
Abstract
The initial seeding density is a critical variable in functional tissue engineering. A sufficient number of cells uniformly distributed throughout the scaffold is a key requirement to achieve homogeneous extracellular matrix deposition in vitro. However, high initial seeding densities might have negative repercussions on nutrient availability, cellular metabolism, and cell viability. In the current study, our aim was to understand the implications of using high seeding densities (3, 5, and 10 million cells/mL) in a human umbilical vein (HUV) tendon model subjected to 1 h of cyclic stretching per day at 2% strain and a frequency of 0.0167 Hz in a mechanostimulating bioreactor, on nutrient availability, cell viability and metabolism, and construct properties. Mechanostimulated constructs seeded with 3 million cells/mL had significantly higher cell number than the static controls and resulted in a 20-fold increase in proliferation rates and a 3-fold increase in tensile strength values after 1 week of culture in the bioreactor. However, higher seeding densities resulted in cell death, degraded extracellular matrix, and poorer mechanical properties. Nutrient and growth factor mass transport limitations are implicated in the inability of the decellularized HUV to support high cell numbers. The effective diffusion coefficient for glucose was measured to be 0.21±0.04 cm2/day. In the absence of convective flow, proteins and growth factors with a molecular radius larger than 4.9 nm could not diffuse through the HUV. Cells seeded in the HUV consumed 10.5±0.5 ng/cell/day of glucose. Glucose diffusion coefficient and glucose consumption rates in the HUV indicated the presence of glucose mass transport limitations when cell seeding densities exceed 3 million cells/mL.
Introduction
The main three components of functional tissue engineering are cells, a biocompatible scaffold to house the cells and give the construct initial mechanical integrity, and a bioreactor that provides the appropriate biochemical and biomechanical signals to regulate cell differentiation and tissue assembly.2–5 The initial seeding density is a critical variable in attaining functional engineered tissue since cells are responsible for tissue deposition and extracellular matrix (ECM) remodeling. A sufficient number of cells uniformly distributed throughout the scaffold is a key requirement to achieve homogeneous ECM deposition in vitro.6–8 Increasing cell seeding densities has been shown to result in more aligned and elongated cell nuclei, higher final cell number per construct, and elevated ECM deposition rates compared with lower seeding densities.9,10 However, high initial seeding densities might have negative repercussions on nutrient availability, cellular metabolism, and cell viability.
In a previous study our group introduced a novel model for tendon tissue engineering composed of culturing the human umbilical vein (HUV) seeded with bone marrow mesenchymal stem cells (MSCs) in a mechanical stimulator for tissue engineering applications (MSTE). 11 In the current study, our aim was to understand the implications of using a range of seeding densities in the HUV tendon model on nutrient availability, cell viability and proliferation, and construct properties. We hypothesize that the initial seeding density will significantly influence cell proliferation rates and distribution within the scaffold and construct tensile properties. We thus investigated the effect of three alternative seeding densities: low seeding density (low SD=3 million cells/mL), medium seeding density (medium SD=5 million cells/mL), and high seeding density (high SD=10 million cells/mL) on the metabolic, morphometric, and mechanical properties of the tissue-engineered tendon. Since cells are seeded within the central portion of the decellularized HUV and the supply of nutrients is from the external surface, transport of key nutrients, including growth factors, is critical to maintain appropriate cell function, and for the constructs final biological and biomechanical properties. To assess diffusion limitations through the decellularized scaffold the effective diffusivity and permeability of glucose and fluorescein isothiocyanate (FITC)-dextran molecules were computed.
Materials and Methods
All materials were purchased from (Invitrogen, Carlsbad, CA) unless specified otherwise.
Preparation of the HUV scaffold
The HUV was dissected out of the human umbilical cord using previously published methods.11–13 Since cells integrated with the rough lathed Wharton's jelly matrix surface better than the smooth luminal surface, 12 HUV sections were inverted inside out such that the Wharton's Jelly matrix is on the central portion of the HUV (Fig. 1). The dissected scaffold was then decellularized using 1% sodium dodecyl sulfate (Baker, Phillipsburg, NJ), sterilized using 0.2% peracetic acid (Sigma Aldrich, St. Louis, MO), and stored in phosphate-buffered saline (PBS; Atlanta Biologicals, Lawrenceville, GA) at a temperature of 4°C.

Schematic cross section showing the main two layers composing the human umbilical vein (HUV): the HUV wall (HUVwa) and the Wharton's Jelly (WJ).
Cell source
Bone marrow MSCs were harvested from the femur and tibia of 6–8-week-old male Wistar rats (Harlan, Indianapolis, IN) using well-documented procedures.14–16 Cells were plated on 75 cm2 culture plates and incubated in a humidifier at 37°C and 5% CO2. All cells used for experimental purposes were from passage 2.
Experimental design
HUVs were seeded and cultured in the bioreactors as described previously. 11 Briefly, MSCs were mixed with type I collagen (Angiotech Biomaterials, Vancouver, BC, Canada) to yield a final cell density of 3, 5, or 10 million cells/mL and a collagen concentration of 2 mg/mL. Custom-designed steel adapters were attached to each end of the HUV facilitating the handling of the scaffold during seeding. The HUV scaffolds were held by the adapters using hemostats. Then, a total volume of 0.6 mL of cellular collagen solution was inoculated in the central portion of each 6.5 cm long section of HUV using a 1 mL pipette, after which constructs were incubated for 1 h at 37°C to permit collagen gel polymerization. Seeded scaffolds were then cultured under sterile conditions in a custom-made MSTE bioreactor that permits controlled application of different mechanical stimulation regimes to triplicate constructs simultaneously. 11 Two cell-seeded groups were assessed: static controls without any mechanical stimulation, and stimulated samples, which were subjected to 1 h of cyclic stretching per day at 2% strain and a frequency of 0.0167 Hz. Constructs were assessed at 1 and 2 weeks to determine cellular density, morphometric appearance, and mechanical properties. Glucose consumption rates were measured and normalized by the number of cells in each construct. To assess diffusion limitations through the scaffold, the permeability of the decellularized HUV to glucose and dextran molecules of a wide range of molecular weights was measured.
Calculation of glucose consumption
Constructs seeded with 3 million cells/mL (low SD) were cultured in the MSTE with fresh media containing 1 mg/mL glucose (alpha minimum essential medium supplemented with 10% fetal bovine serum [Atlanta Biologicals], 100 units/mL penicillin G 100 μg/mL streptomycin sulfate, and 0.25 μg/mL amphotericin B). One milliliter of the fresh medium was stored in a centrifuge tube and placed in the incubator as a control sample. After 24 h, the medium was collected from the MSTE and tested for glucose content using an enzymatic method. About 100 μL of glucose assay reagent (Sigma-Aldrich) was incubated with 10 μL of sample for 20 min. The absorbance was then measured at 340 nm and the glucose content was deduced from a standard curve. The glucose consumption was calculated as the difference between the glucose content of the control and that of the sample. The constructs were also tested for DNA content to compute the glucose consumption rate per cell.
Quantifying cell number
Three ringlets having a length of 0.5 cm were cut from different locations of the HUV (two from each end and one from the middle section) to obtain an average representative number. Sections were then dissected into smaller pieces ≤2 mm2, and incubated with 1 mL of collagenase type I (MP Biomedicals, Solon, OH) for 5 h at 37°C to release cells from ECM. The final cellular mix was sonicated in ice for 1 min and then subjected to three freeze–thaw cycles. To quantify the cell number a pico green DNA quantification kit was used according to the manufacturer's protocol. Fluorescence was read at an excitation/emission wavelength of 480/520 nm. DNA concentrations were converted to cell densities based on an average DNA content per MSC of 3 pg/cell.
Histology
Supplies for conducting histology were purchased from Richard-Allan Scientific (Kalamazoo, MI) unless specified otherwise. Five-millimeter-thick ringlet tissue samples were fixed in 10% neutral-buffered formalin and embedded in paraffin blocks. Seven-micrometer-thick sections were sliced and mounted on Histobond slides (VWR). Tissue samples were then stained using hematoxylin and eosin.
Fiber alignment
Fiber alignment was analyzed using ImageJ. The reference axis was taken as the direction parallel to stretching, from which the angular deviation of fibers was measured. The mean with the standard deviation were reported.
Shape of cells
Using the ImageJ program, it was possible to quantify the dimensions of the cells (length, width, and area) that appear in a longitudinal slide. The shape factor (width/length) was then computed. The closer the shape factor is to 1, the more round the shape of the cell is.
TUNEL assay
The DeadEnd™ Colorimetric TUNEL System kit from Promega (Madison, WI) was used to determined cell apoptosis in the tissue samples. Paraffin-embedded cross sections of the HUV were assayed for apoptotic cells following the manufacturer's protocol. Briefly, tissue slides were deparaffinized in Clear-Rite (Richard-Allan Scientific). The sections were then rehydrated in series in decreasing ethanol concentrations and washed with a sodium chloride solution and PBS. Tissue samples were then fixed in 10% PBS-buffered formalin (VWR) and permeabilized with a proteinase K solution. After fixing, the cells' DNA was attached with biotinylated nucleotides using a recombinant terminal deoxynucleotidyl transferase solution. Sodium chloride and sodium citrate solution was used to stop the reaction, after which, the nucleotides in solution were tagged with horseradish peroxidase-labeled streptavidin. Finally, the nuclei were stained dark brown by a hydrogen peroxide, diaminobenzidine chromogen, and peroxidase substrate solution. The samples were then analyzed using light microscopy.
Mechanical testing
Using a uniaxial tensile testing frame (United Testing Systems, model SSTM-2K, Flint, MI), samples were preconditioned for five cycles and then stretched to failure at a strain rate of 1%/s. 12 Force and extension data were digitally recorded and used to compute stress and strain values. The gauge length and cross-sectional dimensions were accurately measured for every sample using digital calipers. To account for end effects force and extension data were collected only from samples that failed in the region away from the clamps (a minimum of 10% of the gauge length).
Measurement of effective diffusivity and permeability through the HUV
A side-by-side diffusion chamber was designed to measure the effective diffusivity through the HUV matrix. 17 The setup consisted of two cylindrical chambers having a diameter of 9.5 mm, a length of 3.5 cm, and an individual volume of 2.5 mL (Fig. 2a). The decellularized HUV was cut open along its longitudinal axis, and dissected into 1.5×1.5 cm squares having a thickness of 1.0±0.2 mm. Figure 2b shows a histological longitudinal section of the different layers in the decellularized HUV. Sections of decellularized HUV were placed between the two chambers and the molecule of interest was loaded in the first chamber at a predetermined concentration, facing the luminal side of the HUV, and 2.5 mL of nano pure water was loaded in chamber 2 facing the abluminal side of the HUV. This layout for the HUV membrane was chosen because in the bioreactor medium was supplied to the construct at the luminal side of the HUV, and nutrients had to diffuse through the membrane thickness to the central portion where the cells were inoculated (Fig. 2b).
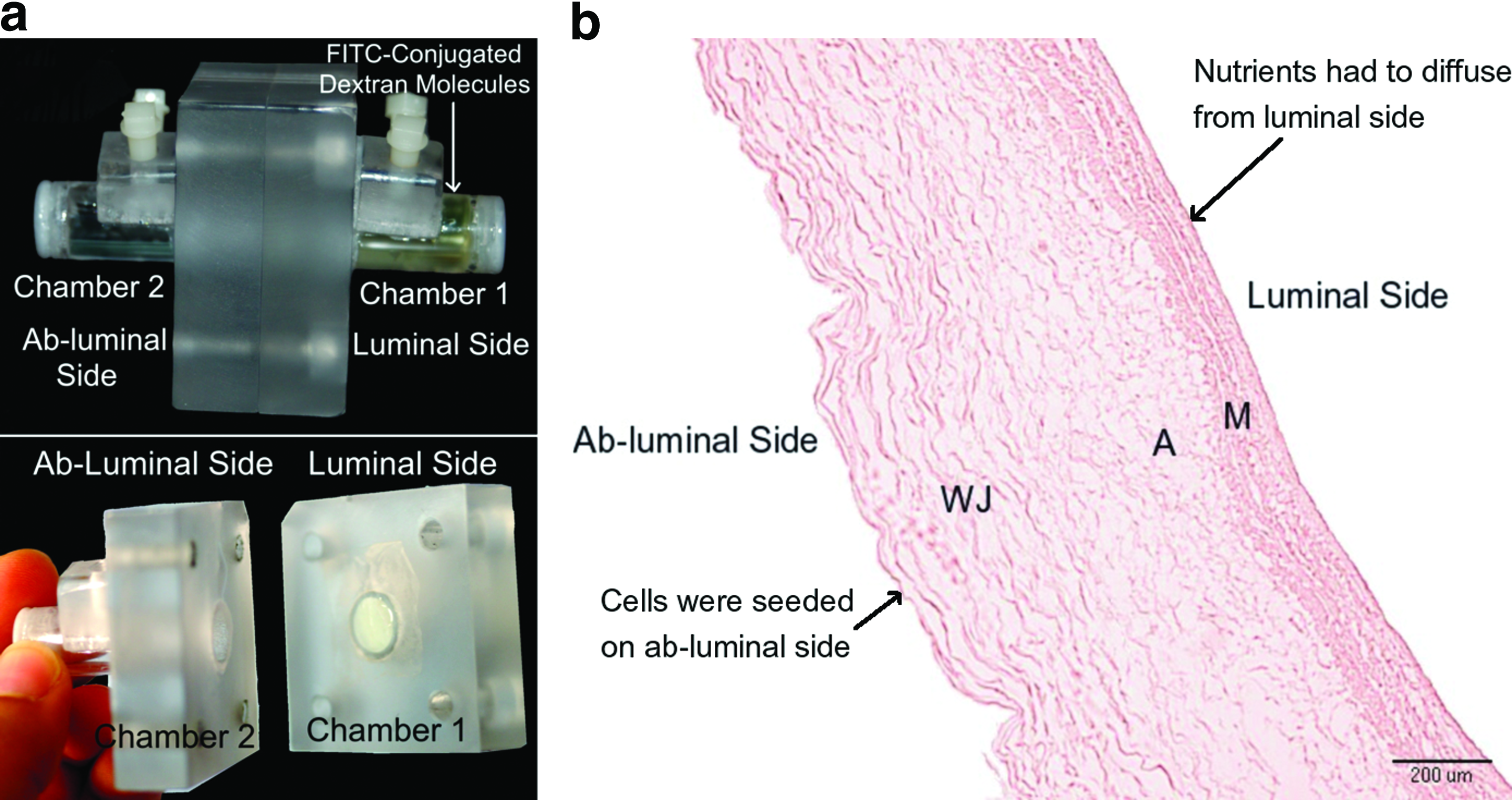
Two different solutions were used for the measurements of the effective diffusion coefficient. The first solution was FITC-conjugated dextran molecules at three different molecular weights: (FITC-4) 4.4 kDa, (FITC-50) 50 kDa, and (FITC-580) 580 kDa (Sigma-Aldrich), and loaded at a concentration of 70 μg/mL. The second solution was glucose (0.18 kDa; Sigma-Aldrich) at a concentration of 100 mg/mL. Ten-microliter samples were collected from both chambers at different time points and were replaced by 10 μL of the original solution at the beginning of the experiment. Collected samples were diluted to 1 mL using nano pure water. The concentration of different molecules at each time point was calculated by measuring the fluorescence (480/520 nm) of FITC-dextran molecules or absorbance (340 nm) of the sample incubated with glucose assay reagent. The coefficient of diffusion (Deff) was calculated assuming a quasi-steady state approximation 18 and considering the HUV as a thin membrane. The permeability (P) of different molecules to the membrane was deduced by dividing Deff by the thickness of the HUV.
Statistical analysis
For each culturing duration and seeding density at least three constructs were dedicated to quantifying cell number and histological evaluations (n≥3), and from each construct three representative sections dissected from different locations along the seeded HUV were considered (n=3). At least six constructs (n≥6) were employed for mechanical testing. All results were expressed as mean values±standard deviation. To determine the statistical significance of different variables the two-way analysis of variance method using the software PRISM was employed, with the Bonferroni post hoc test to make specific comparisons among groups or against a control.
Results
Cell proliferation
Figure 3 shows plots of cellular proliferation for constructs seeded at the low SD of 3 million cells/mL, medium SD of 5 million cells/mL, and high SD of 10 million cells/mL. A total volume of 0.6 mL of cellular collagen mix was originally inserted in the HUV. Thus, with a seeding density of 3 million cells/mL, the total number of cells originally inserted in the HUV was 1.8 million cells. Similarly, 3 million cells and 6 million cells were inserted in the HUV for the medium SD and high SD, respectively.
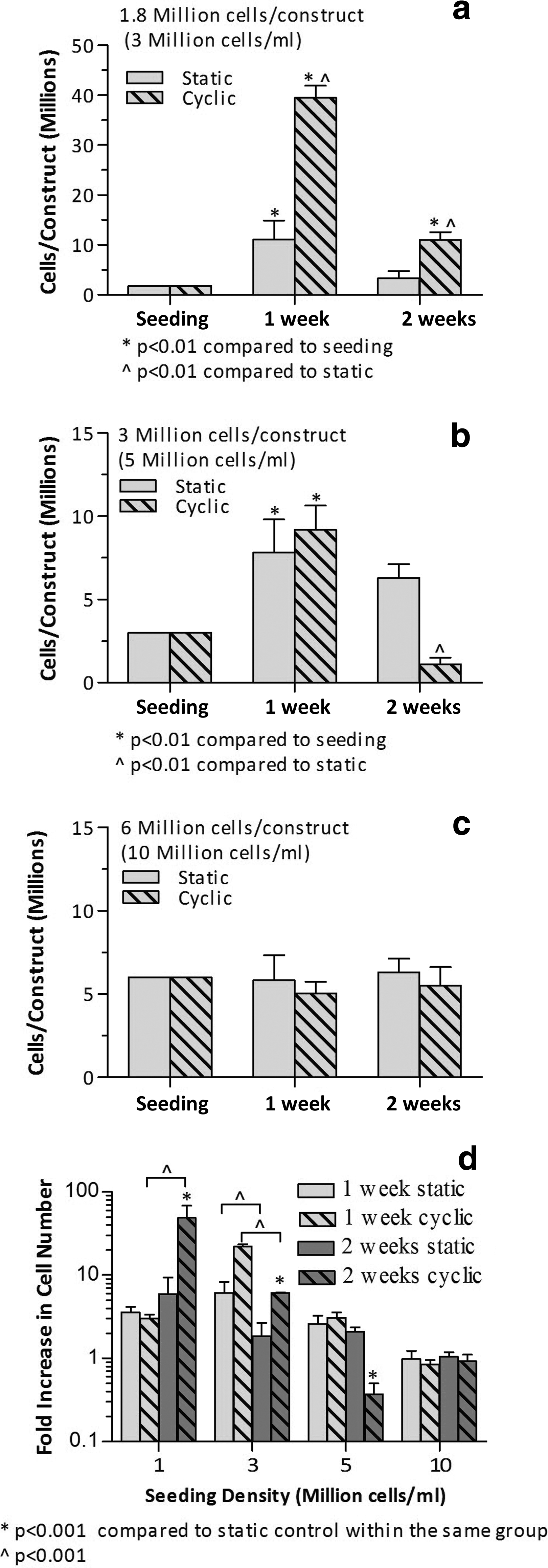
Plots of cell number (in million) per construct versus culturing duration for an initial seeding density of
A significant increase in cell number was measured for the low SD and medium SD (Fig. 3a, 3b) after 1 week of culture for both static and stimulated constructs. Although the pico green assay does not directly measure the number of viable cells, an increase in DNA concentration over time indicates that cells are viable and proliferating. For the low SD the stimulated samples had significantly higher cell number than the static controls. After 1 week of culture, the cellularity of the control samples increased almost 6-fold for the low SD and 2.6-fold for medium SD. This increase was significantly higher for cyclically stretched samples: 22-fold for the low SD and 3-fold for medium SD. No additional increase in cell number was measured for the constructs 2 weeks postculture. Interestingly, there had been a decrease in DNA content after 2 weeks of culture that was more significant for stretched samples. Contrarily to low SD and medium SD results, no significant change in cell number was measured for the high SD for either culturing regimes at the time points tested (Fig. 3c).
TUNEL assay
All seeding densities and conditions showed apoptotic cells after 2 weeks of culture reaching levels between 30% and 50% of the stained cells for the high SD. Most of the apoptotic cells in the low SD were toward the second half of the scaffold closer to the wall of the HUV. Contrarily, for the high SD, apoptotic cells were concentrated in the central portion of the HUV.
Histology
Figure 4 shows the morphology of the stimulated constructs after 1 week of culture. The decellularized HUV has a tortuous spongy matrix (Fig. 4a). MSCs seeded at 3 million cells/mL were able to proliferate and migrate into the matrix of the stimulated HUV (Fig. 4b) after 1 week of culture. It is clear that the scaffold was occupied with numerous cells, some of which had reached the outer wall of the HUV, possibly remodeling the original tortuous and porous matrix. Quantitative histomorphometry revealed 65% of the cells occupied the first half of the HUV thickness, whereas almost 35% migrated deep into the second half of the decellularized scaffold. Longitudinal histological sections showed parallel fiber orientation with an average fiber deviation from the axial direction measured to be 10.58°±°2.83°. Similar to the previous study, 11 after 1 week of culture cell nuclei seemed to be more spindle-like with a shape factor of 0.75±0.03.
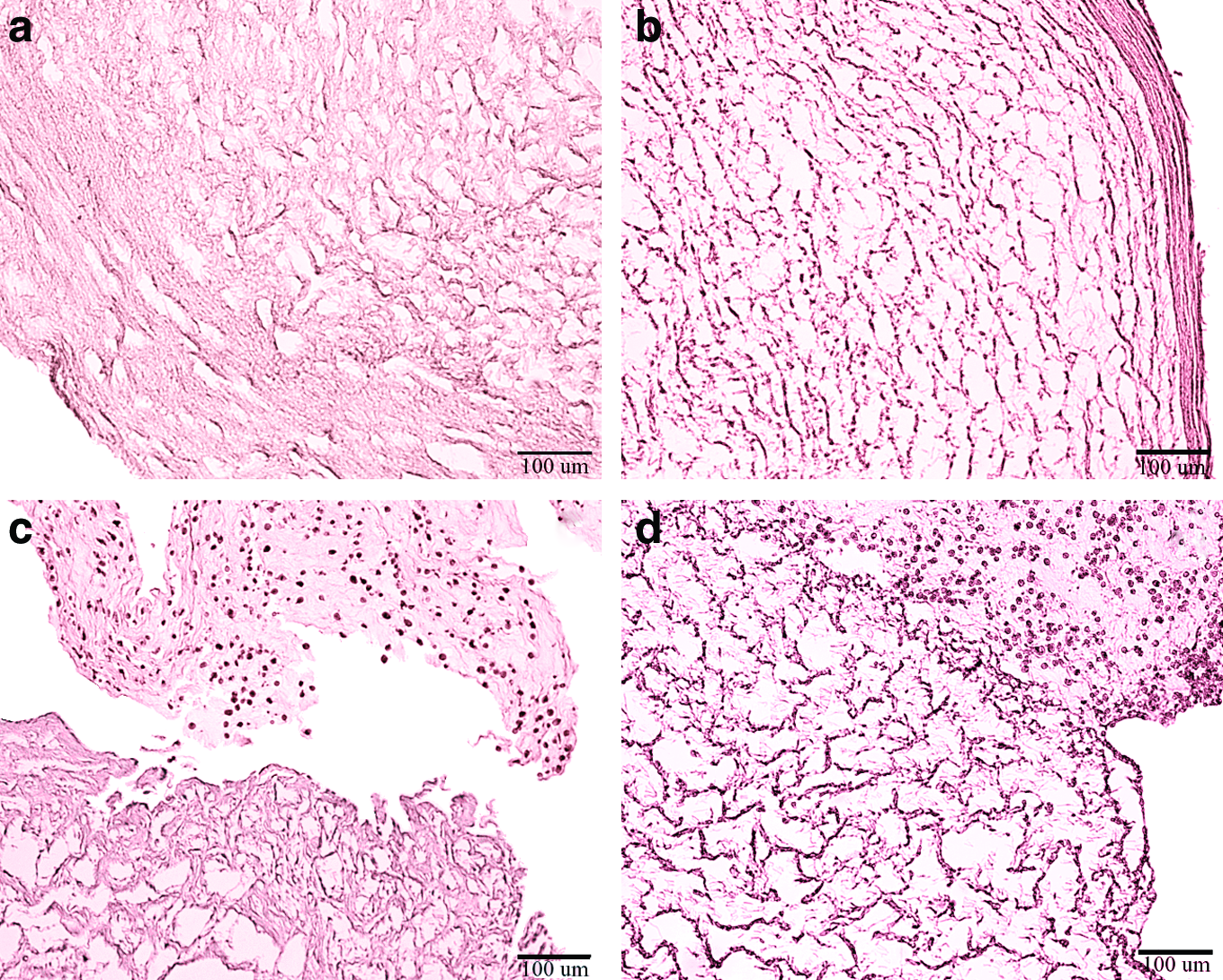
Histologic cross sections at a magnification of 10×of
For the medium SD cells were mostly present in the central portion of the HUV (Fig. 4c). A few cells migrated into the ECM of the scaffold that appeared similar in structure as that of the acellular HUV. Cell nuclei looked more round with a shape factor of 0.89±0.09. Although cells occupy the inside of the HUV seeded at high SD (Fig. 4d), cells appeared to have an irregularly round shape with faded staining that could be an indication of slowed metabolism of apoptotic cells.
A qualitative observation of ECM after 2 weeks of culture is that ECM appears degraded, with fewer fibers surrounding cells compared with the ECM of 1-week cultures (Fig. 5). Fibers were clearly short and disconnected for all culturing conditions and seeding densities after 2 weeks, unlike the connected mesh-like morphology of the original decellularized HUV (Fig. 4a). Moreover, cell fragments that appear in Figure 5 indicated to be lysed cell bodies at a magnification of 40×(figure not shown). These observations are in agreement with the cell proliferation results that indicated a significant decrease in cell number after 2 weeks of culture.

Histologic cross sections of the HUV after 2 weeks of culture at a magnification of 20×for
Mechanical properties
For the low SD an improvement in the tensile strength was observed after the first week of culture (Fig. 6a). A statistically significant increase in the constructs' tensile strength was recorded for stimulated samples (2.7±0.8 MPa) compared with 1.5±0.5 MPa for static, with control samples (acellular) at 0.9±0.1 MPa. Similarly, for the medium SD (Fig. 6b), the tensile strength for stimulated HUVs was 4.1±0.5 MPa compared with 1.0±0.3 MPa for the static constructs. No additional increase in tensile strength was measured after 2 weeks of culture. Contrary to the low SD and medium SD, no significant increase in the tensile strength was observed for any culturing condition at either culturing points for the high SD constructs (Fig. 6c).

Tensile strength of the seeded HUVs for different culture periods for low SD
Glucose consumption, effective coefficient of diffusion, and permeability through the HUV
The average cellular glucose consumption rate of MSC cultured in the HUV was 10.47±0.55 ng/cell/day. The resulting average effective diffusion coefficients for glucose and 4.4 kDa dextran (FITC-4) were 0.21±0.04 cm2/day and 0.05±0.01 cm2/day, respectively. However, large molecules (50 and 580 kDa) did not diffuse through the decellularized HUV during the time frame of 48 h. The permeability coefficients of the HUV to glucose (0.18 kDa) and FITC-4 (4.4 kDa) were 2.5×10−5±5.0×10−6 cm/s and 6.4×10−6 ± 9.0×10−7 cm/s, respectively. Figure 7 shows an inversely proportional relationship between the permeability coefficient and the radius of gyration.

An inversely proportional relationship between the permeability coefficient and the molecular radius molecules diffusing through the decellularized HUV.
Discussion
The seeding density is an important factor that contributes to the success of a functional tissue-engineered construct. In this study we explore the effects of different seeding densities on the morphometric, mechanical, and metabolic properties of a tissue-engineered tendon. Decellularized HUV bioscaffolds were seeded with rat bone marrow MSCs using three different seeding densities (3, 5, or 10 million cells/mL), and cyclically stretched for 1 h/day for durations of 1 and 2 weeks. The constructs were tested for cellularity, histological appearance, and tensile strength. Since high initial seeding densities might have negative repercussions on nutrient availability and cellular metabolism the permeability of the HUV to glucose and FITC-dextran molecules has been computed, and cellular glucose consumption rates were measured.
For the low SD and medium SD constructs, a statistically significant increase in cell number was measured after 1 week of culture for both tensioned and untensioned constructs (Fig. 3). However, no additional increase in cell number was recorded 2 weeks postseeding. On the contrary, the pico green assay reflected a decrease in total DNA content, indicating cell death. Moreover, high SD constructs did not show any increase in cell number over the entire time course. These results were in agreement with histologic slides. Cross sections for the low SD 1 week postculture revealed numerous cells invading the ECM of the scaffold reaching the outer wall of the HUV for stimulated constructs (Fig. 4b). Nuclei of the MSC had a spindle-like appearance similar to that of tenocytes with a shape factor 0.75±0.03. However, for the medium SD and high SD cells appeared more round with a shape factor 0.89±0.09, and were concentrated in the central portion of the HUV (Fig. 4c, 4d), and the ECM of the scaffold appeared similar to the decellularizd HUV (controls), indicating a lack of remodeling (Fig. 4a). By 2 weeks of culture the ECM fibers of the HUV appeared to be degraded, and lysed cell bodies were present in significant numbers for both culturing conditions and for all seeding densities (Fig. 5). The decreased DNA content after 2 weeks could be explained by these histologic slides where DNA was released from the lysed cell bodies and either diffused out of the constructs or was degraded by DNAses. Mechanical testing revealed an increase in the ultimate tensile strength of stretched constructs only 1 week postculture (Fig. 6). After 2 weeks of culture, cell death, evident degradation of the ECM, and no further increase in the constructs' mechanical properties were noted.
Figure 3d shows the fold increase in cell number within the HUV for four different seeding densities: 1 million cells/mL (taken from a previous study 11 ), low SD of 3 million cells/mL, medium SD of 5 million cells/mL, and high SD of 10 million cells/mL. Proliferation rates had a parabolic relationship with the seeding density after 1 week of culture. At a seeding density of 1 million cells/mL, the cellularity within the constructs increased more than threefold 1 week postculture (Fig. 3d). Proliferation rates increased further to more than 20-fold for the low SD. However, for the medium SD, cellularity of the HUV constructs only increased twofold, and for the high SD there was no increase in cell number. Proliferation seized after 2 weeks of culture for the low SD, medium SD, and high SD. Contrarily, cells in stimulated constructs seeded at 1 million cells/mL showed an additional 20-fold increase in proliferation rates. 11 These results are in agreement with other studies in which lower seeding densities using MSCs resulted in higher proliferation rates. 19 It is possible that not enough nutrients were available for the cells to proliferate at higher seeding densities, which in turn induces metalloproteinases (MMPs) production. 20 Moreover, cells could have been attempting to remodel the ECM by secreting MMPs. However, the rate of ECM deposition was not balanced by the rate of MMP secretion. When cells remodel the ECM, they also deposit new matrix that could potentially decrease the effective porosity of the scaffold, and in turn decrease the flow of nutrients through the membrane. These facts would limit mass transport through the HUV causing the cells to experience stressful conditions and eventually lyse.
Another possible explanation for the lack of proliferation is due to the increased metabolic activity of proliferating cells that leads to an increase in metabolic byproducts such as production of lactic acid. The accumulation of lactic acid in the HUV creates an acidic environment that would become toxic to the remaining viable cells accompanied by increased secretion of MMPs 20 eventually causing ECM degradation. The TUNEL assay showed apoptotic cells for all culturing conditions. The majority of apoptotic cells for the low SD were present toward the outer wall of the HUV. Contrarily, for the high SD, between 30% and 50% of stained cells were apoptotic and cells were concentrated in the central portion of the HUV. Hypoxia 21 and acidosis 22 are two possible explanations for induced apoptosis, especially at high SDs.
To assess mass transport limitations, the permeability of the decellularized HUV to glucose and dextran molecules with molecular weights ranging between 4 and 580 kDa was calculated. Although high-molecular-weight dextrans (>50 kDa) did not diffuse through the HUV after 2 days, it could not be concluded that proteins having a molecular weight larger than 50 kDa would not permeate through the membrane. Two molecules having the same molecular weight might have significantly different geometries. For instance, bovine serum albumin has a molecular weight of 65 kDa but a molecular radius R=3.5 nm 23 significantly lower than FITC-50 with an R=4.9 nm. Thus, it was necessary to compare the permeability of the HUV using the molecular radius, or radius of gyration, rather than the molecular weight (Fig. 7). The coefficient of permeability had an inversely proportional relationship with the molecular radius (Fig. 7). Thus, using the resulting relation it is possible to predict the coefficient of permeability of dextrans with different molecular weights through the HUV. Moreover, the proposed relation could provide an approximate diffusivity coefficient of proteins with different chemical properties than dextran. Therefore, large proteins, such as immunoglobulin G antibodies (Mw≈150 kDa), having a molecular radius of 5.0 nm 23 larger than 4.9 nm would probably not diffuse through the wall of the HUV.
To compute a limiting initial seeding density in the HUV above which no glucose would be available for cellular nutrition, Fick's first law of diffusion could be applied
Conclusions
The HUV tendon model seeded at 3 million cells/mL and cyclically stretched for 1 h/day resulted in a 20-fold increase in proliferation rates and a 3-fold increase in tensile strength values after 1 week of culture. However, initial seeding densities higher than 3 million cells/mL resulted in a cyctotoxic environment causing cell death, ECM degradation, and loss of construct mechanical integrity. The seeding density was limited by cellular glucose consumption and glucose diffusivity through the HUV. In the absence of convective flow, the initial seeding density was restricted to 3 million cells/mL. Stimulating the HUV tendon model for longer than 1 h/day may circumvent these restrictions by enhancing mass transport through the scaffold. By understanding the dynamics of cell density and nutrient demand, this engineered tendon could be implanted before complete in vitro remodeling to allow the body to naturally vascularize the tissue within a time frame that maintains seeded cell viability and function during a secondary in vivo remodeling step where a more natural migration and cell density can occur.
Footnotes
Acknowledgments
The authors would like to acknowledge the Women's Center in Norman Regional Hospital, Norman, OK, for supplying the umbilical cords used in the current study. The authors thank Dr. Paul DeAngelis from the University of Oklahoma Health Sciences Center for supplying the FITC-dextran molecules. Finally, the authors would like to acknowledge the Oklahoma Center for the Advancement of Science and Technology (HR09-085) for financial support.
Disclosure Statement
The authors have nothing to disclose.