
Abstract
Apatite biomaterials have potential not only as cell vehicles for engineering bone tissue but also as regulators of calcium (Ca) concentration in situ for controlling osteoblast functions, for example, osteogenic differentiation and fate management of hematopoietic stem cells (HSCs). To design apatite materials having optimal chemical properties for the latter purpose, more detailed investigations into what effect Ca concentrations have on osteoblast functions is crucial. In this study, osteoblasts were cultured at different Ca concentrations, and the temporal alterations in osteogenic differentiation and HSC niche-related protein (angiopoietin-1, 2 [Ang1, 2]) expression were investigated. The different Ca concentrations (1.8–50 mmol/L) in the cell culture medium had no effect on the proliferation of osteoblasts, but did on the cell morphology. The higher Ca concentrations (<6 mmol/L) enhanced the mineralization as well as Ang1 expression. In addition, Ang1 expression in osteoblasts showed higher correlation with expression of connexin43, the major marker of cell–cell interactions, whereas Ang2 related to integrin β1, the major marker of cell–matrix interactions. Thus, the local Ca concentration regulates cell morphology through the cell–cell or cell–matrix interactions, leading to the alteration of Ang1 expression in osteoblasts. Since these changes triggered by Ca are concerned with the osteogenic differentiation or reproduction of HSCs niche microenvironment, the results obtained in this study might be useful for designing apatite materials with optimal chemical properties.
Introduction
On the other hand, osteoblasts in an endosteal microenvironment play an essential role in controlling the fate of hematopoietic stem cells (HSCs). For example, osteoblasts and marrow endothelial cells express chemokine (CXC motif) ligand 12 (CXCL12/SDF-1) so that they can localize human hematopoietic progenitor cells in the bone marrow.14–16 Histologic analysis of the distribution of N-cadherin-positive osteoblastic cells revealed a distribution pattern similar to that of long-term HSCs. 17 Angiopoietin-1 (Ang1) expressed by osteoblasts activated the Tie-2 receptor on the HSCs and promoted tight adhesion of HSCs to their niches. This adhesion resulted in the quiescence and survival of HSCs, leading to the maintenance and self-renewal of stem cells. 18 Thus, the control of osteoblast functions including protein expression relating to the interaction with HSCs would be an effective way of reproducing this HSC niche-like microenvironment in vitro. However, alterations in HSC niche-related protein expression in osteoblasts originating from Ca ion cues have not been well investigated.
On the basis of these considerations, osteoblasts were cultured with different Ca concentrations. The temporal alterations in osteogenic differentiation and HSC niche-related protein expression of osteoblasts were investigated in this study.
Materials and Methods
Cell culture
Osteoblast-like cells (MC3T3E1, passage 30–35; Riken) were cultured in an alpha-minimal essential medium (Wako Pure Chemical) supplemented with 10% fetal bovine serum (Sigma) and 100 U/mL penicillin and streptomycin (Nacalai Tesque), and maintained at 37°C in a humidified atmosphere containing 5% CO2. To investigate the effect of Ca ions on osteoblasts functions, a medium containing 1.8–50 mmol/L CaCl2 (pH 7.4) was used for the osteoblast culture.
Cell proliferation
Twenty thousand osteoblasts were seeded in 24-well nonpyrogenic polystyrene plates (Corning) with different concentrations of Ca ions, and the cell number in each condition was determined using the WST-8 cell proliferation assay (Dojindo). Morphological changes of the cultured cells were observed with a phase-contrast microscope (TE2000; Nikon).
Von Kossa staining
Cells in the well plates were incubated with 1% silver nitrate solution and placed under UV light for 20 min. After several changes in distilled water, unreacted silver was removed with 5% sodium thiosulfate for 5 min, and the cells were rinsed and kept in distilled water. An image of the black stain in the plate was obtained using a digital scanner (GT-9300UF; Epson), and transmitted light was measured with a spectrophotometer (MTP-32; Corona Electric).
Western blotting
Equal amounts of collected protein from the cultured cells with 6 or 50 mmol/L of Ca concentrations were separated through a polyacrylamide gel by electrophoresis, transferred to nitrocellulose, and blocked in Tris HCl buffer containing 3% bovine serum albumin and 0.1% tween 20 (TBS-T) for 1 h. Incubation with a primary antibody (1:1000 dilution) was carried out overnight in the blocking buffer for 1 h at room temperature or overnight at 4°C. After washing with TBS-T, the membrane was reacted with an ABC solution (Vector Laboratories). The bands were observed by enhanced chemiluminescence according to the manufacturer's (Amersham) instructions. Image analysis software (Image J, NIH) was used for the quantitative analysis. Antibodies used in this study were rabbit anti-Ang1, rabbit anti-Ang2 (Alpha Diagnostics International), mouse anti-connexin43, and mouse anti-integrin β1 (BD Biosciences).
Immunofluorescent staining
Immunofluorescent staining was carried out to observe expression of HSC niche-related proteins (i.e., Ang1 and Ang2) in osteoblasts. Cells cultured with different Ca concentrations (6 or 50 mmol/L) were fixed with 4% paraformaldehyde solution on day 19. Antigen was retrived with a pressure cooker at 120°C for 2 min in a citrate buffered solution (Dako). Nonspecific binding was blocked with Blocking One solution (Nakalai Tesque). A 1:500 dilution of rabbit polyclonal antibody against Ang1 or Ang2, followed by Alexa fluor488 goat anti-rabbit secondary antibody (1:400 dilution; Invitrogen), were applied for 30 min at room temperature. Fluorescent images were obtained using a fluorescent microscope (TE2000; Nikon). Ten images randomly selected from four different stainings were used for image analysis to obtain the quantitative data.
Statistical analysis
Quantitative tests were conducted in quadruplicate, and mean values with standard deviations were then calculated. Statistical analysis of data was accomplished by one-factor analysis of variance. Student's t-test was used for comparison at the 99% confidence interval.
Results
Cell proliferation, morphological change, and differentiation
The normal alpha-minimal essential medium used in this study contained 1.8 mmol/L of Ca. A previous study indicated that the Ca concentration in bone marrow increased to 40 mmol/L in the region of bone remodeling. 6 Therefore, concentrations of Ca within the range of 1.8–50 mmol/L were used in this study. The cell proliferation profiles at all Ca concentrations indicated no significant differences in proliferation by day 30. Cells reached confluence at day 11, and the number of cells was maintained until day 30 of the culture period (Fig. 1a). However, microscopic images of osteoblasts at day 19 revealed that some parts of cells had detached from the bottom of the plate, aggregated with one another, and formed island-like cell groups in the culture in the presence of 50 mmol/L Ca (Fig. 1b). With 14 mmol/L Ca, cells did not detach but some cells exhibited a rounded morphology in the culture (Fig. 1c). With 1.8 or 6 mmol/L Ca in the medium, cells still firmly attached to the bottom of the dish (Fig. 1d).

Cell proliferation rate was not altered by the calcium ion concentration (
In vitro mineralizations in osteoblasts culture with different Ca concentrations were evaluated by von Kossa staining. The results indicated that mineralization was highest at 6 mmol/L Ca in the medium, and was significantly higher at 6–50 mmol/L Ca compared to 1.8 mmol/L Ca in the medium at day 30 (p < 0.01; Fig. 2).

von Kossa staining of osteoblasts cultured in the medium to which different concentrations of Ca ions were added. Osteoblasts cultured with 6–50 mmol/L Ca ions showed enhanced mineralization compared with cells cultured in the normal medium. The asterisk indicates a significant difference with p < 0.01 (analysis of variance, ANOVA). Color images available online at www.liebertonline.com/ten.
Alterations in HSC niche-related protein expression in osteoblasts
Since the cells indicated highest mineralization at 6 mmol/L Ca and showed significant morphological changes at 50 mmol/L Ca, expression of HSC niche-related proteins, Ang1 and Ang2, under these Ca conditions was evaluated by Western blotting analysis and immunofluorescent staining. The results indicated that expression both Ang1 and Ang2 changes according to the culture period and Ca concentrations (Fig. 3). Expression of Ang1 substantially increased (20-fold) within 7 days of culture. However, that of Ang2 indicated relatively low (2.4-fold) increments in osteoblasts cultured with 6 mmol/L Ca. On the other hand, Ang1 expression in cells cultured with 50 mmol/L Ca gradually increased with increased culture period, but Ang2 expression decreased under these conditions. Immunofluorescent images indicated that no significant difference in the stained cell number was detected in cells stained with Ang1 or Ang2 when culturing with 6 mmol/L Ca. In contrast, cells expressing Ang1 were significantly higher than cells expressing Ang2 in the culture with 50 mmol/L Ca (Fig. 4).

Expression of angiopoietin (Ang) proteins in osteoblasts cultured in different concentrations of Ca. Data are mean ± standard deviation from four experiments measured in quadruplicate. The asterisk indicates a significant difference with p < 0.01 (ANOVA). (
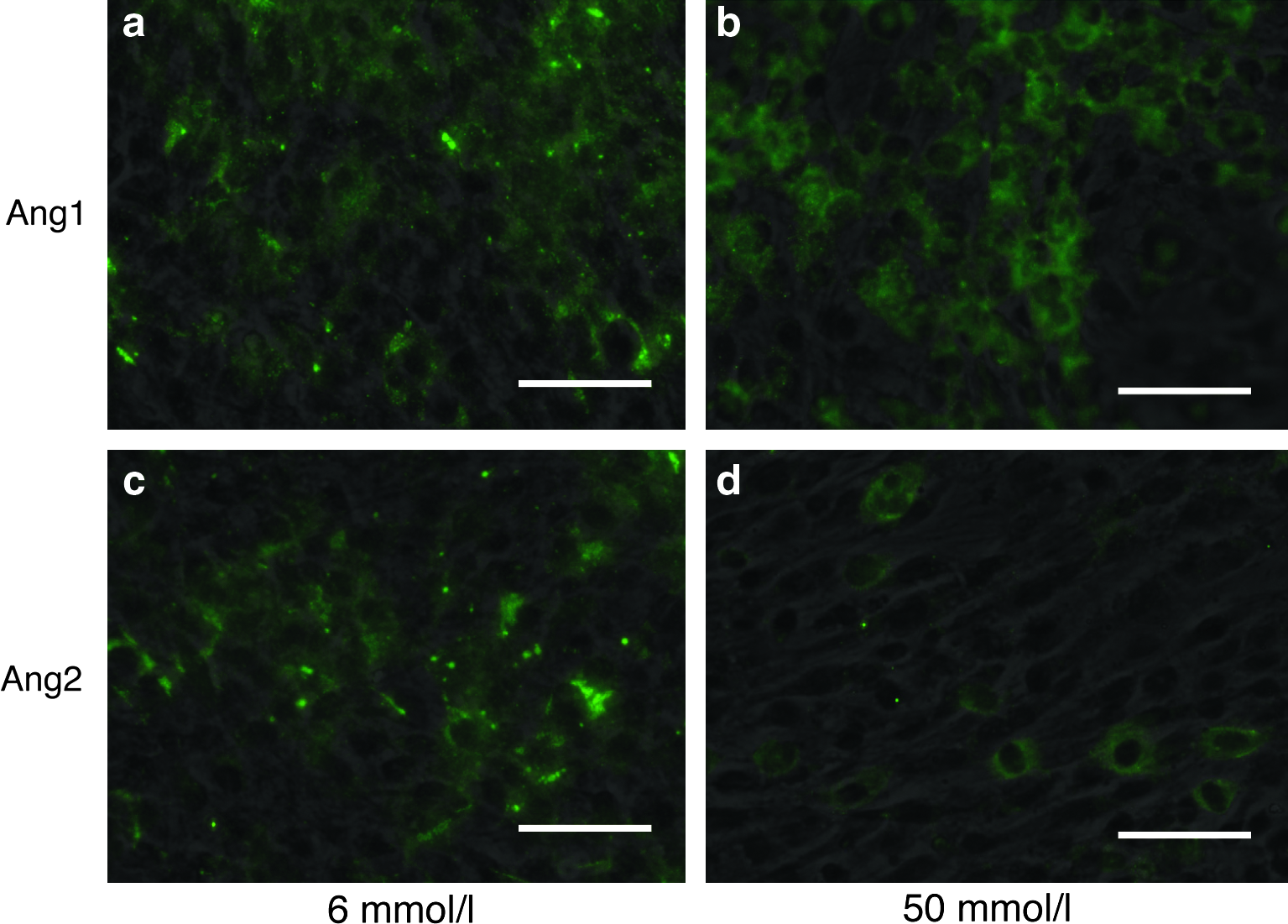
Immunofluorescent staining of Ang proteins in osteoblasts cultured with different Ca concentration at day 19. (Scale bar = 100 μm) (
Possible mechanism of changes in osteogenic differentiation and HSC niche-related protein expression in osteoblasts
Cell–matrix interactions are generally greater than cell–cell interactions during cell proliferation, and then cell–cell interactions gradually increase depending on the area occupied by cells in a two-dimensional culture dish. As seen in Figure 1, osteoblasts reached confluence at day 11 at all Ca concentrations. However, morphological changes in osteoblasts were observed according to Ca concentrations in the medium. The cell morphology is extremely regulated by cell–matrix and/or cell–cell interactions; therefore, these interactions might be related to the alterations in osteoblast functions evaluated in this study. To test and confirm this, expression of integrin β1, the major marker of cell–matrix interaction, and connexin43, the major marker of cell–cell interaction, in osteoblasts cultured at different Ca concentrations was evaluated. The results indicated that the protein expression profile of connexin43 was similar to that of Ang1, and that the profile of integrin β1 was similar to that of Ang2 (Fig. 5).
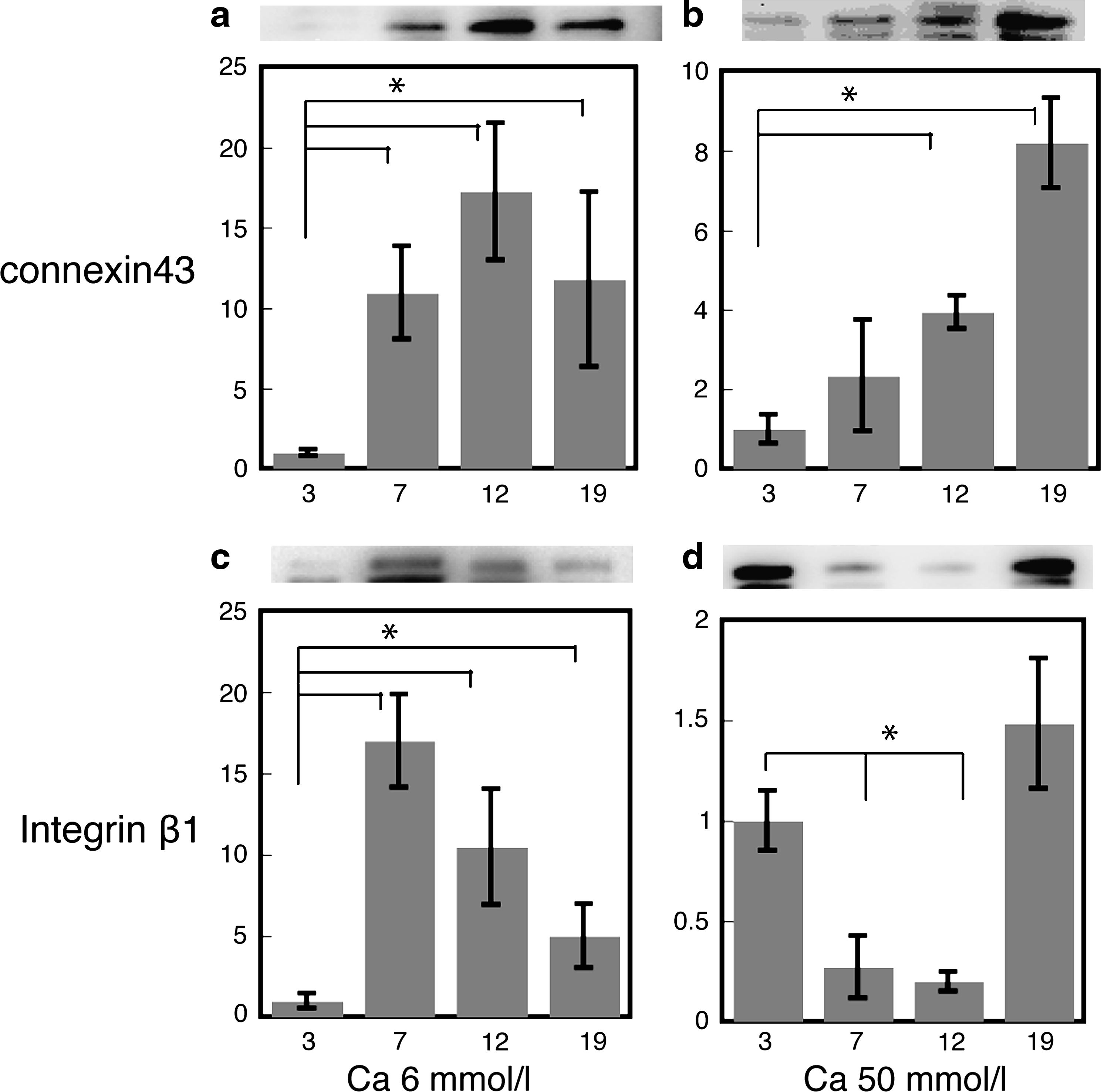
Expression of proteins (connexin43 and integrin β1) in osteoblasts cultured in different concentrations of Ca. Data are mean ± standard deviation from four experiments measured in quadruplicate. The asterisk indicates a significant difference with p < 0.01 (ANOVA). (
Discussion
Osteogenic differentiation and HSC niche-related protein expression of osteoblasts cultured at different Ca concentrations were evaluated in this study. The different Ca concentrations did not affect the proliferation of osteoblasts, but did affect the cell morphology during the long cell culture period. Consequently, Ca concentrations at the range of 6–14 mmol/L in the medium greatly enhanced mineralization. These results are supported by previous reports indicating that the addition of 6–15 mmol/L Ca to the medium promoted the osteogenic differentiation of osteoblasts.19,20 Thus, it was confirmed that the local Ca concentration was an important factor for the regulation of osteoblast functions, including cell adhesion and osteogenic differentiation.
Although the expression profiles of HSC niche-related proteins had specific patterning depending on the time (days) spent and the Ca concentration in the medium, a significant increase of Ang1 expression was totally detected in the cells cultured with the additional Ca ions. Ang1 and Ang2 are ligands of the Tie-2 receptor and are well known for their critical roles in angiogenic remodeling. Ang1 acts as an obligatory agonist promoting the structural integrity of blood vessels, whereas Ang2 functions as a naturally occurring antagonist promoting vessel growth.21–23 In addition to the angiogenic activity, recent studies indicated that Ang1 played crucial roles on osteogenesis. For example, in Ang1-transgenic mice, bone volume and bone parameters increased significantly compared with those of wild-type littermates. 24 Also, treatment or overexpression of Ang1 matrix complex in osteoblasts enhanced alkaline phosphatase activity, osteocalcin production, and mineral deposition. 25 Therefore, increased expression of Ang1 caused by the presence of Ca enhances the osteogenic mineralization in this study. Although higher expression of Ang1 in osteoblasts was detected under each condition, the cells cultured with a higher concentration of Ca (50 mmol/L) did not show higher mineralization in the same way that those cultured with 6 mmol/L Ca did. It is generally understood that higher ionic conditions (high ionic strength) enhance the detachment of proteins from the substrate in the solution. 26 Therefore, higher Ca concentrations in the medium should tend to remove the precipitated matrix proteins secreted by osteoblasts. Because the precipitated and organized matrix proteins supply the region for the mineralization in osteoblast cultures, the balance between the protein secretion by cells and the detachment of precipitated matrix triggered by Ca is crucial for the regulation of mineralization.
A previous report indicated that Ang1 acts as a maturation factor ensuring tight association of adjacent endothelial cells, whereas Ang2 acts as a destabilizing signal and is expressed before vessel sprouting in angiogenesis. 27 Since the sprouting in angiogenesis is the postevent following the degeneration of basement membrane, Ang1 and Ang2 might be concerned with the cell–matrix and/or cell–cell interactions. As shown in Figure 1b and c, indeed, cells with rounded shape were observed when cells were cultured with a higher concentration of Ca (<14 mmol/L). Therefore, expression of integrin β1, the major marker of cell–matrix interaction, and connexin43, the major marker of cell–cell interaction, in osteoblasts cultured at different Ca concentrations were evaluated. Strikingly, the protein expression profile of connexin43 was similar to that of Ang1. In contrast, that of integrin β1 was similar to that of Ang2. These results suggested that expression of these HSC niche-related Ang proteins was closely correlated with the alterations of cell–matrix and cell–cell interactions, triggered by the presence of Ca ions. Previous reports stating that Ang proteins promote cell migration or cancer-cell metastasis through an integrin-related pathway also support this idea.28,29
Recent progress in degradable ceramic biomaterials, such as those made from tricalcium phosphate and octacalcium phosphate, has enabled them to be developed for biomedical applications, including bone tissue engineering and drug and/or gene delivery purposes. 30 Low crystalline and ion-substituted apatite are also considered as excellent candidates for these purposes, because Ca dissolution can be easily regulated in these materials.31,32 As shown in this study, Ca ions play pivotal roles in controlling cell–matrix or cell–cell interactions that alter the cell morphology in osteoblast cultures. These alterations in interactions greatly affect Ang expression that regulates osteogenic differentiation. According to our findings, the chemical properties of these alterations, especially those of the dissolution of these apatite materials at implanted sites, should be severely regulated to obtain good prognoses of bone regeneration or to shorten the recovery period. Moreover, since Ang proteins play pivotal roles in the HSCs' fate control, regulating Ang expression by Ca concentrations should be an effective approach for reproducing artificial HSC niches or for engineering bone marrow.
Footnotes
Acknowledgments
The authors are grateful to Ms. Shiho Ishihara for her technical assistance. This work was supported by the Japan–Korea Basic Scientific Cooperation Program from Japanese Society for the Promotion of Science (JSPS) and Korean Science and Engineering Foundation (KOSEF).
Disclosure Statement
No competing financial interests exist.