
Abstract
The aim of this study was to evaluate whether tissue-engineered skin produced in vitro was able to sustain growth of hair follicles in vitro and after grafting. Different tissues were designed. Dissociated newborn mouse keratinocytes or newborn mouse hair buds (HBs) were added onto dermal constructs consisting of a tissue-engineered cell-derived matrix elaborated from either newborn mouse or adult human fibroblasts cultured with ascorbic acid. After 7–21 days of maturation at the air–liquid interface, no hair was noticed in vitro. Epidermal differentiation was observed in all tissue-engineered skin. However, human fibroblast-derived tissue-engineered dermis (hD) promoted a thicker epidermis than mouse fibroblast-derived tissue-engineered dermis (mD). In association with mD, HBs developed epithelial cyst-like inclusions presenting outer root sheath-like attributes. In contrast, epidermoid cyst-like inclusions lined by a stratified squamous epithelium were present in tissues composed of HBs and hD. After grafting, pilo-sebaceous units formed and hair grew in skin elaborated from HBs cultured 10–26 days submerged in culture medium in association with mD. However, the number of normal hair follicles decreased with longer culture time. This hair-forming capacity after grafting was not observed in tissues composed of hD overlaid with HBs. These results demonstrate that epithelial stem cells can be kept in vitro in a permissive tissue-engineered dermal environment without losing their potential to induce hair growth after grafting.
Introduction
Dermal papilla at the base of the hair follicle is recognized as a key orchestrator of the hair development and their cyclical growth.12,13 In the way of producing new hair follicles, the first attempts were performed in vivo and consisted to insert dermal papillae or cultured dermal papilla cell condensations in proximity of an epithelium.14–19 Another well-used cellular-based approach is the deposition of mouse neonate hair buds (HBs) or keratinocytes combined with uncultured newborn dermal fibroblasts to the dorsal muscle of recipient mouse within a silicon chamber.20–24 Similar cell preparations injected directly into the skin dermis also reform hair follicles. 25
Few hair induction assays involving tissue-engineered matrices have been previously reported. Mouse HBs or human dermal papillae combined with dissociated keratinocytes cultured in collagen gel formed tubular structures but no complete hair follicle.10,26 Human outer root sheath (ORS) cells combined with dermal papilla cells in matrigel gave rise to epidermoid cyst-like inclusions. 27 Even if in these in vitro models, the hair-derived structures present some trichogenic-like features, it is not clear whether normal hairs could actually be formed once the whole tissue is grafted in vivo.
The self-assembly approach is a tissue-engineering method developed at the LOEX laboratory.1,28 It allows the reconstruction of living tissues such as tissue-engineered blood vessels, bladder, cornea, skin, or adipose tissue.7,8,28–35 Dissected human pilo-sebaceous units can be integrated within tissue-engineered skin in vitro. 7
We report herein that the stem cell potential to differentiate into normal hair follicles can be preserved in vitro in the tissue-engineered skin made by the self-assembly approach. Indeed, when appropriate cell populations and culture conditions were used, hair growth was induced after grafting. This work also reveals that the tissue-engineered dermal environment has an impact on the differentiation commitment of stem cell progeny.
Materials and Methods
Tissues and cells
All protocols were approved by the institution's animal care and use committee (Comité de Protection des Animaux de Laboratoire de l'Université Laval, Québec, Canada) and by the institution's committee for the protection of human subjects (Comité d'Éthique de la Recherche du Centre Hospitalier affilié Universitaire de Québec).
Dissociated mouse keratinocytes (dK) were extracted from the epidermis of C3H/HeN newborn (<1 day of age) mouse skin using the method by Yuspa and Harris,
36
with slight modifications. The cells from the epidermal portion were washed in complete keratinocyte medium (cDME-Ham)—a combination of Dulbecco-Vogt modified Eagle's medium (DME) (Invitrogen, Oakville, Canada) and Ham's F12 (Invitrogen) in a 3:1 proportion, supplemented with 24.3 μg/mL adenine (Sigma, Oakville, Canada), 5 μg/mL insulin (Sigma), 2 × 10–9 M 3,3′,5′-triiodo-
Preformed hair follicles, referred to as HBs (from <1-day-old C3H/HeN mice), and mouse fibroblasts (from 2-day-old FVB mice) were obtained by collagenase digestion of the dermal portion remaining after epidermal retrieval of trypsinized skin as previously described.20,21 Three low-speed centrifugations (37 g, 3 min) in cDME-Ham were performed to sediment HBs. The supernatent was removed and set aside, whereas the HB pellet was resuspended in cDME-Ham. Mouse fibroblast-containing supernatant was filtered through a 40-μm cell strainer (BD Falcon). The cells of the filtrate were pelleted by centrifugation (300 g, 10 min). Human fibroblasts were obtained from the dermal portion of adult breast skin biopsies as previously described. 37 Mouse and human fibroblasts were subcultured in DME supplemented with 10% fetal calf serum (FCS; HyClone) and cryopreserved in 90% FCS (HyClone) and 10% dimethyl sulfoxide (Sigma) as previously described. 37 All cultures were incubated at 37°C in a 8% CO2-air atmosphere and the culture media were changed three times a week.
Tissue-engineered skin production and grafting
Murine or human fibroblasts were thawed and used between their 4th and 8th passages for the reconstruction of a 3D extracellular matrix, which was used as dermal substitute. Fibroblasts from two litters of mice (2 days old) and human subjects (21 and 23 years old) were used. Briefly, fibroblasts were cultivated 28–35 days in DME with 10% FCS supplemented with 50 μg/mL of ascorbic acid (Sigma). According to the self-assembly approach of tissue engineering,7,8,37 fibroblast-derived matricial sheets were superimposed (2 or 4 for in vitro experiments and 4 for in vivo experiments) to obtain a thicker 3D matrix (referred as to tissue-engineered dermis) onto which epithelial cells were seeded 1 week later to obtain the tissue-engineered skin. Ten percent of the HBs freshly collected from each newborn mouse per cm2 or 600,000 freshly extracted dK per cm2 were seeded on the tissue-engineered dermis. To study maturation in vitro, the resulting tissue-engineered skin was cultured 7 days submerged in cDME-Ham and then raised at the air–liquid interface in epidermal growth factor-free cDME-Ham. Biopsies were taken after 7, 14, and 21 days of culture at the air–liquid interface. Five to 13 tissue-engineered skins were produced for each condition.
To evaluate the maturation of the tissue-engineered skin in vivo, the tissue-engineered skin cultured submerged in cDME-Ham for 10, 14, 21, or 26 days were grafted onto the dorsal muscular fascia of adult athymic nu/nu mice (Charles River Laboratories, Wilmington, MA) as previously described. 8 As positive control, freshly extracted newborn mouse HBs (50% of the HBs collected from each newborn dermis) were inserted through a 1-cm-diameter silicon grafting chamber placed on the dorsal muscular fascia of athymic nu/nu mice as described by Lichti. 20 Biopsies were taken after 6 weeks or 6 months. Five to 13 mice were grafted for each condition except for the 6-month experiment in which 3 mice were grafted for each condition. Biopsies were embedded in Tissue-Tek® OCT Compound (Miles, Inc., Elkhart, IN), cooled in liquid nitrogen, and conserved at −80°C or processed for histological analysis.
Histological analysis, Oil Red O staining, and endogenous phosphatase activity detection
For histological analysis, the biopsies were fixed with Histochoice™MB® fixative (Amresco, Solon, OH) and embedded in paraffin. Six-micrometer-thick sections were processed for Masson's trichrome staining38,39 using Weigert's hematoxylin, fuchsin-ponceau, and aniline blue stains. To reveal lipid droplets within the tissues, cryosections were stained with Oil Red O, and Harris's hematoxylin was used to counterstain nuclei. The endogenous alkaline phosphatase (AP) activity was detected by the Handjiski's method, 40 followed by Harris's hematoxylin counterstaining.
Immunolabeling
Indirect immunofluorescence staining was performed on 5-μm-thick acetone-fixed (10 min at −20°C) frozen sections of the tissues as previously described. 41 Primary antibodies used were rabbit polyclonal anti-keratin 5 (AF 138; BabCO, Richmond, CA), anti-collagen IV (Millipore, Temecula, CA), anti-loricrin (AF 62; Covance, Princeton, NJ), anti-involucrin (Covance), guinea pig anti-K19, 41 anti-K15 (gp 15.1; Progen Biotechnik, Heidelberg, Germany), anti-β-catenin (Cell Signaling Technology, Beverly, MA), rat monoclonal anti-laminin 5 (Immunotech, Westbrook, ME), and goat anti-Lef1 (Santa Cruz Biotechnology, Santa Cruz, CA). The following secondary antibodies were used: goat anti-rabbit conjugated with Alexa 488 (Invitrogen) or rhodamin (Millipore), goat anti-guinea pig conjugated with FITC (fluorescein; Jackson Immunoresearch Laboratories, West Grove, PA), chicken anti-goat IgG conjugated with Alexa 488 (Invitrogen), or goat anti-rat conjugated to Alexa 594 (Invitrogen). In some instances, Hoechst staining was performed to visualize nuclei.
For immunoperoxidase staining, slides were sequentially incubated with formaldehyde (0.37%, 10 min), methanol (100%, 10 min), and NaOH (0.07N, 15 s) and the Vector® M.O.M Basic Kit (ID Labs, London, Canada) was used to reduce nonspecific staining arising from endogenous mouse immunoglobins in the tissue. Then, the sections were labeled with the following mouse monoclonal antibodies: anti-K10 (RKSE60; Cedarlane Laboratories, Burlington, Canada), anti-K20 (IT-Ks 20.10) (American Research Products, Belmont, MA), anti-Ki-67 (BD Biosciences, San Diego, CA), anti-AE13 (Abcam, Cambridge, MA), and anti-AE15 (Abcam) and were revealed using the Ultra HRP peroxidase detection system (ID Labs). Peroxidase activity was revealed with 3,3′-diaminobenzidine (Sigma). Each incubation was separated by a washing step with phosphate-buffered saline. Nuclei were counterstained with Harris's hematoxylin. The slides were examined under a Nikon Eclipse E600 microscope (Nikon) equipped with a SenSys® digital camera (Photometrics®, Tucson, AZ) for immunofluorescence pictures and a Coolsnap™ digital camera (Photometrics) for color pictures. Multicolored pictures and adjustments of the dynamic range were made using Adobe® Photoshop 7.0 software (San Jose, CA).
Western blot
On ice, the tissue-engineered skins were fragmented into small pieces, lysed in Tris-HCl buffer containing 1% sodium dodecyl sulfate, 10% glycerol, and Complete™ protease inhibitor cocktail (Roche Diagnostics), homogenized (27,000 rpm, 30 s) using Polytron PT 3100 (Brinkmann, Mississauga, Canada), and sonicated with Sonifer 250 (Branson, Danbury, CT). The total protein concentration in the lysates was measured using a Micro BCA Protein Assay Reagent kit (Pierce, Rockford, IL). Proteins were separated under reducing conditions by 10% polyacrylamide gel electrophoresis, and immunoblotting was performed as previously described. 42 Primary antibodies used were mouse anti-AE13 (Abcam) and rabbit anti-K14. 43 Secondary antibodies were goat anti-mouse (Millipore) and goat anti-rabbit (Sigma) conjugated to horseradish peroxidase. Immunoreactive complexes were revealed with enhanced chemiluminescence detection reagent (Amersham, Baie d'Urfe, Canada) and images were captured using the Fusion FX7 acquisition system (Vilbert Lourmat, Eberhardzell, Germany).
Epidermal thickness measurement and statistical analysis
The thickness of the epidermal portion of the tissue-engineered skin was evaluated on histological slide pictures (magnification: 20 × ) using Metaview software version 4.5.R2 (Universal Imaging Corp™, Downingtown, PA). A minimum of 50 measures were taken for each sample. A total of 5–13 tissue-engineered skin were produced per condition (n = 5–13). For statistical analysis, the unilateral Mann–Whitney test was used.
Results
Epidermis and hair follicle formation in tissue-engineered skin cultured in vitro
To evaluate the capacity of epithelial cells to generate hair follicles within tissue-engineered skin produced by the self-assembly approach, dK or neonate mouse HBs were seeded onto dermal constructs obtained after culturing fibroblasts with ascorbic acid. Tissue-engineered dermis elaborated from mouse (referred to as mD) or human (hD) fibroblasts were tested. Maturation of tissue-engineered skin was followed in vitro. After culturing at the air–liquid interface for 7–21 days, dK formed a fully differentiated epidermis when cultivated on mD or hD (Fig. 1A–D). The nuclear antigen associated with cell proliferation, Ki-67, was present in the basal cells of the epidermis (Fig. 2A, B, arrows). The newly generated epidermis of dK/hD was significantly thicker than that of dK/mD after 14 and 21 days of culture at the air–liquid interface (Fig. 3), suggesting a differential influence of mouse and human tissue-engineered dermis on the proliferation/differentiation balance of the overlying epidermal cells. To evaluate whether a thick epidermis arises as a mere consequence of a thick tissue-engineered dermis, Pearson's coefficient correlation between dermal and epidermal thickness measurements in a given sample was determined. No significant correlation was noted (r = 0.22, n = 97).
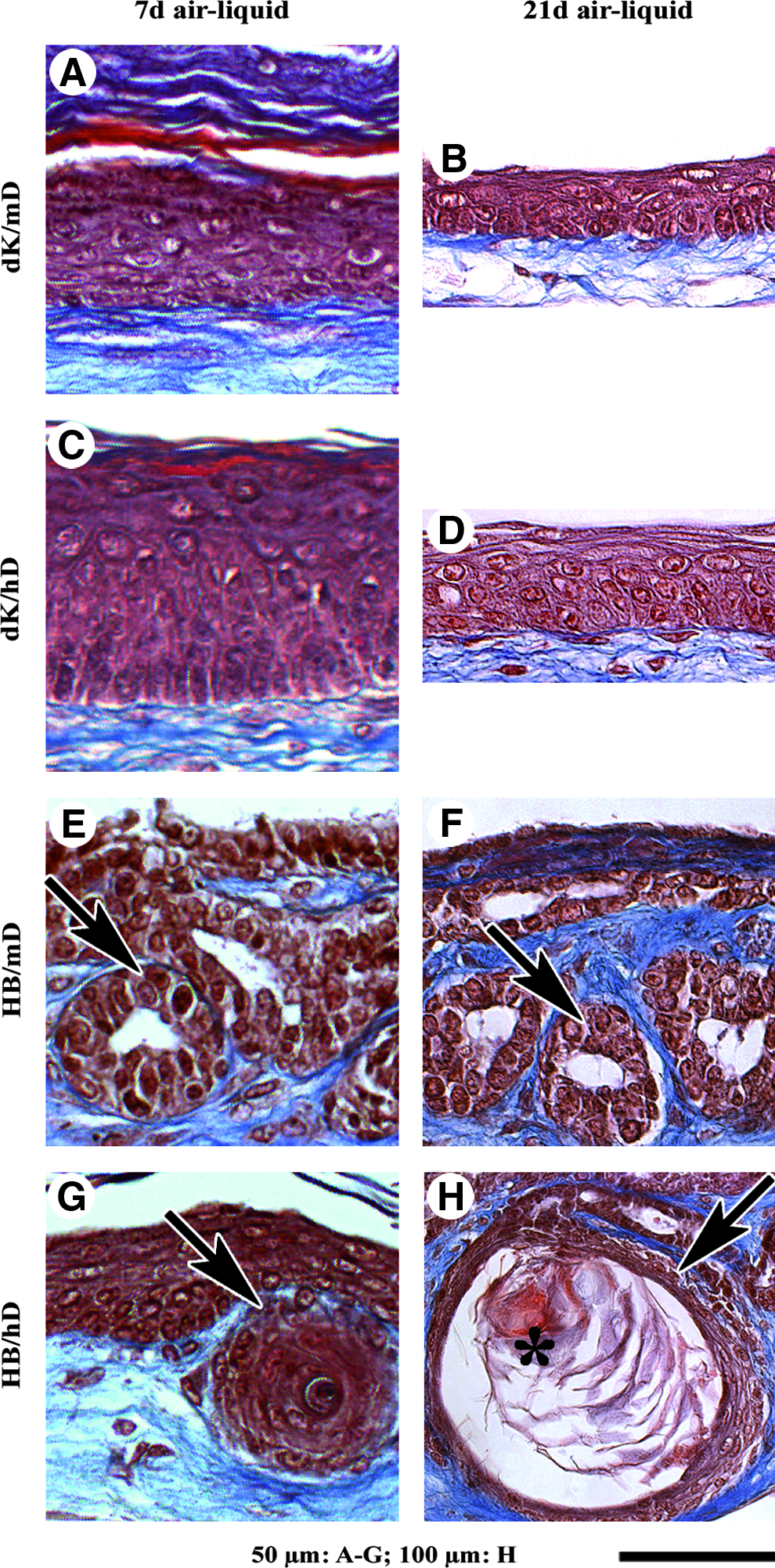
Histological analysis of tissue-engineered skin matured in vitro. Masson's trichrome staining of tissue-engineered skin produced from dissociated mouse keratinocytes (dK) or hair follicle buds (HBs) seeded on mouse or human fibroblast-derived tissue-engineered dermis (mD or hD, respectively) matured for 7

Tissue-engineered skin matured in vitro immunolabeled for skin markers. Tissue-engineered skin produced from dK or HBs seeded on mD or hD matured for 7

Measurement of epidermal thickness. The epidermis of tissue-engineered skin was significantly thicker when dK were cultured on hD compared with mD for 14 or 21 days at the air–liquid interface as well as 6 weeks (42 days) after grafting on an athymic mouse. Data represent the average epidermal thickness (μm) for each tissue-engineered skin condition, ± standard error of the mean. The number of independent experiments (n) is indicated. *p ≤ 0.05; **p ≤ 0.01.
In addition to the epidermis, histological analysis of HB/mD and HB/hD matured in vitro revealed the presence of epithelial cyst-like inclusions in their dermal portion (Fig. 1E–H, arrows). HB/hD epithelial inclusions contained keratinocytes resembling a stratified squamous epithelia. Indeed, basal cuboidal cell nuclei expressing the proliferation-associated Ki-67 44 were located at the cyst periphery (Fig. 2D, arrows), followed inward by spinous and granular-like layers (Fig. 2D, open arrows). The innermost cell layers of the HB/hD epithelial inclusions were formed by anucleate squames detaching in the center (Figs. 1H and 2D, asterisks). The size and differentiation of HB/hD epithelial inclusions appeared to increase with time (Fig. 1, compare G with H), probably as a result of cell proliferation and differentiation. In contrast, HB/mD epithelial inclusions were epithelium-lined cavities maintaining relatively the same size after 21 days of culture at the air–liquid interface (Fig. 1, compare E with F). This feature did not seem associated with a proliferation defect because Ki-67–labeled nuclei were present in several basal cells in the HB/mD epithelial inclusions (Fig. 2C, arrows). However, the innermost differentiated epithelial cell layers typical of a mature hair follicle were not observed.
To further characterize maturation of the tissue-engineered skin, immunolabeling with antibodies directed against proteins associated with different skin compartments was performed. Laminin-5 (Fig. 2E–G, arrows) and type IV collagen (data not shown), two basement membrane components, were detected at the dermo-epidermal junction in all tissue-engineered skins, reflecting the cohesion between epidermis and dermis. In HB/mD and HB/hD, laminin-5 (Fig. 2G and H, open arrows) and type IV collagen (data not shown) were also detected around the epithelial inclusions. K5, which is mainly expressed in basal keratinocytes, 45 was found in the basal cells of the epidermis of the tissue-engineered skin (Fig. 2I–K, arrows) as well as in the cells at the periphery of the epithelial inclusions of HB/mD and HB/hD (Fig. 2K, L, open arrows). As expected, K10 was expressed in the suprabasal layers of the epidermis of the dK/mD and dK/hD conditions (Fig. 2M, N, arrows), whereas loricrin was restricted to the granular and cornified layers (Fig. 2Q, R, arrows). Interestingly, neither K10 (Fig. 2O) nor loricrin (Fig. 2S) was detected in HB/mD epithelial inclusions. In contrast, K10 and loricrin were detected in the internal layers of the HB/hD ones (Fig. 2P, T, respectively, open arrows), suggesting an epidermal differentiation.
We next evaluated the expression of K15 and K19, two known follicular stem cell markers.41,46–48 The K15 expression was higher in the basal cells of the epidermis (Fig. 2U–X, arrows) and in the epithelial inclusions (Fig. 2W, X, open arrows) of the tissue-engineered skin. In the epidermis of dK/mD and dK/hD, K19 was occasionally identified in some basal keratinocytes even after 21 days of culture at the air–liquid interface (Fig. 2Y, Z, arrows), suggesting that stem cells were conserved in the tissue-engineered skin. Interestingly, K19 was expressed in the majority of basal cells in HB/mD epithelial inclusions (Fig. 2AA, open arrow). In contrast, only few K19-expressing cells were observed in the periphery of epithelial inclusions in HB/hD (Fig. 2BB, open arrow). However, HB/mD epithelial inclusions lacked typical inner root sheath (IRS) and hair cortex structures. Accordingly, absence of the hair cortex-associated keratin expression in the four tissue-engineered skins cultured in vitro was confirmed by western blotting using AE13 antibody (Fig. 4A).

Tissue-engineered skin produced in vitro can result in hair growth after grafting onto athymic mice
To test whether the ability to form hair was preserved within tissue-engineered skin despite the failure of hair growth in vitro, they were grafted in vivo. The four tissue-engineered skins, dK/mD, dK/hD, HB/mD, and HB/hD, were maturated 10 days submerged in culture medium and then transplanted onto athymic mice. As positive control, freshly extracted newborn mouse HBs were transplanted directly onto the muscle fascia on the back of athymic mice (without tissue-engineered matrix underneath) according to the method described by Weinberg et al. 21 At about 1 month after grafting, pigmented hair follicles typical of the C3H/HeN mice fur emerged on the grafted site of the control mice (Fig. 5A) as well as on the HB/mD grafts (100% of 13 grafted mice; Fig. 5C, D). Few hairs were observed in dK/mD (Fig. 5B, arrows). The hairs were maintained for at least 6 months (100% of three mice grafted with HB/mD; data not shown), indicating that a functional pool of epithelial stem cells resides in the grafted tissue-engineered skin. Macroscopic examination of dK/mD (Fig. 5B, asterisk), dK/hD (not shown), and HB/hD (Fig. 5E, asterisk)-grafted sites revealed pigmentation (100% of 10, 8, and 8 grafted mice, respectively), confirming the C3H/HeN origin of epithelial cells. No external hair was visible at the surface of dK/hD (not shown) and HB/hD (Fig. 5E). At a histological level, the epidermis of dK/hD was generally thicker than that of dK/mD at 6 weeks after grafting (Fig. 3 and compare Fig. 5G with 5F). In dK/mD tissue sections, numerous hair follicles containing black pigments in their cortex (Fig. 5H, asterisks) and annexed sebaceous glands (Fig. 5H, open arrow) containing cytoplasmic lipid droplets stained by Oil Red O (Fig. 5K, arrows) were observed. In HB/hD, epithelial cyst-like inclusions composed of epithelial cells surrounding enucleate material (Fig. 5I, arrow) or forming an empty cavity (Fig. 5I, open arrow) were observed instead of a structure presenting the typical characteristics of a mature hair follicle. Epidermal invaginations were also present (Fig. 5J).
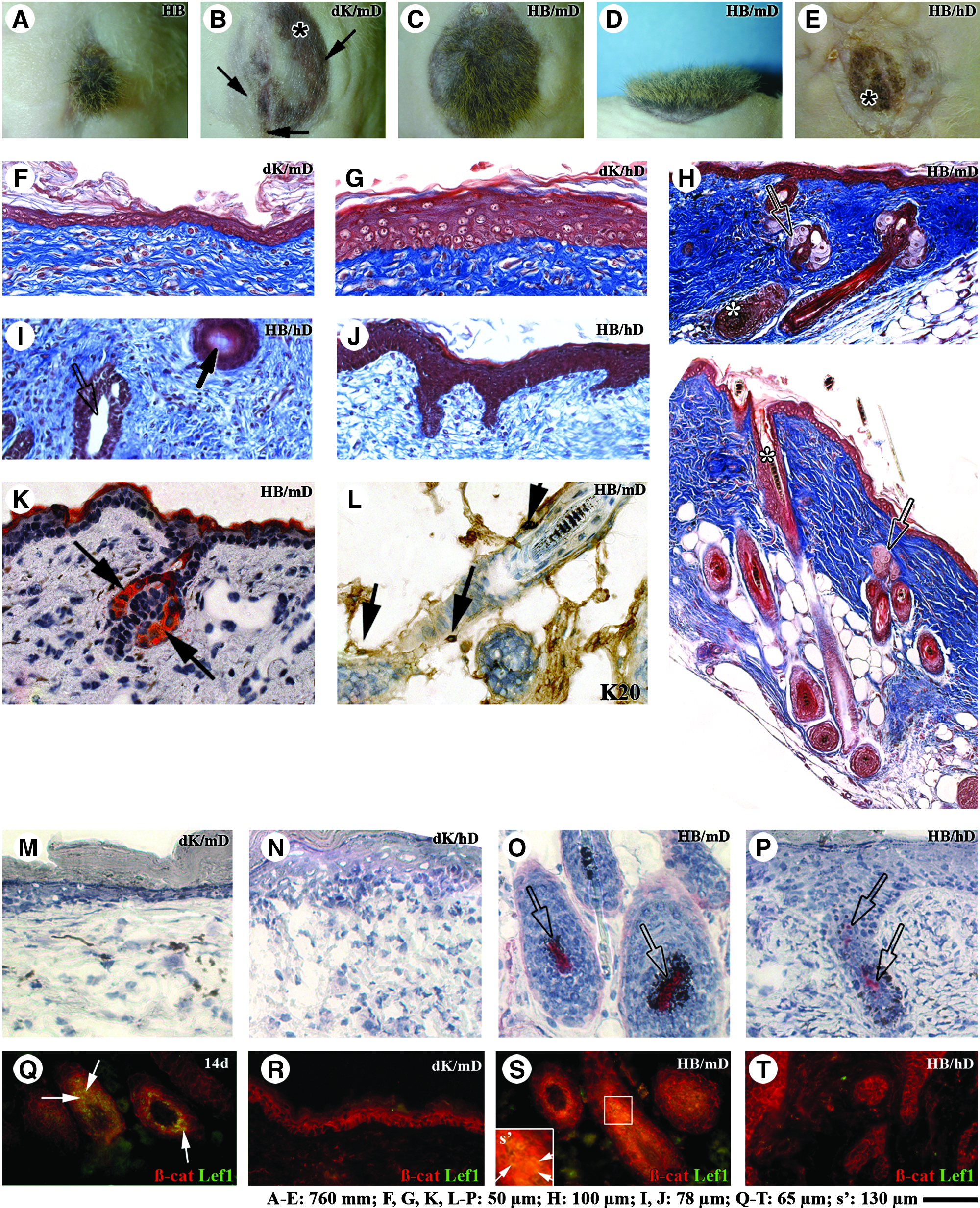
Tissue-engineered skin matured in vivo.
Considering the tactile function of hair follicles, we evaluated the presence of Merkel cells in grafted HB/mD using an antibody directed against K20.49,50 K20-expressing cells were found around the hair follicles (Fig. 5L, arrows), indicating that Merkel cells were present within hair follicles.
The pioneer study of Oliver and Jahoda showed the importance of dermal papilla cells for the orchestration of hair growth.12,13 Given that dermal papilla fibroblasts are known to express AP, 40 we probed the activity of this enzyme in the grafted tissue-engineered skin. Endogenous AP activity was not detected in the grafted tissue-engineered skin made from dK with mD (Fig. 5M) or with hD (Fig. 5N). In contrast, aggregates of cells similar to dermal papilla presenting a strong AP activity were detected in HB/mD (Fig. 5O, open arrows) as well as in HB/hD (Fig. 5P, open arrows), although this latter staining was lower. These observations suggested that the lack of mature hair follicles in HB/hD was not caused by the absence of dermal papilla cells, which remained associated with epithelial cells in HBs.
To better understand the influence of mD and hD on the differentiation of epithelial cells in the grafted tissue-engineered skin, frozen sections (6 weeks postgrafting) were stained with antibodies directed against K10 and involucrin, two proteins associated with the epidermal differentiation program.45,51 In all grafts, K10 was detected in suprabasal layers of the epidermis (Fig. 6A–D, arrows), whereas involucrin expression was restricted to cells of the granular and cornified layers (Fig. 6E–H, arrows). In the hair follicles of HB/mD grafts, K10 (Fig. 6C, I, open arrows) and involucrin (Fig. 6G, J, open arrows) were detected in the infundibulum area. K10 and involucrin expression was noted in some suprabasal cells of the epithelial inclusions within HB/hD (Fig. 6D, H, respectively, open arrows). In the HB/mD grafts, the cortex and IRS of the hair follicles strongly stained with the AE13 and AE15 antibodies, respectively (Fig. 6K, L, respectively, arrows). No significant AE13 labeling was observed in the HB/hD grafts (Fig. 6M), whereas a slight signal in suprabasal cells of the epidermis was observed with the AE15 antibody (Fig. 6N, arrows). Hair cortex differentiation within the dK/mD, dK/hD, HB/mD, and HB/hD grafts was also investigated by western blotting using the anti-AE13 antibody. Signals were detected in dK/mD and HB/mD grafts at the level of 44–46 kDa (Fig. 4B, lanes 3 and 5), whereas the signal was barely detectable in the dK/hD and HB/hD grafts (Fig. 4B, lanes 4 and 6). Collectively, these observations show that the tissue-engineered skin with a mouse fibroblast-derived tissue-engineered dermis can sustain the formation of hair follicles from HBs after grafting. In contrast, cells composing HBs adopted epidermal characteristics rather than differentiating into hair follicles when they were in contact with the human fibroblast-derived tissue-engineered dermis. Thus, the hair differentiation potential was maintained over a 10-day culture period in HB/mD but not in HB/hD.
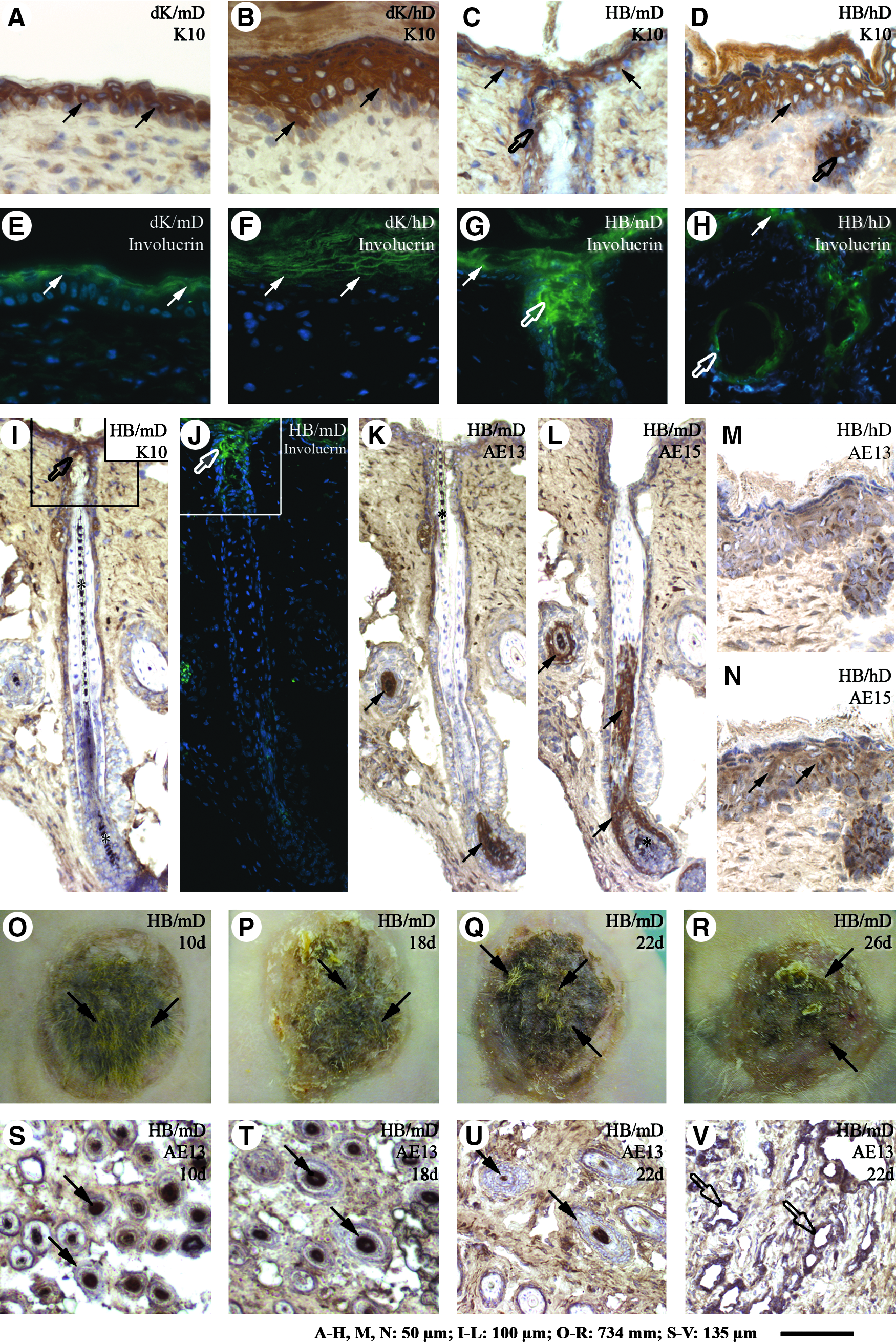
Tissue-engineered skin matured in vivo immunolabeled for skin markers. Cross-sections of grafted (6 weeks) tissue-engineered skin produced with either dK
The Wnt signaling operates as a key regulator of hair follicle differentiation (for review, see ref. 52 ). Thus, the expression of β-catenin and Lef1 in grafted tissue-engineered skin was evaluated by immunofluorescence. As expected, β-catenin was detected at epithelial cell–cell junctions (Fig. 5Q–T). However, its nuclear expression was only observed in the matrix cells of the hair follicles of grafted HB/mD (Fig. 5S, arrows). Further, nuclear β-catenin colocalized with the transcription factor Lef1 strictly within these hair follicle matrix cells (Fig. 5S, s', arrows), as normally observed in normal mouse skin in situ (Fig. 5Q, arrows). In contrast, β-catenin and Lef1 were absent in the nucleus of cells surrounding dermal papilla in HB/hD at 6 weeks after grafting (Fig. 5T), suggesting an interruption of the Wnt signaling pathway.
We next evaluated how long the HB/mD can be kept in culture before transplantation without losing the potential to form hairs after grafting. HB/mD culture time was extended to 18, 22, and 26 days. After grafting, hair appeared in all grafts; however, their number decreased with the augmentation of culture time prior to grafting (Fig. 6O–R, arrows). Accordingly, the expression of the hair cortex keratin marker AE13 was importantly reduced in the HB/mD grafts cultured 26 days submerged in culture medium as revealed by western blot analysis (Fig. 4B, lane 10). AE13 immunolabeling of tissue sections confirmed the presence of histologically normal hair follicles presenting hair cortex differentiation in their innermost cell layers in the HB/mD grafts (Fig. 6S–U, arrows). However, abnormal epithelium-lined cavities were also noted in the dermis of HB/mD, especially in those that have been cultivated for over 18 days before being transplanted (Fig. 6V, open arrows). Altogether, these results indicate that the stem cells of the HBs cultured in vitro within a tissue-engineered environment, with the appropriate fibroblasts and culture conditions, maintain their potential to differentiate into hair and cutaneous appendages such as sebaceous glands, even when grown beyond 26 days.
Discussion
Tissue engineering technology takes advantage of the capacity of cells to form a tissue under appropriate culture conditions and represents a promising research field to elucidate interactions between various cell types and their microenvironment. It is acknowledged that the 3D environment contributes to the cell state modulation.53–56 In this study, tissue-engineered models were designed to examine whether epithelial–mesenchymal interactions present in tissue-engineered skin are permissive to epithelial stem cell differentiation toward hair follicle lineages. Our results show that the type of fibroblasts chosen to elaborate the dermal portion of tissue-engineered skin have an impact on the differentiation fate of keratinocytes and on the hair-forming capacity of epithelial stem cells.
Here, we have developed a tissue-engineered skin model that allows to maintain the hair inductive formation property of HBs in vitro as far as 26 days. About 1 month after transplantation, hair emerged on the surface of grafts when the tissue-engineered dermis was elaborated from neonate mouse fibroblasts. This physiological environment comprising fibroblasts and the matrix they organized over culture in an ascorbic acid-containing medium allowed growth and differentiation, as well as long-term survival of complete pilo-sebaceous units from HBs after grafting. In contrast, the same HBs did not develop into mature hair follicles in the presence of the human fibroblast-derived tissue-engineered dermis. In the present study, HBs were obtained from the dermal portion of newborn mouse. At this stage, immature hair follicles are composed of a dermal papilla surrounded by epithelial cells. 21 Here, even if HBs implanted alone developed hairs (Fig. 5A and see also Lichti 20 and Weinberg20,21), thus containing the cell types as well as the cell contacts required for hair follicle formation, the human fibroblasts and their derived matrix blocked HB growth and differentiation toward the hair follicle lineage in the HB/hD grafts. Histological analysis of HB/hD grafts showed that the HB-derived epithelium rather presented epidermal histological features. The appearance of these cysts was very similar to the structure developed from hair follicles following either β-catenin deletion 57 or expression of Lef1 transgene, which lacks the β-catenin–binding site. 58 The absence of the nuclear complex β-catenin/Lef1 in the cysts observed in the HB/hD grafts suggested an interruption of the Wnt signaling pathway and could explain why stem cells are unable to adopt the fate of hair keratinocytes in the presence of human fibroblasts, in contrast to mouse fibroblasts, which support hair formation after grafting.
The in vitro maturation obtained by culturing tissue-engineered skin at the air–liquid interface further emphasizes the distinct influence of human and mouse fibroblast sheets on HB epithelial cell differentiation. Indeed, different types of HB-derived epithelial inclusions were observed within the tissue-engineered dermis of HB/mD and HB/hD. Inclusions associated with the HB/mD skin featured ORS-like characteristics such as 2–3 layers of cells with cuboidal morphology and keratinocytes expressing K19, a keratin found in a subset of cells presenting stem cell characteristics, localized in the basal cells of the ORS in the bulge region of the hair follicles in situ.41,48 On the other hand, epithelial inclusions associated with HB/hD contained more epithelial layers. Their histological aspect as well as the expression of proteins related to the specialization program of the epidermis, such as K10 and loricrin,45,51 suggested that the human fibroblast-derived tissue-engineered dermis induced epithelial cells within HBs to adopt an epidermal fate. This influence may have abrogated the potential of HBs to form hair follicles after grafting. Further, epidermis regenerated on hD were significantly thicker than those observed on mD, indicating that the type of fibroblasts used to generate tissue-engineered dermis affects the differentiation/proliferation balance of keratinocytes. This is consistent with previous findings on the variation of the epithelial thickness in vivo when human fibroblasts from dermis or corneal stroma were used in the production of tissue-engineered skin, tissue-engineered cornea, or chimeric constructs. 59
In conclusion, tissue-engineered skin with hair can be produced using the self-assembly approach of tissue engineering. Moreover, our works have highlighted that fibroblasts within tissue-engineered dermis participate in the regulation of keratinocyte differentiation program. These models will be very useful in identifying factors that preserve stem cells and regulate epidermis versus hair follicle differentiation.
Footnotes
Acknowledgments
The authors are grateful to Israël Martel, Claudia Fugère, Anne-Marie Moisan, Robert Gauvin, Amélie Lavoie, and Gabrielle Ouellet for their technical support. This work was supported by the Canadian Institutes of Health Research (CIHR). D.L. was the recipient of a studentship from the CIHR. C.P. was the recipient of a postdoctoral scholarship from the Fonds de la Recherche en Santé du Québec (FRSQ). L.G. is the holder of a Canadian Research Chair on stem cells and tissue engineering (CIHR) and was the recipient of fellowships from the CIHR and the FRSQ.
Disclosure Statement
No competing financial interests exist.