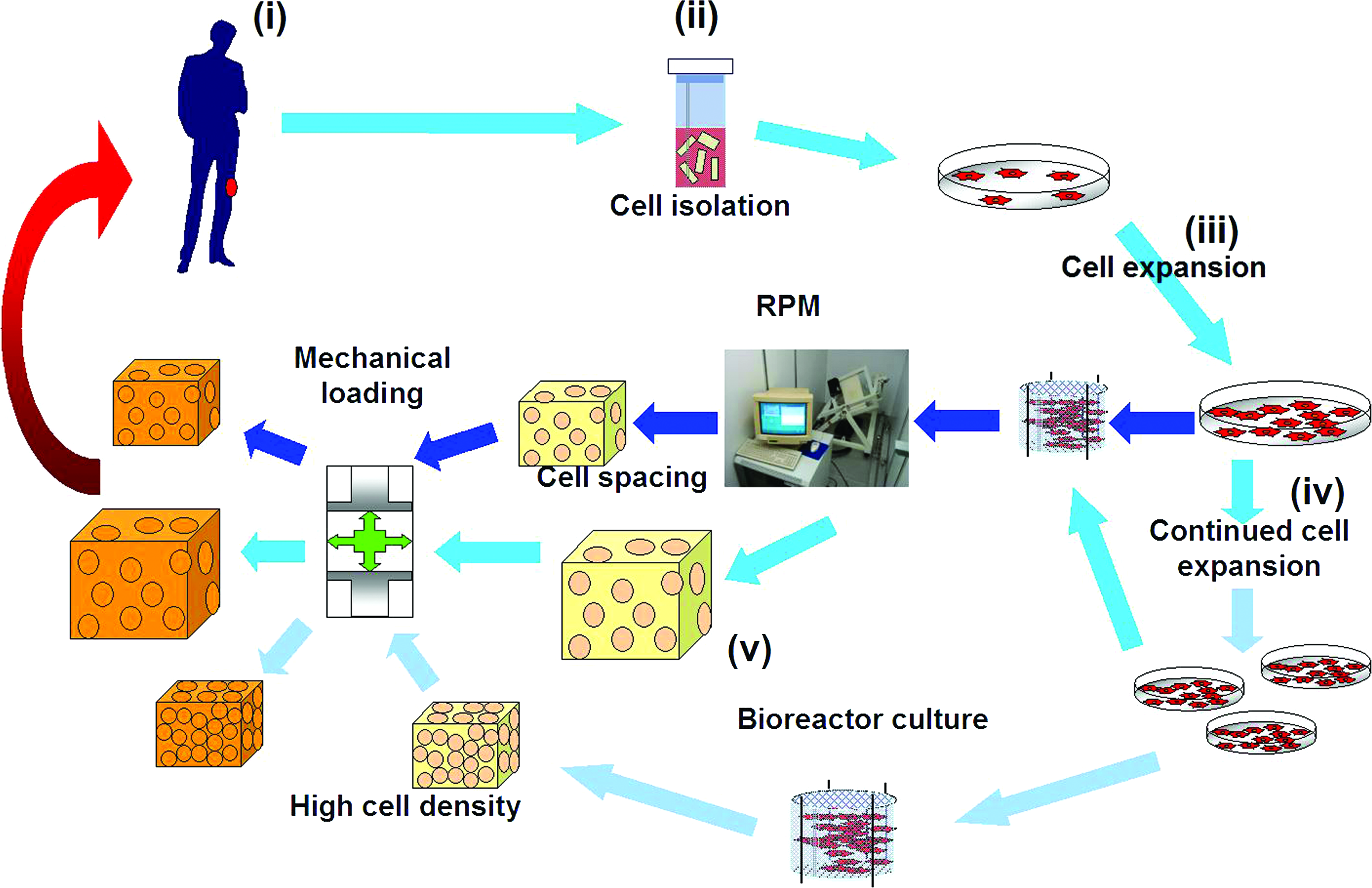
Possible use of the RPM to engineer cartilage implants without scaffolds. Cell-based cartilage repair strategies consist of collecting a biopsy from the donor (i), isolating the cells from the tissue (ii), and expanding the small quantities of cells in monolayer cultures (iii) until sufficient numbers are obtained for implant manufacture in bioreactor systems (iv). The RPM may be employed to produce neotissue with fewer cells because the simulated microgravity spaces the cells further apart. Moreover, if cells are expanded further in culture (v), the RPM may also allow for the production of larger cartilaginous grafts that may be suitable to heal larger defects. Mechanical loading would also contribute to the formation of quality neocartilage tissue for grafting. Color images available online at www.liebertonline.com/ten.
It is important to emphasize that the sample size is very small (eight samples: two microgravity, three RPM, three 1 g); therefore, our conclusions are not based on a sound statistical analysis. But chances to have experiments in microgravity and experimental mass are extremely limited because of the enormous costs involved. The preparation time for our experiment was 2 years following the ESA and NASA regulations and high restrictions. Nevertheless, our results do give an insight into possible connections between different gravitational states. The main goal of this experiment was to provide direct comparison among three different gravity environments from the same cartilage source.
Conclusions
To the best of our knowledge this was the first time that a biological space flight experiment was run in parallel to an RPM reference experiment from the same tissue source. Microgravity leads to the formation of nonuniform neocartilage tissue with reduced ECM and altered gene expression in a manner suggesting dependence on gravity for correct translation to protein. Future studies using the RPM may be useful to elucidate this phenomenon. ISS and RPM tissues possessed similar yet not identical characteristics, as measured by gene expression and cell spacing. Better controls, for instance, freezing/fixing neotissue in microgravity and an onboard 1 g centrifuge, are needed to directly compare these systems. Further exploration as to whether the RPM system could be used as part of a tissue engineering production system may be warranted.
Footnotes
Acknowledgments
The authors are grateful for the support and help of the whole team of the Space Biology Institute of the Swiss Federal Institute of Technology (ETH) Zurich. The authors thank the whole ESA Outreach Office for providing this unique flight opportunity, and the High Energy Physics Institute of the ETH Zurich for building and designing most of the hardware. Their special thanks goes out to Rogier Schonenborg who assisted greatly with the technical development of the hardware and Carmen Adusumalli who was responsible for making all this possible. They are thankful for the expertise of Commander Alexander Kaleri (Russian Cosmonaut), Dr. Michael Foale (NASA Science Officer), and Dr. Pedro Duque (European Astronaut). They are also grateful to Ms. Verena Winkelmann and Ms. Chantal Pauli for processing the histology and the support of Dr. Pierre Mainil-Varlet (Institute of Pathology, University of Bern). The collagen type II antibody (II-II6B3) was obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the National Institute of Child Health and Human Development (NICHD) and maintained by the University of Iowa, Department of Biological Sciences, Iowa City, IA. This research was funded by ESA, ETH Zurich, Centerpulse, and the Holcim Stiftung.
Disclosure Statement
No competing financial interests exist for the following authors: Vlada Stamenković, Georg Keller, Dobrila Nesic, and Shawn Patrick Grogan. Although no competing financial interests exist for Professor Augusto Cogoli, it should be disclosed that he is a cofounder and employee of Zero-g Life Tec.
Work performed at affiliations 1 and 2 and the Russian cosmodrome Baikonur and onboard the International Space Station (ISS).