
Abstract
Scaffold material is expected to play a crucial role in induction of chondrogenic differentiation of mesenchymal stem cells (MSCs) for cartilage tissue engineering. Here we demonstrated the feasibility of a fibrin/hyaluronan (HA) composite hydrogel as a potent scaffold for support of chondrogenesis of rabbit MSCs (rMSCs). rMSCs were prepared in three-dimensional cultures of pellet, alginate layer, and fibrin/HA gel. Specimens in each group were cultured in chondrogenic defined media for 4 weeks in the absence or presence of transforming growth factor β1 (TGF-β1) treatment. Viability of rMSCs was somewhat reduced until 4 weeks, which was less significant in fibrin/HA gels than in the alginate layer (*p < 0.05). The fibrin/HA group showed transient size reduction by about 35% at 1 week, but showed significantly higher mechanical strength than the alginate group. In safranin-O and alcian blue stains, accumulation of sulfated glycosaminoglycans (GAGs) was observed clearly from 1 week, and homogenously in the entire area at 4 weeks in the fibrin/HA group. Of note, TGF-β1 treatment showed no additional effect on GAGs accumulation in the fibrin/HA group. The alginate and pellet groups, however, showed much lower levels of GAGs accumulation only in the presence of TGF-β1. Biochemical assays for GAGs and collagen, and expression of chondrogenic markers also showed much better results in the fibrin/HA group, even without TGF-β treatment than the other groups. These results demonstrated that fibrin/HA composite gel efficiently promoted chondrogenic differentiation of rMSCs, even without TGF-β treatment, and that it could be a useful tool for use in cartilage tissue engineering.
Introduction
Because fibrin possesses important features, such as high biocompatibility, biodegradability, and affinity to biological tissues, it is a useful cell delivery matrix for cartilage tissue engineering.2–4 Hyaluronic acid (HA) is a key component of normal articular cartilage and a highly hydrophilic biopolymer with a large molecular mass. In addition, interactions between chondrocytes and extracellular matrix (ECM) molecules such as HA are known to be critical to maintenance of cartilage homeostasis. 5 When the advantages of the two materials are ideally combined, the hybrid biomaterials could be an attractive scaffold to support better cell survival, proliferation, differentiation, and matrix synthesis, thus being able to reconstitute the hyaline architecture of cartilage. Combination of fibrin with HA has been previously suggested to have a positive effect on cartilage tissue engineering using chondrocytes in our laboratory and in other groups.6,7 We constructed a composite gel of fibrin/HA and observed that it was very efficient in preserving the chondrogenic phenotype of chondrocytes. 6 The potential of fibrin/HA gel was also observed in building of a hyaline-like cartilage construct using chondrocytes, and the structural integrity of the constructs was well maintained throughout the long-term implantation period (4 weeks) in nude mice. Therefore, the fibrin/HA composite gel was expected to provide a favorable environment for chondrocytes for maintenance of their characteristic phenotypes and synthesis of cartilage ECMs.
Considering the major limitations of chondrocytes, such as cell number and loss of phenotype after passage of culture, mesenchymal stem cells (MSCs) are emerging as a promising substitute. Currently, one of the most critical hurdles to use of MSCs in tissue engineering is the question of how to induce and control their differentiation into specific lineages. It is expected that scaffold materials could play an important role in chondrogenic differentiation of MSCs and cartilage tissue engineering.8,9
We presumed that the advantage of fibrin/HA composite gel, which has been demonstrated in cartilage tissue engineering with chondrocytes, could also be applied to chondrogenic differentiation and cartilage tissue formation using MSCs. In this study, the activity of fibrin/HA composite to induce chondrogenic differentiation of rabbit MSCs (rMSCs) was compared with those of alginate layer and pellet cultures in vitro that are representative and widely used methods for cartilage tissue engineering. In addition, the effect of transforming growth factor β1 (TGF-β1) treatment on chondrogenesis of rMSCs in these scaffolds was also evaluated.
Materials and Methods
Preparation of bone marrow-derived rMSCs
Isolation and culture of rMSCs from bone marrow were performed as previously described, with some modification. 10 Bone marrow aspirates were obtained aseptically from the tibia and femur of 2-week-old New Zealand white rabbits (n = 5, Joong-Ang Experimental Animal Center, Seoul, Korea). Aspirates were suspended in 5% acetic acid to remove red blood cells and centrifuged at 500 g for 5 min to remove supernatants. The cell pellet was resuspended in Minimal essential medium Eagle-alpha modification (Gibco BRL, Carlsbad, CA) supplemented with 1% antibiotic–antimycotic and 10% new born calf serum. Cells were seeded at 1.5 × 106 in a culture dish (dia., 150 mm), and cultured in the monolayer for 10 days in a humidified incubator with 5% CO2 supply. Adherent MSCs were then detached using 0.05% trypsin–EDTA (Gibco BRL-Life Technologies, Grand Island, NY) and replated on the dish in the same medium for expansion until the three-dimensional (3D) culture. Expression of CD90 and CD105 on the surface of isolated MSCs was confirmed as described previously. 11
Culture of rMSCs in fibrin/HA composite gel
rMSCs (2 passage) were suspended at 5 × 106 cells/mL in a solution containing 9–18 mg/mL fibrinogen (Green Cross, Young In, Korea) and 10 mg/mL HA of 3000 kDa (LG Chemical Institute, Daejeon, Korea). 6 The rMSC suspension was mixed homogeneously with 110 KIU/mL aprotinin (Green Cross), 60 U/mL thrombin (1000 U/mg; Sigma, St Louis, MO), fibrin stabilizing Factor XIII, and 50 mM CaCl2; 250 μL aliquots of the fibrin/HA mixture with rMSCs were dropped into an empty Petri dish to form a gel. The gels were transferred to six-well culture plates and cultured in chondrogenic defined medium (CDM) with or without 10 ng/mL TGF-β1 (Peprotech, Rehovot, Israel). CDM was composed of high-glucose Dulbecco's modified Eagle's medium (Gibco BRL) supplemented with 1% antibiotic–antimycotic (Gibco BRL), ITS supplement (1.0 mg/mL insulin from bovine pancreas, 0.55 mg/mL human transferrin, and 0.5 mg/mL sodium selenite), 50 μg/mL ascorbic acid, 100 nM dexamethasone, 40 μg/mL proline, 1.25 mg/mL bovine serum albumin, and 100 μg/mL sodium pyruvate. All products were purchased from Sigma.
rMSCs culture in the alginate layer
rMSCs were suspended in sodium alginate (2.4% in 0.15 M NaCl; Sigma) solution at a density of 5 × 106 cells/mL and then filtered using a 0.2 μm filter for sterilization. The alginate/rMSC mixture (300 μL aliquots) was transferred to the transwell insert and spread evenly over the surface. The transwells were immersed in 1 mL of sterile CaCl2 solution (100 mM) for 10 min to produce solid alginate/rMSC layers of 3 mm in thickness. The layers were washed twice with 0.15 M NaCl and rinsed with CDM. The layers were then cultured in chamber wells containing CDM with or without 10 ng/mL TGF-β1 (Peprotech).
Pellet culture
rMSCs were placed in a 15 mL polypropylene tube at a density of 5 × 105 cells/0.5 mL of culture medium, and harvested by centrifugation for 5 min at 1000 g in Dulbecco's modified Eagle's medium with 10% fetal bovine serum. Cell pellets in the tubes were then cultured in CDM with or without 10 ng/mL TGF-β1 (Peprotech). Tube caps were loosened in order to maintain 37°C and 5% CO2. The culture medium was changed every other day after the 3rd day.
Measurement of cell viability by live/dead cell assay
Viability of cells in each scaffold was examined using a Live/Dead assay kit (Molecular Probes, Eugene, OR) according to the manufacturer's instructions. Samples were treated in a solution containing 2 mM ethidium homodimer-1 and 4 mM Calcein AM in phosphate-buffered saline for 40 min. After washing in phosphate-buffered saline, thin slices of stained samples were prepared on glass slides and observed by fluorescence microscope. Live cells were observed in green color, and dead cells were in red color. Viability of cells was measured by dividing the number of viable cells (green cells) with that of total cells (green cells + red cells), and determined using the ImageJ software.
Histological observations
Samples in each scaffold were fixed with 10% formalin for 24 h. Alginate layers were polymerized in BaCl2 solution before fixation. Samples were then embedded in paraffin wax and sectioned serially at 4 μm thickness. Sections were stained with hematoxylin and eosin (H&E) for histological observation, and safranin-O and alcian blue for sulfated proteoglycans.
Biochemical assays for glycosaminoglycans and collagen
Samples were digested for 16 h with papain digestion solution (125 μg/mL papain, 5 mM L-cystein, 100 mM Na2HPO4, and 5 mM EDTA, pH 6.2) at 60°C. The amount of glycosaminoglycans (GAGs) was measured using the 1,9-dimethylmethylene blue assay. 12 Individual samples were mixed with the dimethylmethylene blue solution, and absorbance was measured at 525 nm. Total GAG content in each sample was extrapolated using a standard plot of shark chondroitin sulfate (Sigma) in the range of 0–5 μg/mL. Collagen content was measured using a modified Hride tullberg-reinert method.6,13 Digested samples were dried at 37°C in 96-well plates for 24 h and reacted with a dye solution for 1 h with mild shaking. The dye solution (pH 3.5) was prepared with Sirius red dissolved in picric acid-saturated solution (1.3%) at 1 mg/mL. After washing five times with 0.01 N HCl, the dye–sample complex was resolved in 0.1 N NaOH. Absorbance was read at 550 nm using an ELISA reader (BIO-TEK Instruments, Winooski, VT). Total collagen content of each sample was extrapolated using a standard plot of bovine collagen (Sigma) in the range of 0–10 μg/mL.
Reverse transcriptase–polymerase chain reaction
Total RNAs from each specimen were extracted using Trizol reagent (Invitrogen, Carlsbad, CA). Total RNAs of 1 μg were reverse-transcribed using the Superscript First Strand Synthesis System (Roche, Indianapolis, IN) to produce a complimentary DNA. Complimentary DNA products (2 μg) were amplified using specific primers for chondrogenic markers: type I and II collagens, sox 9, and aggrecan. mRNA level of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was measured as an internal control. Specific primer sequences were as follows: type I collagen (sense: 5′-cttctggcccgctggaaaggatg-3′, antisense: 5′-cccggatacaggtttcgccagtag-3′), type II collagen (sense: 5′-gatattgcacctttggacat-3′, antisense: 5′-cccacaatttaagcaagaag-3′), aggrecan (sense: 5′-gaaaggtgttgtgttccact-3′, antisense: 5′-gtcataggtctcgttggtgt-3′), and GAPDH (sense: 5′-caacttttcagagggacaag-3′, antisense: 5′-ctaaaccatccaatcggtag-3′). Polymerase chain reaction products were separated on a 1.5% agarose gel and stained with ethidium bromide. Signals were observed using a low light image system (GelDoc 2000; BioRad, Hercules, CA). The quantitative data for band intensities were generated by analyzing the gel images on the ImageJ software (Version 1.42).
Analysis of mechanical strength
Specimens (n = 3) were subjected to a conventional open-sided (unconfined) compression test using the Universal Testing Machine (Model H5K-T, Boras, Sweden). Tests were conducted at room temperature. All tests were performed using a displacement control mode at a rate of 1 mm/min. The machine stopped automatically after moving a programmed length in between top and bottom platens. After the tests, the compressive stress and strain were graphed based on the measured sample dimensions: cross-sectional area based on the sample size and sample height (measured automatically at .02 N tare load) and determined by normalizing against sample geometries. Once a peak load was obtained from the load–displacement curve, the compressive modulus was calculated based on a linear regression fitting of a small strain section that precedes an identifiable plateau region and compressive strength was determined using an offset-yield approach. A line was drawn parallel to the modulus line, but offset by 0.5% of the initial sample gauge length. The corresponding stress value at which the offset line crossed the stress–strain curve was defined as the compressive strength of the scaffold.
Statistical analysis
Statistical differences in quantitative biochemical and mechanical analysis were determined using the Mann–whitney U-test (Independent t-test, SPSS). Statistical significance was assigned as *p < 0.05, **p < 0.01, and ***p < 0.001, respectively.
Results
Viability of cells in fibrin/HA composite gel and the alginate layer
rMSCs were differentiated into chondrocytes in the alginate layer and fibrin/HA gel until 4 weeks; morphology and viability of cells were first examined by H&E stain and Live/Dead assay, respectively. H&E images of specimens at 1 week showed that rMSCs were integrated well with their round morphology both in the alginate layer and Fibrin/HA composite gel (Fig. 1a). In the Live/Dead assay performed at 1 day and 1, 2, and 4 weeks, cell viability was slightly higher overall in fibrin/HA gel than in the alginate layer (Fig. 1b). The difference was statistically significant, particularly at 4 weeks (*p < 0.05), and can be observed clearly in the representative images in Figure 1c. Percentage of viable cells (green color) were ∼77% ± 2.6% of the entire cell population (green color + red color) in fibrin/HA gel, whereas it was ∼60% ± 5% in the alginate layer at 4 weeks.

Cell viability.
Gross observation
Gross images of specimens in the alginate layer, fibrin/HA gel, and pellet were observed at 1 day, and 1, 2, and 4 weeks after culture (Fig. 2). Specimens of the alginate layer were ∼12 mm in diameter and appeared to be semi-transparent at 1 day. Their appearance and size did not change significantly until 4 weeks, regardless of TGF-β1 treatment. Specimens of fibrin/HA gel were ∼10 mm in diameter and were also semi-transparent at 1 day. However, they showed a glint of white color and some size reduction (approx. 6.5 mm in diameter) at 1 week, after which no obvious change was observed. Treatment with TGF-β1 showed no significant effect on their appearance and size. Specimens of the pellet culture were ∼1.5 mm in diameter at 1 day, and increased about two times (approx. 3.0 mm), only in the presence of TGF-β1 at 4 weeks. Their color was white at all time points.

Gross observation of specimens after chondrogenic differentiation. rMSCs were cultured in the alginate layer, fibrin/HA gel, and pellet in the CDM with (+) or without (−) 1 ng/mL TGF-β1. Gross images of the specimens were observed at 1 day and 1, 2, and 4 weeks after culture. Scale bar = 1 mm. Color images available online at www.liebertonline.com/tea
Histological observation for sulfated GAGs
The specimens above were examined for the lacunae formation and accumulation of sulfated GAGs by H&E, safranin-O, and alcian blue stainings (Fig. 3). The H&E staining images are not presented in Figure 3 because the distribution of lacunae is also observed in the safranin-O and alcian blue staining images. In the alginate group, the cartilage-specific lacunae formation was increasing with time and evenly distributed in the whole construct area. It was observed in much larger amount and more wildly distributed in the presence of TGF-β1. The alginate layer itself was stained red in the safranin-O staining, and accumulation of sulfated GAGs was not clearly distinguished by intensity of color, but the alcian blue staining of the alginate group also showed that TGF-β1 treatment induced lacunae formation and strong accumulation of sulfate GAGs at early time points, particularly around the peripheral regions. Results from the pellet group at 4 weeks also showed a significant increase in the number of lacunae and accumulation of sulfated GAGs only in the presence of TGF-β1. In contrast, in the fibrin/HA group, the lacunae formation and accumulation of sulfated GAGs were increasing significantly with time both in the presence and in the absence of TGF-β1. This result suggests that fibrin/HA gel provided strong support for chondrogenic differentiation of rMSCs, even without TGF-β1 treatment.
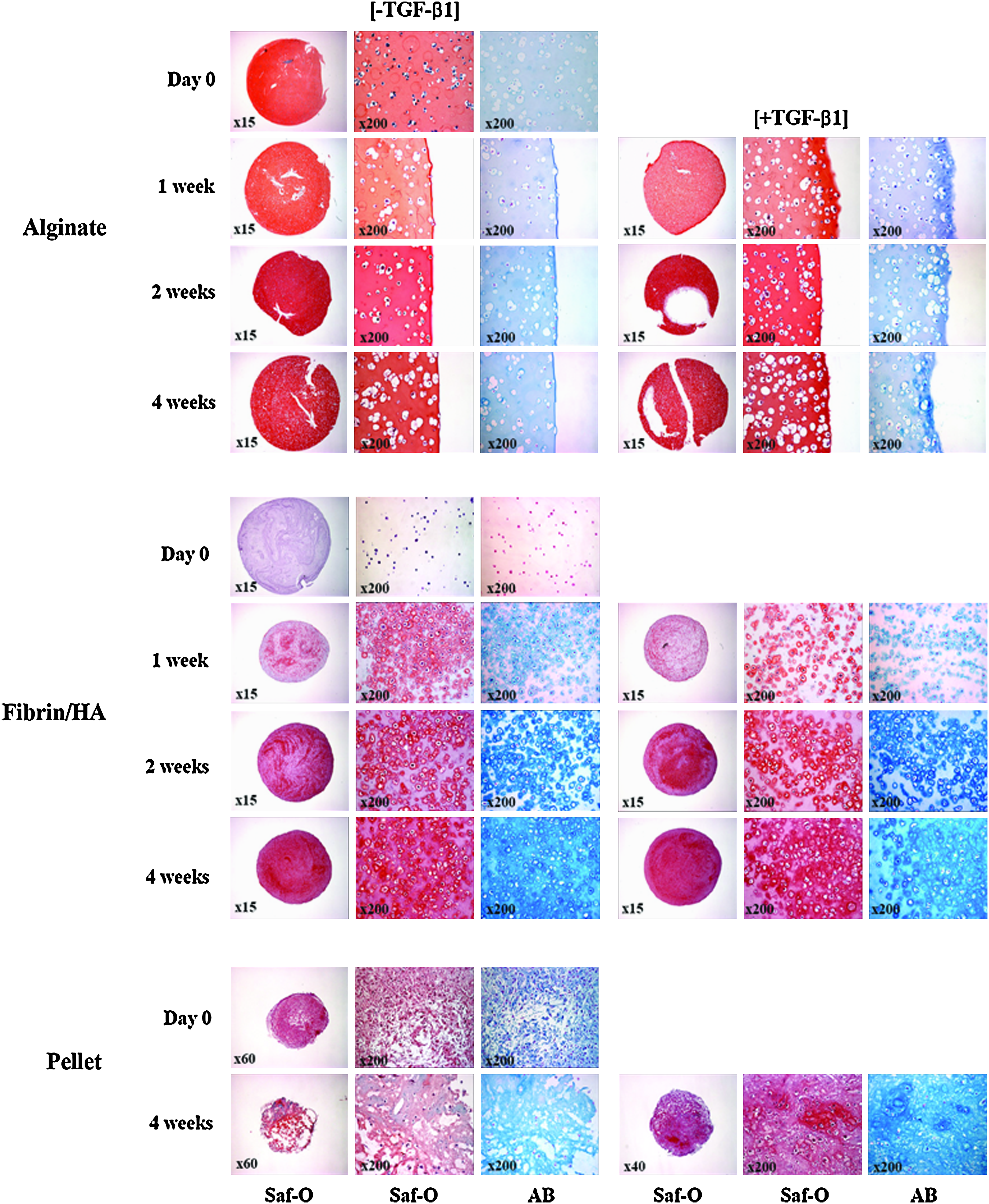
Histological observation of GAG accumulation. rMSCs in alginate, fibrin/HA, and pellet were cultured in CDM with (+TGF-β1) or without (−TGF-β1) 10 ng/mL TGF-β1. Specimens were subjected to Saf O and AB staining at 1 day and 1, 2, and 4 weeks after culture. Whole images of Saf O staining for each group are shown in the leftmost column (low magnification). High-magnification images of Saf-O and AB stains are also presented (×200). GAGs, glycosaminoglycans; Saf O, safranin-O; AB, alcian blue. Color images available online at www.liebertonline.com/tea
Quantitative analysis of GAGs and collagen content
The amounts of sulfated GAGs and collagen were measured in each specimen by chemical assay and presented as μg/mL dry weight (Figs. 4 and 5). In the absence of TGF-β1, the amount of sulfated GAGs increased moderately in the alginate group and only slightly in the pellet group by time, whereas it increased significantly from 1 week in the fibrin/HA group (**p < 0.05, ***p < 0.001) (Fig. 4). TGF-β1 treatment resulted in a clear increase in the amount of GAGs in the alginate (+p < 0.05, ++p < 0.01) and pellet (##p < 0.01) groups; however, no additional effect was shown in the fibrin/HA group compared to the TGF-β1 untreated group. The amounts of GAGs (μg/mg dry weight) in the absence and presence of TGF-β1 at 4 weeks were 53.6 ± 10.9 and 88.5 ± 10.5 in the alginate group (p < 0.01), 25.4 ± 4.1 and 49.8 ± 6.4 in the pellet group (p < 0.001), and 111.3 ± 8.1 and 127.1 ± 9.3 in the fibrin/HA group, respectively.
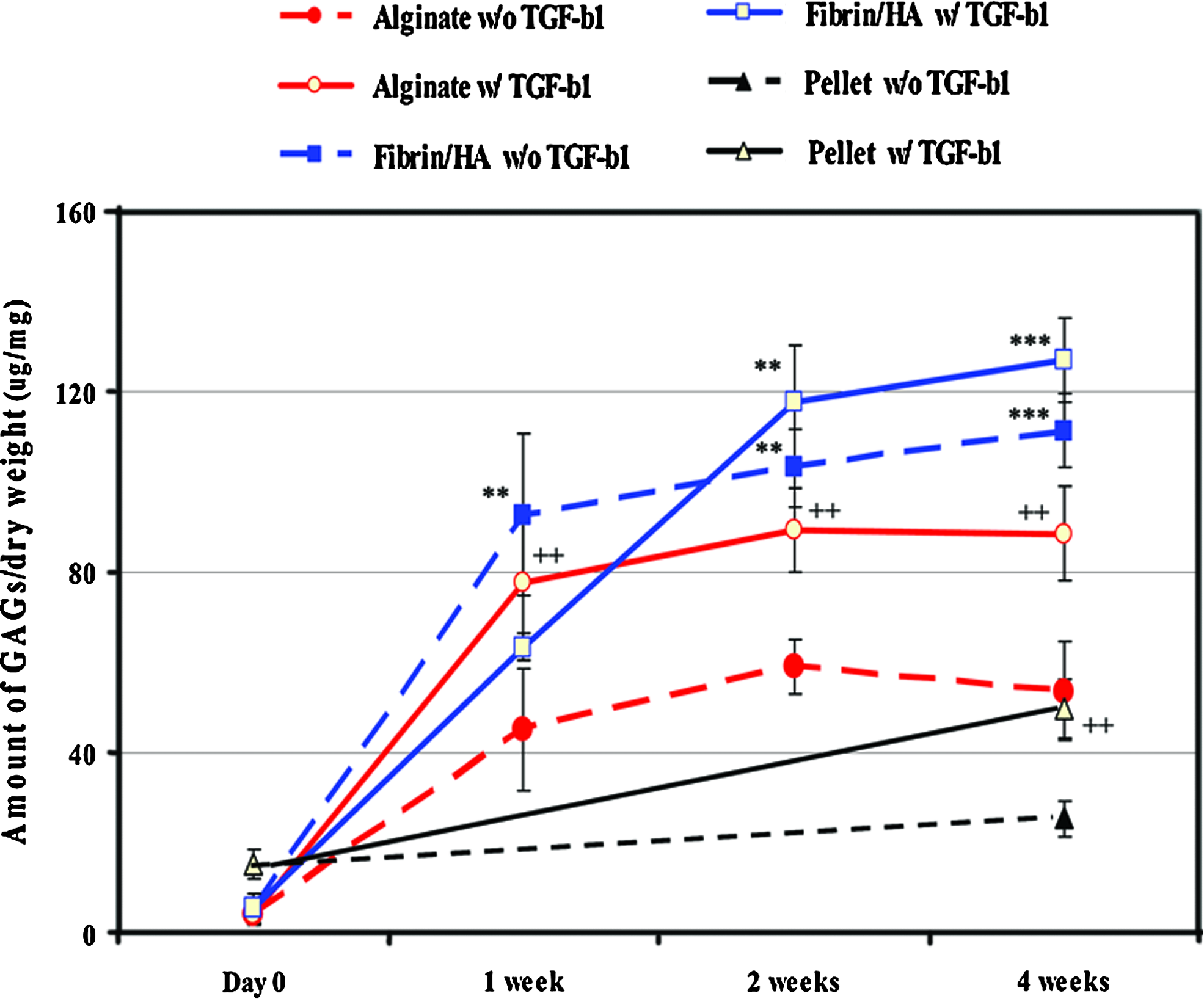
Measurement of GAG contents. Specimens cultured as above were retrieved at the time points indicated and digested in the papain digestion solution, as described in the Materials and Methods section. The amount of GAGs in each specimen was measured by the 1,9-dimethylmethylene blue assay and normalized by its dry weight (μg/mg). Data presented in the line graph represent mean value with SD from three independent experiments. **p < 0.01, and ***p < 0.001, respectively. Asterisk (*) indicates comparison of statistical difference of fibrin/HA gel to other groups. Plus (+) also indicates statistical difference between absent TGF-β1 and present TGF-β1 in the same experimental group. ++p < 0.01. Color images available online at www.liebertonline.com/tea
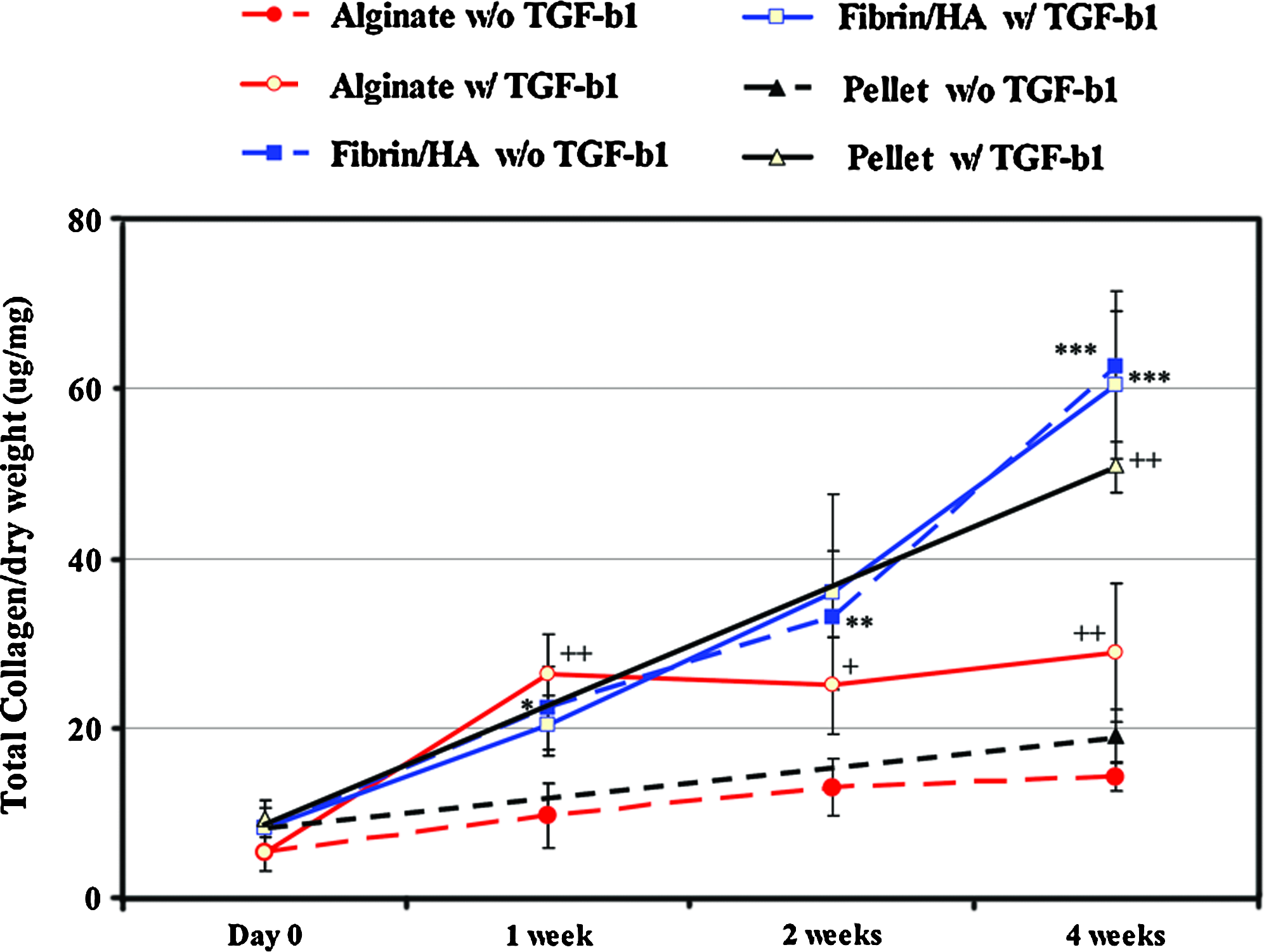
Measurement of collagen content of the specimens. The amount of collagen was measured in each specimen digested with papain solution as above by the Hride Tullberg-Reinert method at the time points indicated. Data are presented as normalized values with SD by the dry weight of each specimen (μg/mg) from three independent experiments. *p < 0.05, **p < 0.01, and ***p < 0.001, respectively. Asterisk (*) indicates comparison of statistical differences in fibrin/HA gel to other groups. Plus (+) also indicates statistical difference between absent TGF-β1 and present TGF-β1 in the same experimental group. +p < 0.05 and ++p < 0.01, respectively. Color images available online at www.liebertonline.com/tea
The amount of collagen also increased depending on TGF-β1 treatment in the alginate and pellet groups, whereas it increased significantly both in the presence and in the absence of TGF-β1 treatment in the fibrin/HA group (Fig. 5). The amount was about two times as much in the fibrin/HA group as in the alginate group in the absence of TGF-β1 (***p < 0.001). The amounts of collagen (ug/mg dry weight) at 4 weeks in the absence and presence of TGF-β1 were 1.4 ± 0.1 and 2.9 ± 0.8 in the alginate group (p < 0.05), 19.1 ± 3.1, and 50.9 ± 3.2 in the pellet group (p < 0.001), and 62.6 ± 3.1 and 60.5 ± 8.7 in the fibrin/HA group, respectively.
Expression of chondrogenic marker genes
To further support results from histological observation, expression of chondrogenic markers, such as aggrecan, type II collagen, and sox 9 was analyzed by reverse transcriptase–polymerase chain reaction (Fig. 6a, b). Expression of GAPDH was examined as a control. In the alginate and pellet groups, mRNA levels of aggrecan, collagen type II, and sox 9 showed a clear or at least slight increase only in the TGF-β1-treated samples. The mRNA level of type I collagen was not significantly different overall by TGF-β1 treatment in the alginate groups and was not clearly detected in the pellet group. In the fibrin/HA group, expression of both chondrogenic marker genes and type I collagen showed a clear increase from 1 week, and did not change significantly with time, regardless of TGF-β1 treatment.

Profile of chondrogenic gene markers.
Analysis of mechanical properties
In the alginate group, the compressive strength (Fig. 7a) and modulus (Fig. 7b) of the alginate group did not change significantly with time in the presence or absence of TGF-β1. In the all fibrin/HA group, however, the compressive strength (Fig. 7c) and modulus (Fig. 7d) both statistically increased with culture. Interestingly, these were somewhat lower in specimens with TGF-β1 treatment in the fibrin/HA group. In addition, fibrin/HA group without TGF-β1 treatment showed 8-folds at 1 week and up to 10-folds at 2 weeks than alginate group without TGF-β1 treatment even though the compressive modulus of the alginate group at day 1 was ∼1.6-folds higher than that of the fibrin/HA group. In the case of the pellet culture, we failed to measure the mechanical strength of the specimens, due to its small size.
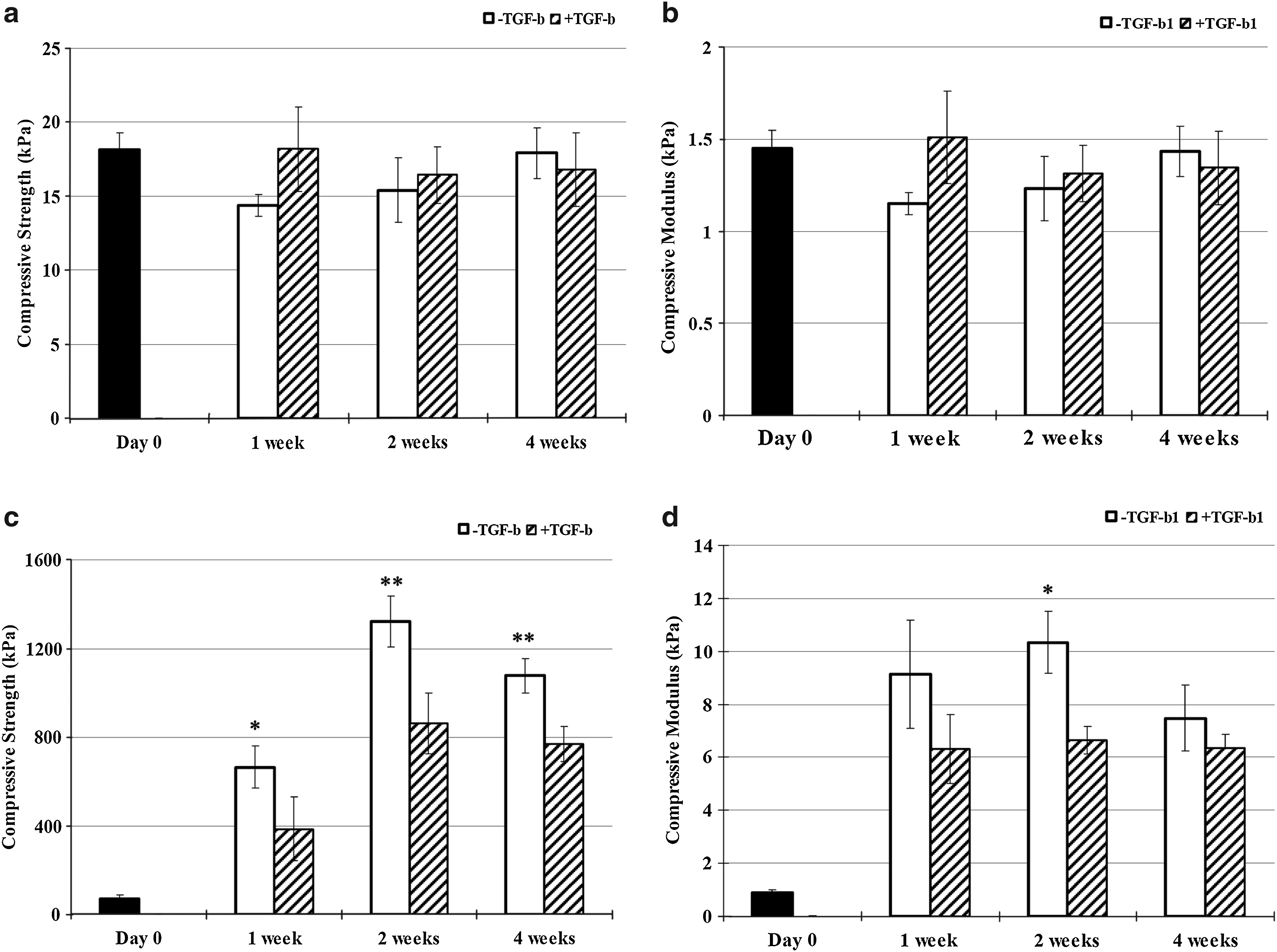
Mechanical properties of the specimens.
Discussion
In this study, we have shown that fibrin/HA composite gel was a promising scaffold supporting chondrogenic differentiation of rMSCs and cartilage-like tissue formation. Constructs in the fibrin/HA gel showed larger sulfated GAG content, higher expression of chondrogenic genes, and better compressive strength than those in the alginate layer and the pellet culture. Pellet and alginate culture systems have been widely used for chondrogenesis of MSCs with successful results.14–16 However, fibrin/HA gel has several strengths over these two culture systems. Fibrin is highly biocompatible and safe for implantation in vivo, which is not fully guaranteed for alginate. HA is a key component of normal cartilage matrix and is expected to provide a favorable environment for chondrogenesis of MSCs. 17 In addition, HA might slow the degradation rate of fibrin, because fibrinogen can bind specifically to HA. 18 Another advantage over alginate and pellet culture is that constructs using fibrin/HA gel can be prepared in a gel type and are easily injectable to the target tissues. Taken together, we speculate that fibrin/HA composite is a highly promising scaffold for cartilage tissue engineering of MSCs in terms of quality of the cartilage tissues produced and its applicability in the clinic.
It is well known that chondrogenic differentiation of MSCs usually requires treatment with growth factors, of which TGF-β is known to be essential.19–21 In vitro or in vivo application of TGF-β upregulates gene expression of chondrogenic markers, produces cartilage-specific ECM molecules, and, in turn, improves repair of full-thickness cartilage defects. 22 However, this study using fibrin/HA gel showed that growth factors like TGF-β might not be essential for chondrogenesis of MSCs. Similar results have also been reported in many previous studies by our group and others using scaffold materials,23,24 and even mechanical stimulation.25,26 In our previous studies, a natural scaffold composed of ECM produced by chondrocytes was successfully used for support of chondrogenic differentiation and cartilage tissue formation using rMSCs and chondrocytes without TGF-β treatment. 23 Chondrogenic differentiation of rMSCs and human MSCs was induced efficiently in vitro and in vivo by mechanical stimulation using low-intensity ultrasound10,27,28 and cyclic compressive loading. 29 These results are supported by those from studies conducted by other groups.30,31 Gelatin-chitosan hybrid materials to mimic the natural environment led to increased ECM synthesis and promoted differentiation of stem cells. 30 Adult rat MSCs showed upregulated extracellular signal-regulated kinases activation in the collagen–GAG blend scaffold, which provided a suitable 3D environment. 31 Compressive forces were shown to promote expression of chondrogenic marker genes and chondrogenesis in mouse embryonic limb bud MSCs10,27,28 and rabbit bone-marrow MSCs. 32 However, conflicting results were reported by Sharma et al. 33 in their study using TGF-β3 and HA; TGF-β3 was shown to be essential for proteogylcan production. Differences in results might be because they undertook the experiment in vivo and used materials that were different from ours. Overall, these results suggest that growth factors like TGF-β might not be essential if optimal conditions for chondrogenesis of MSCs are provided by other means. Differentiation of MSCs without use of biologic factor(s) would be very beneficial, resulting in prevention of a potential harmful effect and reduction of production costs for therapy.
The mechanism of fibrin/HA gel in facilitation of chondrogenic differentiaton of MSCs independently with TGF-β is not clear at this point, but probably involves cellular signal transduction pathways that are mediated by interaction of cells with the ECM environment of fibrin/HA. Cells interact with the surrounding ECM via integrins on the cell surface, which transmit intracellular signals by themselves or by cooperation with nearby growth factor receptors.34,35 Interaction of MSCs with ECM has been shown to induce cellular signals for regulation of cell function and matrix accumulation within tissue-engineered constructs.30,31 Kinases and other signaling molecules associated with focal complexes appeared to mediate transducing signals from the site of cell–ECM interaction. 36 In addition, type II collagen hydrogels have been shown to enhance TGF-β1-induced chondrogenesis of bovine MSCs; however, type I collagen and alginate gels did not. 37 Fibrin/HA composite gel can also mechanically couple itself to MSCs and stimulate cellular signals directly or via upregulation of nearby receptors, which eventually increases chondrogenic differentiation of MSCs. Fibrin has peptide domains that facilitate cell adhesion to the clot, serving as bonding sites for integrin receptors on cells involved in tissue repair. 38 HA also provides a binding site to CD44, which is expressed by chondrocytes. In particular, HA has been shown to influence many processes within the ECM, including matrix assembly, cell proliferation, cell migration, and embryonic tissue development.39,40
The pellet and the alginate culture systems have been widely used for the chondrogenesis of MSCs and have shown promising results of producing cartilage-like engineered tissues.14–16 However, we had some limitation for analysis using these system. In the case of pellet culture, we failed to measure the mechanical strength of specimens due to their small size. In contrast, the fibrin/HA composite gel showed a better mechanical property and uniform cell distribution as well as efficient chondrogenic differentiation of MSCs. The alginate gel also has a limitation of showing the same background color as that of positive signal in the safranin-O staining. The fibrin/HA gel has an additional strength. It is easy to handle because cells can be just mixed with the composites before polymerization to make a solid construct. Also, when combined with its injectable property, it would be one of the most convenient ways to deliver cells into living organs.
In conclusion, this study showed that fibrin/HA composite gel is a promising 3D scaffold for inducing chondrogenic differentiation of rMSCs in vitro. When compared with the alginate layer and pellet culture systems, it provided a stable physical property and an efficient method for inducing chondrogenesis of MSCs, even without treatment of TGF-β. Due to easy manipulation and proven biocompatibility of fibrin and HA, fibrin/HA gel could be used in a variety of therapeutic applications for cartilage repair.
Footnotes
Acknowledgment
This work was supported by a grant from the Korea Healthcare Technology R&D Project, Ministry for Health, Welfare and Family Affairs, Republic of Korea (A091120).
Disclosure Statement
No competing financial interests exist.