
Abstract
Although it has been observed that aggregate size affects cardiac development, an incomplete understanding of the cellular mechanisms underlying human pluripotent stem cell-derived cardiomyogenesis has limited the development of robust defined-condition cardiac cell generation protocols. Our objective was thus to elucidate cellular and molecular mechanisms underlying the endogenous control of human embryonic stem cell (hESC) cardiac tissue development, and to test the hypothesis that hESC aggregate size influences extraembryonic endoderm (ExE) commitment and cardiac inductive properties. hESC aggregates were generated with 100, 1000, or 4000 cells per aggregate using microwells. The frequency of endoderm marker (FoxA2 and GATA6)-expressing cells decreased with increasing aggregate size during early differentiation. Cardiogenesis was maximized in aggregates initiated from 1000 cells, with frequencies of 0.49±0.06 cells exhibiting a cardiac progenitor phenotype (KDRlow/C-KITneg) on day 5 and 0.24±0.06 expressing cardiac Troponin T on day 16. A direct relationship between ExE and cardiac differentiation efficiency was established by forming aggregates with varying ratios of SOX7 (a transcription factor required for ExE development) overexpressing or knockdown hESCs to unmanipulated hESCs. We demonstrate, in a defined, serum-free cardiac induction system, that robust and efficient cardiac differentiation is a function of endogenous ExE cell concentration, a parameter that can be directly modulated by controlling hESC aggregate size.
Introduction
One of the main challenges of aggregate-based hPSC differentiation is that heterogeneity and spatial disorganization leads to inefficient differentiation to specific cell types and a poor understanding of the mechanisms involved in lineage commitment. In studies addressing this challenge, two strategies have emerged. One involves exogenously controlling differentiation by delivering factors that are known or thought to be involved in specification, commitment, and proliferation of the cell type of interest.21–24 The second strategy focuses on controlling physical parameters of aggregate formation, such as aggregate size and shape.24–31 We and others have previously reported the observation that human embryonic stem cell (hESC) aggregate size can be modulated to optimize cardiac induction efficiency25,26,30,32,33; however, the cellular mechanism by which aggregate size and geometry impacts differentiation to specific cell types remains unclear. Given that the surface area-to-volume ratio of a sphere decreases with increasing sphere size, and that an ExE layer forms on the surface of the differentiating aggregates, we examined whether this geometric relationship dictated ExE cellularity during aggregate-based hESC differentiation, and measured the implications of varying ExE concentrations on cardiomyocyte differentiation efficiencies.
We have previously observed that the differentiation propensity of the input population is a function of hESC colony size, a parameter that significantly affects cardiac induction efficiency. 25 Herein, to eliminate heterogeneity of the input hESCs, we tracked ExE and cardiac development in size-specified aggregates formed using forced microwell-based aggregation of a single-cell suspensions. 31 To investigate the effect of ExE differentiation frequency on cardiomyocyte differentiation independently of aggregate size, we manipulated the expression of SOX7, a transcription factor that plays a crucial role in ExE development.34,35 SOX7 silencing leads to inhibition of ExE differentiation of pluripotent cells, 34 and SOX7 overexpression of hESCs leads to the development of an ExE progenitor phenotype. 35 The results presented here demonstrate, for the first time under defined media conditions, 21 that varying hESC aggregate size impacts the efficiency of cardiac development. We show that this aggregate size effect occurs by a mechanism wherein the aggregate surface area-to-volume ratio (specified by its size) determines the amount of cardiac-promoting ExE developed during early differentiation, which in turn specifies subsequent cardiomyogenesis.
Materials and Methods
Cell culture
The HES2 (ES Cell International), and CA1 (Nagy Lab, Mount Sinai Hospital, University of Toronto) hESC cell lines used in these experiments were maintained and expanded as previously described.21,25,35 SOX7 O/E cell lines were generated from CA1 cells as previously described 35 and maintained in the same conditions as their parental CA1 cell line. Aggregate-based differentiation of hESCs was carried out using a protocol for serum-free directed differentiation to the cardiac lineage that has been previously described. 21 hESC aggregate size was controlled by forced aggregation of defined cell concentrations in AggreWell™ inserts (STEMCELL Technologies) containing a textured surface of microwells. 31 Human cardiac myocytes were purchased and cultured in a commercial medium for four passages before extracting RNA for the positive control in quantitative real-time-polymerase chain reaction analysis (ScienCell).
Knockdown of SOX7 gene expression in hESCs by siRNA
The Dharmafect™ 1 (Dharmacon) delivery system was used to transfect hESCs with predesigned siRNA against SOX7 (100 nM, SMARTpool, L-019017-01) according to the manufacturer's protocol in OptiMEM (Invitrogen) base medium. Control cells were similarly transfected with nontargeting siRNA (ON-TARGETplus Non-targeting Pool, D-001810-10-20). Aggregates were formed by forced aggregation of varying ratios of SOX7 siRNA-transfected HES2 cells and SOX7 O/E CA1 cells (hESC-derived ExE progenitors).
Flow cytometry
Dissociated hESC colonies or differentiating aggregates were enzymatically dissociated to single cells using the appropriate dissociation technique. For intracellular staining, cells were fixed and permeabilized with the IntraPrep fixation and permeabilization kit (Immunotech). Primary antibodies for detecting intracellular proteins were directed against FoxA2 (R&D Systems), GATA6 (R&D Systems), and cardiac Troponin T (cTnT; Lab Vision). Antibodies for detecting surface protein expression were directed against KDR and C-KIT. Cells were analyzed using a FACSCanto (BD Biosciences) flow cytometer.
Confocal microscopy
hESC aggregates were fixed and stained for confocal microscopy imaging using a protocol that has been previously described. 36 FoxA2 (1:1000; R&D Systems) and secondary (1:200; Molecular Probes) antibodies were used. Cell nuclei were detected by staining aggregates with Hoechst 33342 (Molecular Probes).
Immunostaining and imaging
Fluorescent images of Sox7 (1:100; R&D Systems), Sox17 (1:100; R&D Systems), and GATA-6 (1:20; R&D Systems) protein expression were obtained with the Cellomics Arrayscan VTI platform and quantitatively analyzed using the Target Activation assay algorithms available with the Cellomics Arrayscan VTI platform (Cellomics).
Statistics
All data shown are the mean of three independent experiments with error bars on plots representing the standard deviation of the mean unless otherwise indicated. To evaluate differences between conditions, statistics were computed in Origin 7.5 using the two sample t-test or one-way analysis of variance as indicated at a significance level of p<0.05.
Results
Establishing the optimal hESC aggregate size to maximize cardiomyocyte differentiation
Cardiac induction and differentiation were assessed in hESC aggregates formed with 100, 1000, and 4000 cells per aggregate (Fig. 1A). It was previously reported 21 that when conventional scraped embryoid bodies (EBs), cultured in the same defined conditions used for the studies presented here, were analyzed for expression of KDR and CKIT on day (D)5/6, three distinct populations emerged: KDRhigh/CKITpos, KDRlow/CKITneg, and KDRneg/CKITpos. In that report, it was demonstrated that the KDRlow/CKITneg population expressed high levels of genes involved in cardiac development and that these cells subsequently gave rise to an enriched population of cells expressing mature cardiac differentiation markers. 21 We therefore assayed the same markers to assess the influence of aggregate size on cardiac induction. Representative flow cytometry dot plots are presented in Figure 1B. The results of the KDR/CKIT flow cytometry analysis demonstrate that the large (4000 cells) and small (100 cells) aggregate sizes were not optimal for cardiac induction, and that the highest frequency of KDRlow/CKITneg cells were detected in the mid-size condition (1000 cells) (Fig. 1C). This observation is consistent with previously published findings suggesting that varying aggregate size modulates the level of cardiac induction during hESC differentiation.25,26,32,33

Of the three embryoid body (EB) sizes investigated, cardiac induction is maximized in EBs generated from 1000 cells.
By day 12 of differentiation, spontaneous contractions were observed in the majority of aggregates under all size conditions. Cardiac differentiation maturity was assessed by measuring the frequency of cTnT-expressing cells in D16 size-specified aggregates using flow cytometry (representative flow cytometry histograms are presented in Fig. 1D). Corresponding to the results of the cardiac induction analysis, the highest frequency (∼0.25) of cTnT-expressing cells was detected in aggregates initiated with 1000 cells (Fig. 1E). Aggregates formed with 100 and 4000 cells exhibited lower frequencies of cTnT expression by D16, ranging between 0.05 and 0.10 (Fig. 1E). We further characterized beating EBs initiated from 1000 cells using optical mapping and quantitative real-time-polymerase chain reaction. Action potentials could be measured from different parts of spontaneously beating EBs (Supplementary Fig. S1; Supplementary Data are available online at www.liebertonline.com/tea). Also we compared gene expression profiles, using commercially available human cardiac myocytes as a positive control, and documented that hESC-derived cardiomyocytes differentiated into at least three different mature cardiac cell types. We used TNNT2, TNNC1, and ACTN2 as common cardiomyocyte markers; MLC2v as a ventricular-like cardiomyocyte marker; MLC2a as an atrial-like cardiomyocyte marker; and HCN2 as a sinusoidal-like cardiomyocyte marker (Supplementary Fig. S2). These data demonstrate that controlling aggregate size with respect to the number of cells per aggregate influences cardiac induction and differentiation even in the defined medium that contains factors that specify cardiac commitment and expansion.
Emergence and spatial organization of endoderm cells in size-controlled aggregates
We next evaluated spatial organization in differentiating hESC aggregates to confirm that endoderm cells develop on the surface of the spheroid during early differentiation. Figure 2A outlines the relationship between the surface area and volume of a sphere as sphere radius doubles. With each radius doubling, the surface area-to-volume ratio of a sphere is halved. If an inverse relationship also applies to the development of the ExE layer on the surface of a differentiating hESC aggregate, then ExE frequency during early differentiation decreases with increasing aggregate size. Confocal imaging revealed that the spatial expression of the endoderm protein marker FoxA2 was primarily on the surface of D4 aggregates (Fig. 2B). This analysis suggested that spatial organization of endoderm cells to the aggregate surface occurs in hESC aggregates formed by forced aggregation from single cells.
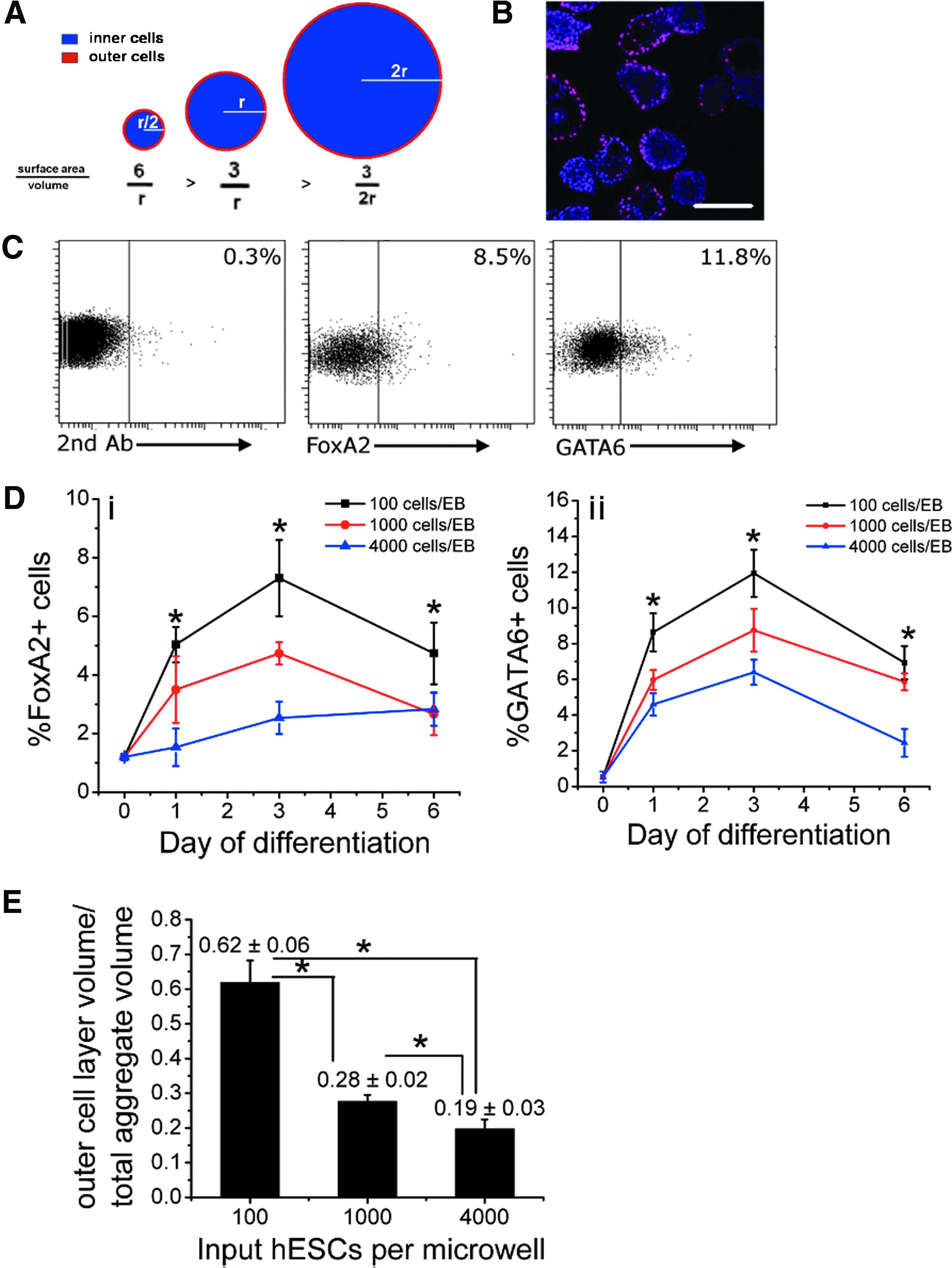
Endoderm cells develop on the surface of size-specified human embryonic stem cell (hESC) aggregates formed by forced aggregation and endoderm differentiation frequency decreases with increasing hESC aggregate size.
The capacity for hESC differentiation along the endoderm lineage increases with decreasing aggregate size
To directly evaluate the effect of aggregate size on endoderm concentration during early differentiation, size-specified aggregates were assessed at D0, D1, D3, and D6 of differentiation for frequency of cells expressing the endoderm protein markers FoxA2 and GATA6 by flow cytometry (Fig. 2C, D). Figure 2C depicts representative dot plots from the flow cytometry analysis. A trend emerged in which the frequency of FoxA2+ and GATA6+ cells increased with decreasing aggregate size at each timepoint, with the smallest aggregates that were formed at an initial size of 100 cells consistently exhibiting the highest frequency of endoderm marker-expressing cells (Fig. 2D). Similar protein expression profiles were observed for both endoderm markers tested at all three aggregate sizes, with maximum frequencies generally reached by D3 of the timepoints that were analyzed. FoxA2 expression reached a maximum frequency of 0.073±0.013 in the smallest aggregate condition (initiated with 100 cells) on D3, whereas the largest aggregates (formed with 4000 cells) consistently displayed the lowest frequencies of FoxA2-expressing cells never exceeding 0.03 (Fig. 2Di). Maximum expression frequencies for GATA6 were also observed on D3, where the highest level of cells expressing the protein (0.1193±0.0132 of cells) was observed in the smallest aggregate size condition (initiated with 100 cells) and the lowest levels (0.064±0.0071) in the largest aggregates (initiated with 4000 cells) (Fig. 2Dii). Supplementary Figure S3 also shows the time course of Foxa2 staining in EBs with 100 cells per aggregate and 1000 cells per aggregate on days 4, 5, and 6. These data clearly indicate the spatial relationship and positioning of the endoderm cells as EBs grow in the size.
Day 3 aggregate diameters were measured by analyzing phase-contrast microscopy images of the aggregates, and these values were then used to estimate the ratio of the volume of the outer layer of cells (OV) to the total aggregate volume (TV). To estimate the OV, the diameter of a single cell was assumed to equal 10 μm. 37 The OV/TV ratios for aggregates generated with 100, 1000, and 4000 cells are presented in Figure 2E. Under all aggregate size conditions, the OV/TV ratios decreased with increasing aggregates size; however, the OV/TV ratios were higher than the corresponding FoxA2-expressing and GATA6-expressing cell frequencies (100 cell aggregates: OV/TV ∼ 0.6, FoxA2pos ∼ 0.07, GATA6pos ∼ 0.12; 1000 cell aggregates: OV/TV ∼ 0.3, FoxA2pos ∼ 0.05, GATA6pos ∼ 0.09; 4000 cell aggregates: OV/TV ∼ 0.2, FoxA2pos ∼ 0.03, GATA6pos ∼ 0.06), indicating either that endoderm cells do not make up a complete layer surrounding the aggregate, or that cells in the outer layer may not be as densely packed as cells located in the interior of the aggregate or may vary in size from inner cells. It can be observed in the confocal microscopy analysis (Fig. 2B) that not all aggregates contain an outer layer of endoderm cells and that in many aggregates endoderm cells do not make up the full layer of outer cells. Importantly, this analysis demonstrated that the OV/TV ratio decreased with increasing aggregates size similar to the trend observed in tracking FoxA2+ and GATA6+ cells on D3, and consistent with our hypothesis.
Varying the frequency of ExE progenitor cells in hESC aggregates influences cardiac differentiation efficiency in size-controlled hESC aggregates independently of aggregate size
As it was observed that cardiac induction and differentiation could be modulated by varying hESC aggregate size and that there was an inverse relationship between aggregate size and the frequency of endoderm cells present during the first 6 days of differentiation, we next examined the possibility that the frequency of endoderm cells can be optimized to maximize cardiomyocyte differentiation efficiency. To vary ExE frequency independently from aggregate size, we manipulated the expression of SOX7, a transcription factor that is required for ExE differentiation,34,35 in the input hESC population (Fig. 3A). To study the effect of increasing ExE frequency, 1000 cell aggregates were generated with varying ratios of normal hESCs to SOX7-overexpressing (O/E) hESCs (ExE progenitors) (Fig. 3Bi). Conversely, the effect of reducing ExE frequency was examined by generating 1000 cell aggregates with varying ratios of normal hESCs to knockdown SOX7 hESCs (kd-SOX7) (Fig. 3Bii). SOX7 expression was induced in genetically engineered CA1 hESC lines (SOX7 O/E) following transient expression of Cre recombinase. An ExE progenitor phenotype in transgenic cell lines was confirmed by expression of SOX7, GATA6, and SOX17 proteins, markers that are associated with an ExE phenotype (Supplementary Fig. S4). siRNA transfection was employed to silence SOX7 expression in hESCs. SOX7 siRNA transfection was validated by demonstrating SOX7 protein knockdown in the SOX7 O/E cells that were transfected with the siRNA (Supplementary Fig. S5). We further validated that knock down effect of SOX7 siRNA maintained during the EB differentiation on day 3, which resulted in down regulation of GATA6 in both 100% SOX7 siRNA-transfected EBs and 50% mixed EBs compared to control (Supplementary Fig. S6). SOX7 O/E hESCs and kd-SOX7 hESCs were mixed at varying ratios with the parental non-Cre-transfected CA1 hESCs transfected with nontargeting siRNA.
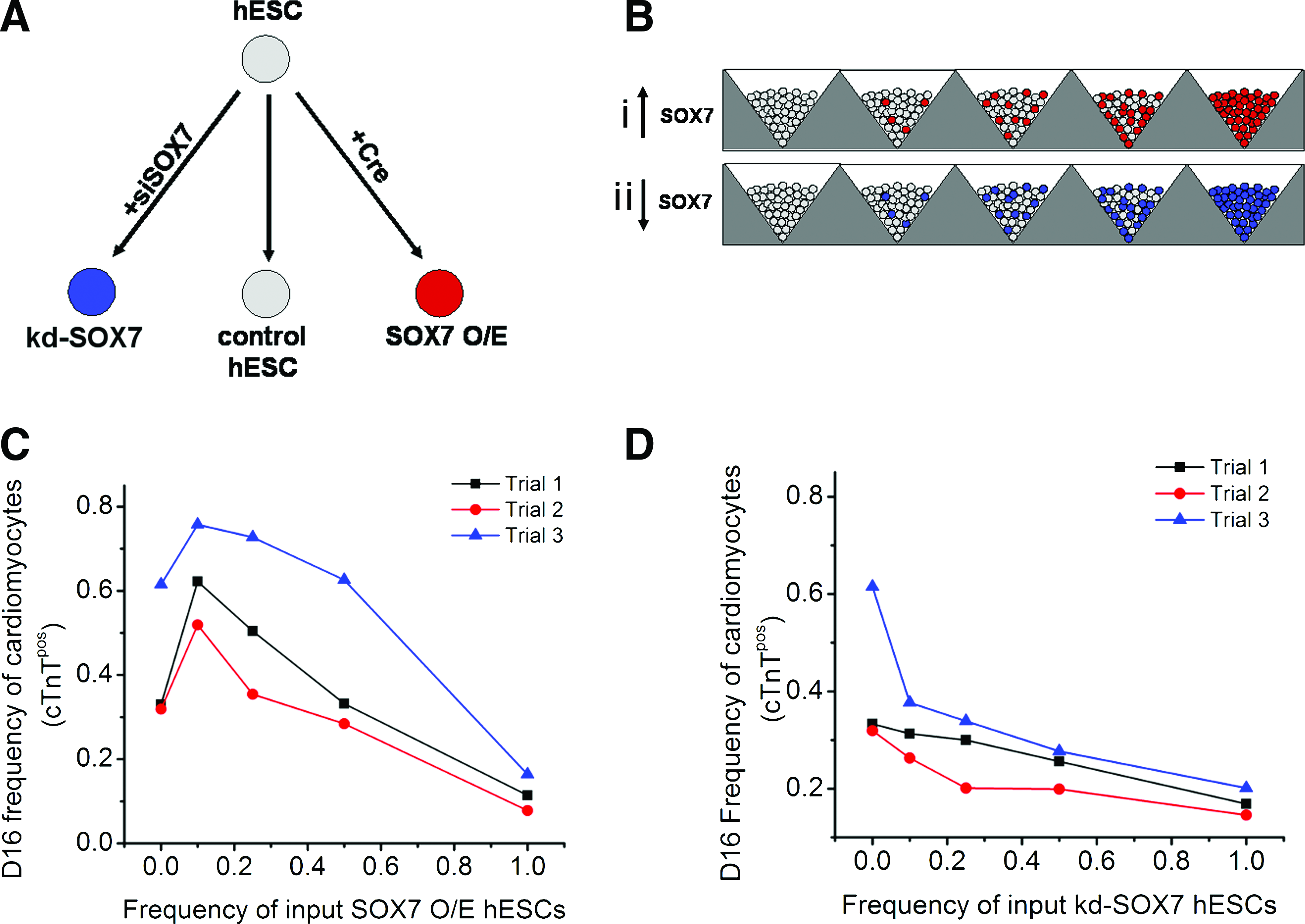
The frequency of input ExE cells has an aggregate size independent effect on cardiomyocyte differentiation efficiency.
Spontaneously contracting aggregates were observed in all conditions, regardless of the ratio of SOX7 O/E or kd-SOX7 hESCs in the starting population, suggesting that all cell lines have the capacity for cardiac induction. The frequency of cTnTpos cells on D16 was measured to track cardiomyocyte differentiation frequency with respect to input ExE progenitor (SOX7 O/E hESC) or kd-SOX7 hESC frequency. It was observed that cardiomyocyte differentiation efficiency varied in response to different input frequencies of ExE progenitors (Fig. 3C). Maximum cardiac differentiation (frequency=0.6327±0.1194) was observed in aggregates generated with a frequency of 0.10 SOX7 O/E hESCs (Fig. 3C). In the condition initiated with 0.50 input ExE progenitor frequency, cardiomyocyte differentiation efficiency returned to control levels (∼0.40). As expected, in aggregates initiated with only ExE progenitors (frequency SOX7 O/E hESCs=1.0), cardiomyocyte differentiation was significantly reduced. A negative response to increasing frequencies of kd-SOX7 hESCs was observed with respect to cardiomyocyte differentiation efficiency (Fig. 3D), wherein all aggregates containing kd-SOX7 hESCs exhibited reduced cardiomyocyte frequencies compared to the control condition reaching a minimum frequency of 0.172±0.0276 cTnTpos cells in aggregates generated with a frequency of 1.0 kd-SOX7 hESCs, indicating that although SOX7-expressing cells do not efficiently differentiate toward the cardiac lineage, their presence promotes cardiogenesis. From these findings it was concluded that by manipulating the input frequencies of ExE progenitors, cardiomyocyte differentiation could be attenuated or increased even in the presence of growth factors that specifically direct cardiac commitment and expansion, suggesting that endogenously generated ExE cells exert an inductive effect on differentiating hESCs in an aggregate size-dependent manner.
Discussion
Using a variety of methods to control aggregate size, a number of reports have demonstrated that this parameter influences hPSC differentiation along the cardiac lineage.25,26,30,33 A significant observation that has been made consistently across these studies is that there is a specific range of aggregate sizes, usually defined by aggregate diameter, in which the frequency of contracting aggregates and cardiac-associated gene expression is maximized. In aggregates generated by either forced aggregation 26 or in microwells, 33 it has been shown that the frequency of contracting aggregates and expression of cardiac-associated genes was maximized in aggregates with diameters in the 250–350 μm range, and decreased in smaller and larger aggregates. Even under conditions in which aggregate size has been optimized, however, cardiac differentiation efficiency appears to be quite low in these studies, with observed frequency of contracting aggregates typically <0.25,26,33 and in the few cases where cardiomyocyte differentiation frequency was evaluated, frequencies of cells that express cardiac-specific proteins are <0.05.25,33
The present study is the first report investigating the effect of hESC aggregate size on cardiomyocyte differentiation under defined conditions that promote cardiogenesis. In addition, more robust metrics for aggregate size and cardiac differentiation efficiency were employed than in the previously described reports. Aggregate size was defined in terms of the number of input cells per aggregate, not aggregate diameter, which can represent different cell numbers between runs depending on cell density. To more accurately determine cardiomyocyte differentiation efficiency, we analyzed the frequency of cells expressing cardiac progenitor and mature cardiac markers, instead of counting the frequency of beating aggregates or determining relative gene expression levels of cardiac markers. Similar to previous reports, we observed a trend wherein cardiomyocyte induction and differentiation was attenuated in aggregates that were too small or too large even in the presence of growth factors that specify the cardiac lineage, and midsize aggregates (initiated with 1000 cells) produced the highest frequencies of D5 cardiac progenitors (∼0.50) and D16 cardiomyocytes (∼0.25–0.40). While the trend we observed was similar to previous studies, under defined conditions that specify cardiac induction we achieved significantly higher levels of cardiomyocyte differentiation in hESC size-controlled aggregates than in any prior reports.
From the findings presented here we propose a cellular mechanism to describe how aggregate size affects cardiomyocyte differentiation efficiency. We reasoned that aggregate size determined the level of endogenous cardiac-promoting ExE that developed by a simple geometric relationship: the ratio of the surface area to volume of a sphere decreased with increasing sphere diameter. In this case, aggregate size could be varied to directly control the frequency of ExE differentiation, which subsequently could impact the efficiency of cardiac differentiation. Tracking endoderm frequency over the first 6 days in size-specified aggregates, we observed a relationship in which the level of endoderm cell development decreased with increasing aggregate size, supporting the hypothesis that the level of endoderm differentiation is related to the ratio of outer cells to inner cells in the cell spheroid. A direct relationship between frequency of ExE cells during aggregate formation and cardiomyocyte differentiation efficiency was confirmed by monitoring cardiomyocyte frequency in size-specified hESC aggregates that were generated with controlled frequencies of cells that either overexpressed SOX7 transcription factor or were transfected with siRNA blocking SOX7 transcription. Our findings support the concept that there is a geometric relationship between hESC aggregate size and the extent of ExE development during early differentiation that subsequently impacts cardiac induction and differentiation efficiency.
It has been suggested that the effect of hESC aggregate size on cardiac differentiation is influenced by the diffusion of inductive molecules throughout the aggregate. 33 Certainly, during embryogenesis spatial gradients of several signaling molecules are involved in guiding cardiac development, many originating from ExE cells as previously described. The inductive characteristic of the endoderm has been attributed to transforming growth factor-β superfamily and fibroblast growth factor family growth factors that have been reported to be involved in cardiac differentiation.12,13,38–40 It has been shown using mouse (m)ESCs that inhibition of primitive endoderm with diffusible leukemia inhibitory factor during cell aggregation attenuates cardiomyocyte differentiation and that cardiomyogenesis could be rescued with parietal endoderm conditioned medium, 18 suggesting that the primitive endoderm secretes factors that contribute to cardiac commitment in a paracrine, leukemia inhibitory factor-independent manner. In mESC aggregates generated by mixing different ratios of Dox-inducible GATA4-overexpressing mESCs with the untransfected parental cell line, 41 GATA4-overexpressing cells develop into SOX17+ cells that secrete bone morphogenetic proteins and Dickkopf-related protein-1 (DKK1) to promote cardiac induction in the non-GATA4-overexpressing cells. It has also been recently observed in the mESC system that Wnt5A gene expression is modulated in an aggregate size-dependent manner wherein lower gene expression in larger aggregates (generated from 450-μm-diameter microwells) corresponded to enhanced cardiogenesis based on frequency of beating aggregates and cardiac gene expression. 32 In the same study it was shown that GATA4 mRNA was highly expressed in larger aggregates. It may be that the GATA4-expressing cells secrete enough Dickkopf-related protein to inhibit Wnt5A in the larger aggregate condition. Secreted protein, acidic, rich in cysteine, a matricellular glycoprotein that is highly expressed in the developing heart,42,43 is another candidate molecule secreted by parietal endoderm that has been shown to promote early myocardial differentiation in mESC aggregates.44,45 mESC aggregates cultured in parietal endoderm-conditioned medium exhibit enhanced cardiac differentiation 44 —an effect that is abrogated with the addition of anti-secreted protein, acidic, rich in cysteine antibodies in a concentration-dependent manner.
The investigation into the effect of aggregate size on endoderm differentiation showed that the frequency of cells expressing the pan-endodermal markers FoxA2 and GATA6 decreased with increasing aggregate size during the first 6 days of differentiation. The subsequent analysis assaying the effect of SOX7 O/E hESC frequency on cardiac differentiation indicated that ExE cells specifically promote cardiogenesis but did not eliminate the possibility that definitive endoderm (DE) cells also contribute to the aggregate size effect observed. Reports specifically examining the effect of DE on cardiac development have been sparse and contradictory.46–52 Similar to the SOX7 O/E hESC cell lines used here, hESCs have also been genetically engineered to overexpress SOX17. 35 It has been demonstrated that SOX17 O/E hESCs are DE progenitor cells exclusively and incapable of ExE differentiation.53–55 Therefore, using the system described herein, the effect of DE cells on cardiac commitment during hESC differentiation can be examined by generating hESC aggregate cocultures containing hESCs and SOX17 O/E cells. It would be of particular interest to contrast the effects of DE and ExE on different cardiac cell types. During gastrulation, cardiogenesis occurs in two separate progenitor cell populations that arise from a common progenitor. 56 The first cardiac progenitors are specified in the anterior lateral plate mesoderm to form the primary heart field and subsequently give rise to cardiomyocytes belonging to the left ventricle and atria. Pharyngeal mesoderm cells give rise to the secondary heart field that is located medial to the primary heart field and contribute to the right ventricle and outflow tract of the heart. Given that there is evidence for both ExE and DE promotion of cardiogenesis and that the two heart fields arise at different times and locations in the embryo, it may be that ExE cells play a role in specifying cardiac induction in the primary heart field, whereas DE cells are involved in guiding commitment of the secondary heart field.
In conclusion, we have established a robust system for hESC aggregate size-controlled cardiac induction and differentiation. Upon directly regulating endogenous signaling by controlling aggregate size, we not only achieved consistent cardiac induction between runs, but also significantly enhanced cardiac induction efficiency achieving yields as high as 75% cTnTpos. Using this system, it was determined that aggregate size influences endoderm differentiation efficiency, and that the concentration of ExE in size-controlled aggregates directly influences cardiac differentiation efficiency. This system establishes a basis to examine the effect of endoderm-secreted factors on cardiogenesis in aggregate size-controlled hPSC aggregates.
Sources of Funding
Heart and Stroke Foundation of Ontario (P.W.Z.) and the Canadian Institute of Health Research (MOP-57885).
Footnotes
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.