
Abstract
Noninvasive monitoring of matrix development in tissue-engineered cartilage constructs would permit ongoing assessment with the ability to modify culture conditions during development to optimize tissue characteristics. In this study, chondrocytes seeded in a collagen hydrogel were exposed for 20 min/day to pulsed low-intensity ultrasound (PLIUS) at 30 mWcm−2 and cultured for up to 5 weeks. Biochemical assays, histology, immunohistochemistry, Fourier transform infrared spectroscopy, and magnetic resonance imaging (MRI) were performed at weeks 3 and 5 after initiation of growth. The noninvasive MRI measurements were correlated with those from the invasive studies. In particular, MRI transverse relaxation time (T2) and magnetization transfer rate (km) correlated with macromolecular content, which was increased by application of PLIUS. This indicates the sensitivity of MR techniques to PLIUS-induced changes in matrix development, and highlights the potential for noninvasive assessment of the efficacy of anabolic interventions for engineered tissue.
Introduction
Mechanical stimulation has been shown to modulate the composition and biophysical properties of both engineered cartilage and cartilage explants4–6 with dynamic compression generally increasing, and static compression generally inhibiting, the synthesis of matrix macromolecules.7–12 Pulsed low-intensity ultrasound (PLIUS) is a form of dynamic compression and has been shown to accelerate union of bone fractures and enhance cartilage metabolism and repair.13–15 Although the effects of PLIUS have been shown to vary with culture systems and conditions,16–18 there is substantial evidence that it stimulates proteoglycan (PG) and collagen synthesis and increases gene expression of these macromolecules in cartilage tissue constructs.19,20
Applications of PLIUS to engineered cartilage have primarily focused on the synthesis of PG and collagen II, the two predominant extracellular matrix components, and their corresponding genes. Fewer studies have examined the effects of PLIUS on markers of degradation, such as matrix metalloproteinases (MMP), or other collagen types, including collagen types I and X, markers of articular cartilage dedifferentiation and cartilage hypertrophy, respectively. PLIUS stimulation was reported to downregulate MMP-1 expression in cultured articular chondrocytes although MMP-1321 and tissue inhibitor of metalloproteinase-1 were unaffected.21,22 Further, PLIUS inhibits the production of collagen type X gene expression in chick chondrocytes from nonossifying cartilage cultured in alginate beads.23,24 This suggests that PLIUS has an effect on a number of genes and gene products, and supports the maintenance of the chondrogenic phenotype in nonossifying cartilaginous tissues.
In addition to identifying appropriate anabolic interventions, development of therapeutics will require identification of appropriate biochemical and physical markers of cartilage quality. Evaluation of cartilage constructs is generally performed using classical techniques that are invasive and require physical disruption of tissue specimens. Histology and immunohistochemistry are semiquantitative and require specimen slicing, fixation, and dehydration. More recently, Fourier transform infrared (FTIR) spectroscopy has been used to quantify the biochemical components of cartilage.25,26 In contrast to these methods, magnetic resonance imaging (MRI), sensitive to the mobility of water protons and to the characteristics and density of tissue macromolecules, has emerged as a potential noninvasive tool for assessment of the growth and development of implanted cartilage constructs.
MR parameters sensitive to tissue properties include the longitudinal and transverse relaxation times, T1 and T2, respectively, which generally increase with tissue hydration and decrease with increasing macromolecular concentration, and the magnetization transfer rate, km, primarily but not exclusively reflecting collagen content.27,28 Correlations have been established between these parameters and cellularity, sulfated glycosaminoglycan (sGAG), and collagen content in engineered cartilage developing in vivo 29 and in vitro.28,30–33
Therefore, the aim of the present study was to evaluate the sensitivity of MR parameters to macromolecular content in engineered cartilage constructs. In addition to control tissue, we studied tissue treated with PLIUS, as a representative anabolic intervention. Although other gold-standard techniques, including FTIR and biochemical analyses, provide highly sensitive and specific data regarding matrix composition, these techniques are invasive and require tissue sampling, and therefore, unlike MRI, cannot be used clinically for cartilage evaluation.
As a model, we used a system of bovine articular chondrocytes cultured in a type I collagen scaffold, which has been shown to support chondrogenesis and prevent dedifferentiation of chondrocytes.34–37 The ultimate goal of this and related work is to further characterize a noninvasive anabolic intervention, PLIUS, for in situ cartilage development, and to apply noninvasive MRI studies to the characterization of the augmented tissue development.
Materials and Methods
Isolation of chondrocytes and fabrication of tissue constructs
Articular cartilage was harvested from 2- to 4-week-old bovine stifle joints. Chondrocytes were isolated with an 18 h digestion using 0.15% collagenase type II (Worthington Biochemicals Co.) in Dulbecco's modified Eagle's medium (Invitrogen) supplemented with antibiotics. The digest was filtered through a 100 μm nylon filter, after which the isolated chondrocytes were washed twice with phosphate-buffered saline (PBS; Invitrogen). Total cell count and viability were determined using the Trypan blue dye exclusion assay.
The isolated chondrocytes were mixed with 0.32% collagen type I gel (Sigma) to yield a final collagen concentration of 0.27% with 2.5 × 106 cells/mL. The collagen gel was prepared for cell seeding using the manufacturer's protocol. The cell-collagen constructs were allowed gel at 37°C, with full polymerization completed within 35 min. Constructs were then cultured within six-well plates in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (Hyclone), 2 mM glutamine (Invitrogen), 0.2% penicillin/streptomycin (Invitrogen), 0.25 μg/mL fungizone (Invitrogen), 50 μg/mL gentamicin (Invitrogen), 0.1 mM nonessential amino acids, 0.4 mM L-proline (Sigma), and 50 μg/mL L-ascorbic acid-2-phosphate (Sigma). Each construct had a total volume of 350 μL, was covered with 5 mL of the culture medium in each well and was cultured at 37°C under a gas mixture of 95% air/5% CO2. The culture medium was changed three times per week.
Ultrasound (US) treatment
PLIUS (Exogen 4000+; Smith & Nephew, Inc.) at a frequency of 1.5 MHz was applied to the chondrocytes after 72 h in culture. The US parameters included a spatial-average temporal-average output intensity of 30 mW/cm2, a burst frequency of 1 kHz, and a burst width of 200 μs, with the signal transmitted via coupling gel to the constructs through the bottom of the well plate. Constructs were treated 5 days per week for 20 min for either 3 or 5 weeks. Control constructs were handled identically, including attachment of the ultrasound transducers to the well plate, but the US device was not turned on.
Biochemical quantification
Constructs were blotted in a consistent fashion for the removal of excess fluid, weighed before and after lyophilization, and digested in 1 mg/mL of Proteinase K (Sigma) in 100 mM of ammonium acetate.
sGAG content was quantified from the tissue digests using the colorimetric 1,9-dimethylmethylene blue dye binding assay with a shark cartilage chondroitin sulfate standard. 38
Total collagen content was quantified using the colorimetric chloramine-T/p-dimethylaminobenzaldehyde assay with a hydroxyproline (HP) standard. 39 One hundred microliters of digest solution was hydrolyzed in 6 N HCl solution in a small sealed tube at 105°C overnight and subsequently neutralized with 6 N NaOH solution. Fifty microliters of standard and digest solution was pipetted into a clear flat-bottomed 96-well plate. Fifty microliters of chloramine-T was added to each sample and incubated in the dark for 20 min, after which 50 μL of p-dimethylaminobenzaldehyde was added to the solution in each well and incubated at 60°C for 30 min. After cooling the samples to room temperature, absorbance was measured at 540 nm on a spectrophotometer. Collagen content was calculated from the HP measurements using a collagen: HP ratio of 10:1. 40
Water content was determined by dividing the difference between the samples' wet and dry weights by the wet weights. The fractional values were reported as percent.
Histology
Constructs were rinsed in PBS, fixed in 10% neutral-buffered formalin for 7 h, and stored in 70% ethanol. Samples were dehydrated using progressively higher concentrations of ethanol, embedded in paraffin, and sectioned into 4 μm slices. Tissue sections were deparaffinized and rehydrated using xylene, decreasing concentrations of ethanol, and distilled water. Deparaffinized sections were stained with Safranin-O to observe sGAG and Alcian blue to observe PG distribution.
Immunohistochemistry
Deparaffinized sections were incubated with hydrogen peroxide (3% in 1 × PBS) for 30 min to block endogenous peroxidases. Sections were treated with hyaluronidase (1% in 1 × PBS) at 37°C for 30 min for antigen retrieval and incubated with Dako Protein Block (Dako) in PBS for 20 min to block nonspecific staining. Tissue sections were stained with monoclonal mouse antibodies against collagen I (1:100), collagen II (1:100) and collagen X (1:100), and polyclonal goat antibody against MMP-13 (1:500) and left to react overnight at 37°C. After washing with PBS, sections were incubated with peroxidase conjugated-anti-mouse immunoglobulin G and peroxidase conjugated-anti-goat immunoglobulin G as secondary antibodies. For both histology and immunohistochemistry, all staining was done simultaneously on paired samples from each group to minimize variability and permit accurate qualitative comparison of the staining intensities.
IR spectroscopy
A Nicolet Continuum FTIR Microscope (Thermo Fisher Scientific Inc.) was used to acquire mid-IR data in the wavenumber range 800–4000 cm−1 with 64 coadded scans on dried full-thickness constructs. PG was assessed using the integrated area of the absorbance centered at 858 cm−1, associated with the PG sulfate absorbance. Collagen content was calculated using the integrated area of the absorbance at 1338 cm−1, associated with collagen side chain vibrations. 41 Grams software (Thermo Fisher Scientific Inc.) was used for data processing.
MRI measurements
MR data were acquired at 4°C using a 9.4T Bruker DMX system operating at 400 MHz (Bruker Biospin, GmbH) and equipped with 1000 mT/m three-axis microimaging gradients. Images were obtained as 0.5 mm-thick sagittal slices through the center of each of the four wells in the sample holder, with all samples in a given well imaged simultaneously. T1 data were acquired using a progressive saturation spin-echo sequence with an echo time (TE) = 12.8 ms and repetition time (TR) ranging between 100 ms and 15 s in 12 steps. Magnetization transfer (MT) images were obtained by applying a 6 kHz off-resonance saturation pulse of amplitude B1 = 12 μT before the spin-echo imaging sequence with TE/TR = 12.8 ms/5 s. The saturation pulse length was incremented from 0.1 to 4.6 s in eight steps. T2 values were measured using a 64-echo Carr-Purcell-Meiboom-Gill imaging sequence with a TE/TR of 12.8 ms/5 s. Signal intensity was averaged over all pixels in a region of interest covering an entire collagen construct, with averaged intensities then fit to appropriate three-parameter monoexponential functions to obtain T1, T2, and the MT rate, km.
Statistical analyses
All values are reported as mean ± standard deviation. Biochemical data (sGAG, collagen, and water content), IR peak areas, and MR parameters were analyzed using two-way analysis of variance to test for the significance of the effects of time in culture (3 and 5 weeks) and treatment (control or PLIUS-treated). Statistical significance was taken as p < 0.05.
Results
Biochemical results
Both time in culture and PLIUS treatment resulted in statistically significant changes in sGAG (Fig. 1), with an increase from 5.71 μg/mg in the 3-week control samples to 8.23 μg/mg in the corresponding 5-week samples. Further, PLIUS application increased sGAG in the 3-week constructs by 50%, to 8.58 μg/mg. There was also a nonsignificant trend (p = 0.1) toward an increase in sGAG with PLIUS treatment at 5 weeks.
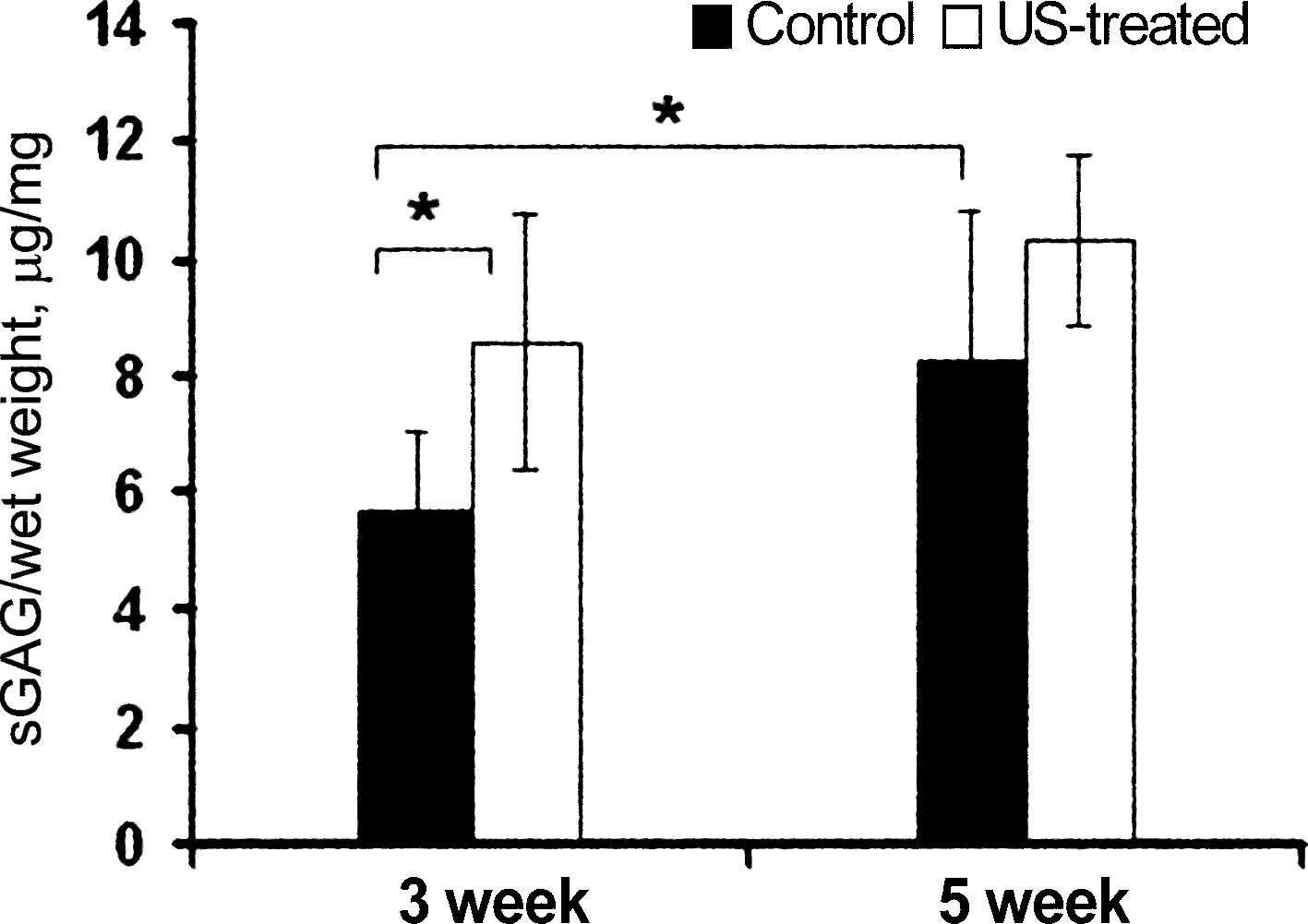
Effect of pulsed low-intensity ultrasound (PLIUS) treatment on sulfated glycosaminoglycan (sGAG) content of chondrocyte-seeded collagen I scaffolds cultured for 3 and 5 weeks (n = 6 per group). There was an increase in sGAG content with time in culture and PLIUS treatment. *p < 0.05 versus 3-week control.
Collagen content also increased significantly as a function of time in culture (Fig. 2). The collagen content of the scaffolds was 0.27% collagen per wet weight at the time of seeding; it increased from 1.6% at the 3-week time point to 3.4% after 5 weeks of growth. The effect of PLIUS on collagen concentration differed between the 3- and 5-week constructs; no statistically significant difference was observed after 3 weeks of PLIUS treatment, whereas at the 5-week time point, the PLIUS-treated group showed a significant decrease in collagen, from 3.4% to 2.5% (p = 0.029).
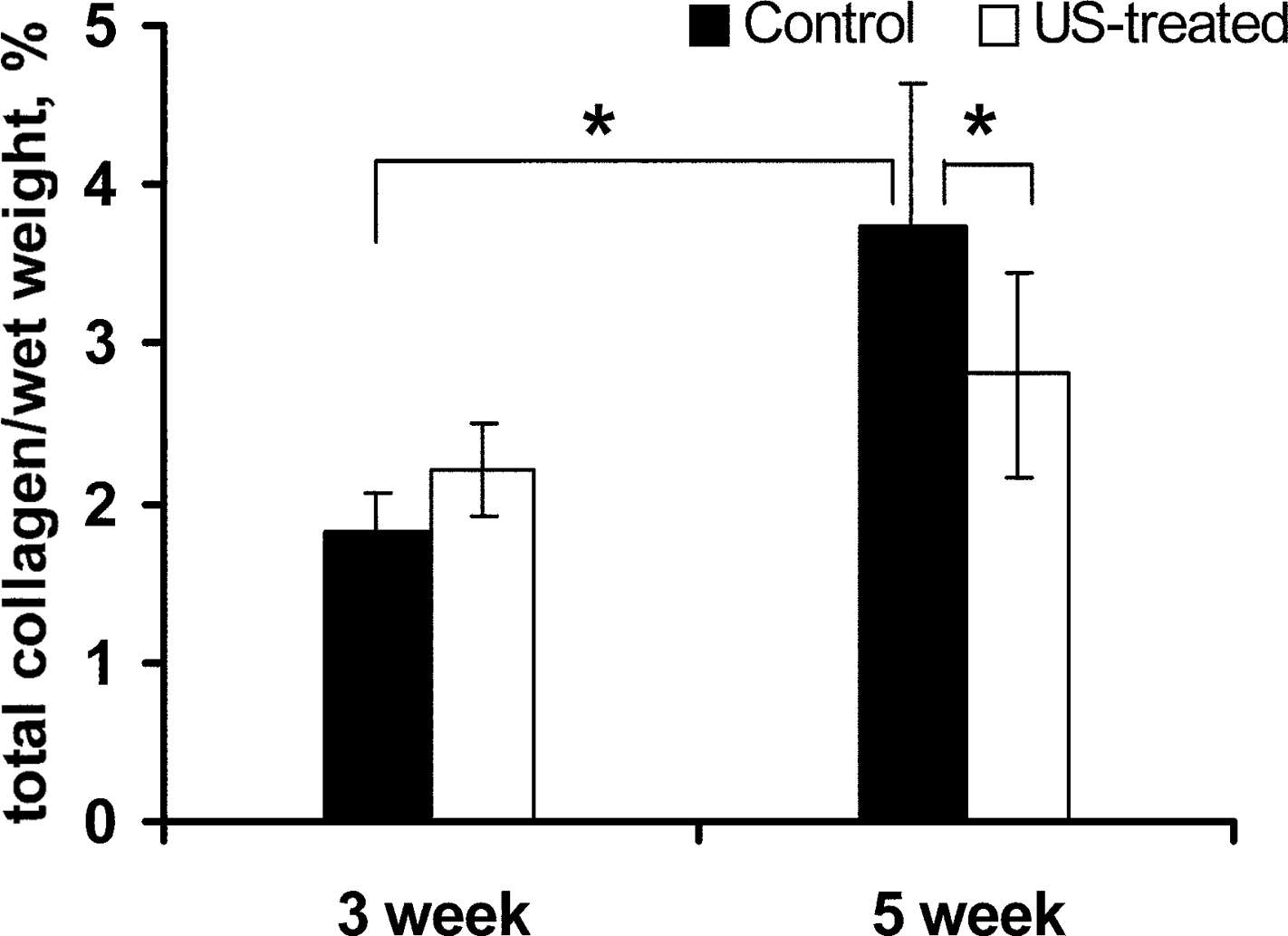
Effect of PLIUS treatment on total collagen (% wet weight) content of control and PLIUS-treated chondrocyte-seeded collagen I scaffolds cultured for 3 and 5 weeks (n = 6 per group). Collagen concentration increased with time in culture; however, the effect of PLIUS on collagen concentration varied with the age of the construct. *p < 0.05.
The water content of the constructs decreased significantly with time in culture, from 95.7% to 90.8% between 3 and 5 weeks in the control samples (p < 0.001), consistent with increased macromolecular synthesis (Fig. 3). However, the effect of PLIUS on water content was again different for 3 and 5 weeks of growth, with no effect being seen in the 3-week constructs but a significant increase in water content at the 5-week time point, from 90.8% in controls to 93.8% in the treated constructs.

Water content (% wet weight) of control and PLIUS-treated chondrocyte-seeded collagen scaffolds cultured for 3 and 5 weeks (n = 10 per group). Water content decreased in the week 3 but increased in week 5 with PLIUS treatment. Overall, water content decreased significantly with time in culture. *p < 0.05.
Histology
Staining intensities, indicating macromolecular content, increased with increased growth time and with application of PLIUS. The intensity of both the Safranin-O (Fig. 4A) and Alcian blue (Fig. 4B) stains increased markedly from weeks 3 to 5 in both control and treated constructs, consistent with increased PG synthesis. In addition, PLIUS-treated constructs showed more pronounced positive staining for both Alcian-blue and Safranin-O stains. In addition, a marked difference was seen in the staining patterns for the 3- and 5-week time points. PG was concentrated within the pericellular regions in the 3-week constructs, whereas the 5-week constructs showed homogeneous staining throughout the scaffold, indicating a more homogeneous PG distribution.
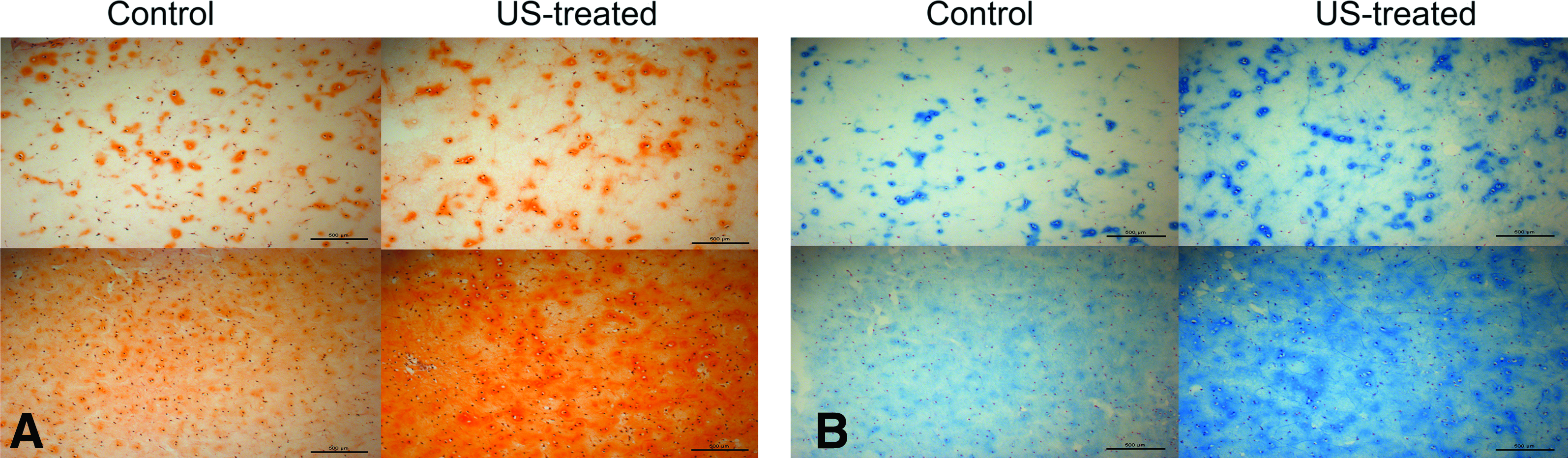
Light micrographs of
Immunohistochemistry
Immunohistochemistry aided in identifying the contribution of various collagen types to the observed PLIUS-induced variation in total collagen content (Fig. 5). Collagen type I immunostaining increased marginally with culture time, although it was minimal in all constructs (Fig. 5A).
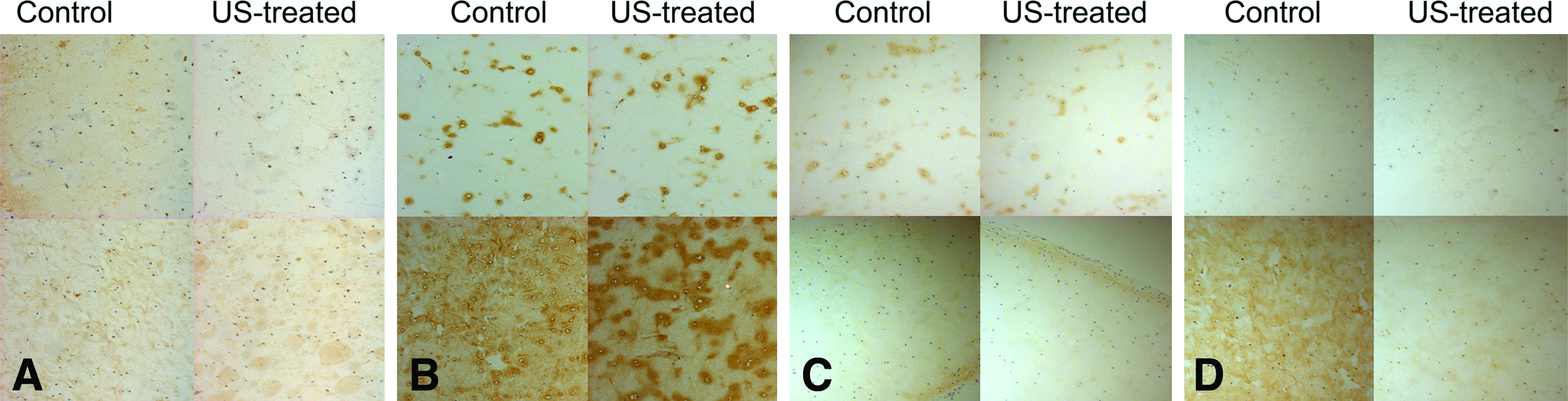
Light micrographs of immunohistochemical stains for
Both control and PLIUS-treated samples showed strong collagen II immunostaining (Fig. 5B), with more intense staining seen at the 5-week than at the 3-week time point. Application of PLIUS did not have a substantial effect in the 3-week constructs; however, there was more intense pericellular staining in the treated 5-week constructs, suggesting that PLIUS may have stimulated collagen II production.
Immunostaining for collagen X was minimal in all constructs (Fig. 5C), indicating that PLIUS does not stimulate collagen X synthesis and thus does not promote differentiation toward a hypertrophic phenotype. Although MMP-13 immunostaining was minimal in both groups, it increased with culture time (from weeks 3 to 5) (Fig. 5D). However, a salutary effect of PLIUS to inhibit MMP-13 synthesis was seen in the 5-week constructs.
IR spectroscopy
FTIR results showed a trend toward an effect of PLIUS to promote PG synthesis in the first 3 weeks of culture (p = 0.085), with a statistically significant effect seen at the 5-week time point (Fig. 6A). For the control samples, there was no significant difference between the PG contents of the 3- and 5-week constructs. However, the constructs showed a trend toward increased collagen synthesis with time in culture (p = 0.06) (Fig. 6B). A trend toward increased collagen synthesis was further seen with PLIUS application in the 3-week constructs (p = 0.07), with a nonsignificant small decrease in collagen due to PLIUS seen at 5 weeks. These results were consistent with those observed from the biochemical measurements.
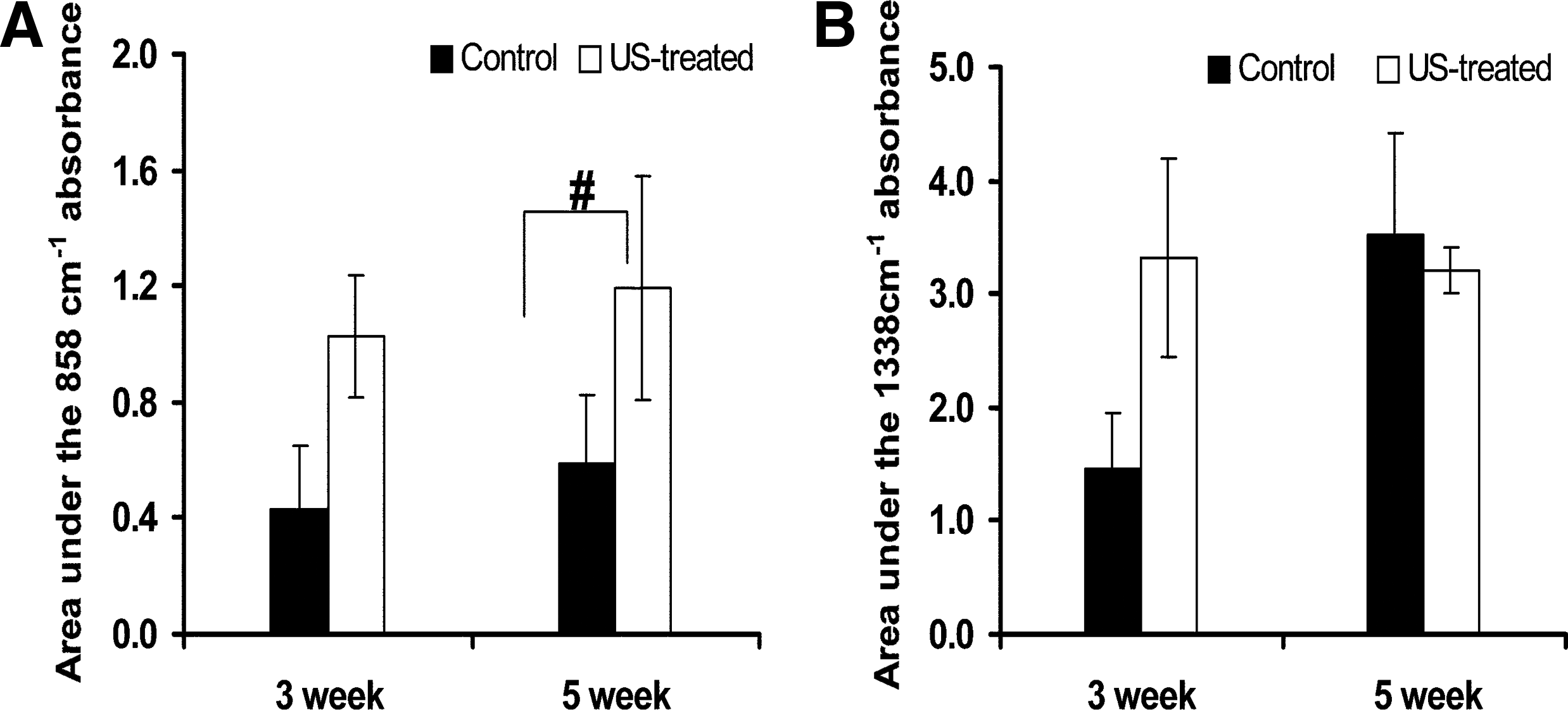
Effect of PLIUS treatment on
MR imaging
T1 values did not vary between groups (Fig. 7A), although the effects of PLIUS and culture time were both statistically significant for the magnetization transfer rate (Fig. 7B). km increased from 0.03 to 0.13 s−1 between the 3- and 5-week time points for the control samples, with PLIUS increasing km in the 3-week constructs by 93% to 0.066 s−1. In the 5-week constructs, application of PLIUS resulted in a significantly lower km, from 0.13 s−1 in controls to 0.066 s−1 in the treated samples. This is consistent with the biochemical results indicating accelerated loss of scaffold collagen with PLIUS. 42 Although the results shown in Figure 5A and B permit an assessment of the relative amounts of collagen I and II in the constructs, neither the HP assay results shown in Figure 2 nor the km results shown in Figure 7B distinguish between these two collagen types. However, the central observation of this aspect of the study is that the measurements of total collagen per wet weight and the MRI parameter km show similar patterns, with a statistically significant increase seen in control samples over time, and a statistically significant decrease with application of PLIUS seen at the 5-week time point.

Effect of PLIUS treatment on
T2 values decreased with time in the control samples, with no change with time being seen in the PLIUS-treated group (Fig. 7C). PLIUS significantly shortened T2 by 15.1% in the 3-week constructs. In contrast, there was a trend toward increased T2 in the PLIUS-treated group at the 5-week time point (p = 0.06). This nonstatistically significant trend may be consistent with the interpretation that at 5-week time point, the rate of scaffold breakdown in the treated samples exceeded the rate of matrix synthesis. Similar to the case for km and total collagen, it is clear that the pattern of T2 changes within the four experimental groups (Fig. 7C) matches that of the biochemical evaluation of sGAG (Fig. 1).
Discussion
Our results indicate that MRI is sensitive to time-associated- and PLIUS-induced changes in tissue-engineered cartilage construct development. This indicates the potential utility of MRI to noninvasively assess tissue engineering protocols during construct development as well as the efficacy of tissue engineering-based in vivo therapeutic interventions.
Previous studies have documented the salutary effects of PLIUS stimulation on chondrocyte metabolism.23,24 In this study, biochemical assays and histology demonstrated an increase in GAG content resulting from PLIUS treatment, consistent with previous work.19,20 The change in GAG distribution from the 3- to 5-week time point, observed by histology, was indicative of construct maturation, also consistent with previous studies. 43
FTIR and MRI have previously been used together to assess PG- and collagen-containing tissues.44,45 The integrated areas under the 858 and 1338 cm−1 absorbances are directly proportional to PG and collagen content, with our results confirming that the PG concentration of the constructs increased significantly with PLIUS treatment. Further, FTIR results verified that the effect of PLIUS on collagen concentration was different for the 3- and 5-week constructs. Thus, the FTIR results were consistent with biochemistry and MRI data.
Type I collagen, a marker for chondrocyte dedifferentiation, is the primary collagen type present in fibrocartilage and is found in repair tissue formed after damage to articular cartilage. 46 The absence of intense pericellular staining in the 3-week constructs, a pattern common with the other stains, indicated that collagen type I synthesis was minimal. Our findings are consistent with previous studies indicating that chondrocytes in 3D culture typically produce minimal collagen type I.47–49 In addition, although the collagen I produced by the chondrocytes was indistinguishable from that of the scaffold, it is evident from the immunostaining results that PLIUS did not stimulate collagen I production.
Type II collagen is the major component of articular cartilage and is required for a mechanically functional collagen network. 50 We found that collagen II increased with time in culture from 3 to 5 weeks. Although application of PLIUS for a full 5 weeks did not have a significant effect on overall collagen II immunostaining, a more intense pericellular pattern is evident. There are conflicting reports about PLIUS’ effect on type II collagen synthesis. Our findings are consistent with studies that have shown an increase in both collagen II and collagen II mRNA with PLIUS stimulation.17,20 The continuous production of collagen II observed in the constructs suggests that the cells maintained their chondrocytic phenotype throughout the culture period.
Collagen X is upregulated during osteoarthritis (OA) and is a known marker for chondrocyte hypertrophy, which is a necessary step in endochondral ossification.51–53 Chondrocyte hypertrophy is also associated with matrix mineralization, which reduces the weight-bearing capacity of articular cartilage and promotes matrix damage.54,55 Therefore, synthesis of collagen X is indicative of damaged and mineralizing cartilage. Given the efficacy of PLIUS in promoting fracture healing, it is essential to establish that PLIUS does not stimulate expression of collagen X mRNA and protein in articular cartilage. In a previous study 23 we showed that PLIUS increased collagen X in ossifying (proximal) sternum, but did not stimulate collagen X in nonossifying (distal) sternum. This suggests that the effect of PLIUS stimulation is dependent on the cell's microenvironment and does not alter the hyaline phenotype. Further, Mukai et al. reported that exposure of a chondrocyte pellet culture to PLIUS resulted in delayed expression of the collagen type X gene. 56 In the present study, collagen X was produced at a very low level in all groups, and was further diminished in the PLIUS-treated samples providing evidence that PLIUS inhibits the differentiation of chondrocytes toward the hypertrophic phenotype, thus potentially ameliorating this aspect of the pathology of OA.
MMPs are associated with cartilage destruction in OA, and MMP protein production and gene expression have been shown to be upregulated in osteoarthritic cartilage.57,58 Although its substrates include aggrecan and collagen types I, II, and III among others, MMP-13 is reported to cleave collagen type II more than any other collagen type. 59 We found that MMP-13 production increased at the 5-week compared to the 3-week time point in control samples. Metalloproteinases are constitutively expressed at low levels in healthy connective tissue, presumably to permit ongoing tissue remodeling. 60 Although we cannot specifically account for the increased MMP-13 production with culture time in the control constructs, Freyria et al. 61 also reported that MMP-13 gene expression was increased in type I collagen gels with culture time. However, this increase was prevented in the treated samples by the application of PLIUS (Fig. 5D), a further indication of its chondroprotective effect. These results are similar to those of Gurkan et al., who showed that PLIUS treatment decreased MMP-13 expression in knee joints of guinea pigs concomitant with the prevention or delay of onset of osteoarthritic cartilage changes. 62
The sensitivity of MRI to the PLIUS intervention indicates the potential for noninvasive assessment of the quality of both engineered and in situ cartilage. The increase in km in the control constructs between 3 and 5 weeks of growth is consistent with the biochemically-determined increase in total collagen. 63 The decrease in total collagen in the 5-week PLIUS-treated samples as compared to the 5-week controls may reflect acceleration of scaffold breakdown. However, km depends on other tissue characteristics besides total collagen, including collagen cross-linking. 64 To our knowledge, the effect of PLIUS on cross-linking has not been investigated; such an effect would complicate the interpretation of km data on PLIUS-treated samples. Application of PLIUS may also have resulted in selective inhibition of the synthesis of other, more minor, collagen types as seen for collagen type X. Further, the increased water content of the 5-week PLIUS-treated samples and resulting decrease in collagen concentration may account for the decrease in km seen by MRI. Nevertheless, at the 5-week time point, the MRI km measurement was consistent with the biochemically measured total collagen. Finally, we note that the relationship between km and collagen content and other matrix characteristics has not been widely investigated in engineered cartilage and may differ from that seen in native cartilage.
T2 is sensitive to a wide range of cartilage matrix characteristics, and generally increases as the proportion of free water to macromolecular-associated water increases. Our finding of a decrease in T2 with PLIUS treatment in the 3-week constructs is consistent with increased macromolecular synthesis from the intervention, as also seen with km. The trend toward increased T2 in the 5-week treated constructs was likely due to increased water content and decreased collagen concentration; breakdown of the collagen scaffold and inhibition of synthesis of collagen X is consistent with this. This finding also reflects the relatively greater influence of collagen than of PG to shorten T2. 65
In summary, we find that PLIUS increases PG and collagen type II synthesis of chondrocytes seeded into collagen I scaffolds, does not stimulate collagen type I production, and inhibits the production of collagen type X and MMP-13. We have established the sensitivity of noninvasive MR techniques to this intervention, and validated these results with the application of FTIR and other invasive assessment modalities to the engineered constructs. We found that the MR parameters T2 and km reflected the biochemically determined total construct collagen and sGAG, respectively. The sensitivity of MRI in this setting establishes this approach as a promising one for evaluation of cartilage repair strategies based on both ex vivo engineering and in situ development of viable cartilage within a diseased joint.
Footnotes
Acknowledgments
This work was supported in part by the Intramural Research Program of the NIH, National Institute on Aging and by NIH R01 EB000744 and AR056145 (NP). We would also like to acknowledge the Hospital for Special Surgery (New York, NY) for providing histology and immunohistochemistry services (NIH AR046121).
Disclosure Statement
No competing financial interests exist.