
Abstract
We used the tarsus of an adult Xenopus laevis frog as an in vivo load-bearing model to study the regeneration of critical-size defects (CSD) in long bones. We found the CSD for this bone to be about 35% of the tarsus length. To promote regeneration, we implanted biocompatible 1,6 hexanediol diacrylate scaffolds soaked with bone morphogenetic proteins-4 and vascular endothelial growth factors. In contrast to studies that use scaffolds as templates for bone formation, we used scaffolds as a growth factor delivery vehicle to promote cartilage-to-bone regeneration. Defects in control frogs were filled with scaffolds lacking growth factors. The limbs were harvested at a series of time points ranging from 3 weeks to 6 months after implantation and evaluated using micro-computed tomography and histology. In frogs treated with growth factor-loaded scaffolds, we observed a cartilage-to-bone regeneration in the skeletal defect. Five out of eight defects were completely filled with cartilage by 6 weeks. Blood vessels had invaded the cartilage, and bone was beginning to form in ossifying centers. By 3 months, these processes were well advanced, and extensive ossification was observed in 6-month samples. In contrast, the defects in control frogs showed only formation of fibrous scar tissue. This study demonstrates the utility of a Xenopus model system for tissue engineering research and that the normal in vivo mechanism of endochondral bone development and fracture repair can be mimicked in the repair of CSD with scaffolds used as growth factor delivery mechanisms.
Introduction
Current treatments for repair of large bone defects include distraction osteogenesis, or bridging the defect with a bone graft, or a natural or synthetic scaffold. 5 Distraction osteogenesis, based on the Ilizarov technique, is frequently employed in clinical practice, but it has its drawbacks, including a prolonged wearing of a heavy frame and fixator, substantial demands on patient compliance, high rates of complications, and other issues.6–11 New surgical techniques for distraction osteogenesis have been proposed. 12 Autogenous (i.e., from same human) bone grafts have been used with varying degrees of success, but this process is traumatic due to harvesting of bone and subsequent high morbidity at the donor site.13–16 Allogenic bone (i.e., from other humans) 17 or xenogenic bone (from nonhuman species) grafts have also been used, but they have other drawbacks such as a risk of rejection and disease transmission. 18
Osteoinductive scaffolds, the product of tissue engineering research, are one of the most promising techniques for long bone CSD repair. Currently, most experimental approaches seek to directly deposit new bone on scaffolds. The scaffolds can be treated with growth factors to encourage ingrowth of bone-forming cells from surrounding tissues, or seeded with osteogenic stem cells or osteoblasts before implantation.19–22 For this approach, scaffolds are required (i) to be three-dimensional (3D) and highly porous with an interconnected pore network for cell growth and flow transport of nutrients and metabolic waste; (ii) to be biocompatible and bioresorbable with a controllable degradation and resorption rate to match cell/tissue growth in vitro and/or in vivo; (iii) to have suitable surface chemistry for cell attachment, proliferation, and differentiation; and (iv) to have mechanical properties to match those of the tissues at the site of implantation. 23 A scaffold of this type provides a necessary support for bone-forming cells to proliferate and differentiate, and its geometry defines the ultimate shape of new bone. Biological or synthetic scaffolds have been used combined with variety of growth factors22,24–30 and seeded with osteoblasts or mesenchymal stem cells.31–36 Other methods, including the use of cellular transplantation and bioactive molecule treatments, have also been considered and explored in various combinations to address the problem of large bone defect repair.20–22,37–49
Normal fracture repair of long bones is essentially a recapitulation of the normal embryological developmental of long bones—an endochondral ossification, in which a cartilage template is first formed and is subsequently ossified and remodeled into a mature lamellar bone. 50 Thus, attempts to directly regenerate bone within a scaffold, while appropriate for CSDs of intramembranous bones, may not be ideal for the repair of endochondral bones.
Several growth factors have been investigated for their ability to enhance bone healing in bone injury studies, most notably the bone morphogenetic proteins (BMP) 2, 4, and 7.51–53 These factors have been shown to promote ectopic bone formation in a number of systems, as well as playing critical roles in fracture healing.50–53 BMP-4 expression is induced in the periosteum very early postfracture in mice and is important for cartilage callus formation.50,54,55 Angiogenic factors, most notably the vascular endothelial growth factors (VEGF), also play critical roles in endochondral ossification and fracture repair.50,56 VEGF activity is critical for angiogenesis and remodeling of hypertrophic cartilage for ossification to take place in the later stages of endochondral ossification, 56 and it plays an important role in early events of bone healing. 53
In this article, we have developed a novel experimental model system for studying the repair of long bone CSDs, the tarsus of the adult frog Xenopus laevis. Like mammalian species, Xenopus is also unable to regenerate CSD of long bones. Although seemingly unorthodox as a choice for a model system for bone CSD repair, use of Xenopus as a model organism has some advantages over mammalian models. Moreover, long bone development and fracture repair in Xenopus also take place by an endochondral ossification process that is essentially the same as in mammals. 57 Using this system, we have been able to induce endochondral bone ossification across a CSD by using a scaffold to deliver two growth factors known to be important regulators of endochondral ossification and bone healing, BMP-4 and VEGF.
Materials and Methods
Scaffold
Fabrication
1,6 hexanediol diacrylate (Sartomer Company, Inc.) scaffolds were fabricated by an innovative 3D microfabrication technology, Projection Micro Stereolithography (PμSL). 1,6 hexanediol diacrylate was chosen for our initial study due to its ease of use in the PμSL process, and its ability to support cell growth in culture without adverse effects. 58 These biocompatible scaffolds were designed as hollow cylinders, with a 1.2 mm outer diameter and a 0.8 mm inner diameter, and a porous wall structure with 50 μm pores as shown in Figure 1. These dimensions were chosen to match the transverse dimensions of tarsus in the diaphysis region. The process of PμSL starts by generating 3D structures using Computer Aided Design software and then slicing the structure into a sequence of bitmap images according to the desired spatial resolution. Each image represents a thin layer of microstructure. During one fabrication cycle, one image is displayed on the micro display chip. The image is delivered and captured by a reduction lens. The reduced image is focused on the UV photo curable liquid surface. A whole layer (usually 2–20 μm thick) is polymerized simultaneously. After one layer is solidified, the polymerized part is immersed into the liquid surface to allow a new thin liquid layer to form on top and a new fabrication cycle starts. By repeating the cycles, a 3D microstructure is formed from the stack of layers.

Scanning electron microscopy image of the porous hollow cylinder scaffold used in this study.
Preparation of scaffolds for implantation
Upon completion of fabrication, the scaffolds are immersed and stored in acetone. The combination of UV photocuring and acetone immersion results in sterile scaffolds. After manufacture, scaffolds were handled under aseptic conditions to maintain sterility. Scaffolds were rinsed several times and soaked in sterile distilled water for at least 48 h before experimental usage to remove all traces of acetone. On the day of surgical implantation, scaffolds were soaked with a solution of 10 μg/mL human BMP-4 (B2680; Sigma) and 25 μg/mL human VEGF (V7259; Sigma) in sterile phosphate-buffered saline (PBS) containing 0.1% bovine serum albumin at room temperature for 2 h before implantation, and were trimmed to match a surgically created CSD in one segment of the tarsus bone of the left hind limb.
In vitro growth factor release profile determination
A 5- mm-long scaffold was soaked in 20,000 pg/mL human BMP-4 solution for 12 h at 2°C. The scaffold then was submerged in 500 μL calibration diluent solution stored in at 2 mL polyethylene vial. A 50 μL sample was taken from the solution surface using a 100 μL micropipette every 10 min, and 50 μL fresh diluent was added back into the vial. The BMP-4 concentration in each sample was measured using the Quantikine© Human BMP-4 immunoassay kits from R&D Systems. The release profile is shown in Figure 2.
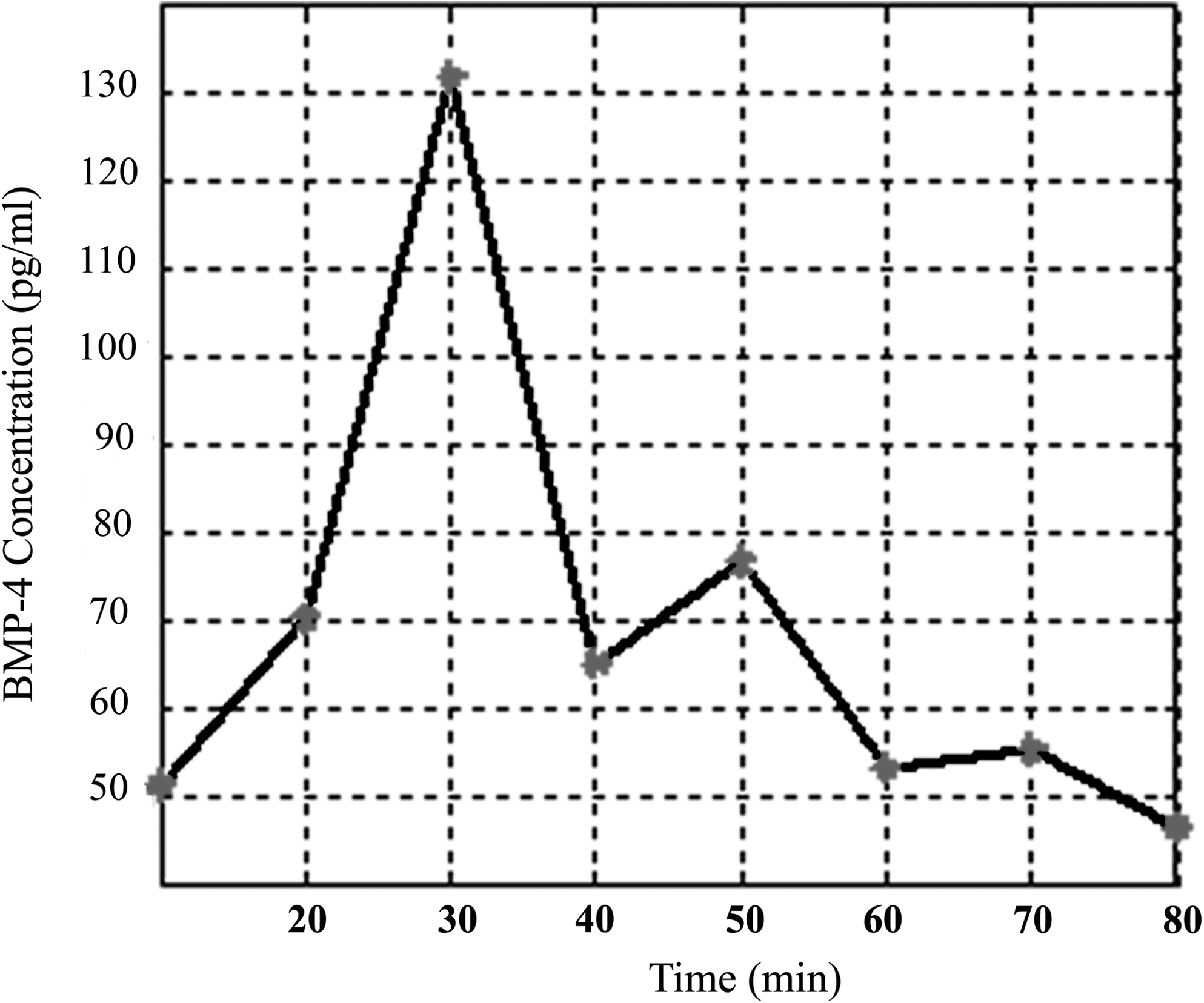
In vitro BMP-4 release profile of porous 1,6 hexanediol diacrylate scaffolds. 1,6 hexanediol diacrylate scaffolds used for this study were loaded with BMP-4 and, growth factor release was profiled in vitro. After an initial burst of growth factor release, the remaining growth factor was depleted after 3 h. BMP, bone morphogenetic proteins.
In vivo experiments
Animals
Sexually mature X. laevis frogs were obtained from Xenopus Express, Inc. The frogs were allowed to acclimate to local conditions for 1 week before surgery.
Surgery
All surgeries and animal care were performed in accordance with the University of Illinois at Urbana–Champaign Institutional Animal Care and Committee procedures and approved protocols. Animals were divided into three groups: CSD determination group (n = 12), control scaffold group (n = 9), and treated scaffold group (n = 19). The frogs were anesthetized in an osmotically balanced, dechlorinated water containing 0.15% ethyl 3-aminobenzoate methanesulfonate salt (MS222; Sigma), a standard amphibian anesthetic. Frogs were left in the anesthesia until pinching the toes with a forceps caused no response.
To determine the CSD, central (diaphyseal) segments of posterior tarsus bone of left hind limbs were excised, ranging from 1.5 to 8 mm in length (the length of the tarsus is 12 mm on average). The anterior tarsus bone was left intact to maintain structural support and also to act as an internal control. This process also damaged lateral diaphyseal muscles, allowing us to observe the interaction between regenerating bone and muscle. The wounds were then closed with 6–0 Ethilon 0.7 metric sutures from Ethicon. A schematic diagram showing the area of tarsus removal is shown in Figure 3.
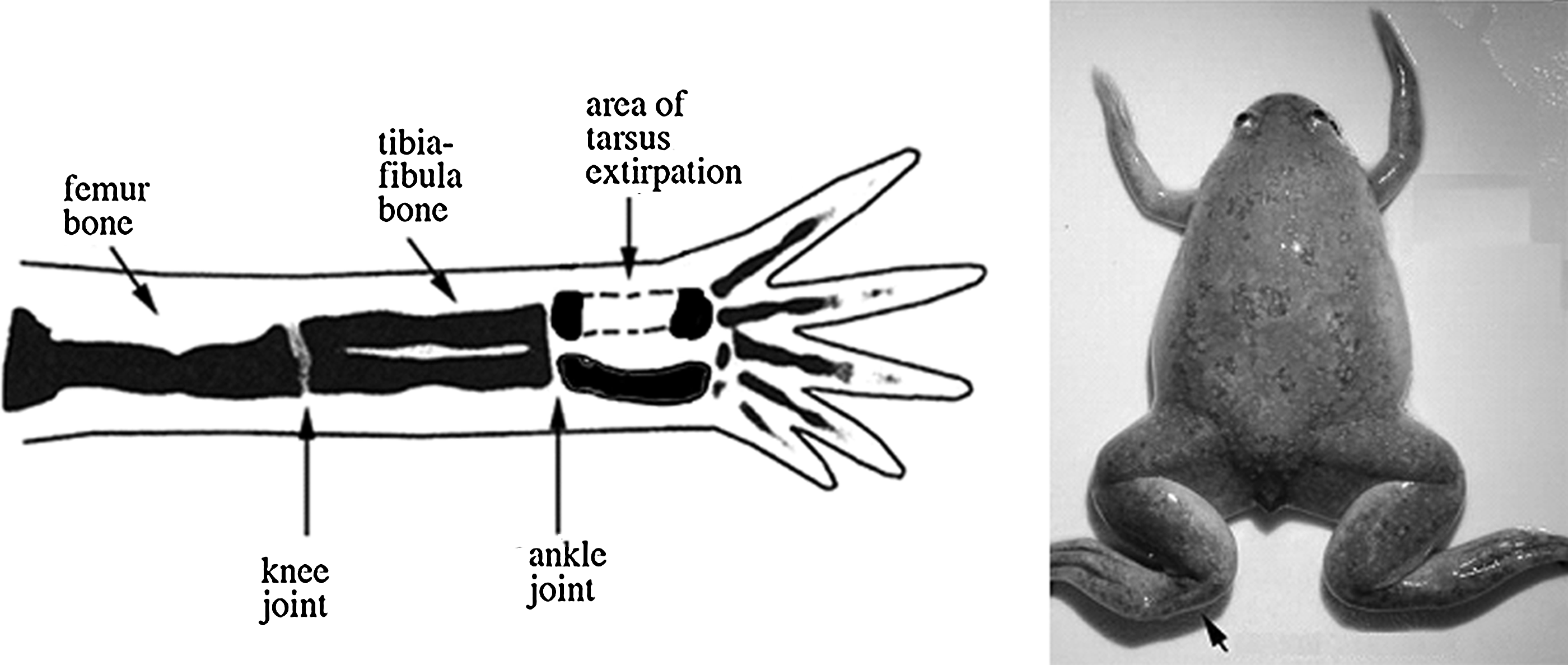
Schematic of bone removal surgery performed on Xenopus laevis hind limbs.
For the other two groups, CSDs were made in the posterior tarsus, and the defects received one of the following treatments: implantation of distilled water-soaked scaffolds (control scaffold group) and implantation of growth factor-treated scaffolds (treated scaffold group). The excised segmental pieces were measured and scaffolds were cut to corresponding lengths. The treated scaffold group underwent implantations with scaffolds that had been soaked for 2 h in a growth factor solution containing BMP-4 and VEGF (see above). After scaffolds were placed in the defect space, the wounds were closed with sutures. The frogs were then placed in osmotically balanced Holtfreter's solution in dechlorinated water to recover from the anesthesia. Frogs were closely observed throughout the wound healing process.
Sample collection and fixation
All samples in the critical-size determination group were collected at 3 months after surgery, whereas samples in the other two groups were collected at the following time steps: 3 weeks, 6 weeks, 3 months, and 6 months after surgery. After frog euthanization by over-anesthetization with a 0.3% MS222 solution in osmotically balanced dechlorinated water, the tarsus limb segments were collected using a single-sided razor blade to cut just distal to the tarsal–metatarsal joint and just proximal to the ankle joint. The tarsus samples were washed for 10 min in 1 × PBS in three subsequent washes, and then fixed in 4.0% paraformaldehyde in 1 × PBS overnight.
Micro-computed tomography imaging
Tarsus samples were put into Eppendorf tubes and scanned with a Sky Scan 1172 MicroCT (SkyScan) at the 11μm voxel resolution to quantitatively assess the status and ingrowth of the calcified tissues in the tarsus. After scanning, Amira 4.0 software was used to postprocess the imaged data. The thresholds were selected in each sample to isolate the calcified tissues while excluding the soft tissues and polymer scaffolds and they were used to generate 3D images of the tarsus bone. The actual sizes of the tarsus bone and defect were measured using these 3D images. The defect percentages were calculated as the ratios of the defect size to the whole tarsus size.
Histology and immunostaining
After fixation and micro-computed tomography (micro-CT) imaging, samples were prepared for cryosectioning. The samples were decalcified in Calci-Clear Rapid (National Diagnostics) solution for 6 h, washed with 1 × PBS, and then cryoprotected by sequential incubation in sterile solutions of 10%, 20%, and 30% sucrose in 1 × PBS. Samples were incubated in each sucrose solution for at least 24 h. Samples were then transferred into embedding molds with a 50:50 solution of 30% sucrose in 1 × PBS and Neg 50 frozen section medium (Richard-Allan Scientific), and frozen in isopentane cooled by liquid nitrogen. The embedded samples were then cryosectioned using a Leica CM 1900 cryostat. Sections were cut at a thickness of 20–30 μm, as this thickness provided the best preservation of tissue morphology and minimization of sectioning artifacts. All samples were cut completely through the defect site, and virtually all sections were collected and used for analysis. After collection, sections were stained with hematoxylin and eosin using standard methods. Images were captured using either a Nanozoomer digital pathology system (Hamamatsu Photonics) or a Zeiss Stereolumar v12 stereomicroscope (Carl Zeiss Microimaging).
A few sections were set aside and used for immunostaining for β1 integrin and sarcomeric myosin heavy chain. Immunostaining was performed as described previously. 59 Mouse monoclonal antibodies for β1 integrin (8C8) and sarcomeric myosin heavy chain (MF20) were obtained from the Developmental Studies Hybridoma Bank (University of Iowa, Iowa City, IA). The secondary antibody used was AF488-labeled goat anti-mouse IgG (Invitrogen). Fluorescent images were captured using an Axiocam MRM high resolution camera on an Axiovert 200M microscope (Carl Zeiss Microimaging).
Results
CSD determination
Twelve frogs were used for the determination of CSD in X. laevis (Table 1; Fig. 4). Central (diaphyseal) segments of the posterior tarsus bone were excised, ranging from 1.5 to 8 mm in length, with the length of intact tarsus being 12 mm. Three months postsurgery, micro-CT images revealed that the small gaps in the CSD determination group had significant re-ossification filling in the gap (Fig. 4a), whereas the large gaps demonstrated no ossification in the gap, and slight re-ossification near the cut ends of the tarsus bone (Fig. 4b). As expected from the micro-CT images, the histological sections demonstrated a substantial skeletal repair involving the development of cartilage, woven bone, and cancellous bone in 8 out of 12 frogs. The CSD was calculated as 35.2% by averaging the defect sizes of the four cases, which failed to regenerate, as listed in Table 1. In samples with defects greater than critical size (4 out of 12), the defect areas were occupied with fibrotic scar tissue and muscle bundles (Fig. 5). The identity of muscle bundles was confirmed by staining for sarcomeric myosin heavy chain (not shown). Cartilage formation and ossification, if any, was limited exclusively to the cut ends of the tarsus bone.
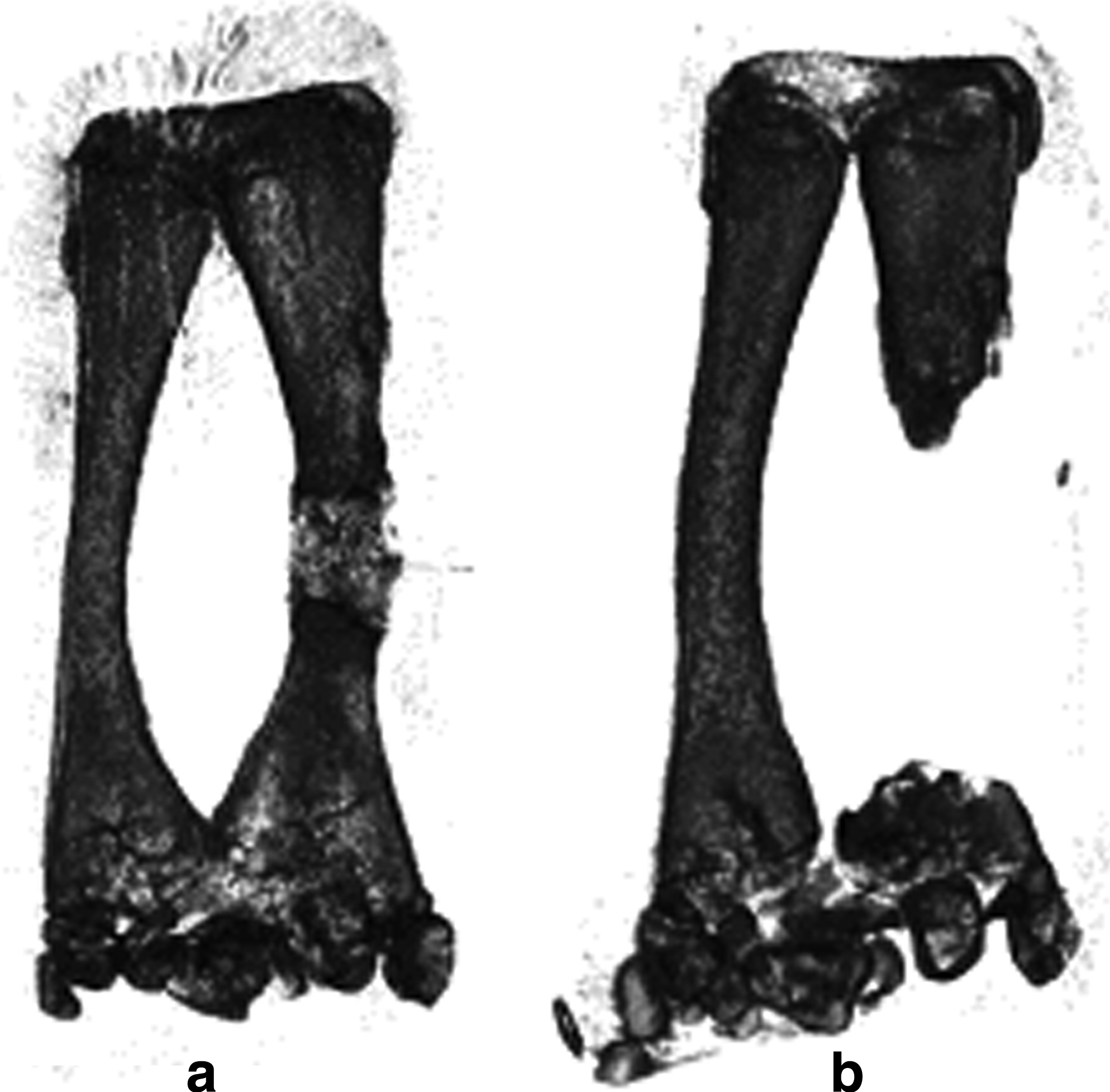
Determination of the critical-size defect for Xenopus tarsus bone. Significant re-ossification of the gap is observed after removal of a short (14.9% of total length) segment in
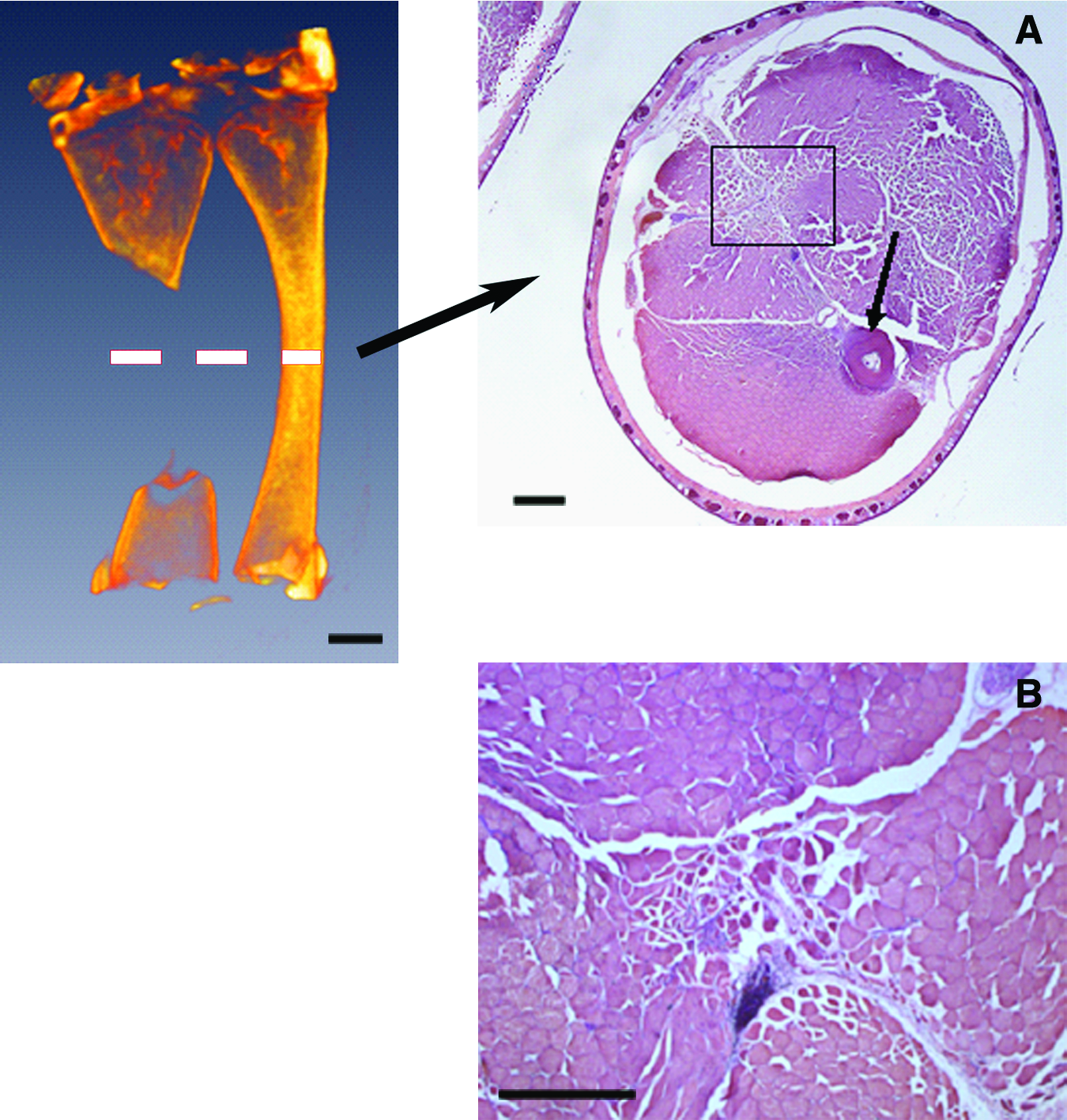
Histology of critical size gap samples. Dashed line in micro-CT image indicates the approximate plane of the displayed histological sections. Left: Micro-CT image showing approximate location of histological panels to the right.
Summary of critical-size gap determination samples. Gap size is given as a percentage of intact tarsus length. Gap tissue formation is listed as either scar or cartilage, depending on the type of tissue found at the center of the gap at 3 months postsurgical removal of tarsus bone segments. Rows highlighted in gray are frogs that did not generate cartilage to fill the gap and bridge the cut ends of the bone, thus defining the critical size for proper tarsus defect repair.
Implantation of control scaffolds
The control scaffold group (scaffolds soaked only in distilled water) consisted of nine frogs. Surgical manipulation in six of these resulted in CSD. Micro-CT and histological analysis revealed that in all six frogs (100%), the segmental defects implanted with control scaffolds did not generate significant cartilage formation or undergo ossification. Instead, the gap was filled with fibrotic scar tissue and muscle, as shown in Figure 6c and f. As in the CSD determination group, cartilage formation and re-ossification was limited to the areas immediately adjacent to the cut ends of the tarsus bone (Fig. 6a, b) The scaffold was observed adjacent to the defect region, and was filled with a scar tissue and blood vessels (Fig. 6d, e). These cases further confirmed that the CSD size was 35% or greater of the tarsus length. The presence of the scaffold alone did not promote cartilage or bone formation in the tarsus gap. The experimental results for this group are summarized in Table 2.
Histology of subjects implanted with control scaffolds. Red dashed lines in micro-CT images indicate the approximate plane of the displayed histological sections.
Summary of results for frogs implanted with control scaffolds. Gap size is given as a percentage of intact tarsus length. Gap tissue formation is listed as either scar or cartilage, depending on the type of tissue found at the center of the gap at the time of sample collection. Defects >35% of intact tarsus length are considered critical-size defects.
Implantation of scaffolds soaked in BMP-4 and VEGF
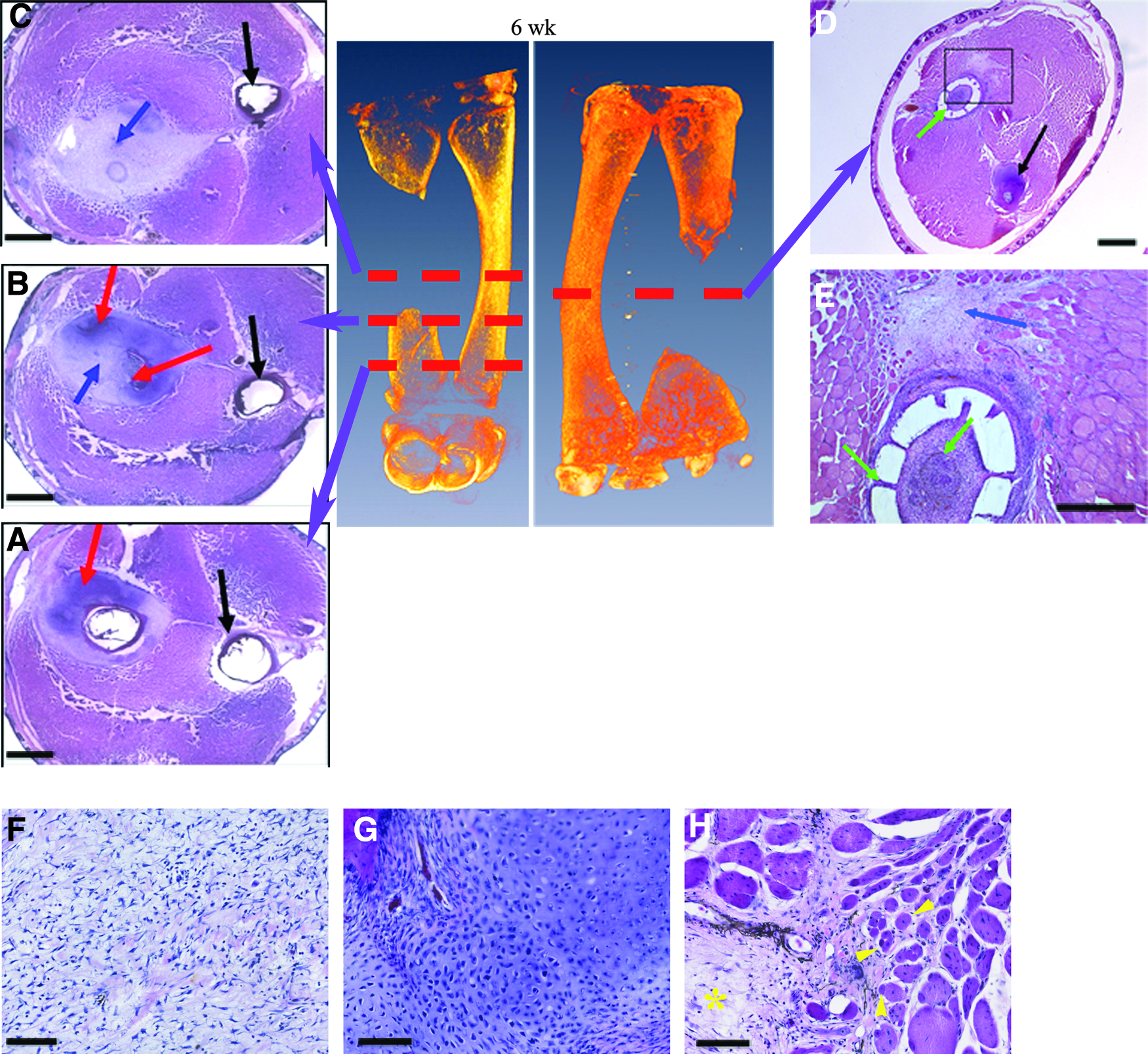
Nineteen animals were implanted with scaffolds loaded with BMP-4 and VEGF. Micro-CT images revealed that 11 of these surgical manipulations generated defects greater than CSD, whereas the other eight surgeries generated no CSD. In 8 of 11 (73%) animals with CSD, the micro-CT and histological analysis revealed that the defect site developed a rod of cartilage extending across the gap, joining the two cut ends of the bone. Moreover, by 3 months postsurgery, foci of ossification were observed in the cartilage rod, and by 6 months, ossification was observed occurring throughout the entire length of the cartilage rod. In all cases the scaffolds were moved aside, leaving the defect space open for regeneration. Figures 7 and 8 display the micro-CT and histological observations from the animals implanted with growth factor-treated scaffolds. In addition to cartilage rod formation bridging the surgically generated bone gap and subsequent ossification of the cartilage rod, we also observed regenerating diaphyseal skeletal muscle attachments and interosseous ligament attachments to the regenerating skeletal element in growth factor-treated cases by 3 months (Fig. 7f–h). Immunostaining of sections with an antibody to β1 integrin show enhanced staining for the integrin at junctional attachments of diaphyseal muscles to the unaltered right tarsus segment (Fig. 7g). Localization of this integrin is enhanced an myotendinous junctions.60,61 Staining of the regenerating tarsus segment (Fig. 7h) shows muscle fibers making attachments to the connective tissue surrounding the regenerating skeletal element. The experimental results for this group are summarized in Table 3.

Histology of subjects implanted with growth factor-treated scaffolds. Red dashed lines in micro-CT images indicate the approximate plane of the displayed histological sections
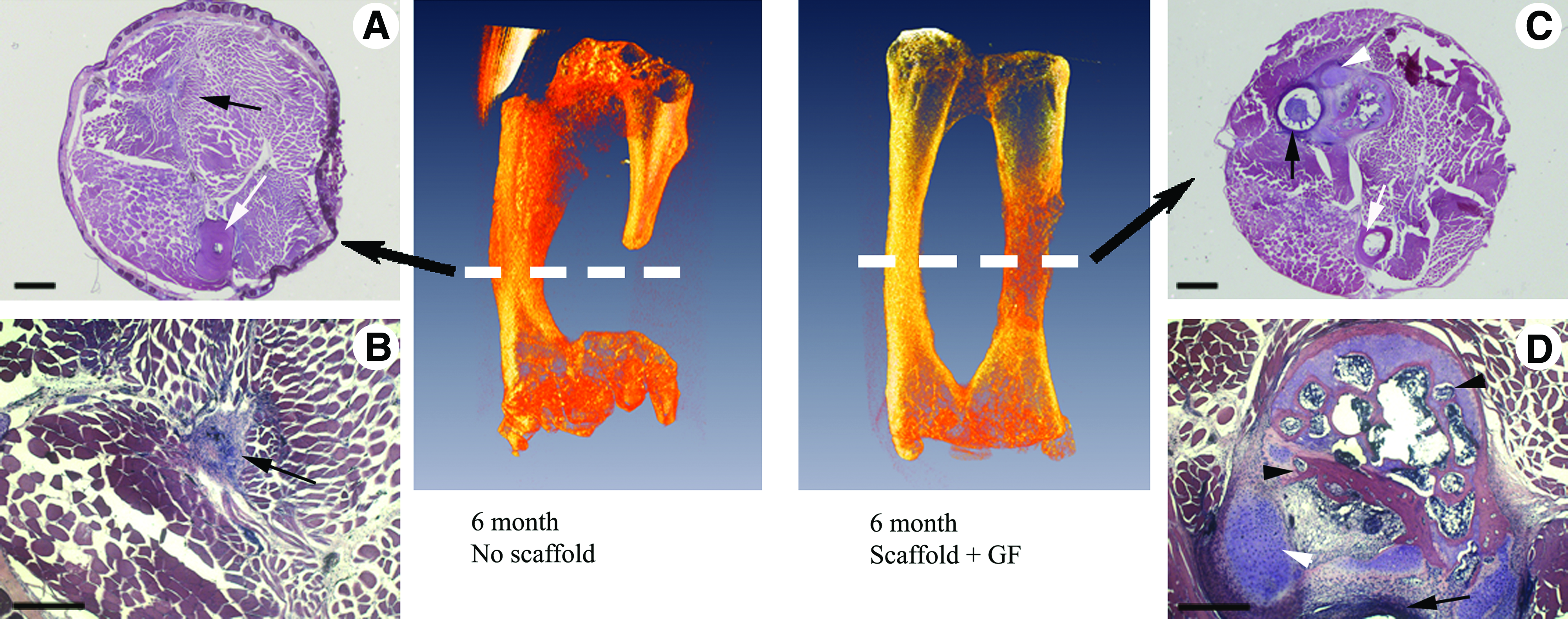
Histology of frog tarsus segments 6 months post-surgery. Dashed white lines in micro-CT images indicate the approximate plane of the displayed histological sections.
Summary of results for frogs implanted with scaffolds loaded with BMP-4 and VEGF. Gap size is given as a percentage of intact tarsus length. Gap tissue formation is listed as either scar or cartilage, depending on the type of tissue found at the center of the gap at the time of sample collection. Defects >35% of intact tarsus length are considered critical-size defects.
BMP, bone morphogenetic proteins; VEGF, vascular endothelial growth factors.
Discussion
Animal model
As shown in our study, adult Xenopus, like mammals, are unable to repair fractures across critical-size bone gaps. This similarity to the mammalian response to these injuries gives us the opportunity to use this animal as a model system to investigate scaffold and growth factor-assisted repair of CSD. While mammalian models have been the standard for experiments using tissue engineering approaches to repair large bone defects, a Xenopus tarsus system has some advantages. First, compared to large mammals such as sheep and pigs (and even smaller mammalian systems such rats or mice), general animal care, surgery, and postoperative care for frogs are simple and inexpensive. Second, because of the dual skeletal element structure of the tarsus bone, we avoid the surgical and logistical complications of an external stabilization device, which would be absolutely required in other experimental systems. Elimination of this requirement is potentially important, as there have been reports of fixation plates affecting tissue repair. 22 Studying a dual skeletal element in an organism that is primarily aquatic avoids these issues. Finally, the size, ease of use, affordability, and bone biology similarity to mammals 57 makes adult Xenopus a very convenient animal model for use as a testing system for screening scaffold designs, growth factor choices, and scaffold–growth factor pairings to identify potentially successful combinations before attempting their use in larger, more expensive, and more experimentally complex mammalian animal models. Additionally, it will eventually be possible to perform experiments using cell-seeded scaffolds, as we have been able to isolate and culture periosteal cells and satellite cells from Xenopus bone and muscle (Milner, D.J., and Cameron, J.A., unpublished results).
Potential drawbacks to our system include comparatively slow injury repair seen in frogs, likely due to the slower overall metabolic rate in amphibians compared to mammals, which may necessitate longer completion times for some experiments. Additionally, there is a lack of detailed molecular characterization of the fracture repair process in Xenopus. We are currently performing studies designed to characterize the temporal and cell-type-specific expression of growth factors, extracellular matrix molecules, and transcription factors in Xenopus fracture repair to determine the extent of similarity of the fracture repair process in frogs to that in mammalian species and possible limitations of the model for studying mammalian systems.
In this work, we have focused on a single scaffold type and a single growth factor combination to establish this small animal model and demonstrate its regeneration potential. We show that this model can be conveniently used to assess or compare different parameter choices for promoting regeneration. Then, after such initial screening one can proceed with experiments using more complex mammalian models.
Critical size gap and intercalary regeneration
The first step in our study was to determine the size of the CSD for Xenopus laevis. We determined the size of the CSD to be at least 35% of the length of the whole tarsus, based on the data from 12 animals. This value is higher than those reported for mammals, which are about 20% depending on species. Unfortunately, most articles on long bone critical-size defect repair do not explicitly report or determine the critical size of defect before their studies, even though this information is of paramount importance. We found that 73% of the animals implanted with a growth factor-loaded scaffold bridged the CSD with cartilage, which was then ossified. This was a significant improvement compared to the 0% of the control group that successfully bridged the gap. Further, in addition to the intercalary regeneration repair of the skeletal element, the regenerated structure made connections with attached skeletal muscle and the interosseous ligament, as shown in Figure 7.
In this study we took an alternate approach to regeneration across CSDs in long bone. Instead of attempting to promote regeneration by direct bone deposition on a scaffold, we induced the intercalary regeneration of the missing bone by implanting a biocompatible scaffold loaded with growth factors specifically selected to induce cartilage to form in the defect, bridge the bone gap, and then convert this cartilage template to bone. We selected BMP-4 and VEGF as the growth factors to be used because BMP-4 is essential for the development of cartilage templates and is induced early in fracture repair, and VEGF induces blood vessel formation and is critical for the conversion of hypertophic cartilage to bone.24,25,50,56 In addition to these factors, there are likely other growth factors that could be used that would lead to successful outcomes.50–52
In our study, the scaffold had freedom to move within the constraints imposed by the surrounding tissues because we did not fix it within the gap zone or glue it to the cut ends of the bone. Thus, instead of acting as a support and a template for direct bone deposition, the scaffold functioned as a 3D carrier that released the growth factors along the length of the defect region. Our in vitro measurements suggest that scaffolds release growth factors within the first few hours after implantation, with an initial burst release of growth factors. We speculate that in vivo, the growth factors may stay longer in the scaffold and are more slowly released into the defect site, but we currently have no measurements to support this statement. More complex scaffold designs leading to longer release times for growth factors, or sequential release of factors, could potentially further improve regeneration outcomes.
The enhanced presence of the growth factors provided by our scaffolds in the initial stages of repair immediately post-injury is sufficient to trigger the processes that lead to bone formation. For BMP-4, this is not surprising, as previous studies have shown the important role BMP-4 plays in endochondral ossification and in the early stages of fracture repair,50,54,55 with BMP-4 expression induced as early as 12 h postfracture in the periosteum of mice. 54 Of further interest is the role played by VEGF. The results reported here using our Xenopus system seem to support the findings of Peng et al. that early ectopic production of BMP-4 and VEGF promote healing of a murine bone gap by endochondral ossification, and that in addition to its important role in the later stages of endochondral ossification, VEGF is important for the early stages of this process as well, acting synergistically with BMP-4. 53 In our system, the growth factors have been released from the scaffold rapidly after implantation, so it seems highly unlikely that the VEGF delivered by our scaffold is directly influencing the latter stages of endochondral ossification (blood vessel invasion and chondrocyte hypertrophy) that rely on VEGF activity. 56 It will be of interest to perform experiments in our system comparing bone repair in the presence of scaffolds loaded with BMP-4 and VEGF to repair with scaffolds loaded with either BMP-4 or VEGF alone to determine if these growth factors are acting synergistically to enhance bone healing, as reported by Peng and coworkers. 53
Conclusions
We have established that the adult X. laevis tarsus bone can serve as a good model for studying the effect of regeneration-promoting growth factors on the process of bone CSD repair. A diaphyseal gap comprising 35% or greater in X. laevis posterior tarsus is the CSD that cannot be bridged by natural fracture repair. There was a striking difference in the reaction of the limb tissues to the growth factor-treated versus the control scaffolds. The BMP-4 + VEGF-treated scaffolds caused hyaline cartilage regeneration across the gap that was followed by ossification. In the control scaffold samples, cartilage formed only at the injury site and scar tissue grew in the gaps. We used our scaffold in an atypical way, as a growth factor delivery mechanism rather than as a template for bone formation. Our future work will use this newly developed small animal model to study different scaffold materials, geometries, growth factor combinations, and release profiles for their effects on long bone CSD repair, and we will develop Xenopus stem cell cultures for use in scaffold-assisted repair. We will also attempt to use live-imaging technology to follow cartilage and bone formation in living individuals over time for a more thorough analysis of regeneration.
Footnotes
Acknowledgments
We thank Paul Miller for his assistance in performing surgeries and Deepika Chitturi for literature searches and valuable discussions. We gratefully acknowledge support of W.M. Keck Foundation (J.A.C. and D.L.S.) and Gauthier Exploratory Fund and LDRD grant by Livermore National Lab (N.F.), Grainger Foundation and National Science Foundation (CMMI 0927909- Dr. Ken Chong) (I.J.), and National Science Foundation (ECCS-1002351- Dr. Rajinder Khosla) (N.F., I.J., and J.A.C.)
Disclosure Statement
No competing financial interests exist.