
Abstract
Complete and predicable regeneration of complex periodontal structures, which include cementum, periodontal ligament (PDL), and alveolar bone, has been a great challenge for periodontal researchers. It is generally believed that human PDL from the root surface contains stem cells (r-PDLSCs), which can enhance cementum/PDL-like tissues regeneration in vivo. In this work, PDL was found to possess asymmetrically distributed stem cells observed by long-term bromodeoxyuridine (BrdU) labeling. Putative stem cells from human PDL on the alveolar bone surface (a-PDLSCs) were then isolated and characterized. It was shown that a-PDLSCs exhibited strong proliferation capability and expressed high percentages of mesenchymal stem cell markers. Comparatively, a-PDLSCs had higher multilineage differentiation potential than r-PDLSCs with regard to both osteogenic and adipogenic differentiation. Alkaline phosphatase activity and the expression of mineralization-related markers of a-PDLSCs were also higher than those of r-PDLSCs. In vivo, a-PDLSCs could regenerate bone/PDL-like structures and repair critical-size defects created in calvarial bone of NOD/SCID mice. Autologous PDLSC-mediated periodontal regeneration showed that a-PDLSCs could accomplish reconstruction of alveolar bone more perfectly than r-PDLSCs. Our data suggest that PDLSCs may have quite different characteristics depending on locations. a-PDLSCs may take a synergistic effect with r-PDLSCs in periodontal regeneration.
Introduction
The heterogeneous nature of fibroblasts that populate the periodontium, together with the complex interactions that occur between hard and soft tissues during periodontal wound healing, is a poorly understood concept which is of paramount importance in periodontal regeneration.7–11 The key of complete periodontal regeneration is dependent, to a great extent, on the ability of cells to reconstitute the mineralized tissues of cementum and bone. It has been well recognized that repair of periodontal tissue appears to involve progenitor cells, which are present in the PDL, capable of forming fibroblasts, osteoblasts, and cementoblasts. 12 Up to now, it is still not entirely defined whether a single progenitor cell type can give rise to all the tissues of the periodontium. In other words, it remains unclear whether cementoblasts and osteoblasts have a common precursor in the PDL. Although the identification of stem cells within the human PDL represents a significant development in the progress toward predictable periodontal regeneration, in a heterotopic transplantation model of immunodeficient mice, PDLSCs are merely capable of differentiating into cementoblast-like cells and forming cementum/PDL-like tissues in vivo. 3 It is generally believed that mesenchymal stem cells are inclined to develop tissues compatible with their origin in an ectopic site. Numerous studies have repeatedly demonstrated that in conditions devoid of any influence derived from relevant cell types and matrix normally present in periodontium in situ,5,13 PDLSCs isolated from PDL on the root surface could recapitulate most of the basic features of cementum/PDL tissue formation when transplanted into immunodeficient mice. Lang's study also indicated that the regenerative capacities of cells originating from different periodontal cell populations were various. 14 PDLSCs from root surface play limited role in alveolar bone regeneration. Therefore, it is imaginable that stem cells derived from PDL on the alveolar bone surface might form bone/PDL-like tissues. In fact, previous studies have pointed out differences between root-related periodontium and that associated with alveolar bone. 15 The tooth-related ligament appeared to contain the cementoblast progenitors and the bone-related ligament osteoblast progenitors.
In the present study, for the first time we postulated that there was a population of stem cells residing in PDL proximal to the alveolar bone surface, which were distinctly different from putative PDLSCs derived from root surface (r-PDLSCs). Meanwhile, r-PDLSCs and bone marrow stem cells (BMSCs) were used as control to evaluate the characteristics of a-PDLSCs and their effect on periodontal regeneration.
Materials and Methods
Histological analysis
To assess the distribution of PDL after extraction, alveolar bone in the molar area of a 4-week-old Sprague-Dawley rat was acquired after the mandibular first molar was extracted as previously reported. 6 Alveolar bone was fixed in 4% paraformaldehyde for 1 day and then decalcified for 10 days in 10% EDTA before embedding in paraffin. The sections of alveolar bone were prepared and stained with hematoxylin and eosin for histological analysis.
Cell culture
All experimental protocols were approved by the Committee of Ethics of Fourth Military Medical University (FMMU) and conducted in accordance with the Guidelines. Eighteen patients aged 19–28 undergoing third molar extractions at the Dental Clinics of the Fourth Military Medical University were included in this study. After extraction, PDL was gently scraped from the middle part of the alveolar socket wall (surface of alveolar bone) with curette. The tissue was then washed repeatedly with sterile phosphate-buffered saline (Gibco-BRL) before bone scraps were removed by tweezers. The other part of PDL was gently scraped from the middle part of the root as previously reported.16,17 Both parts of PDL were minced into small pieces and placed into six-well culture plates (Costar). Bone marrow that had been acquired from the residual sample of the fresh and healthy bone marrow after the bone marrow aspiration in the hematology department, Xijing Hospital, the Forth Military Medical University, was washed twice with sterile phosphate-buffered saline and cultured in six-well culture plates. All primary tissues or cells were cultured with α-minimum essential medium (α-MEM; Gibco BRL) supplemented with 10% fetal bovine serum (FBS), glutamine 0.292 mg/mL, penicillin 100 U/mL (Gibco BRL), streptomycin 100 μg/mL (Gibco BRL), and ascorbic acid 100 μM/L (Sigma). The cultures were incubated at 37°C in a humidified atmosphere of 5% CO2.
To isolate putative stem cells, single-cell suspensions (1 × 104 cells) were seeded into 90 cm culture dishes (Costar) as previously described and cultured in α-MEM containing 10% FBS.3,16,17 Multiple colony-derived PDLSCs at two to five passages were used. For each experiment, a-PDLSCs, r-PDLSCs, and BMSCs at the same passage were used.
Proliferation ability assay
a-PDLSCs, r-PDLSCs, and BMSCs at a density of 2 × 103 cells/well were plated in 96-well culture plates for MTT assay. At 8 consecutive days, the optical density (OD) values were determined by a multiplate reader (Bio-Tek) at wavelength of 490 nm.
To assess colony-forming efficiency, 2 × 103 a-PDLSCs, r-PDLSCs, and BMSCs were under standard culture conditions for 10 days. Then, all cultures were fixed with 4% formalin and stained with 0.1% toluidine blue. Aggregates of over 50 cells were counted as colonies. Each experiment was repeated at least thrice.
After 4-day culture, single-cell suspensions of a-PDLSCs, r-PDLSCs, and BMSCs were prepared for cell cycle analysis as previously reported.16,18
Cell phenotype analysis
Cell phenotype was analyzed by flow cytometry profile of CD34 (Chemicon), CD44, CD90, CD45, CD14, CD31, CD105, CD29 (eBioscience), STRO-1, and CD146 (R&D Systems) according to the manufacturer's protocol.17,18
a-PDLSCs, r-PDLSCs, and BMSCs cultured with α-MEM containing 5% FBS at passage 4 were harvested for immunofluorescence staining. After incubated with primary antibodies including goat anti-osteocalcin (OCN, 1:100; Santa Cruz Biotechnology), rabbit anti-collagen type I (COL I; 1:100; Millipore) for 2 h, cells were incubated with rabbit anti-goat or goat anti-rabbit secondary antibodies of FITC (1:200; Santa Cruz Biotechnology) for 45 min. The images were collected by DP controller (Olympus) and DP manager.
Labeling cells with bromodeoxyuridine in vivo and Immunohistochemistry analysis
To evaluate the distribution of the proliferating cells in PDL, we injected a single dose of bromodeoxyuridine (BrdU) (50 mg/kg; Sigma) twice daily into four 4-week-old Sprague-Dawley rats at 4 consecutive days and killed the animals 1 month after injection. Bilateral maxillary molars and periodontal tissues were fixed and decalcified as just described. Total eight samples were incubated with mouse anti-BrdU antibody (1:200; Millipore) at 4°C overnight and then incubated with goat anti-mouse secondary antibodies that were conjugated to Rhodamine (1:200; Chemicon) for 45 min at 37°C. The images were collected by the DP controller (Olympus) and the DP manager.
Osteogenic and adipogenic differentiation
The osteogenic and adipogenic induction medium were prepared as previously described.17,19 After 2 weeks of osteogenic induction and 3 weeks of adipogenic induction, the cells were stained with alizarin red (pH 4; Sigma) and fresh oil red O solution (Sigma) for 15 min at room temperature. To determine the spontaneous mineralization potential, cells were plated in 75 cm2 culture flask and cultured with standard culture medium (α-MEM supplemented with 5% FBS). Six weeks later, cells were stained with alizarin red (pH4; Sigma) for 15 min at room temperature. The nodule and lipid area were quantitatively measured by an image analysis system (Image-Pro Plus 5.0; Media Cybernetics) as previously described. 18
Alkaline phosphatase activity assay
Single-cell suspensions of a-PDLSCs, r-PDLSCs, and BMSCs were seeded at a density of 2 × 103 cells/well into 96-well plates. After 7 and 14 days of culture, alkaline phosphatase (ALP) activity was detected with a commercially available assay kit (Zhongsheng Co.). 17
Real-time PCR
Total cellular RNA was isolated from 5 × 105 r-PDLSCs, a-PDLSCs, and BMSCs with Trizol reagent (Invitrogen). First-strand cDNA syntheses were performed as previously described. 20 The Quantitect Sybr Green Kit (Toyobo) and ABI Prism 7700 Sequence Detection System (Applied Biosystems) were used. Primer sets in this study are shown as below: Runx2 sense, 5′-CCCGTGGCCTTCAAGGT-3′, and Runx2 antisense, 5′-CGTTACCCGCCATGACAGTA-3′; COL I sense, 5′-CCAGAAGAACTGGTACATCAGCAA-3′, and COL I antisense, 5′-CGCCATACTCGAACTGGAATC-3′; OCN sense, 5′-AGCAAAGGTGCAGCCTTTGT-3′, and OCN antisense, 5′-GCGCCTGGGTCTCTTCACT-3′; ALP sense, 5′-GGACCATTCCCACGTCTTCAC-3′, and ALP antisense, 5′-CCTTGTAGCCAGGCCCATTG-3′; collagen type III (COL III) sense, 5′-CTGGTCAGCAGGGTGCAATC-3′, and COL III antisense, 5′-TGGTTCCATCTTGCCAGGAG-3′; scleraxis (SCX) sense, 5′-AGCCCAAACAGATCTGCACCTT-3′, and SCX antisense, 5′-TTCTGTCGCGGTCCTTGCT-3′. The gene expression of GAPDH was used as a reference gene expression in all applications.
Differentiation assay in vivo
All procedures were performed under institutionally approved guidelines for the use of animals in research. Total six 6-week-old NOD/SCID mice (purchased from the Laboratory Animal Research Centre of Fourth Military Medical University) were used for allogenic transplantation. Transplantation procedures were performed as previously reported.16,17 About 5 × 106 a-PDLSCs, r-PDLSCs, and BMSCs at passage 4 were mixed with 15 mg ceramic bovine bone powders (Research and Development Center for Tissue Engineering, Fourth Military Medical University, Xi'an, Shaanxi, China), covered by fibrin gel (Beixiu Co.), and then transplanted into the subcutaneous pockets on the dorsal surface of NOD/SCID mice. A total of 18 grafts were implanted. Each mouse received a set of three different grafts. The transplants were recovered at 8 weeks post-transplantation, fixed with 4% paraformaldehyde, decalcified with buffered 10% EDTA, and then prepared for hematoxylin and eosin staining. The mineralization area was quantitatively measured by an image analysis system (Image-Pro Plus 5.0; Media Cybernetics).
Repairing of critical-size defects in calvarial bone
The calvarium of 6-week-old NOD/SCID mice was exposed. The critical-size defect (6 mm in diameter) was surgically created (Fig. 1A). Twelve 6-week-old NOD/SCID mice were divided into three groups at random. r-PDLSCs and a-PDLSCs with fibrin gel as carrier were made into flat-plate type of grafts and implanted into the defects, respectively. Fibrin gel alone was used as a control.
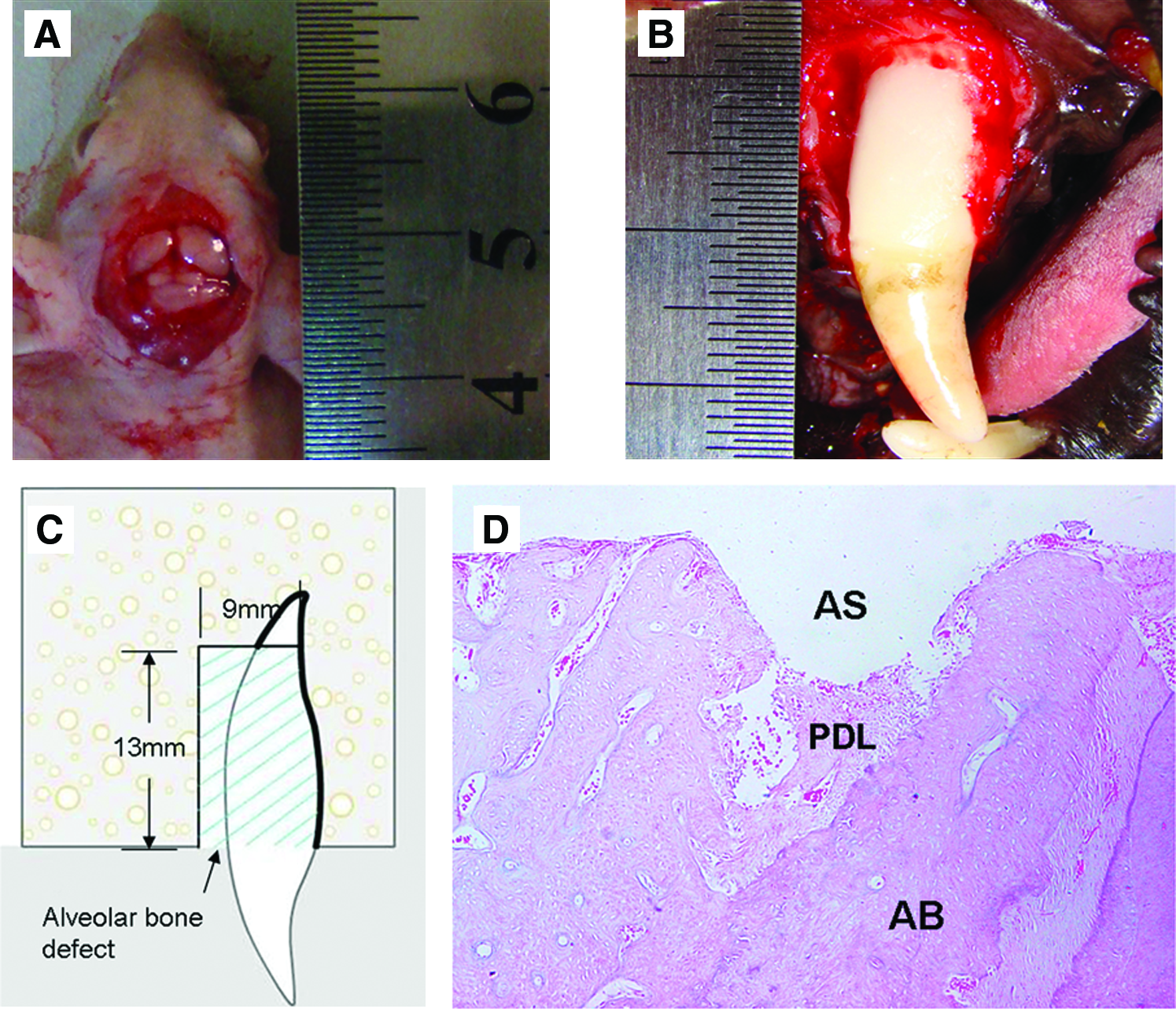
Artificial defect models and histological analysis of PDL in situ.
The animals were observed from 0 to 10 weeks by microsopic computerized tomography (CT) (Inveon MM Gantry-STD; Siemens). The percentage of new bone area in the total defect area was taken to assess the repairing capability. The new bone formation area was quantitatively measured by an image analysis system (Image-Pro Plus 5.0; Media Cybernetics). After 10 weeks, histological sections of the defects were prepared and stained with hematoxylin and eosin for histological analysis.
Repairing of periodontal defects
The anterior teeth of six 20-month-old Beagle dogs were extracted for the isolation of a-PDLSCs and r-PDLSCs. Alveolar bone and cementum in mesial and buccal region of bilateral maxillary canines were removed to create experimental alveolar bone defects (Fig. 1B, C). Autologous a-PDLSCs with fibrin gel were transplanted into left maxillary periodontal defect region; r-PDLSCs with fibrin gel or fibrin gel alone were transplanted into right maxillary periodontal defect region. After surgery, the defects were identified by CT analysis from 0 to 10 weeks.
Statistical analysis
All data were expressed as mean ± SD and analyzed by two-tailed unpaired Student's t-test using SPSS software (version 12.0, SPSS). p-Values less than 0.05 were considered statistically significant.
Results
Distribution of PDL
In the rat alveolar socket, there was partial PDL remaining on the alveolar bone surface after extraction (Fig. 1D).
Morphological feature and proliferative potential
Human r-PDLSCs, a-PDLSCs, and BMSCs all showed a typical fibroblast-like spindle appearance (Fig. 2A–C) and had the ability to form adherent clonogenic cell clusters (Fig. 2D–F). Meanwhile, there was a statistically significant difference with PDLSCs having an increased colony-forming unit behavior as compared with BMSCs (Fig. 2G).

Morphological characteristic and colony-forming capability. r-PDLSCs
The consecutive test for 8 days indicated that a-PDLSCs possessed a higher proliferative potential than BMSCs and the difference became significant from day 4 (Fig. 3A). The results of cell cycle analysis further confirmed that the proliferation abilities of r-PDLSCs and a-PDLSCs were higher than the ability of BMSCs, which was shown by the fact that a-PDLSCs and r-PDLSCs exhibited significantly higher percentages of cells in G2+S phases than BMSCs (Fig. 3B–D).
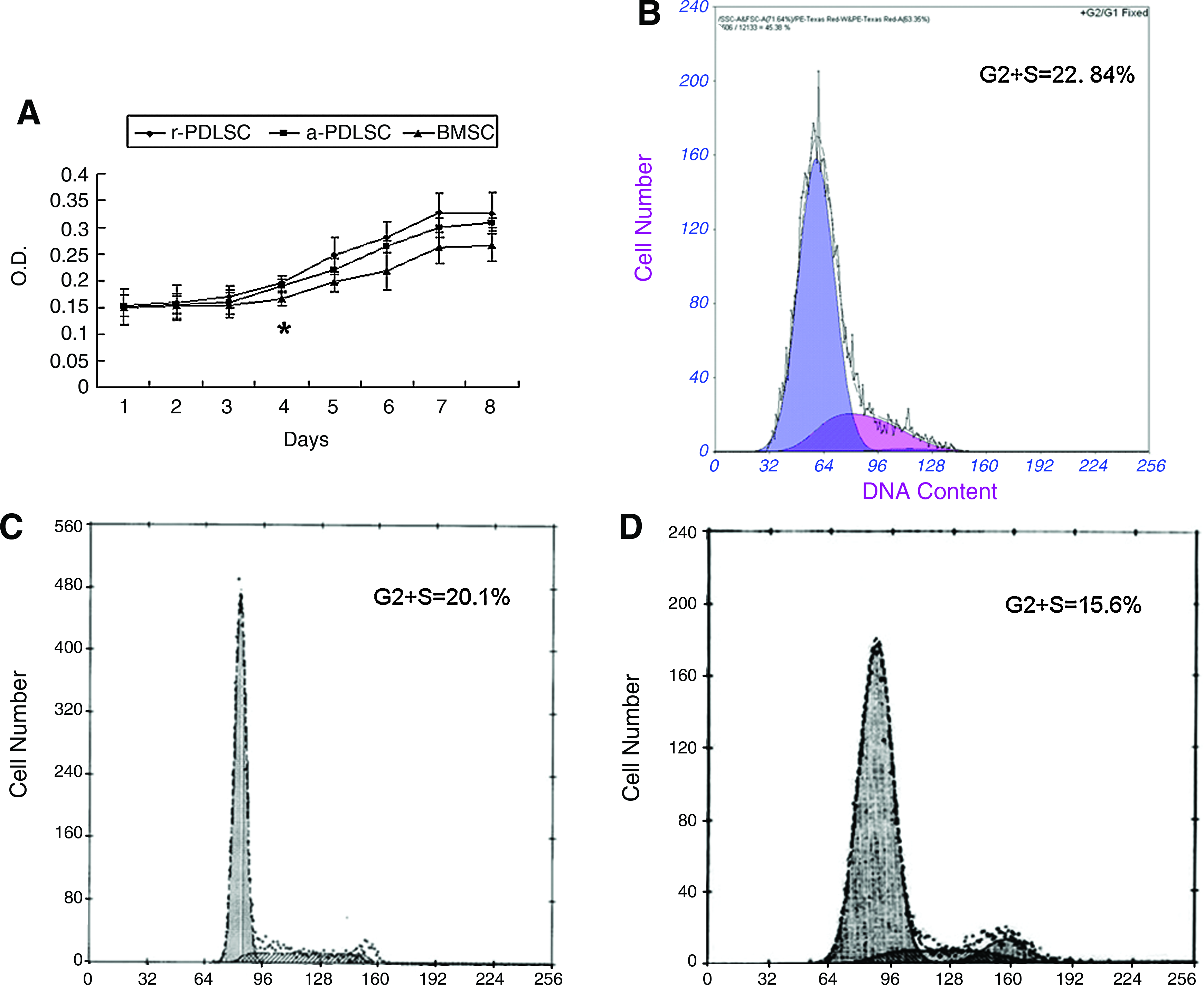
Proliferative potential.
Characterization of epitope profile
Fluorescence-activated cell sorting analysis for epitope profile showed that a-PDLSCs, similar to r-PDLSCs and BMSCs, expressed CD29, CD44, CD90, CD105, CD146, and STRO-1, the putative mesenchymal stem cell markers, at the high levels of 100%, 99.9%, 100%, 99.1%, 12.4%, and 9.2%, respectively, whereas expressed CD31, CD34, CD45, and CD14, the hematopoietic stem cell markers, at the low levels of 1.6%, 2.2%, 2.1%, and 2.4%, respectively (Table 1).
BMSCs, bone marrow stem cells; PDLSCs, periodontal ligament stem cells
Distribution in situ of r-PDLSCs and a-PDLSCs
The expression of BrdU in PDL was observed by immunofluorescent staining in situ. After 1 month, the whole PDL exhibited a high uptake of BrdU. However, the distribution of BrdU-positive cells was asymmetrical in PDL. The BrdU-positive cells in PDL on the alveolar bone surface were more than those in PDL on the root surface (Fig. 4).
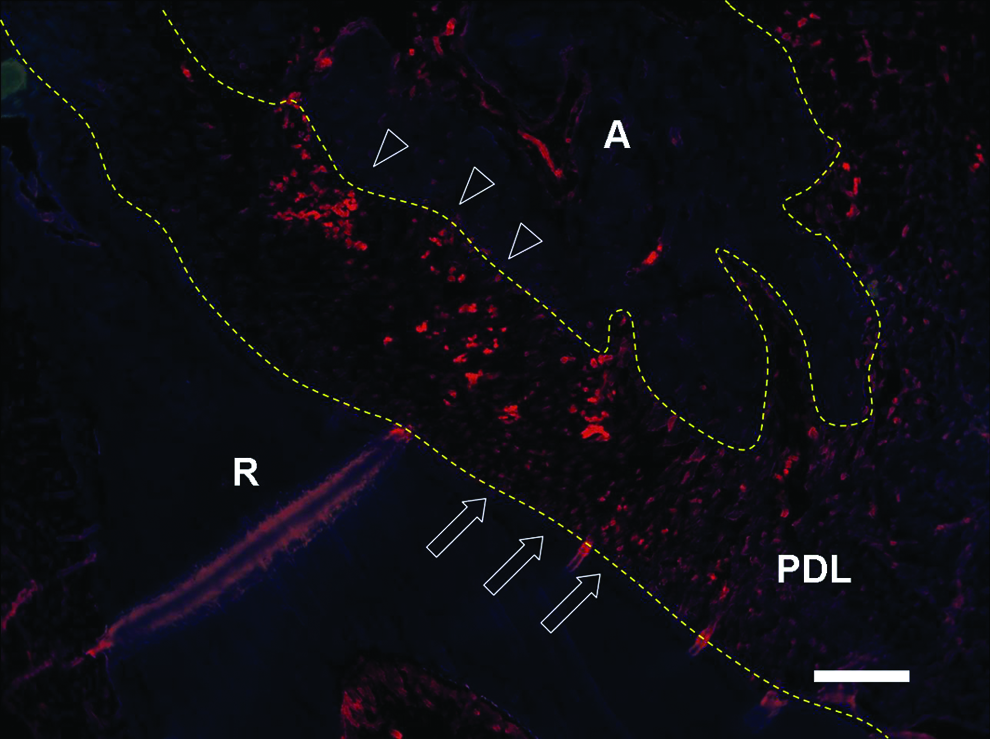
Distribution of BrdU-positive cells in PDL. The bromodeoxyuridine (BrdU)-positive cells in PDL on the alveolar bone surface (triangles) were more than those on the root surface (arrows). A, alveolar bone; R, root. The structure between the two dotted lines indicates PDL. Scale bar represents 500 μm. Color images available online at www.liebertonline.com/tea.
In vitro multipotential ability of PDLSCs
Both human r-PDLSCs and a-PDLSCs expressed mineralization markers OCN and COL I (Fig. 5A–D), suggesting that a-PDLSCs possessed a mineralization potential. After osteogenic induction for 2 weeks, calcified nodules could be observed in all three cell populations (Fig. 6A–C). Quantification of the alizarin red–positive area showed that the calcified nodule per microscopical field was 1.3% in r-PDLSCs, 2.8% in a-PDLSCs, and 4.5% in BMSCs (Fig. 6D). All three cell populations formed oil red O-positive lipid clusters after 3 weeks of adipogenic induction (Fig. 6E–G); and the oil O-positive area per microscopical field was 1% in r-PDLSCs, 5% in a-PDLSCs, and 4.43% in BMSCs (Fig. 6H). Calcified nodules were also formed in all of the populations after 6-week culture in standard medium (Fig. 6I–K). Comparatively, alizarin red–positive area per well of a-PDLSCs was much higher than that of either r-PDLSCs or BMSCs (Fig. 6L). All things considered, a-PDLSCs exhibited much higher osteogenic and adipogenic potential than r-PDLSCs. There were statistically significant differences among all three cell populations on the differentiation potential.
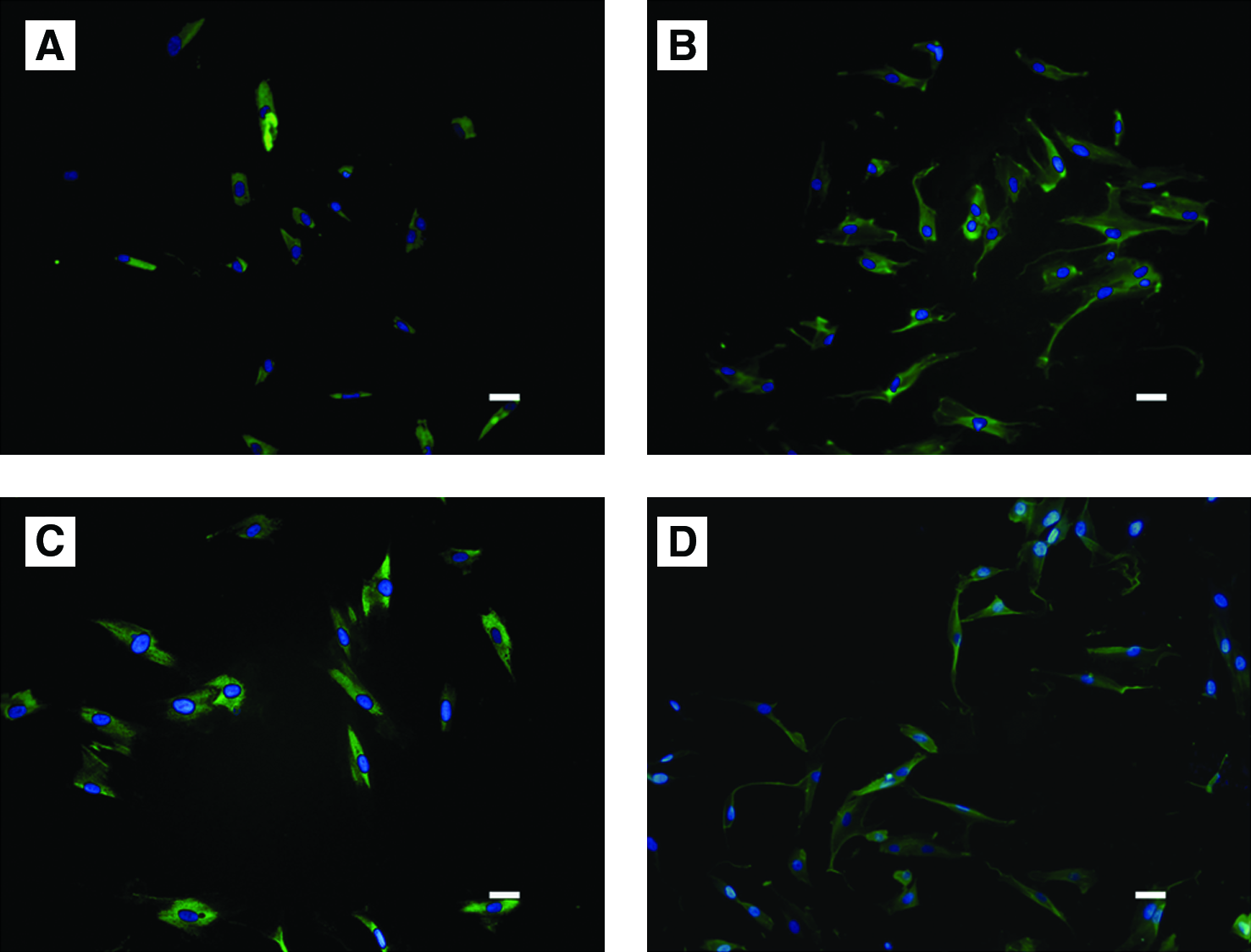
Immunofluorescence staining for r-PDLSCs
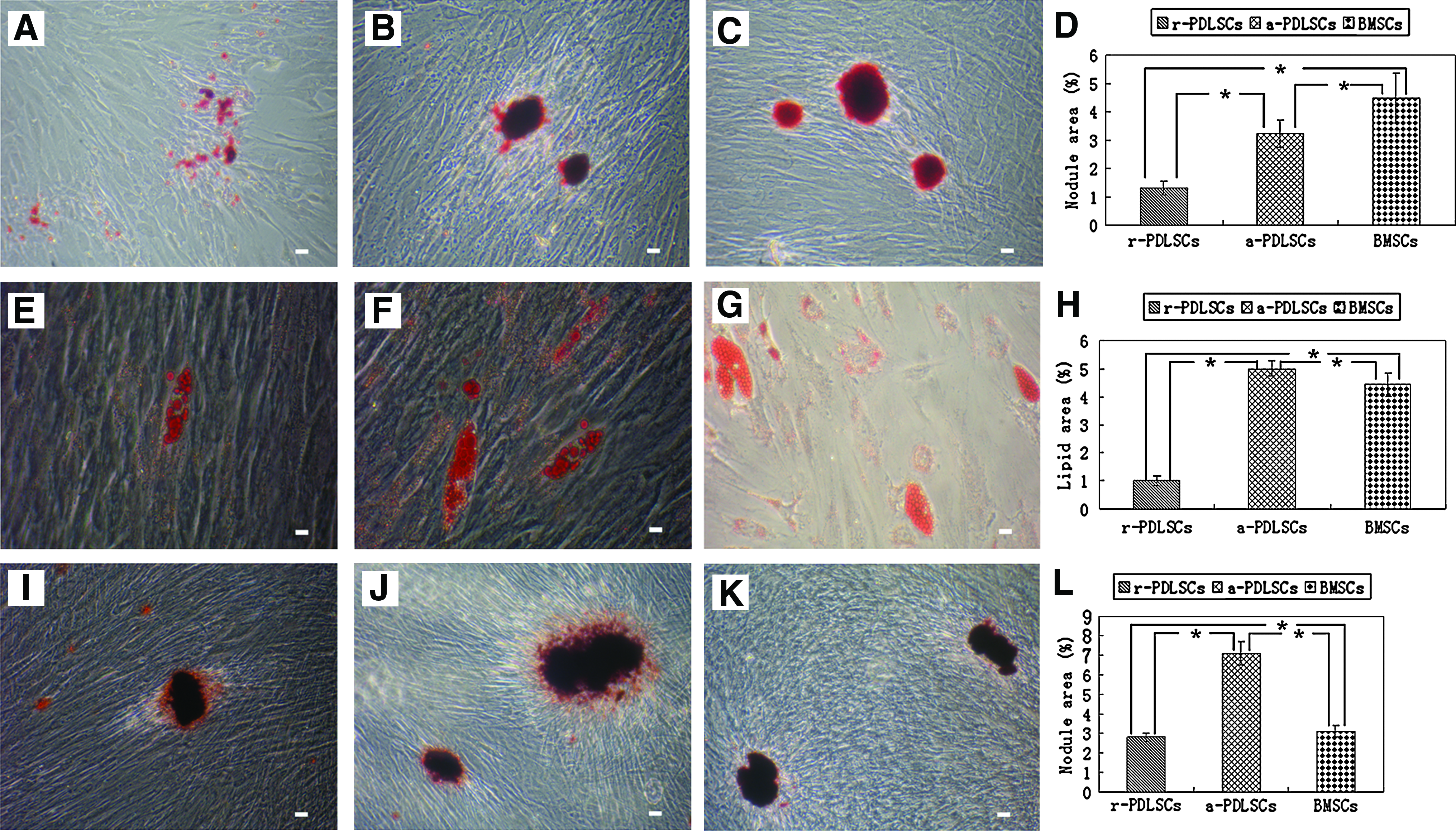
Differentiation potential in vitro. After osteogenic induction for 2 weeks, mineralized deposits could be observed in r-PDLSCs
ALP activity assay indicated that the ALP activity of a-PDLSCs was much higher than that of r-PDLSCs (Fig. 7G). Further research on mRNA relative intensities suggested that a-PDLSCs expressed mineralization markers (ALP, COL I, COL III, OCN, and Runx2) at higher levels in comparison with r-PDLSCs. There was a statistically significant difference between PDLSCs and BMSCs on the expression of SCX, a tendon-specific transcription factor (Fig. 7A–F).
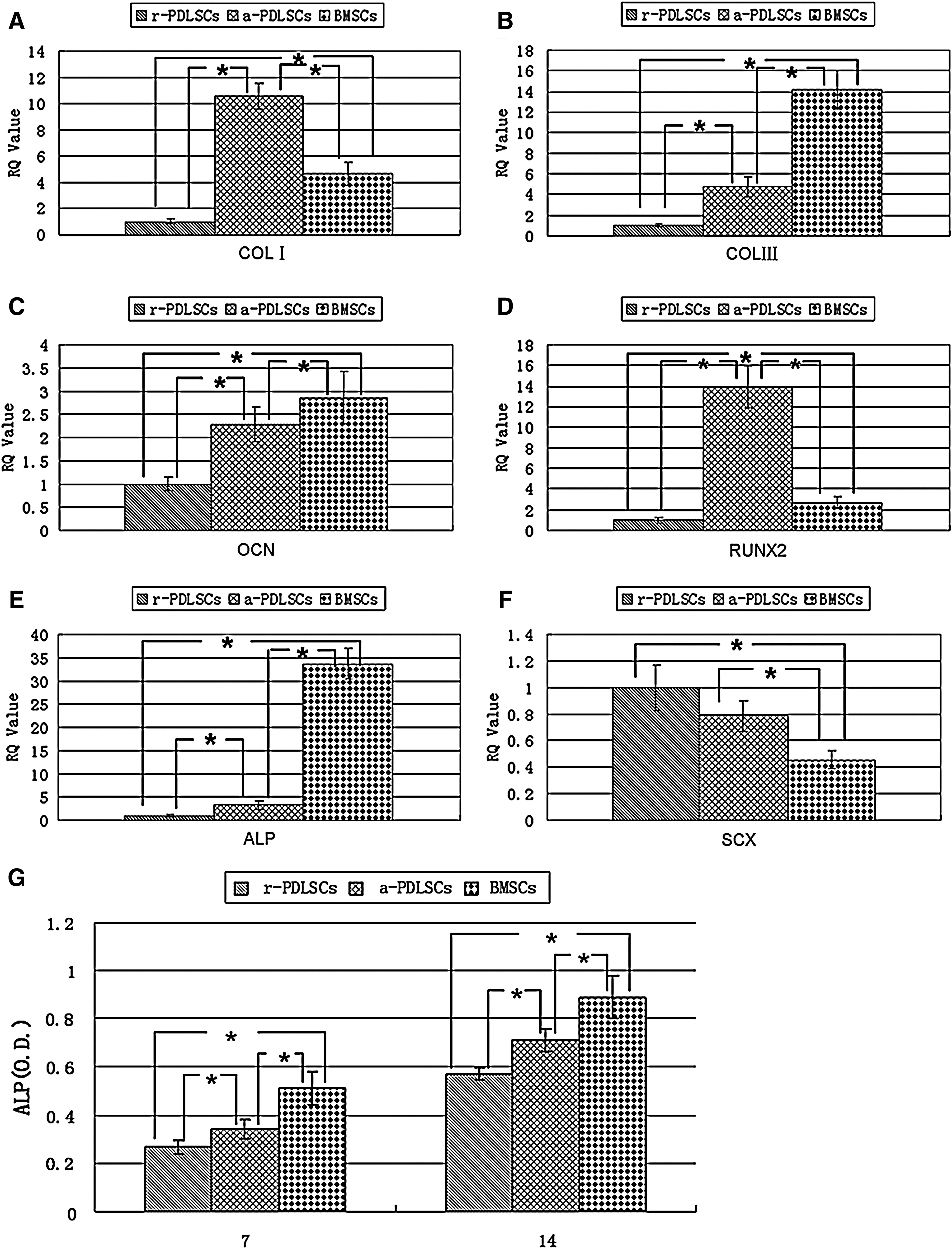
Real-time PCR and ALP activity analysis. The expression of mineralization-related genes including COL I
Differentiation potential in vivo
In vivo, human r-PDLSCs implants generated a thin layer of cementum-like structures on the ceramic bovine bone surfaces and a great amount of PDL-like collagen fibers (Fig. 8A). In contrast, human a-PDLSCs implants generated a large area of bone-like tissues and PDL-like structures (Fig. 8B, C). The area of the mineralized tissues formed by a-PDLSCs was significantly higher than that formed by r-PDLSC, but similar to that formed by BMSCs (Fig. 8D, E). Microsopic CT images of repairing of critical-size defects in calvarial bone showed that there was continued new bone formation in the experimental area. At 5 weeks, new bone formation was observed at the edge of the defect. After 10 weeks, the tissue was further mineralized. A mass of new bone was clearly evident in the marginal and the central region in the r-PDLSCs and a-PDLSCs group. In contrast, very little new bone formation was observed at the defect edge in the control group at 5 and 10 weeks (Fig. 9A). Further quantification of the new bone area showed that it was 21.94% in r-PDLSCs and 31.77% in a-PDLSCs at week 5; whereas 41.39% in r-PDLSCs and 50.72% in a-PDLSCs at week 10 (Fig. 9B). There was a statistically significant difference between the r-PDLSCs and a-PDLSCs group. Histologically, lots of new bone and osteocytes were generated in the surgical defect (Fig. 9C, D). No new bone was observed in the control group (Fig. 9E).
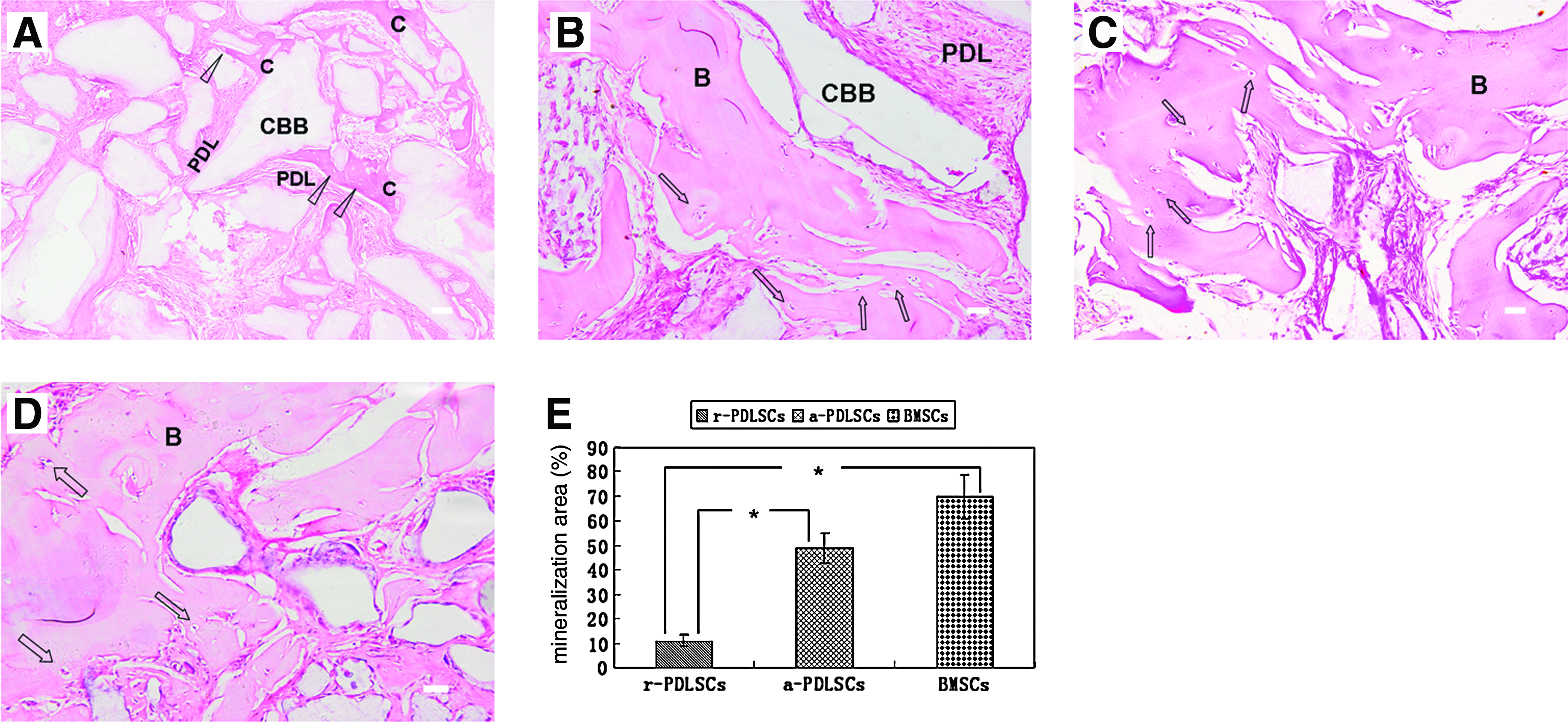
Differentiation potential in vivo.
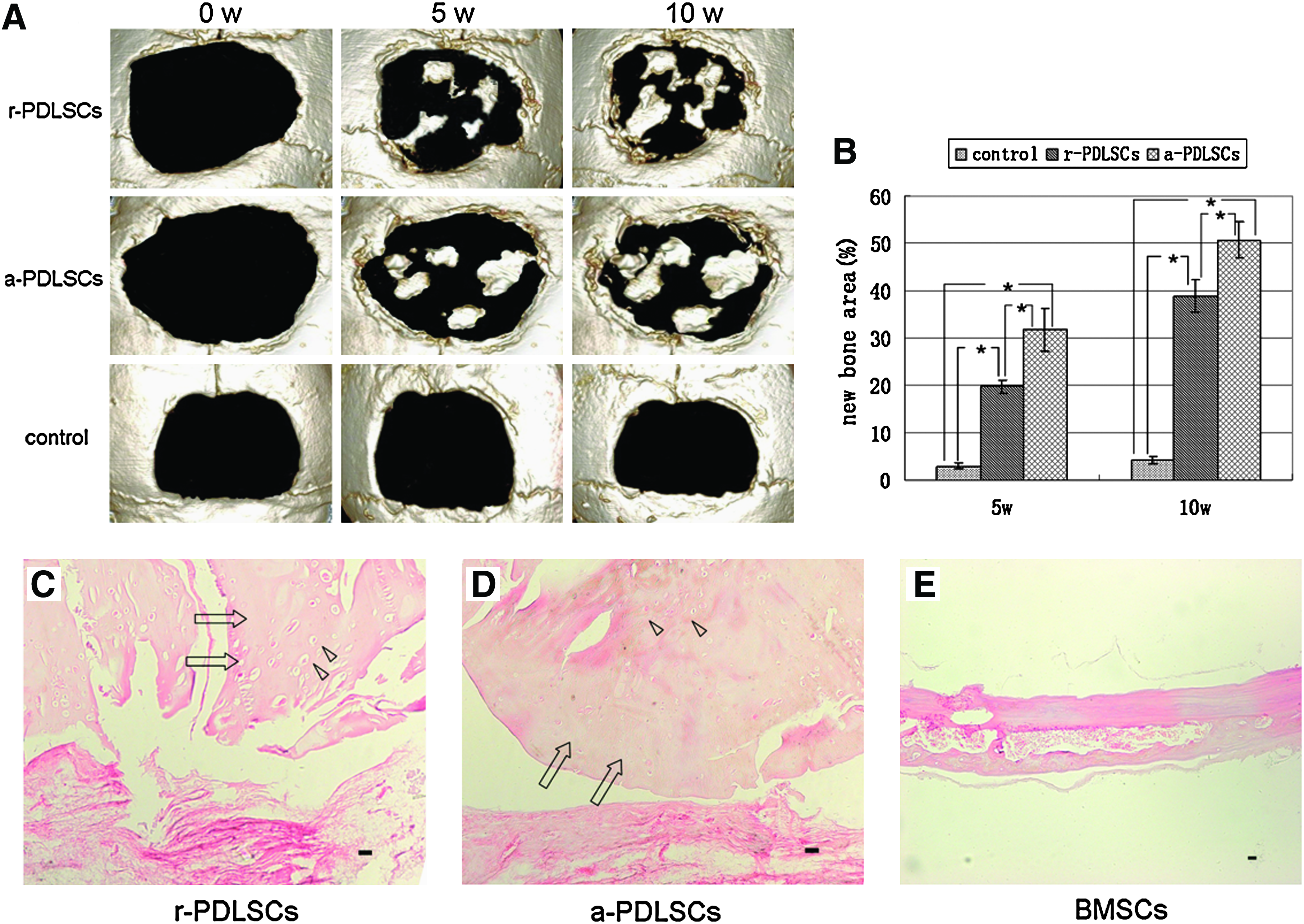
Repairing of critical-size defects in calvarial bone.
Periodontal defects repairing
About 5 weeks after operation, both autologous r-PDLSCs and a-PDLSCs had enhanced bone healing response of periodontal defects created in dogs to varying degrees. In contrast, there was no new bone formation in the control group. Statistically, the amount of the new bone formed in the a-PDLSCs group was more than that in the r-PDLSCs group. However, defects in the control group almost remained unchanged (Fig. 10).
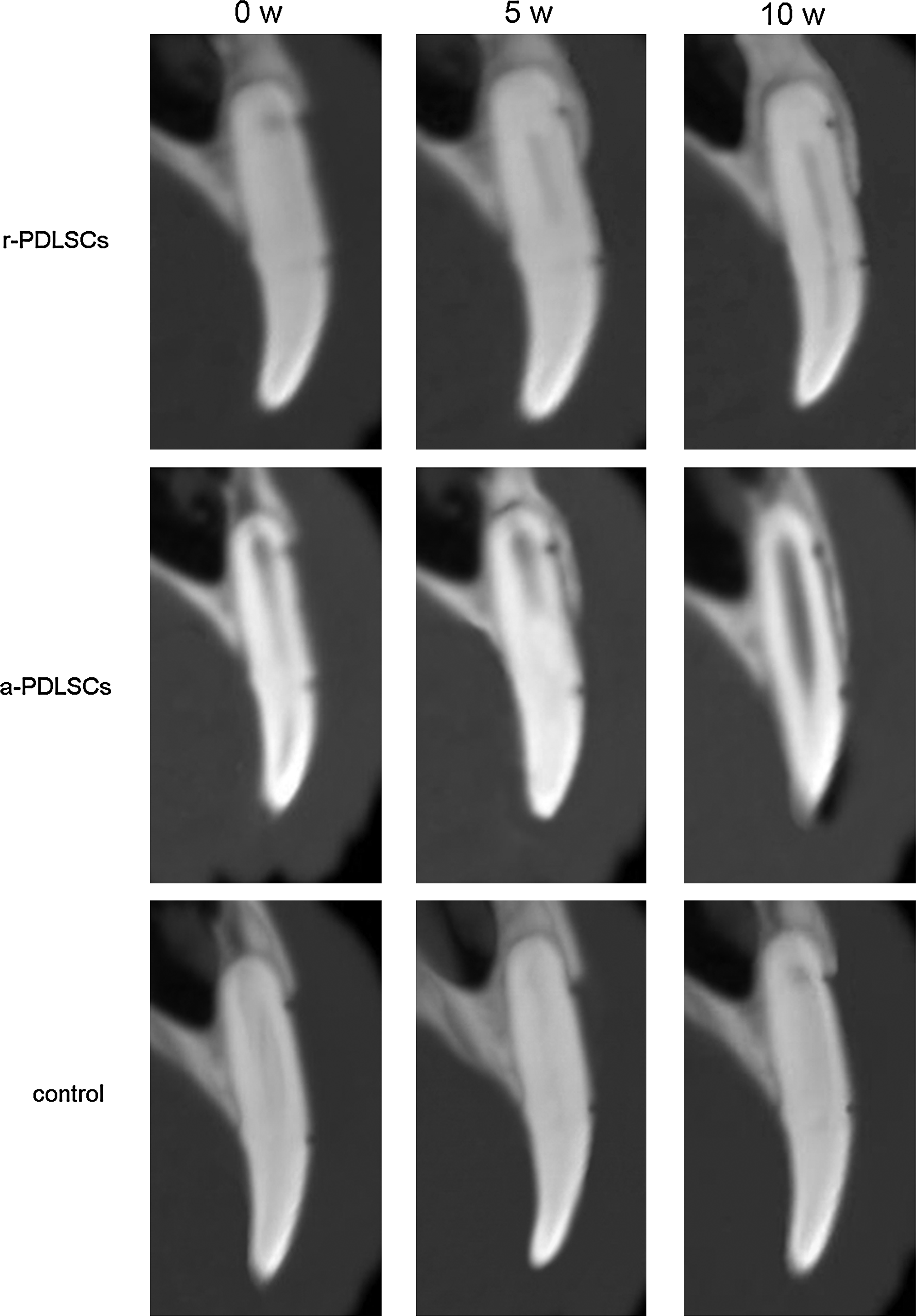
Computerized tomography images of the repairing of the periodontal defect. New bone was generated gradually in all the three groups from 0 to 10 weeks. The repairing capability of a-PDLSCs was better than that of r-PDLSCs.
Discussion
The periodontium is known to contain a heterogeneous population of periodontal progenitor cells that can differentiate into fibroblasts, cementoblasts, and osteoblast.10,21,22 The analysis of the long-term incorporation of BrdU objectively show that there is a distribution difference of periodontal stem/progenitor cells in PDL. Generally, BrdU is incorporated into proliferative cells and the uptake is decreasing gradually with time. Stem cells can retain their long-term proliferation potential due to their slow-cycling properties. 23 In the present study, it is found that there is an asymmetrical distribution of BrdU-positive cells in PDL, with more stem cells possessing long-term potential distributed mainly in the PDL on the alveolar bone surface.
In this study, our findings show that there is partial PDL remaining on the alveolar bone surface after extraction. Through scraping the alveolar socket, a-PDLSCs are successfully isolated from a new accessible tissue resource of PDL embedded on the surface of the alveolar socket and expanded in vitro. Similar to other mesenchymal stem cells, such as r-PDLSCs and BMSCs,5,24–26 a-PDLSCs exhibit the self-renewal capacity to form clonogenic clusters and undergo extensive proliferation in vitro. Meanwhile, a-PDLSCs express mesenchymal stem cell makers CD29, CD44, CD90, CD105, CD146, and STRO-1, lacking expression of hematopoietic markers CD34, CD45, and CD31. Further analysis of growth potential of a-PDLSCs indicates, consistent with the previous results, 3 mesenchymal stem cells derived from PDL have a higher proliferation rate than those derived from bone marrow, suggesting their advantages for periodontal regeneration with the possibility of being easily required with rapid expansion in vitro.
Multilineage differentiation potential has been considered a key factor in stem cell-mediated tissue regeneration. 21 Compared with other tissues in the body, periodontal tissues are unique, which include two mineralized tissues in close proximity—bone and cementum, and nonmineralized tissue—PDL. So, the ultimate goal of periodontal therapy is the regeneration of attachment apparatus lost due to periodontitis with the formation of new bone, cementum, and functionally oriented PDL. As is known, specific phenotypic characteristics of seeding cells determine the structure and function of regenerated tissues. 27 Although it has been recognized that the identification of human PDLSCs within the PDL represent a significant development in the progress toward predicable periodontal regeneration, our results indicate that a-PDLSCs with excellent proliferation rate and higher differentiation potential, in contrast to r-PDLSCs, might be a competent candidate source for periodontal regeneration. Not only are mineralization-related genes including ALP, COL I, COL III, OCN, and Runx2 highly expressed in a-PDLSCs, but also SCX, a specific transcription factor associated with tendon cells,3,5,28 is also detected at a higher level, indicating that a-PDLSCs have the capability to differentiate into two main cell lineages in periodontium, fibroblastic tissue-forming cell lineage, and mineralized tissue-forming cell lineage.
Over the past decade, strategies for periodontal repair are mainly based on anti-infectious measures such as scaling and root planning, guided tissue regeneration (with or without bone grafts), or the use of polypeptide growth factors. Although these procedures may result in some gain in clinical attachment levels and have value in inducing PDL and cementum, careful histological evaluation has indicated that both predictability and extent of healing response, especially the osteoinductive capability obtained, are highly limited and variable. Therefore, stem cell-mediated periodontal regeneration has been advocated in recent years, which greatly depends on the regenerative capacity of the stem cells involved. 29 In the present study, in vitro-expanded a-PDLSCs implants are capable of generating collagen fibers and a large area of mineralized tissues, which are apparently different from thin-layered cementum-like structures generated by r-PDLSCs, while similar to bone-like structures generated by BMSCs. Further, repairing of critical-size defects in calvarial bone and in situ transplantation into periodontal defects show that a-PDLSCs exhibit much higher osteogenic capability than r-PDLSCs, indicating that PDLSCs derived from alveolar bone surface (a-PDLSCs) are more inclined to develop bone/PDL-like tissues compatible with their origin and shed light on their unique potentials in the treatment of periodontal defects in contrast to r-PDLSCs.
In summary, previous studies have provided numerous evidences on the use of r-PDLSCs in the periodontal tissue regeneration.3–5,22 Our current study demonstrates that a-PDLSCs, possessing high self-renewal capacity and multilineage differentiation potential, are able to generate bone/PDL-like tissues in vivo and exhibit a superior effect on repairing periodontal defects, thus strongly implying their potential of synergistic use with r-PDLSCs for periodontal tissue regeneration.
Footnotes
Acknowledgments
This work was supported by grants from the Nature Science Foundation of China (30725042 and 30900316) and the National Basic Research Program (973 Program) (2010CB944800).
Disclosure Statement
No competing financial interests exist.