
Abstract
The nanoscale spatial organization of collagen fibrils as major constituents of extracellular matrices is believed to be crucial for neurite guidance in neural development and repair. To systematically study the influence of collagen fibril alignment, length, and density on human neuronal cell behavior, we used our novel technology to produce aligned collagen matrices by shear flow deposition using a microfluidic channel system and applied these surfaces to functional human neurons and glia derived from white matter neural stem cell cultures. Neurites on aligned collagen were highly oriented in the direction of the underlying fibrils, whereas neurites on nonaligned collagen or poly-
Introduction
Synthetic tubular nerve guides were designed to replace the need for nerve autografts.6,7 Nerve guidance channels/tubes provide global direction to regenerating axons and prevent their escape into neighboring tissue; however, they do not ensure local direction of axons and cells within the tube. Hence, so-called guidance channels or fibers aligned along the longitudinal axis within the tube were introduced to improve axon regeneration. 8 In this context, extracellular matrix (ECM) components are of great importance, mainly collagen, laminin, and fibronectin, as well as matrix-bound trophic factors.9,10 Collagen type I is of special interest because it has been shown to favor neuronal rather than glial cell growth.3,5 Transplantation of a collagen matrix into the transected cortico-spinal tract of adult rats induced neuronal fiber ingrowth while inhibiting invasion of reactive astrocytes or microglial cells, which are thought to cause glial scar formation. 11 In a self-assembly process, the triple-helical collagen I forms fibrils 20–70 nm in diameter, which in turn associate into fibers and fiber bundles.12,13 In many tissues, collagen fibrils/fibers are organized in parallel arrays. In peripheral nerves, for instance, the endoneurium surrounding the axons with Schwann cell sheets is predominately composed of orientated collagen fibers. 3 Oriented collagen fibrils aligned within gels by magnetic fields in vitro are known to increase neurite extensions in comparison to randomly oriented collagen fibrils. 8 In addition, aligned electrospun fibers or grooves profoundly affect the direction and guidance of cells and their processes.14–18 It is now clear from a growing body of evidence that neurite growth strongly depends on substrate topography, including dimension and density of guidance cues.19–21
Although it is straightforward to prepare randomly oriented collagen networks in vitro, creating aligned structures is more challenging. Several groups have introduced methods to align collagen matrices in vitro22–28 ; however, these do not allow for systematic adjustment of fibril alignment, density, and morphology. Therefore, we have recently developed a new method using a microfluidic system to create well-aligned native collagen fibril matrices of variable density and morphology. 29 To assess the potential of these aligned collagen fibrils as bioartificial scaffolds for biomedical applications in humans, we studied the behavior of primary human adult neuronal cells cultured on aligned collagen with differing fibril alignment, length, and density. As primary human neurons are not available for obvious reasons, we used adult human neural stem cells (hNSCs) as a source for functional human neurons and glia. In detailed analyses of adult hNSCs from various brain regions including subcortical white matter, it has been recently shown that these adult hNSCs behave similar to their murine counterparts both during expansion and differentiation and differentiate into functional nerve cells with similar neurite outgrowth.30–33 We show that aligned collagen matrices with tunable structural characteristics have a great potential to guide and enhance neurite outgrowth.
Materials and Methods
Cell culture
Adult human subcortical white matter tissue was obtained from routine epilepsy surgery (anterior temporal lobectomy or resection of focal cortical dysplasias; six samples; individuals 28–49 years old; five women and one man) following informed consent of the patients. All procedures were in accordance with the Helsinki convention and approved by the ethical committee of the Dresden University of Technology (EK 47032006). Specimens were taken from the removed tissue and stored in ice-cold Hank's balanced salt solution supplemented with 11 mM glucose and 1% penicillin/streptomycin while being transported from the operating theater. All patients underwent high-resolution magnetic resonance imaging to exclude tumors and were screened for the presence of infectious disease. In all cases, tissue and neuropathological examination did not reveal evidence of tumor formation. For expansion of neurospheres consisting of hNSCs, white matter was isolated from tissue samples and then cut into small pieces with a scalpel, incubated in 0.1% trypsin (Sigma Aldrich, St. Louis, MO) for 30 min at room temperature, incubated in DNase (40 mg/mL; Sigma, Steinheim, Germany) for 10 min at 37°C, and homogenized to a quasi single-cell suspension by gentle triturating.30,31,34 The cells were added to 25-cm2 flasks (2–3 × 106 viable cells per flask) in knockout Dulbecco's modified Eagle medium (DMEM) (Gibco BRL–Life Technologies, Tulsa, OK), supplemented with 10% serum replacement (Gibco), 0.5 mM glutamine, 1% penicillin/streptomycin, and 20 ng/mL of both epidermal growth factor (EGF) and fibroblast growth factor-2 (both from Sigma) at 5% CO2, 92% N2, and 3% O2 using an incubator equipped with an O2-sensitive electrode system (Heraeus, Hanau, Germany). After 10–20 days, neurosphere formation was observed, and these spheres were expanded for additional 5–8 weeks (in total, 7–12 weeks) before differentiation was initiated. The medium was changed once a week and the growth factors were added twice a week.
Differentiation conditions
Induction of neural differentiation was initiated by plating the cells on poly-
Preparation of glass substrates
For experiments with glass substrates coated with PDL and poly(octadecene-alt-maleic acid) (POMA; Polysciences, Warrington, PA), freshly cleaned 22 × 22 mm glass coverslips (Menzel Gläser, Braunschweig, Germany) were oxidized in a 1:1 mixture of aqueous ammonia solution (Acros Organics, Geel, Belgium) and hydrogen peroxide (Merck). PDL solution was pipetted onto the glass substrate, kept at room temperature for 30 min, and rinsed with phosphate-buffered saline (PBS). For POMA coating, coverslips were functionalized by reaction with 3-aminopropyl-dimethylethoxy-silane (ABCR, Karlsruhe, Germany). POMA was dissolved in tetrahydrofuran (Fluka, Deisenhofen, Germany) in a concentration of 0.08 wt% and spin coated (RC 5; Suess Microtec, Garching, Germany) onto the amine-modified coverslips at 4000 rpm for 30 s. Stable covalent binding of the polymer films was achieved by annealing at 120°C to form imide bonds with the amino-silane on the glass substrate.
Microfluidic system and flow conditions
Aligned collagen matrices were prepared by a novel approach using a microfluidic setup (for further information about the microfluidic system, please refer to Refs.29,35). The flow conditions were performed under conditions of laminar flow. In all experiments, the Reynolds number (as a criterion for laminar flow) was smaller than 1, and a flow rate of 11 μL/min corresponding to a wall shear rate of 203.1 s−1 was applied.
Collagen solution
Bovine dermal collagen I solution (purified and pepsin-solubilized in 0.012 N HCl; PureCol; Inamed, Fremont, CA) was brought to physiological pH by mixing eight parts of acidic collagen solution (3.0 mg/mL) with one part of 10-fold concentrated PBS (Sigma) and one part 0.1 M NaOH. All components were kept in an ice bath before and after mixing. Appropriate volumes of chilled 1× PBS were added to adjust the final concentration of the collagen solution.
Nonaligned collagen matrices
POMA-coated glass slides were coated by contacting the substrates to the collagen solution and then subsequently raising the temperature. 36 To precisely apply collagen coating in designated areas, a silicone-based culture insert (Ibidi, Martinsried, Germany) consisting of two wells was placed onto a POMA-coated glass slide. A 100 μL drop of chilled nonfibrillar collagen solution (0.4 mg/mL) was pipetted into each well and kept in a CO2-free incubator at 37°C to initiate fibril formation. After 90 min the resulting fibrillar collagen gel and the culture insert was removed from the surface, leaving two 0.22-cm2 rectangular areas covered with a thin collagen layer on the substrate. The modified surface was rinsed with PBS for at least three times and subsequently with Milli-Q water (Millipore, Molsheim, France).
Aligned collagen matrices
To produce collagen matrices containing long fibrils at a low density, a low-concentration collagen solution (3 μg/mL) was used as described previously. 29 Briefly, collagen solution (0.8 mg/mL) was prepared at 4°C in a centrifuge tube (Roth, Karlsruhe, Germany), placed in a CO2-free incubator (Sanyo, Bensenville, IL) at 37°C, and allowed to form fibrils for 24 h. The resulting gel was homogenized for 4 min (T-8 Ultra Turrax Tube Drive from IRA, Staufen, Germany) and centrifuged (Heraeus) at 1000 g for 6 min. The supernatant (2 mL, 3 μg/mL) was drawn into a syringe that was subsequently installed into the syringe pump. The solution was then pumped for 1 h with a flow rate of 11 μL/min through the microfluidic channel. Afterward, the channel was rinsed with PBS for 20 min to flush away loose fibrils, and the coated coverslip was removed from the poly(dimethylsilotane) (PDMS) channel plate and kept in PBS for cell culture. Aligned collagen matrices containing short fibrils at low density were prepared with a solution concentration of 0.2 mg/mL. 29 Briefly, the microfluidic system was placed in a CO2-free incubator (Sanyo) at 37°C. The collagen solution was prepared at 4°C and drawn into a chilled syringe (Roth), which was quickly installed into the syringe pump to avoid early fibrillogenesis. The streaming process was started immediately by pumping the chilled collagen solution from the cooled storage reservoir through heated tubing, followed by streaming through the microfluidic channel for 1 h. The sample was then rinsed with PBS as described earlier. Aligned collagen matrices containing long fibrils at high density were prepared according to the same protocol for collagen matrices containing short fibrils at a low density using a solution concentration of 0.8 mg/mL. Samples were sterilized under UV light for 5 min.
Confocal reflection microscopy
Confocal reflection microscopy (CRM; TCS SP; Leica, Bensheim, Germany) analysis was performed at a wavelength of 488 nm (Ar laser) to visualize unstained collagen fibrils as described elsewhere. 37 CRM pictures of fibril matrices were obtained using a 40× oil-immersion objective.
Atomic force microscopy
Surface topography of air-dried collagen matrices was investigated via intermittent contact scanning force microscopy with a PicoSPM (Molecular Imaging, Phoenix, AZ) using silicon cantilevers (Tap300; BudgetSensors, Sofia, Bulgaria) with a 300 ± 100 kHz resonant frequency, 40 N/m force constant, and tip radius <10 nm.
Scanning electron microscopy
Samples for scanning electron microscopy (SEM) analysis were fixed with 2% glutaraldehyde, dehydrated, and critical point dried (CPD 030; Baltec, Schalksmühle, Germany). The samples were gold coated with a sputter coater (SCD 050; Baltec) to allow sample analysis under high vacuum by SEM (XL 30 ESEM FEG; FEI-Phillips, Eindhoven, The Netherlands) equipped with a scanning electron detector and operating at 7.5–10 kV, Spot Size 3. Micrographs of hNSCs on the area covered with aligned collagen fibrils or the corresponding area (5 mm × 1 mm) on the controls were taken. For each condition (long or short fibrils at high or low density), three independent donors were analyzed.
Image analysis to quantify collagen fibril alignment
The quantification of fibril alignment was performed as described previously. 29 Briefly, CRM images of three samples containing two channels with collagen fibrils were analyzed for each condition and quantified using the NIH ImageJ 1.37v software. One sample contained a minimum of 200 collagen fibrils. A threshold value of the reflection intensity was defined to isolate fibrils from background. By fitting an ellipse to the major axis of each collagen fibril, the angle of the fibrils to the flow direction was determined. The orientation of fibrils parallel to the alignment direction corresponded to an angle of 0°. The angular distribution of collagen fibrils was determined based on the relative frequency of orientation angles (classified into bins of 10° angles) and by a fit to a Gaussian full width at half maximum (FWHM).
Image analysis to quantify hNSC extension alignment and length
SEM images were analyzed to quantify the extent of alignment and length of cell extensions. The quantification of extension alignment was performed as described for quantification of collagen fibril alignment. The angular distribution of extensions was determined based on the relative frequency of extension angles (classified into bins of 10° angles) of three donors. Only cells/extensions on the area covered with aligned collagen fibrils or the corresponding area (5 mm × 1 mm) on the controls were measured. The length of each extension extending from neurospheres was measured using NIH ImageJ 1.37v (plugin NeuronJ) software. Each extension was measured from its origin at the edge of the neurosphere to its end. Changes in direction along its course were taken into account by tracing along the entire length of each extension. Extensions that were solitary and clearly isolated were measured only to exclude the possibility of mix-up with other extensions.
Immunocytochemistry
Cell cultures were fixed with Accustain (Sigma). Immunocytochemistry was carried out using standard protocols described previously.30,38 Cell nuclei were counterstained with 4,6-diamidino-2-phenylindole (DAPI). The following primary antibodies were used: mouse anti-GalC 1:750 (Chemicon International, Temecula, CA), rabbit anti-Tuj1 1:2000, mouse anti-Tuj1 1:500 (both from Covance, Richmond, CA), chicken anti-glial fibrillary acidic protein (GFAP) 1:1000 (Abcam, Cambridge, UK), rabbit antiactivated caspase3 1 μg/mL (BD Pharmingen, San Diego, CA), and secondary antibodies conjugated to Alexa 488, 568, or 647 1:500 (all from Invitrogen–Molecular Probes, Carlsbad, CA). Images were captured using a fluorescence microscope (Leica DM IRE2; Leica Microsystems, Wetzlar, Germany). Living cells were marked for analysis using fluorescein diacetate (FDA) and dead cells were stained with propidium iodide (PI; Sigma Aldrich). A solution of 100 μg/mL FDA was prepared by dissolving in acetone. A working stock solution containing 100 ng/mL of FDA was prepared in PBS and was added directly to the cell cultures (i.e., no change of culture medium) at a 1:1 volume ratio, and PI was added at a concentration of 1:50 (1 μg/mL). Samples were incubated for ∼5 min and visualized using epifluorescent microscopy (Leica DM IRE2; Leica Microsystems) with excitation filters at 490/20 nm and emission at 527/30 nm for fluorescein. As neither PI nor activated caspase3 staining worked on the hNSCs on the substrates, dead cells were quantified by counting condensed DNA using DAPI staining correlated to all nuclei.
Cell counting and statistics
To quantify the percentage of cells producing a given marker in a given experiment, the number of positive cells on the coated well surface was determined relative to the total number of DAPI-labeled nuclei. In a typical experiment, a total of 500–1000 cells were counted per marker. Statistical comparisons were made by ANOVA with post hoc t-test and Bonferroni adjustment or Dunnett's t-test as appropriate. All data are presented as mean values ± standard error of the mean. All quantifications were done by an independent blinded investigator.
Results
Alignment of collagen fibrils
As part of neuronal tissue regeneration in both central and peripheral nervous tissue, targeted axodendritic outgrowth is a major challenge for reconstructing neuronal tracts and circuitries. We thus tested the ability of aligned collagen fibrils generated using a microfluidic system 29 to guide neurites during the differentiation process of adult white matter hNSCs (see Fig. 1 for schematic representation of the technology). Collagen fibrils were deposited on planar POMA-coated substrates from collagen solutions streaming through a microfluidic channel. By varying collagen solution concentrations and gelation times, fibril length and density were tuned and varied. In brief, lower concentrations generated shorter fibrils at lower density, whereas higher concentrations resulted in aligned matrices containing long fibrils at high density. Atomic force microscopy was used to observe the resultant aligned collagen matrices: long fibrils at high density (Fig. 1A), long fibrils at low density (Fig. 1B), and short collagen fibrils at low density (Fig. 1C).
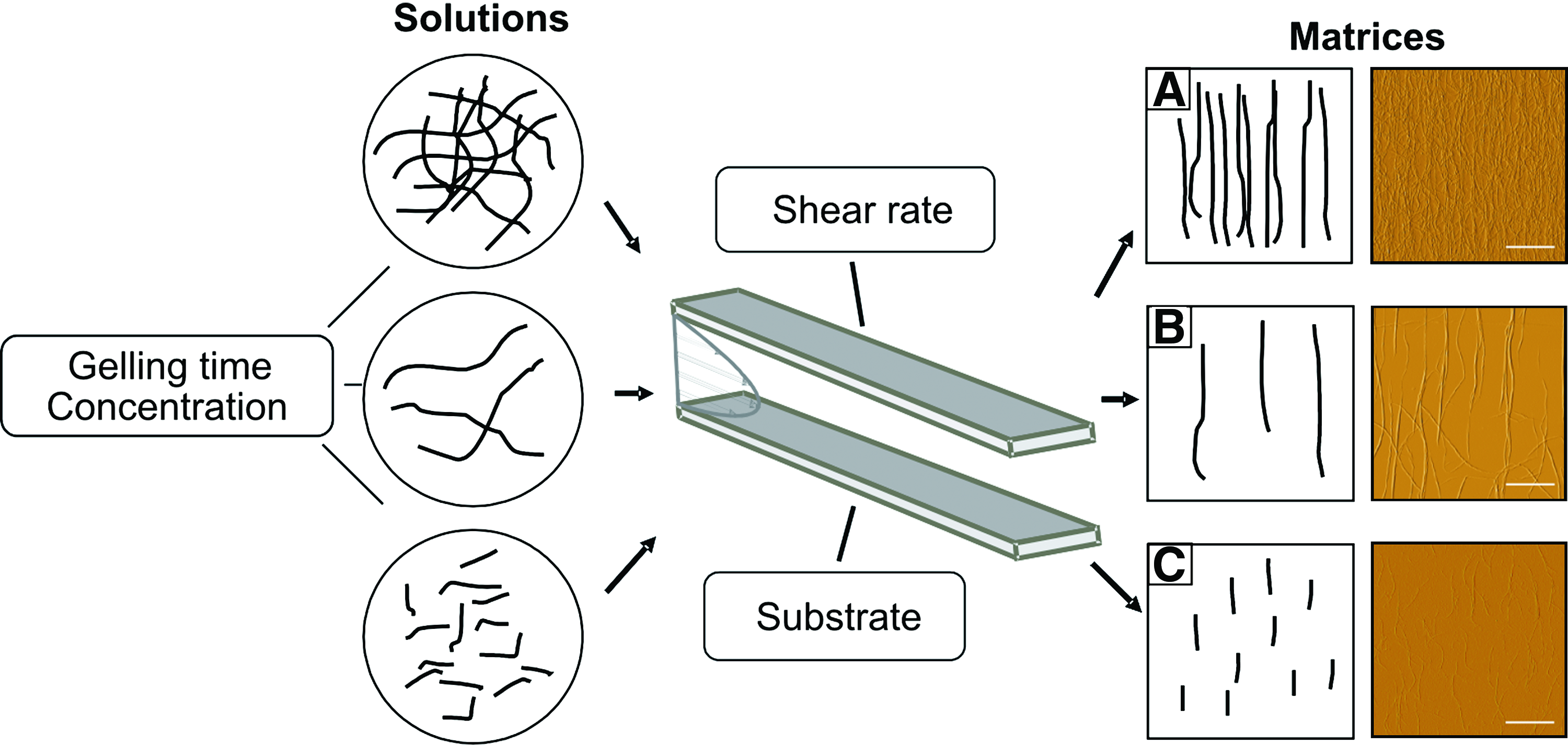
Scheme of the approach to prepare collagen type I fibril matrices that are adjustable in orientation, density, and morphology by varying collagen solution concentration, gelling time, shear rate, and substrate. Atomic force microscopy images show the resultant aligned collagen matrices with long fibrils at high density (
Influence of aligned collagen with varying structural characteristics on extension alignment
To investigate the significance of the newly developed alignment approach for biomedical application, we attempted to prove its potential in a human nerve cell system. As human neurons are not routinely available without harming the patient or the cells themselves, we used neurons differentiated from adult hNSCs. Adult hNSCs were isolated from cortical white matter tissue obtained from neurosurgery interventions as recently described.30,32 Triple immunofluorescence staining of neuroectodermal markers confirmed typical trilineage neuroectodermal differentiation capacity of the adult hNSCs with 30% ± 5% Tuj1+ neurons, 10% ± 4% GFAP+ astrocytes, and 4% ± 2% GalC+ oligodendrocytes after 7 days of differentiation on the standard differentiation surface PDL (data from three independent experiments from three different donors). Adult hNSCs are thus a valuable human cell model for investigating axodendritic outgrowth of human Tuj1+ neurons. We used cells from at least three different donors for all further experiments.
To assess the effect of aligned collagen matrices with varying structural properties on neurite outgrowth and extension alignment (Fig. 1), hNSCs were differentiated for 7 days on rectangular areas (1 mm × 6.5 mm) with aligned collagen fibrils (long fibrils at high density, Fig. 2A; long fibrils at low density, Fig. 2B; short fibrils at low density, Fig. 2C) deposited on a POMA-coated glass surface. Nonaligned collagen resulting from static fibril formation and immobilization on a POMA-coated surface served as the control surface (Fig. 2D). SEM and immunocytochemistry were used to analyze hNSC morphology. Fibril and cell alignment was quantified by determining the angular distribution of collagen fibrils and cell extensions. Based on this analysis, the orientation angle distribution was plotted as a histogram, and the corresponding FWHM was determined by a Gaussian fit (see also Lanfer et al. 29 ). Fibril alignment was highest on matrices containing long fibrils at low density (FWHM, 18° ± 1°; Fig. 2B, F), followed by long fibrils at high density (FWHM, 30° ± 3°; Fig. 2A, E), and short fibrils at low density (FWHM, 41° ± 5°; Fig. 2C, G). Nonaligned collagen showed random distribution of fiber angles (Fig. 2D, H). On all substrates, differentiated hNSCs displayed typical unipolar, bipolar, and multipolar neuronal morphologies with axodendritic outgrowth (Fig. 2I–L). We measured the orientation of a minimum of 166 extensions per independent experiment. On substrates containing aligned fibrils (Fig. 2I–K), aligned extension outgrowth in the direction of the underlying collagen fibrils was observed, whereas extensions on nonaligned collagen exhibited no preferred orientation (Fig. 2L). In contrast to fibril alignment degree, extension alignment was highest on long collagen fibrils at high density (FWHM, 38° ± 3°; Fig. 2I, M), then successively decreased on long fibrils at low density (FWHM, 50° ± 19°; Fig. 2J, N) and short fibrils at low density (FWHM, 53° ± 11°; Fig. 2K, O). No preferred extension alignment direction was observed on nonaligned collagen (Fig. 2L, P). These findings emphasize the importance of both fibril alignment and fibril density for the orientation of axodendritic outgrowth of neurons derived from hNSCs. hNSC differentiation was confirmed using immunofluorescence staining for the neuronal marker Tuj1 and the astroglial marker GFAP. On all substrates, hNSCs acquired morphologic and phenotypic characteristics of Tuj1+ neurons and GFAP+ astrocytes (Fig. 2Q–T).

Influence of fibril alignment, length, and density on extension alignment of adult white matter-derived hNSCs. (
Directional outgrowth of hNSC extensions on aligned collagen matrices
Quantification of cell alignment revealed that the highest extension alignment occurs on matrices containing long collagen fibrils at high density. More detailed SEM studies were performed on these substrates to investigate extension morphology and cell–matrix interactions. Extensions on aligned collagen matrices containing long fibrils at high density were highly oriented, and extension alignment was consistent along the whole width and length of the collagen-coated area (Fig. 3A–C, E, dotted lines mark the collagen matrix border). In contrast, on the adjacent POMA-coated glass surface, extensions showed no preferred direction of orientation, emanating in radial fashion (Fig. 3A, D). Some neurospheres adhered at the border of the aligned collagen layer, extending processes on both the collagen surface and the adjacent POMA-coated glass. While extensions in contact with the POMA surface grew without any bias in alignment, those on the side with aligned collagen followed the direction of the underlying fibrils (Fig. 3E). Once in contact with the fibrils, the processes extended parallel to the fibrils, always in the direction of the aligned collagen (Fig. 3F, including inset). Immunocytochemistry staining demonstrated that the alignment cues from collagen fibrils acted almost exclusively on Tuj1+ neurites, while GFAP+ astroglial extensions were rather uninfluenced by collagen fibril alignment (Fig. 3G–J).
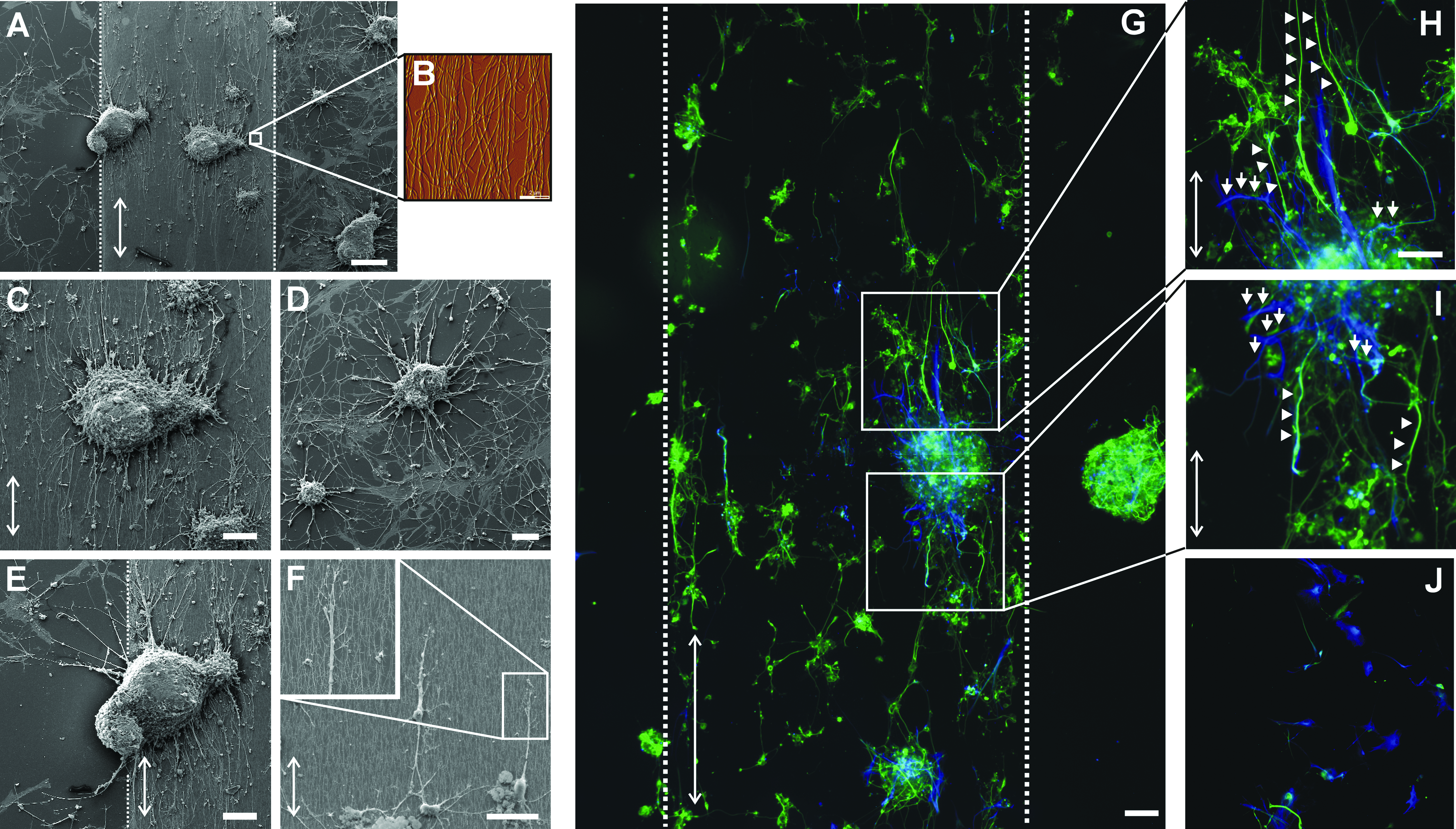
Aligned collagen fibrils guide and direct extensions of adult white matter-derived hNSCs. (
Influence of collagen fibril alignment on hNSC behavior
To further characterize the synergistic influence of topographical and biochemical cues from aligned collagen on hNSC behavior, we compared neuroectodermal differentiation on substrates with the highest cell alignment (long fibrils at high density) with nonaligned collagen and the standard differentiation surface PDL: Trilineage neural differentiation capacity was equivalent on nonaligned collagen and aligned collagen (long fibrils at high density), and no significant differences were observed when compared with PDL (p = 0.068, F-value = 4.06 for Tuj1+ cells; p = 0.90, F-value = 0.11 for GFAP+ cells; p = 0.37, F-value = 1.15 for GalC+ cells; ANOVA; n = 3; Fig. 4A, C). Interestingly, on POMA-coated surfaces, we found additional flat GFAP+ astrocytes that were not detectable by immunocytochemistry on PDL, nonaligned, or aligned collagen fibrils (Figs. 2 and 3). The influence of aligned structures on cell survival was quantified by counting condensed and inhomogeneous DNA using DAPI staining correlated to all nuclei. There were no differences in cell survival, with 5% ± 2%, 5% ± 2%, and 6% ± 1% pycnotic cell nuclei on PDL, nonaligned, and aligned collagen fibrils, respectively (p = 0.85, F-value = 0.17; ANOVA; n = 3; Fig. 4B, D). Thus both hNSC differentiation and survival showed no difference with respect to growth surface. This contrasts to the highly specific influence of aligned collagen type I on the direction of neurite outgrowth.
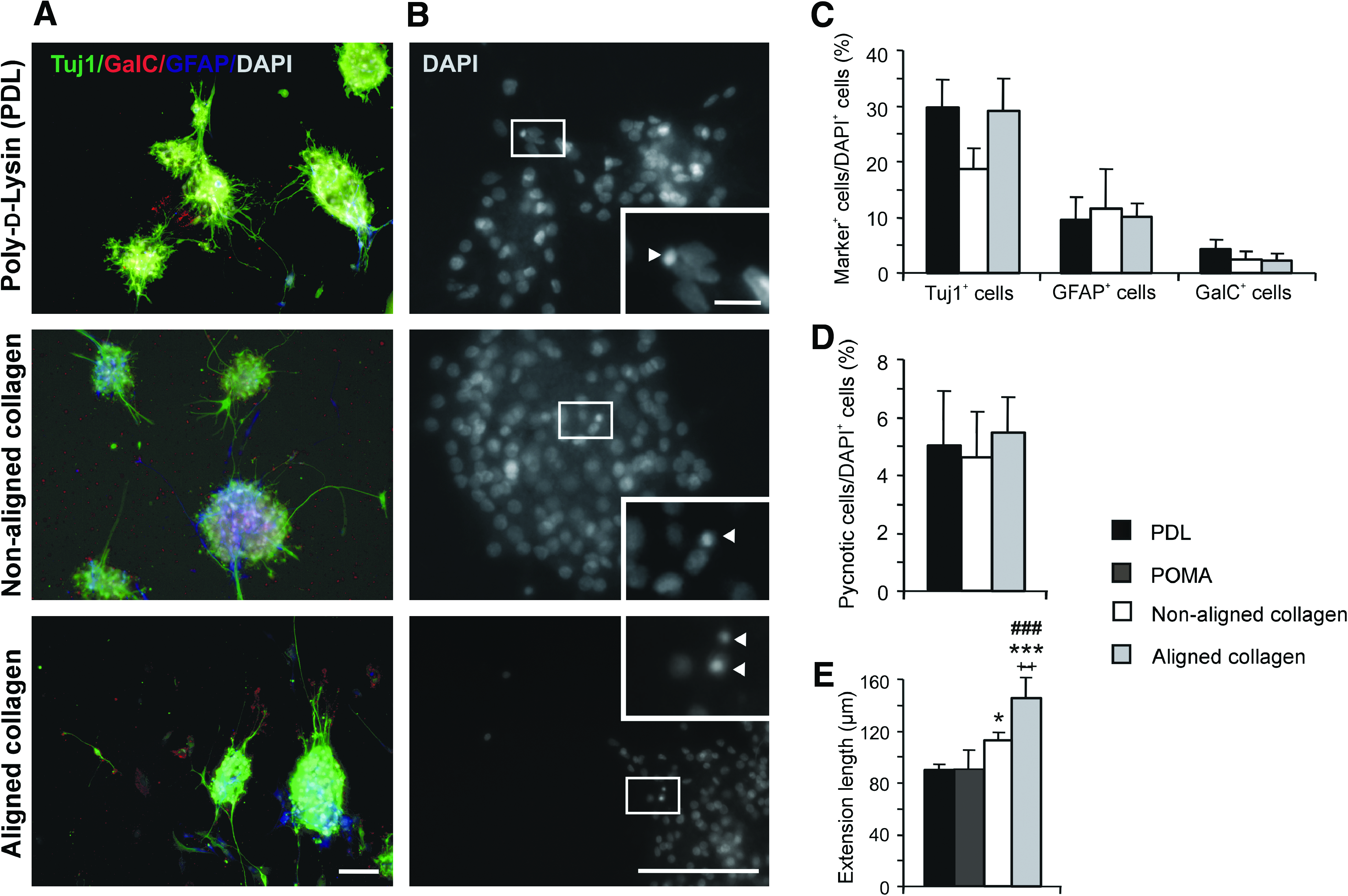
Influence of aligned collagen matrices on hNSC differentiation and survival. (
To determine the effect of fibril alignment on neurite outgrowth, extension length was quantified in SEM photographs by measuring each extension along its entire length from the neurosphere to its end. We measured 109–422 extensions per independent experiment. hNSC extension length significantly varied between seeding surfaces (p = 0.0014, F-value = 17.01; ANOVA; n = 3 independent experiments; Figs. 2 and 4E). Extensions on aligned collagen fibrils (long fibrils at high density) were significantly longer (145 ± 8 μm) compared with those on PDL (86 ± 8 μm; p = 0.0003; post hoc t-test), POMA (88 ± 4 μm; p = 0.0008; post hoc t-test), and nonaligned collagen (111 ± 3 μm; p = 0.0073; post hoc t-test). On nonaligned collagen, extension outgrowth was significantly higher compared with PDL surfaces (p = 0.029). No significant differences were observed between PDL and POMA substrates.
Discussion
Biomaterials are especially relevant for biomedical approaches that aid nervous system regeneration. Current interventions are hindered by insufficient or undirected axodendritic outgrowth, which leads to a lack of synaptic integration into the host brain circuitries.3,5 As we describe here, guidance cues provided by biomaterials may prove essential in repair of central and peripheral nervous system disease/damage. Even though axonal recovery after transection is occasionally observed, this is often hampered by the lack of guidance materials, leading to glial scar formation with subsequent loss of function. 3 As an example, in spinal cord regeneration, the major obstacles to overcome, beyond simply guiding axons in the right direction, are the local inhibitory molecules (e.g., Nogo) and glial scar formation resulting from invasion of reactive astrocytes or microglial cells.3,5 Peripheral nerve regeneration pathophysiologically differs from the recovery of central neurons, depending mostly on sufficient guidance cues to direct axonal regrowth to its target. We demonstrated for the first time that neurite outgrowth of adult human nerve cells derived from white-matter NSCs (hNSCs)3,30–32 can be directed by aligned collagen matrices of particular fibril length and density without affecting cell fate decisions or survival. The exact interplay of collagen fibril alignment, density, and length was found to be crucial, thus proving the importance of our recently developed shear flow deposition method to produce aligned collagen matrices of varying structural properties. 29
There is a general consensus that longitudinally aligned structures improve nerve regeneration and directed neuronal outgrowth. 3 Growth cones at neurite tips respond to aligned structures via contact guidance, resulting in axially directed neurite projections and thus improved structural alignment.39–41 Our aligned matrices indeed served as guidance cues to direct neurite outgrowth of hNSCs along aligned collagen fibrils (Figs. 2 and 3). This was especially true for matrices containing long fibrils at a high density (Fig. 2I, M). Although the best alignment of collagen fibers was seen using long fibrils at low density (Fig. 2A–H), the best neurite orientation was achieved on long fibrils at high density (Fig. 2I–P). It is likely that the close proximity of adjacent fibrils prevents neurites from migrating in all directions when reaching the end of a fibril. Thus, not only the alignment, but the interplay of aligned collagen fibrils of the right size together with the right density is crucial for axonal sprouting of hNSC-derived neuronal extensions. It has been demonstrated in several studies that dimension and size of substrate topography strongly influence neurite guidance and growth. Further, there is substantial evidence from studies on planar substrates using patterned surface chemistry and topography to suggest that feature sizes in the submicron range, in particular, affect cell behavior. Neurite lengths of C17.2 NSCs on nanofibers (700 nm) were significantly higher than on aligned microfibers (3.5 μm). 18 However, in contrast, it has been reported that neurites extending from chick cerebral neurons do not follow grooves of 130 nm width. 42 Xenopus laevis neurites grew parallel to grooves of 14 nm depth and 1 μm width, but hippocampal neurons grew perpendicular. 43 These examples emphasize the importance of nanoscale topography for guidance of neurites, but substrate topography has to be tuned depending on the cell type as well the species of origin. This might be due to individual growth cone characteristics that impact the remarkable abilities to detect directional cues. 44 These data clearly demonstrate that every bioartificial material applied in human medicine has to be tested in human-derived cell or tissue systems.
The second major effect of collagen alignment was enhanced neurite outgrowth, with longer neurites compared with nonaligned collagen or PDL substrates (Fig. 4E). Increased mean extension length on aligned substrates has also been observed for neurites from rat and chick dorsal root ganglia on aligned electrospun nanofibers.14,17 Nonoriented surfaces obviously slow down neurite growth, likely because of time spent deciding which path/cue to take.14,45 Thus, topographical cues seem to influence axon outgrowth, but additional biochemical cues, here collagen type I, might also affect the speed of axonal outgrowth. In recent studies, the incorporation of porous collagen beads in multichannel nerve guides enhanced neurite outgrowth and two-dimensional surfaces of collagen type I improved Schwann cell attachment and adhesion.46,47 It has also been shown that functionalization of aligned electrospun polycaprolactone fiber surfaces with collagen type I increases cell adherence, proliferation, and process extension.48–50 Together, these results suggest that synergistic effects of anisotropic distribution of biochemical and topographical/physical cues enable faster nerve regeneration by exploiting the differential response of growth cones to changes in structural and biochemical features. 51
Interestingly, hNSC differentiation was not affected by aligned or nonaligned collagen (Fig. 4A, C). It has been reported that neuronal differentiation of adult rat hippocampal progenitor cells cocultured with astrocytes is enhanced on aligned polystyrene microgrooves compared with substrates without a pattern. However, their differentiation is not affected when cultured on aligned or nonaligned substrates without astrocytes. 16 Neuronal differentiation of C17.2 NSCs occurs faster on aligned/nonaligned electrospun nanofibers (300 nm) than on aligned/nonaligned microfibers (1.5 μm). Fiber alignment, however, does not seem to influence differentiation. 18 Even though collagen was shown to inhibit glial cell invasion, 11 nonaligned and aligned collagen did not favor neuronal differentiation in our study. Collagen matrices have another major advantage for the use in neural repair: They allow axonal growth, but inhibit glial cell invasion, thereby abolishing glial scar formation. 11 Consistently, our immunocytochemistry data showed that collagen fibrils do not change glial cell differentiation of hNSCs (Fig. 4A, C). Finally, the immunocytochemistry studies proved that the guidance cues of the aligned collagen mostly influence the orientation of neurites rather than glial processes (Figs. 2 and 3).
The fibrils in the aligned collagen matrices we used closely resemble those found in nerve ECM structures. For example, the 20–100-nm-diameter fibrils in our matrices are comparable to the 30–110-nm-diameter oriented collagen fibers found in the endoneurium that surrounds axons and thereby guides neurite migration.3,52,53 Together, our results demonstrate that—by mimicking the size and fashion of endoneurium collagen alignment—the optimal orientation, density, and length of collagen fibrils is effective in guiding and promoting hNSC-derived neuronal extensions without hampering cell behavior and survival. The flexibility of our method to adapt collagen matrices to the nerve type of interest is considered a major prerequisite to mediate these effects. Although the data presented only show the in vitro application of aligned collagen matrices, all data were obtained using adult human neurons, providing a proof-of-principle for induction of adult human neurite outgrowth by engineered ECM. Adult nerve cells were chosen rather than fetal or embryonic stem cell-derived neurons to avoid problems posed by the high developmental potential of the fetal/embryonic cells. The use of adult human dorsal root ganglion and/or spinal cord neurons would be highly intriguing in our system; however, these cells are unattainable for obvious reasons. Future attempts are warranted to develop three-dimensional scaffolds to apply these aligned collagen matrices in vivo to reconstruct axonal pathways in the central or peripheral nervous system.
Footnotes
Acknowledgments
The authors thank Sylvia Kanzler for excellent technical assistance. The work was supported in part by the Deutsche Parkinson-Gesellschaft (to A.H.), by a GlaxoSmithKline fellowship (to A.H.), by the Deutsche Forschungsgemeinschaft through the DFG-Research Center for Regenerative Therapies Dresden (to A.H. and A.S.), and by the Bundesministerium für Bildung und Forschung (Verbundvorhaben “Tissue Engineering”; AZ 0312126 to A.S).
Disclosure Statement
No competing financial interests exist.