
Abstract
Tissue-engineered heart valves are prone to early structural deterioration. We hypothesize that cell–scaffold interaction and mechanical deformation results in upregulation of genes related to osteogenic/chondrogenic differentiation and thus changes extracellular matrix (ECM) composition in human bone marrow mesenchymal stem cell (hBMSC)-derived tissue-engineered grafts. hBMSC were expanded and seeded onto poly-glycolic acid/poly-lactic acid scaffold for 14 days. Seeded tissue-engineered constructs (TEC) were subjected to cyclic flexure for 24 h, whereas control TEC was maintained in roller bottles for the same duration. hBMSC, TEC, and mechanically deformed TEC were subjected to gene-array and histological analysis. Expression levels of RNA and/or protein markers related to chondrogenesis (Sox9, MGP, RunX2, Col II, Col X, and Col XI) and osteogenesis (ALPL, BMP2, EDN1, RunX1, and Col I) were increased in TEC compared to unseeded hBMSC. Histological sections of TEC stained positive for Saffranin O, alkaline phosphatase activity, and calcium deposits. The expression levels of the above gene and protein markers further increased in deformed TEC compared to static TEC. Cell–scaffold interactions and mechanical stress results in gene expression suggestive of endochondral-ossification that impact upon ECM composition and may predispose them to eventual calcification.
Introduction
Although the mechanisms underlying calcification of tissue-engineered heart valves (TEHV) have not been well described, cell–scaffold interactions and mechanical stress have been implicated. 5 Seeding of BMSC onto PGA and PLA scaffolds stimulates deposition of sulfated glycosaminoglycans, whereas cyclic flexure enhances production of collagen type I. 6 Mechanical stress contributes to calcification of bioprosthetic valves, and may also play a role in calcification of TEHV constructs. 7
Several recent studies have demonstrated osteogenic differentiation of human BMSC (hBMSC) upon mechanical stimulus. 8 The effect of mechanical stimulus on changes in gene regulation of hBMSC-seeded scaffolds has not been explored. We hypothesize that cell–scaffold interaction and mechanical deformation increase expression of genes specific to osteogenic- and chondrogenic-extracellular matrix (ECM) formation in tissue-engineered constructs (TEC) constructed from hBMSC seeded onto PGA/PLA polymer scaffold. Further, we hypothesize that changes in gene regulation occur early during development of TEC. This study was designed to evaluate the early changes in gene regulation resulting from interaction of hBMSC with PGA/PLA scaffold and cyclic mechanical flexure.
Materials and Methods
Cell culture and seeding of TEC
Commercially available hBMSC (Lonza) were expanded for 3–4 passages in culture medium (Dulbecco's modified Eagle's medium low-glucose media [Invitrogen]), with 10% fetal bovine serum (Sigma-Aldrich), 1×antibiotics, and 1% HEPES (Invitrogen) at 37°C and 5% CO2. Nonwoven PGA/PLA (50:50 blend) scaffolds (Concordia Medical) were cut into 1×3 cm sections and fixed onto a 2×5 cm poly-tetra-fluoro-ethylene plastic sheet. The fiber diameter is 12–15 μm for PGA and 20 μm for PLA. The pore size for the fabric is 40–60 μm. The scaffold tabs (1×3 cm) were gas sterilized with ethylene oxide. hBMSC were seeded onto the sterilized scaffold constructs at a density of 2×106/cm2 and maintained in roller bottles at 37°C and 5% CO2 for 12–14 days with fresh culture media supplemented with beta-fibroblast growth factor (2.5 μg/500 mL) and vitamin C (AA2P) 41 mg/500 mL (Sigma-Aldrich) replacement every 2–3 days. TEC properties and histological analysis have been previously described.9,10
Mechanical deformation using STREX device
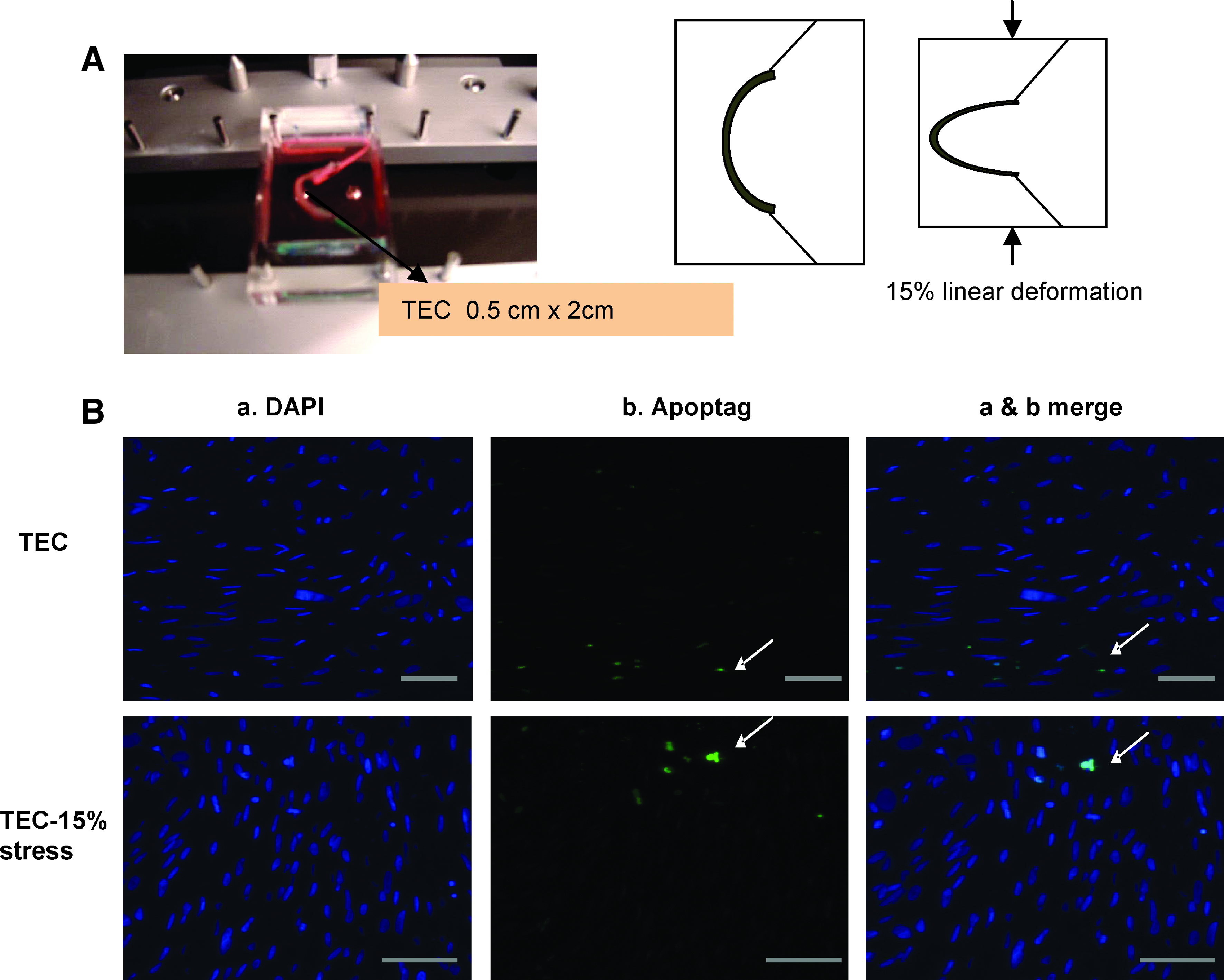
The seeded TEC were transferred onto a flexible platform designed for cyclic mechanical deformation of TEC strips (Fig. 3A) in seeding media at 37°C and 5% CO2, and mounted onto the STREX device ST-140-10 (STREX Inc.). Cyclic axial mechanical deformation of TEC (0.5×2 cm section) was performed for 24 h at a frequency of 1 Hz at 15% linear deformation, resulting in change in curvature (Δκ) of 0.7 mm–1. The deformation parameters were selected to be within the range of previously reported estimations of normal physiologic valve deformation conditions. 8 Control TEC was maintained in seeding media for the same duration.
Phenotypic and functional characteristics of hBMSC
Before seeding of TEC, hBMSC were analyzed for the expression of cell surface markers (CD29, CD90, CD44, CD105, CD31, CD34, and CD14 from R&D Systems) using flow cytometry as previously described. 11 Briefly, hBMSC were trypsinized and washed twice in cold phosphate-buffered saline (PBS). About 1×105 hBMSC were labeled with 2 μg/mL fluorescein isothiocyanate-conjugated monoclonal antibody for each tested marker and incubated for 30 min on ice. Labeled hBMSC were washed three times in cold PBS followed by resuspension in 100 μL of 2% paraformaldehyde and analyzed on a BD FACSCalibur Flow Cytometer (BD Biosciences). Multi-potency of hBMSC was determined using commercially available kits for induction of adipogenesis, osteogenesis, and chondrogenesis (Lonza) as per manufacturer's protocols. Briefly, for adipogenesis, hBMSC were cultured for 3 weeks in adipogenesis induction/maintenance media on 6-well plates with media changes every 2–3 days and assessed for adipocytes containing lipid droplets using oil-O red staining. For osteogenesis, hBMSC were cultured in osteogenesis induction media for 17 days on 6-well plates and stained with alizarin red to detect osteocytes containing calcium deposits. About 2×105 hBMSC were centrifuged into pellets, cultured in chondrogenesis induction media containing transforming growth factor (TGF)-β1 for 28 days, sectioned onto slides, and stained with Saffranin-O for detection of cartilage matrix. To assess cartilage matrix development on the seeded scaffold, histological sections of TEC were deparaffinized and stained with Saffranin-O.
Alkaline phosphatase activity
Alkaline phosphatase activity was determined in unseeded hBMSC and TEC subjected to mechanical deformation or static conditions utilizing a commercially available kit (Sigma-Aldrich). Briefly, plated hBMSC or hBMSC-seeded tissue sections were fixed for 30 s in Citrate-acetone-formaldehyde fixative solution at room temperature and gently rinsed for 45 s in de-ionized water. Slides were stained with alkaline–dye mixture for 15 min and counterstained with hematoxylin solution. Stained sections were examined by microscopy.
Immunohistochemistry
Cellular collagen II (COL II) expression was assessed in the histological sections obtained from hBMSC pellets, and TEC subjected to deformation or static conditions using a commercially available COL II staining kit (MDBIOSciences). Briefly, sections were deparaffinized and hydrated with PBS. Antigen retrieval step using 10 mM citrate buffer (pH: 6.0) was performed before staining with biotinylated monoclonal antibody to COL II for 1 h followed by incubation with Strep-horseradish peroxidase (HRP). Stained sections were developed using 3,3′ diaminobenzidine.
Apoptosis assessment
The rate of apoptosis of TEC subjected to mechanical deformation or static conditions was determined using ApopTag In-Situ Apoptosis detection kit (Chemicon International, Millipore Co.). Briefly, the paraffin-embedded sections were deparaffinized using xylene and absolute ethanol, and treated with protein digesting enzyme. Sections were incubated in terminal deoxynuecleotidyl transferase at 37°C for 1 h. The sections were exposed to fluorescein-labeled anti-dioxigenin antibody conjugate for 30 min at room temperature, counterstained with 4′,6-diamidino-2-phenylindole, and observed by florescence microscopy. Apoptosis rate was calculated as the number of apoptotic cells per high power field, averaged over 3 separate fields (apoptosis rate=[number of dead cells per field]×100/number of live cells per field). t-test was performed (n=3) on rate of apoptosis in deformed TEC compared to control TEC.
Microarray analysis
To examine the effect of cell–scaffold interaction and cyclic mechanical deformation upon changes in gene expression profile in TEC, cDNA gene chip array was utilized. Comparisons of gene profile were performed between hBMSC versus unstressed TEC and static TEC versus TEC subjected to 24 h of mechanical deformation. Total RNA was isolated from specimens using RNeasy kit (Qiagen Inc.). After quantitative and qualitative assessment of extracted RNA, double-stranded cDNA was synthesized. Biotin-labeled cRNA was obtained by in vitro transcription of synthesized double-stranded cDNA utilizing a linear amplification kit (Applied Biosystems). Chip hybridizations, washing, Cy3-streptavidin (Amersham Biosciences) staining, and scanning were performed on the Illumina Bead Station 500×platform (Illumina Inc.) using reagents and protocols supplied by the manufacturer. cRNA samples were hybridized in duplicates on Illumina Human WG-6 v 3.1 BeadChips. All basic expression data analyses were carried out using the manufacturer's BeadStudio (3.1) software. Raw data were background-subtracted and normalized using the “rank invariant” algorithm. Values below the detection limit were arbitrarily set to the level of threshold detection to avoid nonsense values for expression ratios. Fold change ≥1.8 with p-value <0.01 were considered significant. Pathway and Gene Ontology analyses were carried out using DAVID Bioinformatics resources 2008 (National Institute of Allergy and Infectious diseases [NIAID], NIH) and Ingenuity systems software (Ingenuity systems, Inc.).
Real-time polymerase chain reaction (PCR)
Gene upregulation detected by microarray analysis was confirmed by real-time PCR. One step real-time PCRs were performed using SYBR Green detection kit (Applied Biosystems) and the Stratagene Mx3000P® QPCR system (Stratagene). The specific primers for the genes of interest were designed using Oligoperfect™ software (Invitrogen). The following conditions were used to run the reaction: 30 min at 48°C, 10 min at 95°C for 1 cycle, and 15 s at 95°C and 1 min at 60°C for 40 cycles. Relative expression levels of the genes were analyzed using 2−ΔΔCT method in comparison to the expression of the endogenous control gene GAPDH (Invitrogen). Each sample was analyzed in triplicates.
Western blotting
Specific genes upregulated in the microarray analysis were further analyzed for protein expression using immunoblotting. Protein lysates were obtained from hBMSC, unstressed TEC, and TEC subjected to mechanical deformation using lysis buffer (Sigma) and separated on 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Transferred membranes were blocked and labeled with human monoclonal Col II, Col I, Sox9, and tubulin primary antibodies (1:1000 dilution; R&D Systems) for 3 h at room temperature and stained with HRP-conjugated secondary antibodies for 1 h at room temperature. Bound peroxidase activity was detected using Western Blot Luminol reagent as per manufacturer's instructions (Santa Cruz Biotechnology). For quantification, the band intensities were determined using ImageJ software (NIH). Protein band intensities were normalized to tubulin and fold-change was calculated comparing cells to TEC and TEC to mechanically deformed TEC.
Statistical analysis
Rosetta Resolver system (Rosetta Biosoftware) statistical software was utilized to calculate fold-change for analyzing differential gene expression by microarray. Fold change ≥1.8, and values of p<0.01 were considered significant. For other statistical analysis, results were expressed as mean±standard error of the mean. Comparisons between groups were made with a Student's t-test. p-values <0.05 are considered significantly different.
Results
Phenotypic and functional characteristics of hBMSC
hBMSC (n=6 sets) just before seeding onto scaffolds were assessed for phenotypic and functional characteristics by flow cytometry and multi-potency assays, respectively. Cells expressed surface markers CD29, CD90, CD44, and CD105, but not endothelial progenitor cell markers CD31 and CD34, and leukocyte markers CD14 and CD45 (Table 1). Functional assays confirmed the multi-potency features of the hBMSC as detected by their ability to differentiate into adipocytes, osteocytes, and chondrocytes (Fig. 1).
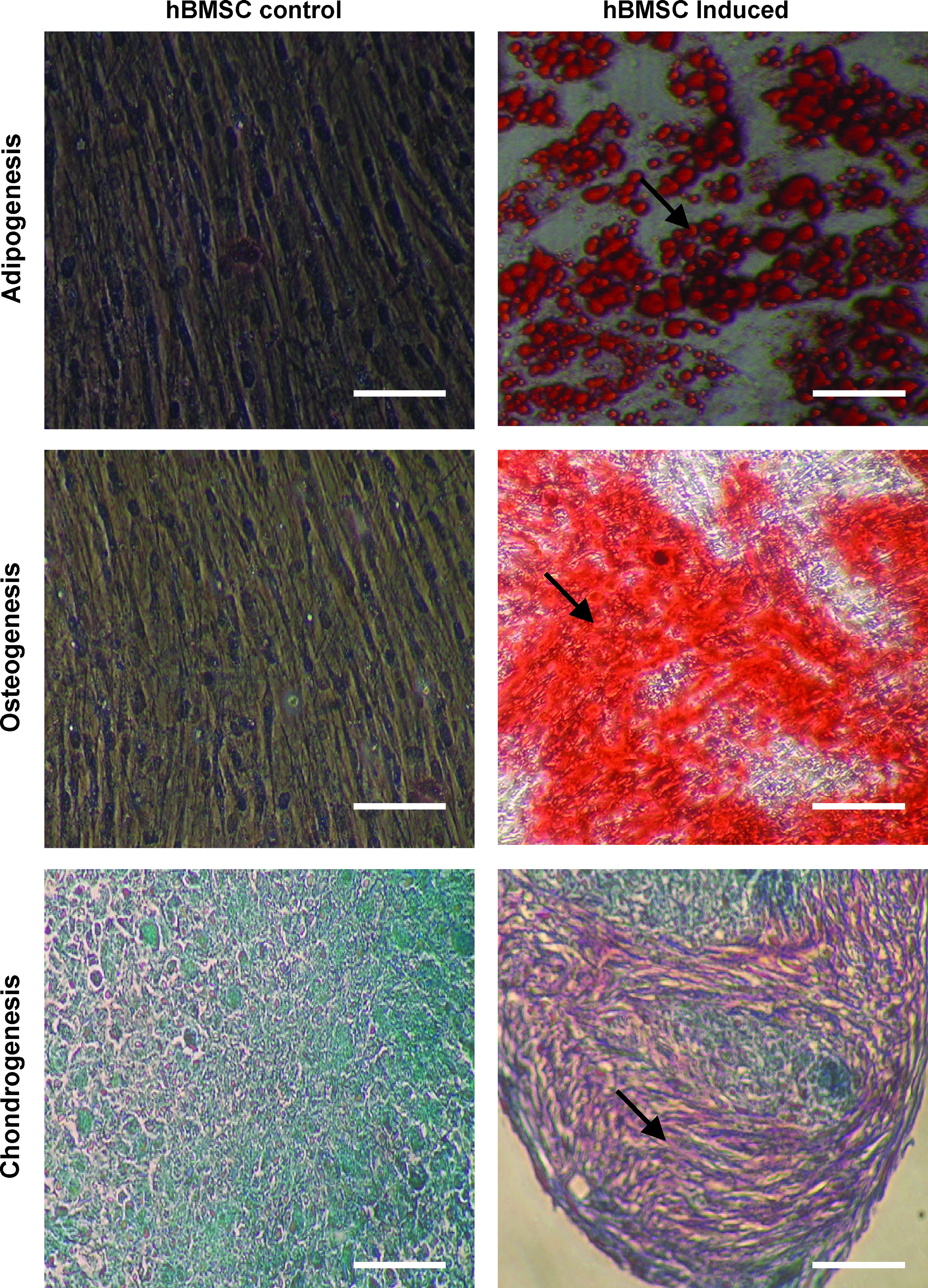
Demonstration of multi-potency features of hBMSC before seeding onto PGA/PLA scaffold. Induction of adipogenesis (lipid droplets are stained red [indicated by arrows] with oil-O-red staining); osteogenesis (staining with alizarin red [arrows] indicates calcium deposition upon induction; and chondrogenesis (sectioned pellet stained for cartilage tissue [arrows] with Saffranin-O). hBMSC culture before induction (left) and after induction (right). 20×magnification for all images. Scale bars, 100 μm. hBMSC, human bone marrow mesenchymal stem cells; PGA, poly-glycolic acid; PLA, poly-lactic acid.
Functional clustering of upregulated genes in unstressed TEC compared to unseeded hBMSC
Comparison of micro array gene profile from unstressed, hBMSC-seeded TEC (n=6) to that from unseeded hBMSC (n=6) revealed 1354±124 upregulated genes and 186±32 downregulated genes. The genes that were upregulated ≥1.8-fold (with p<0.01 by ANOVA) in TEC compared to hBMSC were clustered based on gene function using DAVID 2006 Bioinformatics database. The ECM forming genes and cartilage/bone development gene clusters were predominantly affected by cell–scaffold interaction. The list of genes in these clusters is shown in Table 2.
↑ indicates gene upregulation of >1.8-fold in unstressed TEC compared with unseeded human bone marrow mesenchymal stem cells (p<0.01 by ANOVA). ANOVA, analysis of variance; hBMSC, human bone marrow mesenchymal stem cells; TEC, tissue-engineered constructs.
Many of the genes that were upregulated by seeding of hBMSC onto scaffold were also upregulated by osteogenic and/or chondrogenic induction of hBMSC. Genes that were upregulated during osteogenic and chondrogenic differentiation of hBMSC are displayed for comparison (Table 2). The upregulated genes from the osteogenic/chondrogenic clusters (transcription factor SOX9, growth factor TGF-β3, ECM proteins MGP, TNC, and Col X) detected by microarray were further confirmed by real-time PCR (Fig. 2A). Western blot analysis confirmed overexpression of the above proteins for Sox9 (n=3; 3.4-fold increase; p=0.002) and Col II (n=3; 3.8-fold increase; p=0.006) in TEC compared to hBMSC (Fig. 2B).

Confirmation of gene and protein upregulation using real-time PCR and western blotting.
Gene expression profile in TEC subjected to mechanical deformation
The rate of apoptosis was assessed in TEC subjected to mechanical deformation compared to static TEC. Cyclic flexure of 15% deformation at 1 Hz resulted in minimum apoptosis compared to unseeded hBMSC (8.8% vs. 6.8%, respectively, p=0.42) (Fig. 3B). To examine the effect of cyclic mechanical deformation upon gene expression profile in TEC, gene microarray was compared between TEC subjected to mechanical deformation or static conditions. When the genes that were significantly upregulated (>1.8-fold, p<0.01) by mechanical deformation were organized into functional gene clusters, the ECM and osteogenic/chondrogenic gene clusters emerged as the most prominent clusters. The list of upregulated genes are shown in Table 3. Real-time PCR further confirmed upregulation (Fig. 4A). By western blot analysis, collagen I protein, known to be expressed predominantly by osteocytes, was also expressed at elevated levels (n=3; 12.4-fold; p=0.0004) in mechanically deformed TEC compared to static TEC (Fig. 4B).

Confirmation of gene and protein upregulation for TEC versus mechanically stressed TEC using real-time PCR and western blotting.
↑ indicates gene upregulation of >1.8-fold in TEC compared with mechanically deformed TEC (p<0.01 by ANOVA).
Histological analysis of TEC
To assess for cartilage and bone matrix deposition, histological sections of TEC subjected to cyclic flexure or static conditions were stained with Saffranin O and alizarin red, and assessed for alkaline phosphatase activity. Saffranin O staining was detected in TEC 2 weeks after seeding, and TEC subjected to mechanical deformation for 24 h demonstrated more robust staining compared to static TEC (Fig. 5A). By immunohistochemical staining, Col II was detected in TEC under static conditions or mechanical deformation (Fig. 5B).
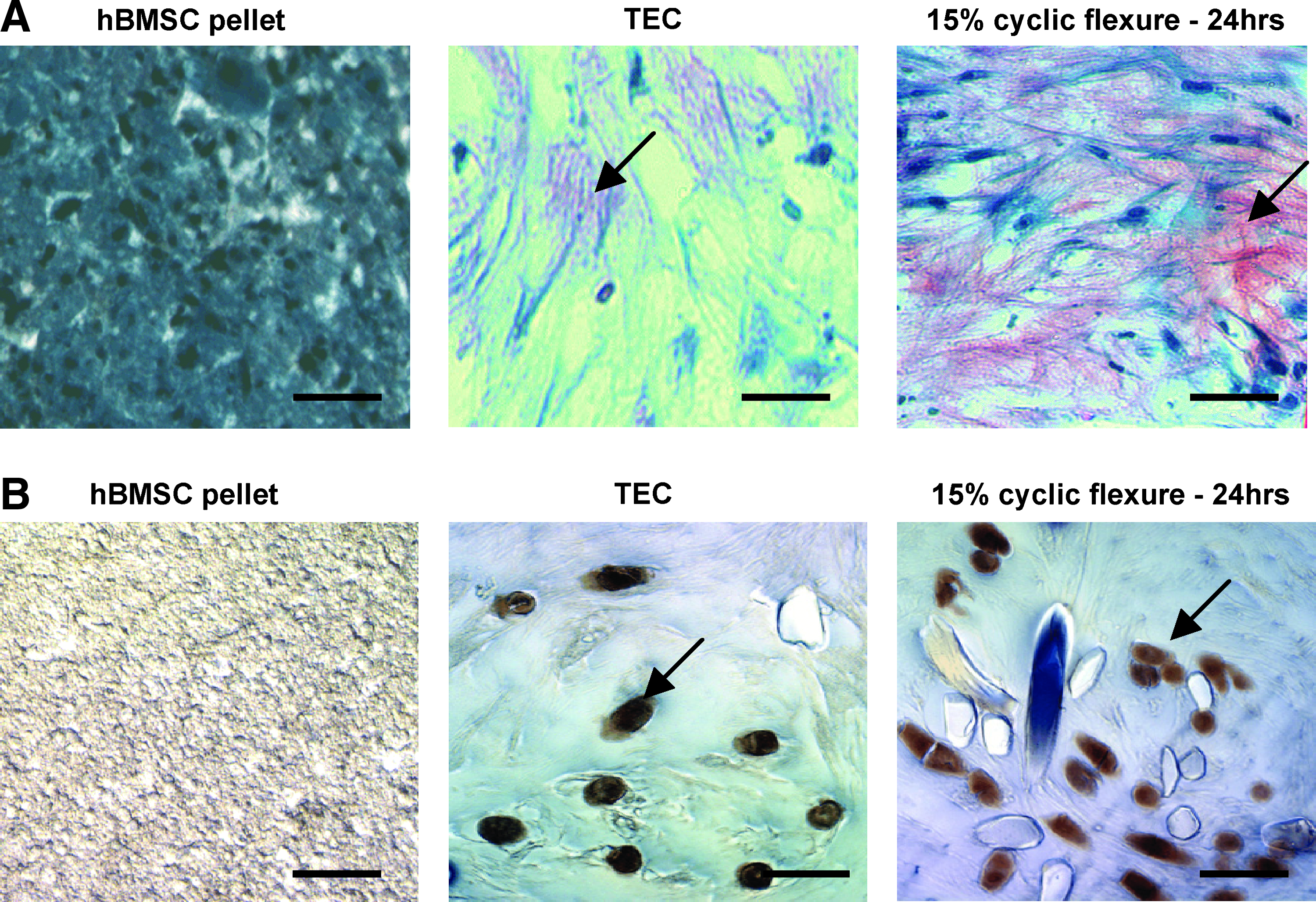
hBMSC pellets, static TEC, and mechanically deformed TEC were stained for cartilage matrix formation with
Alkaline phosphatase, an early marker of osteogenic matrix formation, was more prominent in TEC compared to hBMSC, as well as in TEC subjected to mechanical deformation compared to static conditions (Fig. 6A). Histological sections were also tested for calcium deposition using alizarin red staining in TEC subjected to mechanical deformation or static conditions (Fig. 6B). Calcium deposition was observed in both groups of TEC.

hBMSC-plated culture, static TEC, and deformed TEC were assessed for alkaline phosphatase expression
Discussion
This study demonstrates by gene expression and histological analysis that cell–scaffold interaction and mechanical deformation of hBMSC-derived TEC results in an ECM composition and gene expression profile commonly associated with chondrogenic and osteogenic matrix formation. This process is initiated in the early stages of tissue-engineered valve construction merely by seeding hBMSC onto scaffold PGA/PLA polymer material. The cell–scaffold interaction and mechanical deformation may be stimuli for deposition of matrix of a cartilaginous/osteogenic phenotype, which may predispose the TEC to biomineralization and early failure.
Comparison of unseeded BMSC to seeded TEC revealed increased expression of chondrogenic and osteogenic genes (transcription factor SOX9, growth factor TGF-β3, ECM proteins MGP, TNC, and Col X) and proteins (SOX9 and Col II) in the latter. In addition, Saffranin O staining, which is used to quantify proteoglycan content in cartilaginous tissue, was increased in TEC compared to unseeded BMSC. Together, these findings suggest that BMSC are undergoing chondrogenic and osteogenic differentiation and deposition of corresponding matrix merely by the process of seeding onto scaffold material. Importantly, this process occurs within 10–14 days after seeding.
Since differentiation of hBMSC into osteogenic or chondrogenic phenotype occurs early during the TEC seeding process, the ultimate progression to unfavorable matrix deposition and calcification post-implantation may be inevitable. TEHV deterioration in vivo has been attributed to host coagulation and inflammatory responses. However, this study suggests that long-term TEC graft dysfunction in vivo may not be merely attributable to host responses, but initiated by the cellular microenvironment at the time of inception of the graft. Thus, therapies to prolong graft durability that target inflammatory or immune pathways after graft implantation may be ineffective in a mature TEC that has undergone transdifferentation at an early phase of development, before implantation.
The choice of cell type, scaffold material, and seeding conditions are important as the initial interaction impacts upon the gene expression profile and type of ECM deposited. In this study BMSC were utilized since many laboratories engaged in development of TEHV have focused on this cell source.12,13 Advantages of BMSC include the ease of isolation and expansion, and ability to obtain and utilize autologous cells for TEHV. Autologous BMSC can be readily isolated by bone marrow aspiration, making BMSC-based tissue engineering clinically feasible. BMSC seeded onto PGA/PLA was chosen since previous work in our laboratory demonstrated excellent cell adhesion, tissue formation, and characteristics favorable for in vivo implantation.6,12 If cell–scaffold interaction provides the stimulus for transdifferentiation of BMSC, then careful testing of various scaffold materials is necessary to determine the optimal material with minimal effects on chondro/osteogenesis for the development of heart valve graft. Cartilage and bone tissue formation has been demonstrated on various types of biodegradable materials, including PGA, PLA, and polycaprolactone (PCL). 14 Scaffold properties such as pore size and strand tension have been shown to alter the differentiation potential of seeded MSC. 15 Alternatively, novel technologies are emerging that eliminate the need for scaffold materials, and may eliminate the deleterious response seen with scaffold seeding. 16
Pharmacologic manipulation aimed at preventing osteo/chondrogenic development during the early phases of construct development may be considered for BMSC-seeded scaffolds. TGF-β has been shown to enhance tissue formation during TEC development, but is also likely to enhance the osteogenic and chondrogenic differentiation of BMSC.17–19 Thus, inhibitors or antagonists of TGF-β such as interleukin-10 may prevent the osteogenic differentiation and deleterious response to scaffold seeding.20,21 Platelet-derived growth factor is another potential candidate since it promotes myofibroblastic differentiation. 22 Co-seeding of endothelial progenitor cells with BMSC at different ratios has been utilized in TEC design to modulate ECM production and cell differentiation.23,24
Comparison of TEC subjected to static conditions versus mechanical deformation revealed increased expression of genes (BMP2, EDN1, MMP3, and CXCL5), proteins (Col I), and alkaline phosphatase in the latter. This suggests an important direct role of mechanical deformation upon the phenotype of engrafted cells, which predisposes to TEHV calcification and degeneration. Although mechanical compression has been found to increase mineralization of BMSC-seeded scaffolds designed for tissue-engineered bone, it has not been associated with expression of osteogenic or chondrogenic genes. 25 This finding implies that deformation-mediated cell signaling and gene expression changes, independent of the host inflammatory and coagulation responses, play a significant role in tissue stiffening and calcification.
The association of mechanical deformation to osteo/chondrogenic gene expression and ECM deposition has several implications. Novel strategies for TEC development utilize bioreactors that subject TEC to various forms of mechanical stress. The observation that bioreactors enhance tissue deposition has led to increased application of pulsatile fluid flow and deformation-based bioreactors. 9 Although such bioreactors may increase tissue formation in BMSC-based TEC, they may enhance expression of osteogenic or chondrogenic genes and ECM. The implications of this study are not limited to the field of tissue engineering, as there is a well-known association between elevated wall stress, turbulent flow, and native or bioprosthetic valvular calcification. Deformation-mediated cell signaling and gene expression changes may play a significant role in native and bioprosthetic valve calcification.
In conclusion, the interactions between seeded hBMSC and PGA/PLA scaffold result in expression of genes and proteins suggestive of endochondral-ossification and mechanical deformation further enhances this process. Cell signaling and gene expression changes, mediated by cell–scaffold interactions and mechanical deformation, may play a significant role in tissue stiffening and calcification in vivo, independent of the host inflammatory and coagulation responses. Future investigation into interventions that limit the cartilaginous and osseous transformation may reduce the risk of TEC calcification and deterioration.
Footnotes
Disclosure Statement
No competing financial interests exist.