
Abstract
We have previously isolated and identified stem cells from human anterior cruciate ligament (ACL). The purpose of this study was to evaluate the differences in proliferation, differentiation, and extracellular matrix (ECM) formation abilities between bone marrow stem cells (BMSCs) and ACL-derived stem cells (LSCs) from the same donors when cultured with different growth factors, including basic fibroblast growth factor (bFGF), epidermal growth factor, and transforming growth factor-beta 1 (TGF-β1). Ligament tissues and bone marrow aspirate were obtained from patients undergoing total knee arthroplasty and ACL reconstruction surgeries. Proliferation, colony formation, and population doubling capacity as well as multilineage differentiation potentials of LSCs and BMSCs were compared. Gene expression and ECM production for ligament engineering were also evaluated. It was found that BMSCs possessed better osteogenic differentiation potential than LSCs, while similar adipogenic and chondrogenic differentiation abilities were observed. Proliferation rates of both LSCs and BMSCs were enhanced by bFGF and TGF-β1. TGF-β1 treatment significantly increased the expression of type I collagen, type III collagen, fibronectin, and alpha-smooth muscle actin in LSCs, but TGF-β1 only upregulated type I collagen and tenascin-c in BMSCs. Protein quantification further confirmed the results of differential gene expression and suggested that LSCs and BMSCs increase ECM production upon TGF-β1 treatment. In summary, in comparison with BMSCs, LSCs proliferate faster and maintain an undifferentiated state with bFGF treatment, whereas under TGF-β1 treatment, LSCs upregulate major tendinous gene expression and produce a robust amount of ligament ECM protein, making LSCs a potential cell source in future applications of ACL tissue engineering.
Introduction
Although adult fibroblasts retain many of the phenotypic qualities necessary for collagen synthesis, they are relatively quiescent and have limited potential for further differentiation. 22 Therefore, bone marrow stem cells (BMSCs) are an attractive candidate for ligament engineering, and they are currently under investigation for this purpose. BMSCs are derived from a source that maintains certain degree of self-renewal and has the potential to differentiate into cells of multiple mesenchymal lineages. Moreover, the synthetic and proliferative systems of these cells are robust, and the cells can be rapidly expanded in cell culture media and stimulated to differentiate into fibroblastic progenesis.23,24 A few comparative studies of BMSCs and ACL fibroblasts have been undertaken to evaluate the optimal cell source for ACL tissue engineering. 23 However, the harvest methods, culture techniques, and experimental results of these studies were quite different. The answer to the question of which being a better cell source for ligament tissue engineering remains uncertain.
In recent years, tissue-specific mesenchymal stem cells have been isolated from a variety of adult mesenchymal tissues in addition to bone marrow.25–30 In our previous study, we successfully isolated stem cells from human ACL and posterior cruciate ligament with multilineage differentiation ability and identical surface immunophenotype to BMSCs. 31 The differences in capacity of in vitro multilineage differentiation, cell proliferation, and ECM production between BMSCs and ACL-derived stem cells (LSCs) have not been previously evaluated. Therefore, the purpose of this study was to evaluate these important differences between BMSCs and LSCs from the same donors and further elucidate their potentials in ligament tissue engineering.
Materials and Methods
Isolation of BMSCs and LSCs
ACL tissues and bone marrow from the same donors were collected aseptically during primary total knee arthroplasty (TKA) procedures in four patients and arthroscopic surgeries for ACL rupture in two patients with informed consent. The diagnosis was advanced osteoarthritis in all patients undergoing TKA; there were two male and two female patients, aged 67–77 years. The two patients who underwent arthroscopy surgery were 25- and 39-year-old men. An institutional review board approval was obtained before the commencement of the study. Human LSCs and BMSCs were harvested from the same donor as previously described.31–33 Briefly, the ACL tissues were washed repeatedly in phosphate-buffered saline (PBS; Gibco, Grand Island, NY). The ACL tissues were then minced and digested with 1% collagenase in alpha-modified Eagle's medium (α-MEM; Gibco) containing 10% heat-inactivated fetal bovine serum (FBS; Sigma-Aldrich, St. Louis, MO). Following overnight incubation at 37°C, cells were collected by centrifugation and washed repeatedly in PBS. The cells were plated at 5000 cells/cm2 in T75 flasks with growth medium (α-MEM supplemented with 10% FBS) as passage 0. To isolate BMSCs, bone marrow was aspirated from distal femur during the surgical procedure, and nucleated cells were isolated with density gradient (Ficoll-Paque; Pharmacia, Uppsala, Sweden) and resuspended in α-MEM supplemented with 20% FBS. After 24 h, nonadherent cells were discarded and adherent cells were washed with PBS. These adherent cells were passage 0 cells.
Maintenance and culture expansion
Once adherent passage 0 cells from ACL and bone marrow reached semiconfluence, cells were detached with 0.25% trypsin–ethylenediaminetetraacetic acid (Gibco), washed twice with PBS, centrifuged at 1000 rpm (200 g) for 4 min, and replated as passage 1 at 50 cells/cm2 in each T75 flask. After an additional 14 days of growth, most of the committed and differentiated cells would fade out, and the active proliferative cells were harvested and cryopreserved. These cells were defined as LSCs and BMSCs. The cells were harvested by washing aliquots of the trypsin-released cells twice with PBS and cryopreserved in liquid nitrogen in FBS with 10% dimethyl sulfoxide (Sigma-Aldrich) (passage 1). To expand the cells, a vial of cryopreserved cells was thawed and plated at a concentration of 5000 cells/cm2 in a T75 flask. Serial passage of the cells was performed at the same concentration.
Colony-forming assay
A portion of the passage 1 LSCs and BMSCs from each donor was diluted and replated onto 100 mm culture plates at a concentration of 200 cells per plate. The experiment was performed in triplicates for each donor. After 14 days, colonies that had formed from single cells were visualized under a light microscope. The colonies were fixed with 4% formaldehyde and then stained with 0.5% crystal violet in methanol for 5 min. The plates were washed twice with distilled water, and the number of colonies per dish was determined. Colonies <2 mm in diameter and faintly stained colonies were ignored. 33
Population doublings of cells
To examine the proliferation and population doubling (PD) capacity of LSCs and BMSCs, the cells from passage 1 of each donor were replated at 5000 cells/cm2 in six-well plates in growth medium, and the cells were serially passaged. Medium changes were carried out twice a week. The cells were counted and replated at the same density when confluence was reached. We calculated the PD for each passage using the following formula: PD = log2[NC/N0], where N0 is the inoculum cell population and NC is the number of cells at confluence; then the PD values from all the passages were added together to obtain the overall PD capacity.
Effects of growth factors on cell proliferation
The effects of growth factors on the proliferation of LSCs and BMSCs were determined using a commercially available MTS [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt] kit (CellTiter 96 AQ; Promega, Madison, WI). Briefly, 10,000 cells were placed in each well of a 96-well plate. After stimulation with growth medium supplemented with 10 ng/mL basic fibroblast growth factor (bFGF), 10 ng/mL epidermal growth factor (EGF), or 10 ng/mL transforming growth factor-beta 1 (TGF-β1) for 72 h, the relative cell number was estimated by measuring the absorbance at 490 nm on a microplate reader (U3300 Pro; Amersham Biosciences, Freiburg, Germany).
Effects of growth factors on cell densities
Four thousand LSCs and BMSCs were placed in each well of six-well plates and cultured with growth medium and growth factors. The experimental group consisted of growth medium supplemented with 10 ng/mL bFGF, 10 ng/mL EGF, or 10 ng/mL TGF-β1. The control group was growth factor free. The culture medium was changed twice per week. Cell number was determined on days 7, 14, 21, and 28 by trypan blue exclusion in a Neubauer counting chamber. Three independent experiments were performed for each donor at each time point.
In vitro differentiation and histologic and cytochemical analysis
To induce chondrogenesis, osteogenesis, and adipogenesis, second to fifth passage cells were seeded in six-well plates and cultured by methods previously reported in the literature, with slight modifications.29–31,34,35 For histochemical analysis of osteogenesis, cells were rinsed twice with PBS, fixed with 4% formaldehyde, and washed with distilled water. Alkaline phosphatase (Alk-P) histochemical analysis was performed using the nitroblue/5-bromo-4-chloro-3-indolyl phosphate (BCIP) solution (Roche Molecular Biochemicals, Mannheim, Germany), according to the manufacturer's instructions. To evaluate the mineralized matrix, cells were fixed with 4% formaldehyde and stained with 1% Alizarin-Red S (Sigma-Aldrich) solution in water for 10 min. For Oil-Red O staining, cells were fixed with 4% formaldehyde and stained with Oil-Red O (Sigma-Aldrich) for 10 min. Chondrogenic differentiation was evaluated after pellets were fixed with 4% formaldehyde, dehydrated in serial ethanol dilutions, and embedded in paraffin blocks. Blocks were then cut and sections were stained with Alcian blue.
RNA extraction and semiquantitative reverse transcription–polymerase chain reaction analysis
Total RNA from the cells was extracted using a commercially available kit (S.N.A.P.; Invitrogen, Groningen, The Netherlands), according to the manufacturer's instructions. Complementary DNA (cDNA) was obtained by reverse transcription (RT) of 1 μg total RNA using Advantage RT-for-PCR (Clontech, Palo Alto, CA), according to the manufacturer's instructions. cDNA was amplified using an ABI GeneAmp polymerase chain reaction (PCR) system 2400 (Perkin Elmer Applied Biosystems, Boston, MA). The primers used for amplification are listed in Table 1.
AGC1, aggrecan 1; COL2A1, collagen type IIA1; COLI, collagen type I; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; FABP, fatty acid-binding protein; PPAR-γ, peroxisome proliferator-activated receptor gamma; OC, osteocalcin.
Real-time RT-PCR analysis
Four thousand LSCs and BMSCs were placed in each well of six-well plates and cultured with growth medium and growth factors. The experimental groups consisted of growth medium supplemented with 10 ng/mL bFGF, 10 ng/mL EGF, or 10 ng/mL TGF-β1. The control group was growth factor free. The culture medium was changed twice per week. Cells were harvested at days 7, 14, 21, and 28. Total RNA from the cells in each group was extracted using a commercially available kit (S.N.A.P.), according to the manufacturer's instructions. cDNA was obtained by RT of 1 μg total RNA using Advantage RT-for-PCR (Clontech), according to the manufacturer's instructions. Quantitative PCR was conducted on a Roche LightCycler® 480 (Roche Diagnostics, Laval, Canada) real-time PCR system. The primer sequences were designed using the Universal ProbeLibrary for Human (Roche Applied Science, Laval, Canada) with nucleotide sequences deposited in the GenBank database. Primer sequences and PCR conditions are described in Table 2. Calculations for determining the relative level of gene expression were made using the cycle threshold (Ct) method. The mean Ct value from duplicate measurements was used to calculate expression of the target gene, which was expressed as fold change in comparison with the house-keeping gene glyceraldehyde-3-phosphate dehydrogenase. Negative controls without RNA template and without addition of the RT enzyme were also performed.
COL1A1, collagen type IA1; COL3A1, collagen type IIIA1; α-SMA, alpha-smooth muscle actin; Tm, melting temperature (°C).
Quantification of collagen and total ECM proteins
Collagen and proteins were quantified by colorimetric analyses, as described previously.36,37 The method used the selective binding of Sirius Red F3BA to collagen and Fast Green FCF (Sigma-Aldrich) to noncollagen protein when both are dissolved in aqueous saturated picric acid. Briefly, 2000 LSCs and BMSCs were placed in each well in 12-well culture plates and cultured with growth medium and growth factors. The experimental groups consisted of growth medium supplemented with 10 ng/mL bFGF, 10 ng/mL EGF, or 10 ng/mL TGF-β1. The control group was growth factor free. The culture medium was changed twice per week. After cultured for 7, 14, and 21 days, each well was incubated with 1 mL of saturated picric acid solution that contained 0.1% Sirius Red F3BA and 0.1% Fast Green FCF. The plates were incubated at room temperature for 30 min in a rotary shaker. The fluids were then carefully withdrawn, and the plates were washed repeatedly with distilled water until the fluid was colorless. After washing with water, 1 mL of 1:1 (vol/vol) 0.1% NaOH and absolute methanol was added to the plates to elute the color. The eluted color was immediately read using a spectrophotometer at 540 and 605 nm (U3300 Pro).
Statistical analysis
Results are shown as mean values ± standard deviation. Statistically significant differences between groups were analyzed by independent-sample t-test and multivariate analysis of variance, followed by univariate analysis of variance and the Scheffé test (SPSS version 11.5; SPSS, Chicago, IL). A p-value of less than 0.05 was considered statistically significant. To indicate the p-value between different groups, multiple alphabetical expressions were used in Figures 2E, 4A–E, and 5B.
Results
Isolation and expansion of cells
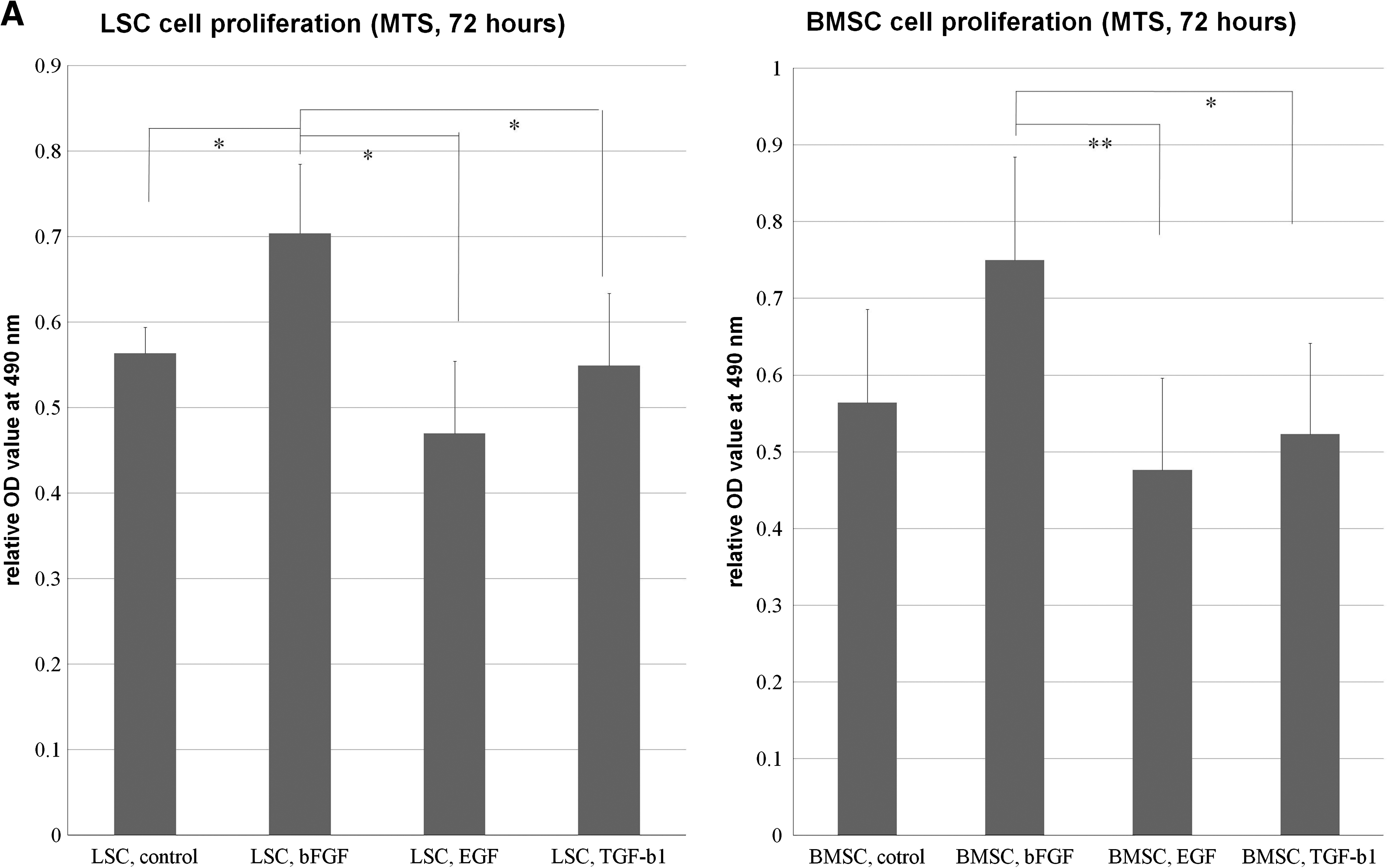
LSCs and BMSCs were able to be extensively subcultivated. The cells were plastic adherent and spindle shaped (Fig. 1). The morphology of bone marrow and ACL cells from patients undergoing TKA or ACLR were not different. In MTS experiment, the proliferation rate of LSCs was significantly increased with the bFGF treatment than control, EGF, and TGF-β1 treatment (0.01 < p < 0.05). The proliferation of BMSCs was significantly increased with the bFGF treatment than TGF-β1 (0.01 < p < 0.05) and EGF (p < 0.01) groups (Fig. 2A). Both BMSCs and LSCs could be serially passaged in vitro for more than 80 days with more than 25 PDs. The PD time was estimated to be 64.5 h for BMSCs and 69.5 h for LSCs, and there was no statically significant difference (p = 0.234; Fig. 2B).
Morphology of LSCs and BMSCs. The morphology of clonally expanded cells derived from ACL (left) and bone marrow (right). The cells are spindle shaped, fibroblast-like, and plastic adherent. Scale bar indicates 100 μm. ACL, anterior cruciate ligament; LSCs, ACL-derived stem cells; BMSCs, bone marrow stem cells.
Comparison of LSCs and BMSCs. (
Ligament-derived cells are clonogenic
The average colony-forming efficiency per 200 cells was determined to be 40.7 ± 10.1 for LSCs and 29.8 ± 4.8 for BMSCs. The results are shown as the mean ± standard deviation of six donors in each group. The difference was statistically significant (p = 0.039). LSCs have significantly higher colony-forming ability than BMSCs (Fig. 2C). The colony-forming efficiency of LSCs and BMSCs from patients undergoing TKA (n = 6) and ACLR (n = 2) were also compared but without significant difference.
Effect of growth factors on cell proliferation and density
In both LSC and BMSC cultures, the cells treated with bFGF developed smaller cells with short cell processes. Cells cultured in TGF-β1 had a flattened cell shape with longer cell processes; these cells tended to aggregate together to form linear or circular multiple cell complexes. In the EGF and control groups, the cell size was between that of the aforementioned two groups (Fig. 2D). In the first week, only LSCs treated with bFGF had significantly more cell number than BMSC treated with EGF and TGF-β1 (p < 0.01). BMSCs and LSCs treated with bFGF in the second week had significant higher cell numbers than all other groups (p < 0.05) except BMSCs treated with TGF-β1, which had significant more cells than LSCs in control and EGF-treating group (p < 0.05). BMSCs and LSCs treated with bFGF and TGF-β1 had significant higher cell number than other groups (p < 0.01) in the third week. In the fourth week, LSCs treated with bFGF and TGF-β1 had significant higher cell numbers than BMSCs treated with bFGF and TGF-β1 (0.01 < p < 0.05) and all other groups (p < 0.01), whereas the cell number in BMSCs treated with bFGF and TGF-β1 were higher than LSCs and BMSCs treated with EGF (p < 0.05). The cell number and cell density continued to increase in the 4-week culture period in all groups (Fig. 2E).
Multilineage differentiation
Under osteogenic induction, BMSCs showed typical osteogenic differentiation and were positive for Alk-P and Alizarin-Red S stain. LSCs showed similar Alk-P staining but weak Alizarin-Red S staining under the same osteogenic condition (Fig. 3A). Under adipogenic induction, LSCs had faster fat droplet formation in the first and second weeks. After 3 weeks of induction, both LSCs and BMSCs showed typical fat droplet formation and were positive for Oil-Red O stain (Fig. 3B). Under chondrogenic condition, the cells aggregated to form pellets. The pellets from both LSCs and BMSCs were positive for Alcian blue stain (Fig. 3C). Specific PCR products of bone (type I collagen, osteocalcin) (Fig. 3A), fat (peroxisome proliferator-activated receptor gamma, fatty acid-binding protein) (Fig. 3B), and cartilage (type II collagen, aggrecan) (Fig. 3C) were expressed in cells treated for 3 weeks with osteogenic, adipogenic, and chondrogenic induction media, respectively.
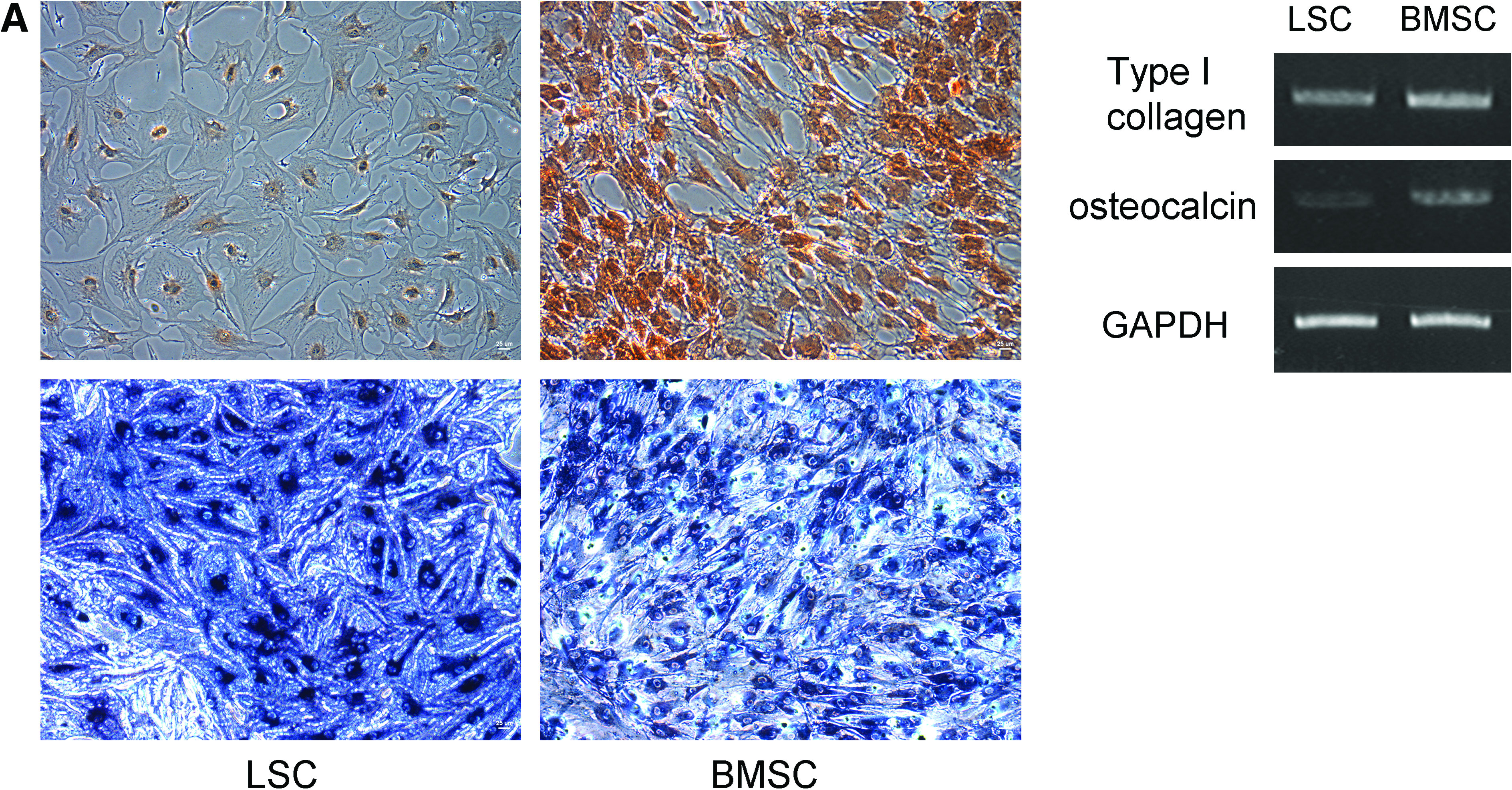
Multilineage differentiation of LSCs and BMSCs. (
Gene expression analysis
The relative gene expression of tenascin-c, fibronectin, alpha-smooth muscle actin (α-SMA), type I collagen, and type III collagen with growth factor treatment from 1 to 4 weeks is shown in Figure 4A–E. All these molecules are important in human cruciate ligaments. The mRNA expression of type I collagen was significantly increased in LSCs with TGF-β1 treatment in comparison with the control group in the first, second, and third weeks (p < 0.01), whereas in BMSCs only increased type I collagen expression was observed in the first week after TGF-β1 treatment (0.01 < p < 0.05) (Fig. 4A). Type III collagen mRNA expression was increased in the first week of LSC control group and LSCs treated with TGF-β1 from 1 to 4 weeks. After 2 weeks, TGF-β1 treatment of LSCs significantly increased type III collagen gene expression compared with that of control. In contrast, TGF-β1 treatment of BMSCs had no significant effect on type III collagen expression (Fig. 4B). The mRNA expression of fibronectin was highest in LSCs with TGF-β1 treatment from 1 to 4 weeks, but significant difference with control group only was noted in the third week. BMSCs with different treatments express significant low-level fibronectin than LSCs with TGF-β1 except in the first week (Fig. 4C). Expression of α-SMA was highest only in LSCs with TGF-β1 treatment for 4 weeks (p < 0.01; Fig. 4D). The mRNA expression of tenascin-c was increased in the BMSC control and EGF groups. Tenascin-c expression was low in all four groups of LSCs and BMSCs treated with FGF-2 (Fig. 4F).
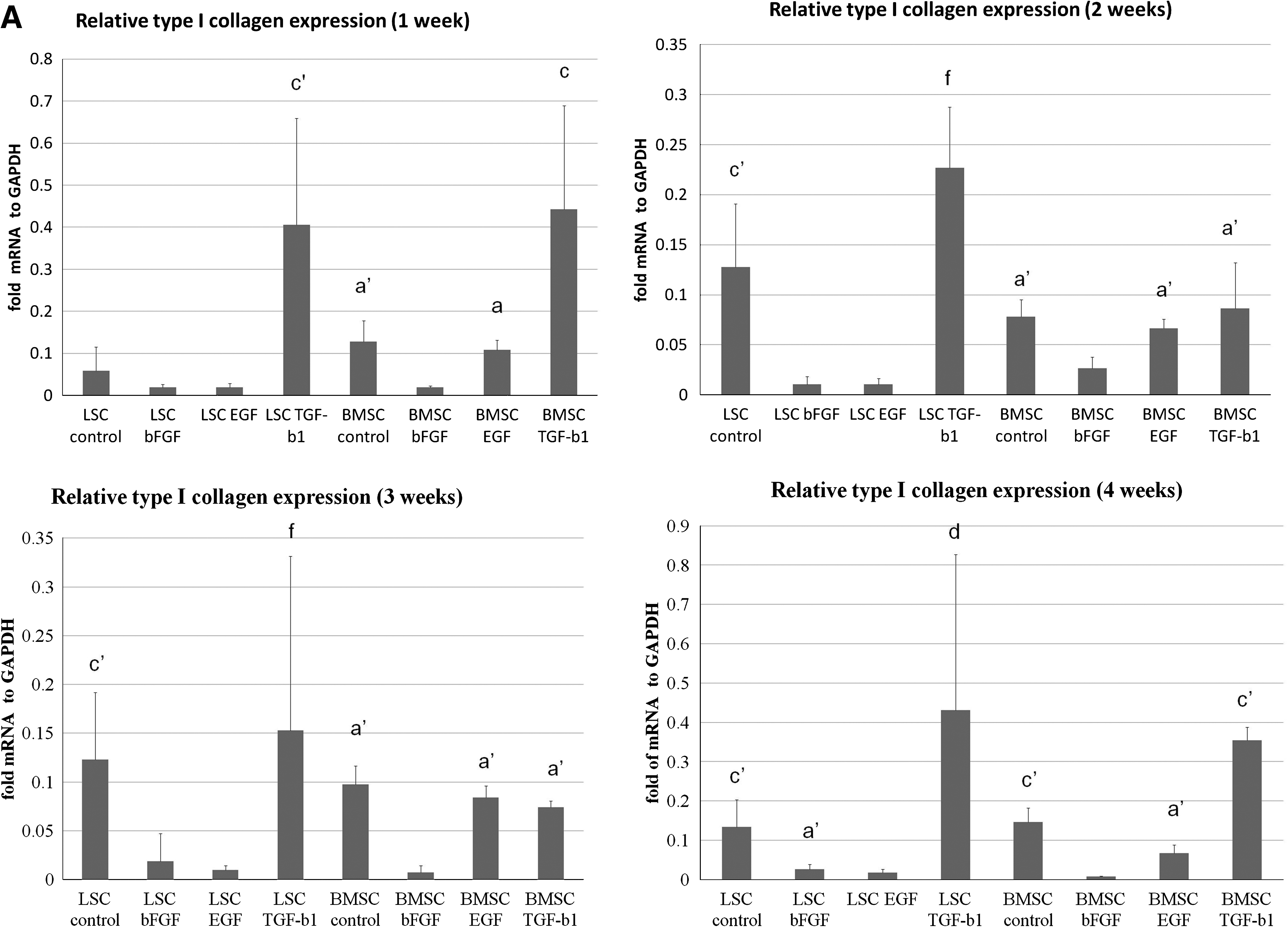
Relative transcript level of genes with different treatments. Four thousand LSCs and BMSCs were placed in each well of a six-well plate and cultured in 10 ng/mL bFGF, 10 ng/mL EGF, or 10 ng/mL TGF-β1 for 4 weeks. The control group had no growth factor. Cells from each donor were harvested weekly and the relative levels of collagen type I (
Quantification of collagen and total ECM protein production
In phase contrast microscopic examination, generally deep staining was noted in the TGF-β1 group (Fig. 5A). With spectrophotometer quantification, there was no significant difference between different treatments in the first week. LSCs and BMSCs treated with TGF-β1 significantly increase noncollagen protein production than other groups after 2 weeks (p < 0.01). In the second week, LSCs with TGF-β1 produce significant more collagen than other groups (p < 0.01) except LSCs with bFGF and BMSCs with TGF-β1. In the third week, LSCs with TGF-β1 produce significant more collagen than other groups except BMSCs with TGF-β1 treatment. After 3 weeks of culture, TGF-β1 treatment increased total ECM collagen and noncollagen protein production to 3.9 and 4.9 times that of the control group for LSCs, compared with 2.9 and 4.3 times that of the control group for BMSCs (Fig. 5B).
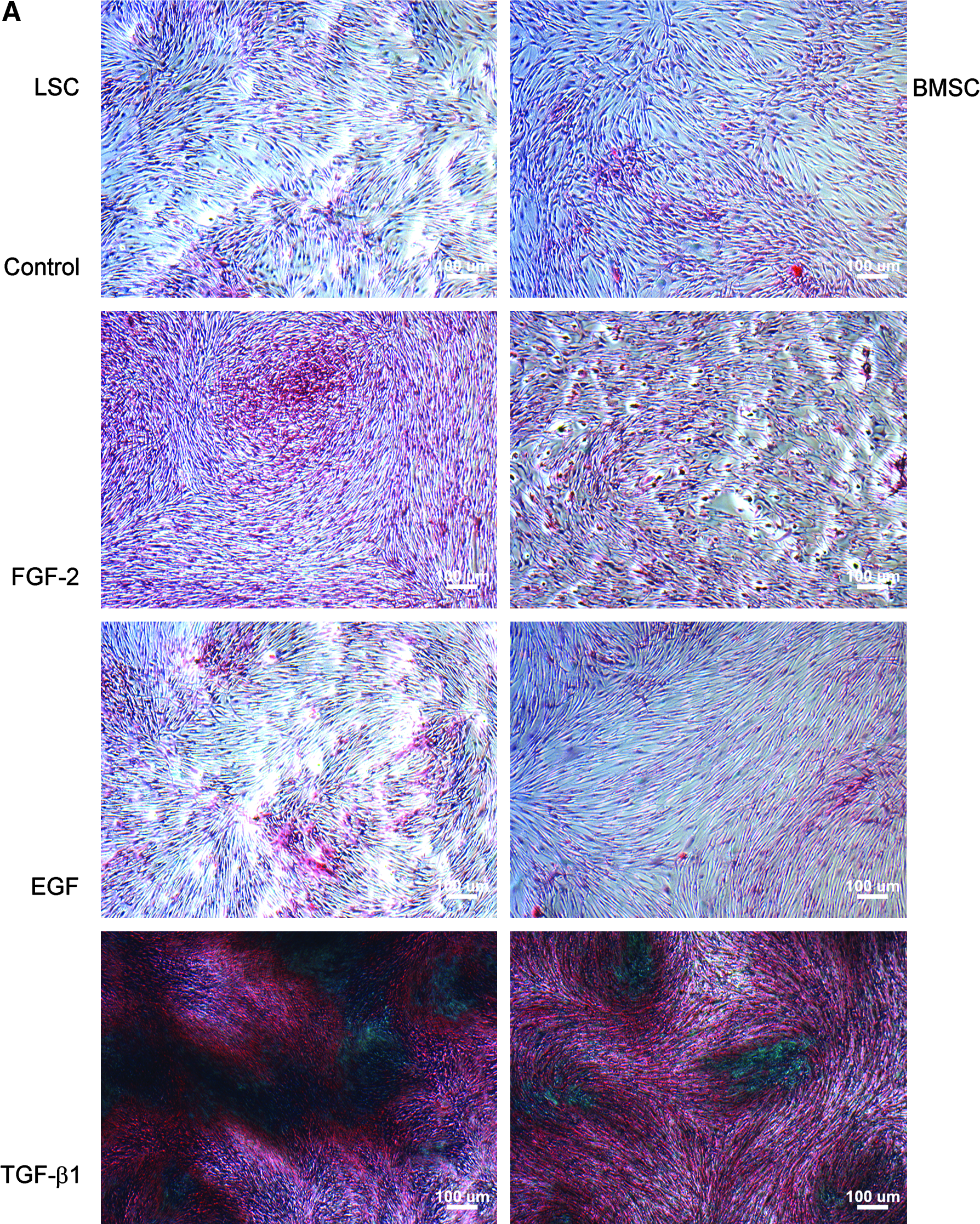
Collagen and noncollagen protein production. (
Discussion
Tissue engineering using stem cells is a fascinating approach to the clinically significant situation of ACL injury. Use of an appropriate cell source is one of the most important factors in the success of tissue engineering. Multiple reports have been published comparing fibroblasts or stem cells from different sources for ACL engineering. Nagineni et al. evaluated the difference between fibroblasts from rabbit ACL and MCL. They concluded that the lower proliferation and migration potential of ACL cells might contribute to the inferior healing potential of ACL compared with MCL. 38 Cooper et al. studied the proliferation and gene expression of fibroblasts from rabbit ACL, patellar tendon, Achilles tendon, and MCL. 39 They reported that ACL fibroblasts have inferior proliferation but superior type I and type III collagen gene expression compared with fibroblasts from other sources. Vunjak-Novakovic et al. compared goat BMSCs, ACL fibroblasts, and skin fibroblasts to evaluate the optimal cell source for ACL engineering. 23 They found that BMSCs showed the highest DNA content and collagen production. 23 Collectively, these early studies suggested that animal ACL fibroblasts had inferior proliferation and migration ability, as well as lower gene expression and collagen production, compared with BMSCs. However, the results of these animal studies have limited significance in human tissue engineering. To determine whether human LSCs possess the same properties as animal ACL fibroblasts, which have inferior proliferation, gene expression, and collagen production compared with BMSCs, we conducted a donor-matched comparison study of human LSCs and BMSCs.
In this study, significant differences from previously published results were found. First, human LSCs have inferior proliferation capacity compared with BMSCs only in PDs. The PD time of LSCs is slightly longer than that of BMSCs but without significant difference. However, no significant difference was found in proliferation ability between LSCs and BMSCs in the control medium at all time points. The effect of growth factors on cell proliferation was somewhat similar in LSCs and BMSCs, with the fastest proliferation in the bFGF group, followed by the TGF-β1, control, and EGF groups. In the 4-week continual cell proliferation and density study using six-well plates, the proliferation ability and cell density of LSCs and BMSCs in control medium were similar at all time points. Cells treated with bFGF and TGF-β1, however, had significantly higher proliferation after the second week. Interestingly, LSCs and BMSCs treated with TGF-β1 were low in cell number than control only in the initial 72 h and the first weeks, but LSCs and BMSCs treated with bFGF had higher cell number than control at all time points. To summarize the MTS and cell density study, we found that with lower cell density and limited culture time, bFGF significantly enhanced cell proliferation. With increased cell density and culture period, TGF-β1 had higher proliferation potential than FGF-2 for both LSCs and BMSCs. EGF treatment was associated with significant proliferation inhibition for both LSCs and BMSCs in both 72-h and 4-week studies.
The reasons that many researchers use BMSCs for ligament tissue engineering are that these cells maintain some degree of self-renewal and have the potential to differentiate into cells of multiple mesenchymal lineages. The synthetic and proliferative abilities of these cells are robust. The cells can be rapidly expanded in cell culture media and stimulated to differentiate into fibroblastic progenesis.23,24 In contrast, although adult ACL fibroblasts retain many of the phenotypic qualities necessary for collagen synthesis, they are relatively quiescent and have limited potential for further differentiation. 22 In previous research, we isolated human LSCs with the capacity for multilineage differentiation, colony formation, anchorage-independent growth, expression of self-renewal genes, and identical surface immunophenotypes to BMSCs. In this study, we further compared the proliferation, multilineage differentiation, colony formation, and PD properties of LSCs and BMSCs in a donor-matched fashion. We found that human LSCs possessed similar multilineage differentiation potentials as BMSCs, although LSCs seemed to have the propensity toward faster adipogenic differentiation and slower osteogenic differentiation in comparison with BMSCs. The results showed that although stem cells isolated from different tissues share most major important stem cells characteristics, certain properties of stem cells can be affected by their origins. This finding was previously pointed out by Sakaguchi et al., who found that stem cells from synovium have higher chondrogenic differentiation potential than those from bone marrow. 34 This result is further confirmed by our study, evidenced by the superior fibroblast-lineage gene expression of LSCs compared with BMSCs. In real-time PCR, TGF-β1 treatment significantly increased the type I collagen, type III collagen, fibronectin, and α-SMA gene expression of LSCs compared with that of BMSCs treated with TGF-β1. The collagen and noncollagen protein quantification study confirmed the results of real-time PCR. Upon TGF-β1 treatment for 3 weeks, total ECM collagen and noncollagen protein production increased to 3.9- and 4.9-fold than that of the control group for LSCs, compared with the increase between 2.9- and 4.3-fold than that of the control group for BMSCs. All these data confirmed the hypothesis that tissue-specific stem cells may retain some tissue-specific differentiation properties. LSCs have both ACL fibroblast and stem cell characteristics, which may make them more advantageous than BMSCs in ligament tissue engineering.
Colony formation is one of the most important characteristics of stem cells. Surprisingly, the average colony-forming unit for LSCs was significantly higher than that for BMSCs in this study. There are two possible explanations for this finding. The first is that LSCs were better maintained with this cell isolation technique and culture condition. The other possible reason is the old age of our donors. The frequency of BMSCs declines with the donor age, and the number of colonies formed by cells from old donors was fewer and the total colony area was less than those from young donors.40,41 The same age-related effect may have less impacts on LSCs, as previously reported by Mochizuki et al., 42 who found that the proliferation and differentiation potentials of synovium-derived stem cells were not influenced by donor age. Although there is no strong evidence to support this hypothesis so far, to further compare the age-related effect on intraarticularly residing stem cells and BMSCs would be of scientific interest.
Ligaments contain parallel bundles of ECM composed mainly of type I collagen and a small portion of type III collagen. 43 In the process of early ligament healing, however, most of the newly synthesized collagen is type III collagen, with production of a small amount of type I collagen. 44 The type III collagen is also increased in the remodeling process after tendon grafting, termed “ligamentation.” 45 In our study, TGF-β1 treatment for 4 weeks significantly increased the gene expression of type III collagen and type I collagen in LSCs (7.76- and 3.22-fold, respectively). In contrast, TGF-β1 treatment significantly increased the gene expression of type I collagen but decreased the gene expression of type III collagen in BMSCs (2.41- and 0.25-fold, respectively). The results showed that TGF-β1 treatment of LSCs much better recapitulates the process of ligament healing or ligamentation than does TGF-β1 treatment of BMSCs.
Numerous studies have investigated the effects of different growth factors on a ligament engineering system with respect to cell proliferation, ECM synthesis, neovascularization, and mechanical properties. 46 Most investigations have focused on the growth factors known to have mitogenic effects in musculoskeletal systems. In previous studies, FGF, TGF, platelet-derived growth factor, EGF, insulin-like growth factor, and growth and differentiation factor have all shown the capacity to improve cell proliferation or matrix production. Moreau et al. reported that media supplemented with three growth factors—EGF, FGF-2, and TGF—enhanced fibroblast differentiation. 47 In our study, however, fibroblast differentiation was strongly enhanced in both BMSCs and LSCs with the addition of TGF-β1, but not EGF or FGF-2. In our experiment, both LSCs and BMSCs were relatively low in ECM gene expression and ECM production with EGF treatment. Moreover, EGF also had no significant effect on cell morphology and proliferation; its main function may be to maintain LSCs and BMSCs in an undifferentiated state. FGF-2, a strong mitogen of stem cells, significantly increased the proliferation and cell density of LSCs and BMSCs and kept the gene expression at low levels, both of which are very important for stem cell expansion in vitro for further tissue engineering. The precise mechanism of action, temporal demand, and physiologic concentration of these agents, as well as the incorporation of effective factors into biodegradable polymers to achieve sustained release in vivo warrant future investigation.
Numerous experiments have been performed to evaluate the optimal conditions and material for ligament tissue engineering.18–20,22–24,39,48–50 No studies to date, however, have compared the properties of proliferation, differentiation, gene expression, and ECM production of human LSCs and BMSCs to determine the optimal cell source for ACL tissue engineering. In this study, we found that the PD time of BMSCs is shorter than that of LSCs but without significant difference, whereas LSCs have significantly greater colony formation ability than BMSCs. TGF-β1 and bFGF both increase cell proliferation in LSCs and BMSCs, with significantly more cells noted in LSC groups at 4 weeks. Real-time RT-PCR revealed that TGF-β1 treatment significantly increased type I collagen, type III collagen, fibronectin, and α-SMA mRNA expression in LSCs, but the effect of TGF-β1 on BMSCs is limited to type I collagen and tenascin-c upregulation. Protein quantification confirmed the result of real-time RT-PCR and suggested that LSCs have higher potential for ECM production with TGF-β1 treatment than do BMSCs. In summary, in comparison with BMSCs, LSCs proliferate faster and maintain an undifferentiated state with bFGF treatment, whereas upon TGF-β1 treatment, LSCs upregulate major tendinous gene expression and produce a robust amount of ligament ECM protein, making LSCs a potential cell source in future applications of ACL tissue engineering.
Footnotes
Acknowledgments
The authors acknowledge financial support from the Taipei Veterans General Hospital (VGH98E1-001) and the National Science Council, Taiwan (NSC98-2314-B-010-001-MY3, NSC97-2627-B-010-010, and NSC98-3111-B-010-003). This study was also supported by a grant from the Ministry of Education, Aim for the Top University Plan.
Disclosure Statement
No competing financial interests exist.