
Abstract
Mesenchymal stromal/stem cells (MSC), when used in combination with biomaterial scaffolds, have been shown to contribute at varying efficiencies to bone and cartilage regeneration in preclinical large animal models and human clinical trials. In an orthopedic context, identification of the optimal scaffold, which is capable of inducing tissue regeneration, has been the subject of numerous studies. In the present study, we show that ex vivo-expanded MSC from human and ovine bone marrow display similar phenotypic properties, but exhibit differences in their ability to form bone in vivo when transplanted with different biocompatible scaffold composites. We found that the ovine MSC formed ectopic bone on all scaffolds tested with the exception of collagen-based demineralized bone matrix. In contrast, human MSC in general formed less bone and only on those biomaterials composed of ceramic particles containing at least 15% hydroxyapatite. This study demonstrates the differences in bone formation potential between human and ovine MSC in vivo based on the osteoconductive properties of different bioscaffolds currently being used for orthopedic clinical applications.
Introduction
We have previously described the use of expanded autologous ovine BM stromal precursor cells to regenerate and repair critical-sized long bone defects without the requirement of a biomaterial carrier. 10 However, bone bridging across the whole defect occurred between a 6- and 9-month period. Findings from numerous other studies examining the efficacy of cell-based therapies to repair skeletal tissue defects suggest that tissue regeneration may be enhanced using the structural and mechanical support of suitable biocompatible scaffolds.6,8,11–15 Numerous scaffold composites have been examined in different skeletal defect models, including synthetic and natural biodegradable scaffolds that exhibit varying osteoconductive and osteoinductive properties.16–21
A high proportion of studies reporting on preclinical large animal orthopedic models have utilized sheep or goats to examine the efficacy of cell-based therapies to repair critical-sized bone defects that have employed porous ceramic scaffolds composed of hydroxyapatite (HA) and/or tricalcium phosphate (TCP) scaffolds.22–28 In the present study we performed a comparative study of ex vivo-expanded human and ovine MSC to form ectopic bone in vivo when transplanted with different commercially available scaffolds used in clinical orthopedic applications. The rationale for this study was to identify an appropriate bioscaffold that exhibited optimal osteoconductive properties when seeded with either human or ovine MSC in vivo, which could form the basis of comparative orthopedic large animal preclinical and human clinical trials.
Materials and Methods
Processing of human and ovine BM aspirates
BM aspirates (40 mL) were obtained from the posterior iliac crest of three normal healthy adult volunteers (20-year-old woman, 27-year-old man, and 35-year-old man), after informed consent, in accordance with procedures approved by the ethics committee of the Royal Adelaide Hospital, South Australia. BM aspirates were obtained from the posterior iliac crest of three 2-year-old Merino wether sheep according to procedures approved by the animal ethics committee of the University of Adelaide and the Institute of Medical and Veterinary Science, South Australia. BM mononuclear cells (BMMNC) were prepared by gradient density separation as previously described. 2
Establishment of primary MSC cultures
MSC isolation and culture was performed as described previously.2,29 Briefly, ∼1–3 × 108 adult human BMMNC were incubated with blocking buffer (Hank's balanced salt solution [HBSS] supplemented with 1% human serum, 1% bovine serum albumin, and 5% fetal bovine serum), and then sequentially incubated with the anti-human and ovine HSP90β monoclonal antibody (STRO-4), 29 anti-immunoglobulin g (IgG)-biotin, and streptavidin microbeads (Miltenyi Biotec) before being separated on a Mini MACS magnetic column (Miltenyi Biotec), in accordance with manufacturer's recommendations. Stromal cultures were established by culturing the STRO-4+ MNC in α-minimum essential medium supplemented with 20% fetal calf serum (FCS), 2 mM L-glutamine, 100 μM L-ascorbate-2-phosphate, 1 mM sodium pyruvate, 50 U/mL penicillin, and 50 μg/mL streptomycin.2,29
Primary antibodies
Monoclonal antibodies—mouse IgG1 anti-human CD14 (BD Biosciences), BD mouse IgG1 anti-ovine CD14 (Serotec), mouse IgG1 anti-human CD31 (BD Biosciences), mouse IgG1 anti-ovine CD31 (Serotec), mouse IgG1 anti-human CD29 (BD Biosciences), mouse IgG1 anti-ovine CD29, mouse IgG1 anti-CD44, mouse IgG1 anti-HSP-90β, 29 mouse IgG1 anti-human CD45 (BD Biosciences), and mouse IgG1 anti-ovine CD45 (Serotec)—were used at 10 μg/mL. Isotype-matched control of irrelevant specificity, 1B5 (IgG1), was kindly provided by Prof. L.K. Ashman, University of Newcastle, New South Wales, Australia.
Flow cytometric analysis
Before immunolabeling, passage 2 (P2) cultures of human or ovine MSC were liberated with trypsin/ethylenediaminetetraacetic acid (EDTA) and single-cell suspensions subsequently incubated in blocking buffer (HBSS + 20 mM Hepes, 1% normal human AB serum, 1% bovine serum albumin, and 5% FCS) for 20 min on ice. Approximately, 1 × 105 cells were resuspended in 100 μL of either neat supernatant or 10 μg/mL purified primary antibody for 45 min on ice. The isotype-matched, nonbinding control antibodies were used under identical conditions. The cells were then washed twice in wash buffer (HBSS with 5% FCS) and incubated with a goat anti-mouse IgG (γ-chain specific) phycoerythrin (1/50; Southern Biotechnology Associates) for 45 min on ice. Before analysis, cells were washed twice in wash buffer and resuspended in 1% paraformaldehyde in phosphate-buffered saline. Flow cytometric analysis was performed using a Coulter Excel flow cytometer (Coulter). Positivity for each antibody was defined as the level of fluorescence >99% of that observed when isotype-matched, nonbinding control antibodies were used. About 20,000 events were collected per sample as list mode data and analyzed using Coulter ELITE software.
In vitro differentiation assays
Passage 3 ovine and human MSC were cultured (1 × 104 cells per cm2) under osteogenic induction conditions for 4 weeks (α-minimum essential medium media supplemented with 10% FCS, 100 μM L-ascorbate-2-phosphate, dexamethasone 10−7 M, and 3 mM inorganic phosphate) as previously described. 2 Extracellular mineral deposits were identified by Alizarin red staining (1% [w/v] Alizarin Red S in distilled water) in 24-well plates. In replicate wells, adipogenesis was induced in the presence of 0.5 mM methylisobutylmethylxanthine, 0.5 mM hydrocortisone, and 60 mM indomethacin as previously described. 2 Oil Red O staining (0.5% [w/v] Oil Red O in isopropyl alcohol/distilled water 1:2) was used to identify lipid-laden fat cells after 3 weeks of induction. Chondrogenic differentiation was assessed for P3 ovine and human MSC in aggregate pellet cultures (1 × 106 cells) in the presence of 10 ng/mL TGF-β3 for 3 weeks. 2 Cross sections of paraffin-embedded pellets were de-waxed and hydrated before being stained with 1% Alcian blue in 0.1 M HCl for 1 h to detect sulfated proteoglycans.
Ectopic bone formation assay
Ex vivo-expanded P4 MSC cultures from all human and sheep donors were liberated by trypsin/EDTA treatment and resuspended in regular growth medium before being mixed with either 40 mg of 0.5-mm-diameter particles or 27 mm3 sections of each scaffold (Table 1) and implanted into subcutaneous pockets created in the dorsal surface of 8-week-old SCID mice as previously described. 2 Transplants were held together with a fibrin clot through the addition of 12 μL of mouse fibrinogen (30 mg/mL in phosphate-buffered saline; Sigma) and 12 μL of mouse thrombin (100 U/mL in 2% CaCl2; Sigma). Each transplant and the corresponding control transplant comprised of either scaffold alone or cells alone were implanted under the same conditions per mouse in triplicate mice. These procedures were performed in accordance with an ethics-approved protocol (IMVS AEC#33/05). Implants were recovered after 8 weeks, fixed for 2 days is 4% paraformaldehyde solution, decalcified for a further 10 days in 10% EDTA, and embedded in paraffin. For histological analysis, 5 μm sections of each implant was cut from the middle and either end of each 3–4 mm transplant and stained with hematoxylin/eosin as previously described. 2 The amount of new bone formation was calculated as a percentage of the total biomaterial surface area in cross sections within the transplants. Twelve tissue sections were analyzed for each of three replicate transplants per scaffold. Measurement of new bone formation was assessed using Scion Imaging Software as previously described. 30 The mean surface area of bone within each corresponding transplant was averaged from three donor samples for both ovine and human MSC.
HA, hydroxyapatite; TCP, tricalcium phosphate.
Statistics
All statistical analysis was performed using SigmaStat for Windows version 3.0 (SPSS) using the paired Students' t-test. In all cases, p < 0.05 was considered significant.
Results
Comparative analysis of human and ovine MSC
We have previously shown that clonogenic MSC and their precursors can be isolated from both human and ovine BM aspirates using an antibody directed against HSP90β. 29 As shown in Figure 1, ex vivo-expanded populations of human and ovine MSC exhibit almost identical expression of mesenchymal cell-associated antigens, CD29, CD44, and STRO-4, and lack expression of macrophage (CD14), leukocyte (CD45), and endothelial (CD31)-specific markers (Fig. 1). Ex vivo-expanded human and ovine MSC also exhibit a similar ability to differentiate into mineral-forming osteoblasts, lipid-containing adipocytes, and chondrocytes under appropriate inductive conditions in vitro (Fig. 2).

Ovine and human MSC exhibit a similar immunophenotype. Single-color immunofluorescence and flow cytometry was performed on cell suspensions of passage 2 (
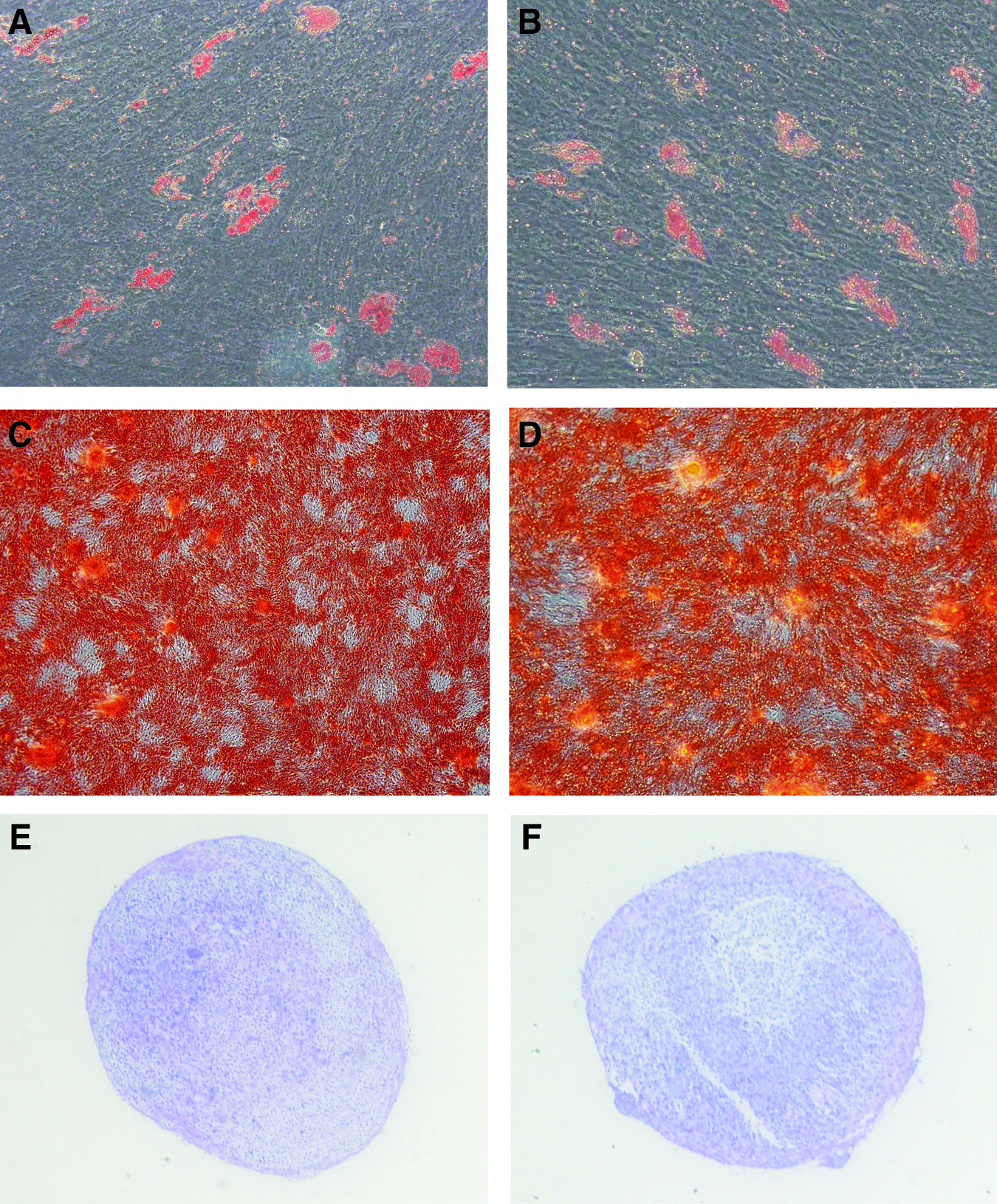
Differentiation potential of ovine and human cultured MSC. Passage 3 ovine and human MSC were cultured in either adipocyte-, osteoblast-, or chondrocyte-inductive culture conditions for 3–4 weeks. The presence of clusters of lipid-containing adipocytes were detected by Oil red-O staining of adipogenic induction in the human (
Differential capacity of human and ovine MSC to form ectopic bone in vivo on different scaffolds
Previous studies have reported the osteogenic potential of both human and ovine MSC in vitro and in vivo.2,31 In this study, we examined whether human and ovine MSC could be induced to form ectopic bone on a variety of bioscaffolds routinely used for orthopedic preclinical or clinical applications. Ex vivo-expanded human and ovine MSC at the same cell passage number were seeded onto different scaffolds (Table 1) before transplanting them subcutaneously into immunocompromised NOD/SCID mice. Each transplant contained either 40 mg of ceramic particles with a diameter size of 0.5–1.0 mm isolated using a sieve shaker (CSC Scientific), or cut into 27 cm3 pieces for collagen-based scaffolds. The transplants were harvested at 8 weeks posttransplantation and processed for histological analysis to determine the osteoconductive nature of each HA/TCP scaffold composite for both human and ovine MSC (Fig. 3). Representative hematoxylin/eosin-stained sections of each transplant were assessed for ectopic bone formation using Scion Imaging as described in the Materials and Methods section (Fig. 3). As shown in Figure 4, ovine MSC displayed the capacity to form ectopic bone on all scaffolds tested with the exception of collagen-based demineralized bone matrix. In contrast, human MSC only demonstrated significant bone formation on scaffolds composed of at least 15% HA. Like ovine MSC, human MSC failed to form a mineralized matrix on a collagenous demineralized bone matrix (Fig. 4). Interestingly, compared to the same number of human MSC, ovine MSC exhibited a three- to fivefold greater ability to form ectopic bone when seeded onto the HA-composite biomaterials. Corresponding control transplants containing scaffold alone failed to generate ectopic bone (data not shown).
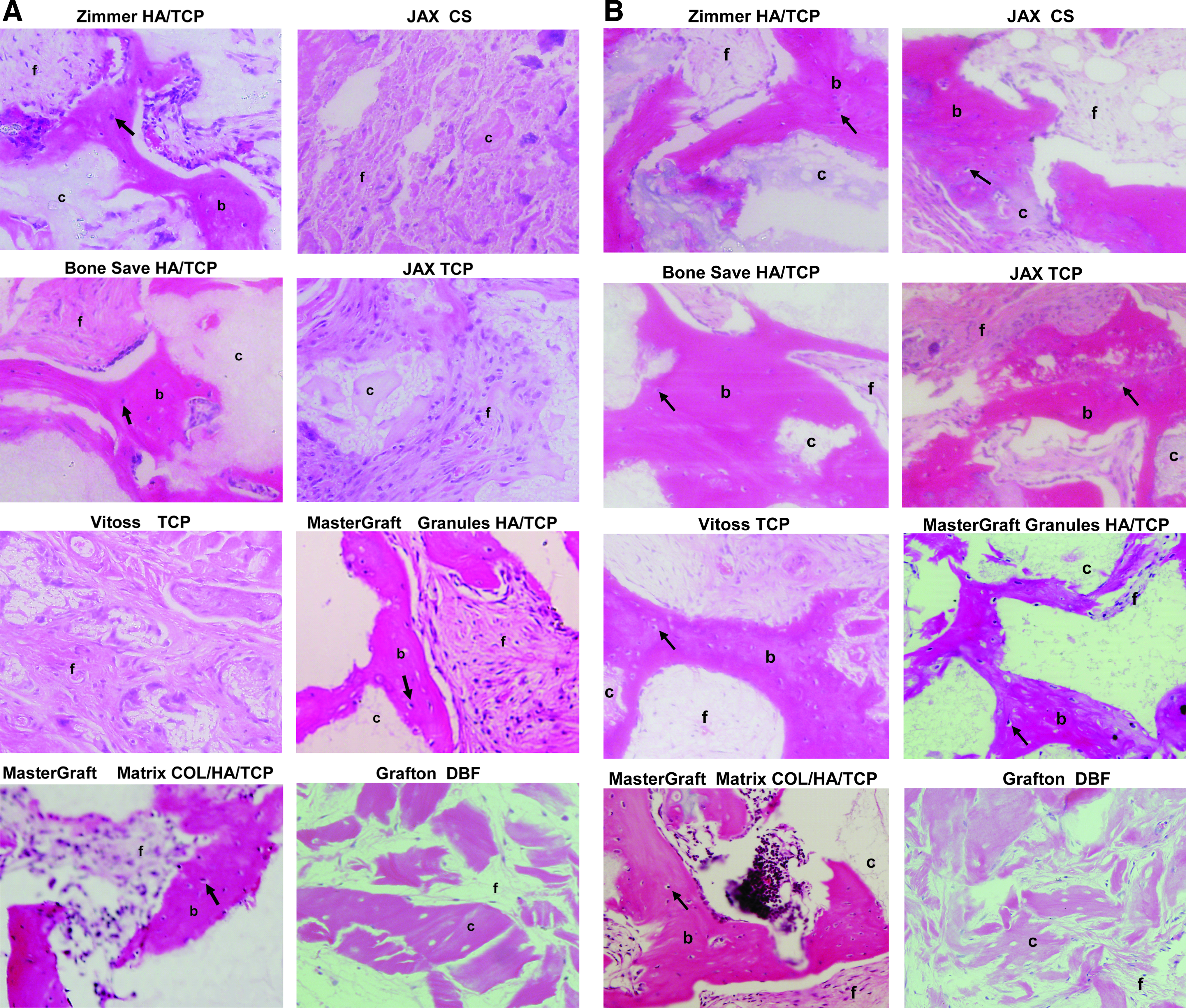
Differential capacity for bone formation in vivo by human and ovine MSC seeded onto different biomaterials. Culture-expanded passage 4 human and ovine MSC cultures were seeded onto biomaterials (Table 1) and then implanted subcutaneously into NOD/SCID mice. Cross sections representative of 8-week-old human transplants stained with H&E are shown (
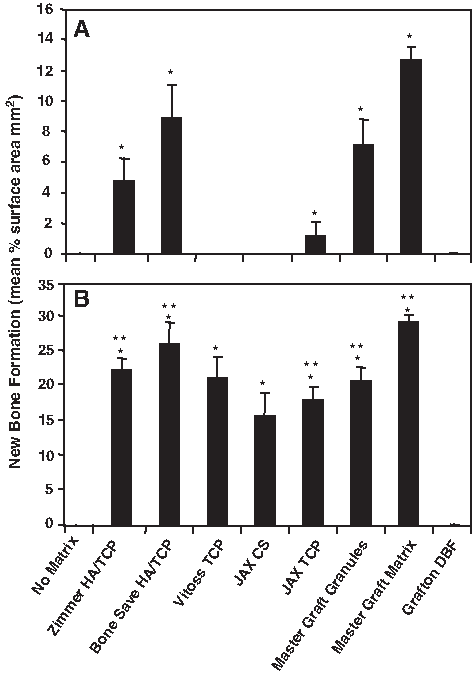
Quantitation of ectopic bone formation. Single-cell suspensions of passage 4 (
Discussion
The present study demonstrates that while ex vivo-expanded ovine and human MSC display comparable phenotypic and functional properties in vitro, they exhibit differences in their ability to form bone when seeded onto biomaterial scaffolds that are routinely used as bone filler substitutes in the clinic.32,33 Similarly, various studies have reported species differences in the growth and differentiation of MSC-like populations derived from various tissues in vitro and when seeded onto different biomaterials.34–37 Biomaterial particles of equivalent size and transplant dimensions were used in this study to ensure a comparable and optimal surface area, previously shown to maximize ectopic bone formation in vivo. 38 This variation may be due to subtle differences in the properties of ovine MSC that are known to have a higher proliferation rate and exhibit a 20% longer life span than human MSC during ex vivo expansion. This may contribute to the increased survival of ovine MSC within the transplants. Further, ovine and human MSC may respond differently to endogenous osteogenic factors in the host and may not be sufficiently conserved to elicit equivalent responses in the human and ovine MSC transplants. The inclusion of species-specific growth factors such as bone morphogenetic protein-2 and −7 bound to the scaffolds may enhance the osteogenic development of the seeded MSC populations.39–41
Our studies showed that the ovine MSC formed ectopic bone on all scaffolds tested with the exception of collagen-based demineralized bone matrix. Collagen-based scaffolds would allow more efficient binding of MSC populations that express the appropriate subclasses of β1-integrin (CD29) cell surface adhesion molecules that act as receptors for collagen. 42 Our previous studies have shown that CD29/CD49 integrin subclasses are critical for MSC colony formation on collagen and other extracellular matrix components, where disruption of these interactions leads to inhibition of proliferation and osteogenic differentiation. 42 While human and ovine MSC were found to bind to the demineralized bone matrix, both failed to form ectopic bone in the absence of osteogenic conductive signals elicited by HA and TCP.
Previous studies have reported differences in the functional properties of murine and human MSC populations based on their differential capacity to form ectopic bone in immunocompromised mice, when seeded onto different biomaterials. 16 Specifically, murine MSC exhibit the capacity to form ectopic bone in vivo on a wider range of scaffolds, including collagen-based gels, in the absence of HA or TCP when compared to human MSC transplants. While demineralized bone matrix and collagen gels have been used in preclinical and clinical orthopedic indications, both scaffolds lack structural strength for load-bearing applications and exhibit varied levels of osteoinductivity in the absence of cells or growth factors.43–45 From our studies, both ovine and human MSC displayed a greater and more consistent capacity for ectopic bone formation when seeded onto Mastergraft Matrix collagen scaffolds impregnated with ceramic particles composed of 15:80 HA/TCP. These findings support previous reports that show that human MSC display a greater ability to form ectopic bone in vivo when seeded onto scaffolds composed of 20:80 HA/TCP compared to granules comprised of 60:40 HA/TCP, despite being of similar porosity and morphology.12,18
Both resorbable ceramics (TCP and calcium sulfate) and nonresorbing ceramics (HA) are now widely used as bone fillers in orthopedic clinical settings with comparable clinical outcomes to autogenous grafts in the treatment of acute long bone fractures 46 and as supplements with autografts or allografts due to their mechanical strength and osteoconductive properties.47–49 In two studies, utilizing either autologous ovine or caprine mesenchymal cell preparations, a significant increase in bone formation was observed when the cells were transplanted in conjunction with porous ceramic scaffolds composed of either TCP or HA into critical-sized bone defects.22,26 Importantly, mechanical stability appeared to be maintained, with treated bones exhibiting similar mechanical strength under loading conditions when compared with the corresponding uninjured control bones.22,26
In a clinical context, case studies by Quarto and colleagues 50 highlight the potential use of cell-based therapies to repair nonunion bony defects in humans. In their study, three patients who presented with nonunion fractures in various limbs (4–7 cm critical-sized defects) 50 were transplanted with autologous MSC seeded onto a macroporous HA scaffold shaped to fit the individual defects. In these patients, no adverse effects were noted and callus formation and integration of the graft was evident at 2 months posttransplantation with a gradual gaining of limb function over time. The authors concluded that the recovery time would have been ∼12–18 months if traditional methods of bone grafting were employed. 50 In a similar study by the same group, long-term follow-up revealed that the defects had healed with complete fusion and integration of the graft by an average of 7 months. 51 However, it was noted that the HA scaffold exhibited low resorption even after 7 years of posttransplantation and suggested that the lack of resorption may compromise the biomechanics of the newly formed bone over time. Interestingly, the present study favored the use of a collagen-based scaffold impregnated with HA/TCP particles that may offer a higher degree of resorption over time 52 with similar osteoinductive properties compared to autogenous grafts. 46
Other animal-based bone regeneration studies have assessed the efficacy and resorbability of different ceramic scaffolds consisting of silicon and TCP. In ovine53,54 or rodent14,55 studies, reduction in the scaffold was observed by 3 months to 2 years postoperatively, coinciding with new bone formation for bone regeneration. Moreover, the efficacy of the using resorbable ceramic calcium sulfate or TCP composites as carrier scaffolds for MSC-based therapies has shown potential benefit in the clinic.56,57 However, in the absence of appropriate control subjects in these studies, the efficacy of stem cell-based therapies must be confirmed with randomized controlled trials. Other studies proposed that scaffolds composed of specific polymers may convey additional benefits, ranging from biodegradable natural materials, including silk, fibrin, and extracellular matrix materials such as collagen and Matrigel and synthetic polymers, including poly glycolic acid, poly lactic acid, and poly DL-lactic-glycolic acid.13,15,21,55,58–60 However, many of these synthetic polymer substitutes may be more effective in mediating cartilage matrix regeneration rather than bone repair,61–66 with one study demonstrating repair of large articular cartilage defects in a sheep model using implants of autologous MSC seeded onto TCP scaffolds. 67 Our study demonstrates the differences in human and ovine MSC-mediated bone regeneration potential and highlights differences in the osteoconductive properties of various commercially available bioscaffolds currently used in orthopedic applications. Therefore, caution should be taken in extrapolating animal studies involving stem cell therapies to human trials before performing appropriate comparative in vitro and in vivo analyses. The present study provides appropriate rationale for the future use of a particular biomaterial for comparative orthopedic large animal preclinical and human clinical trials.
Footnotes
Acknowledgments
This work was partly funded by the National Health and Medical Research Council (NHMRC) project grant no. 453497 and Mesoblast Ltd., Melbourne, Victoria, Australia.
Disclosure Statement
Prof. Itescu is the Executive Director of Mesoblast Ltd.