
Abstract
This study investigated the effect of thrombospondin-1 (TSP-1) on the formation of cartilage repair tissue in combination with stimulation by osteogenic protein-1 (OP-1). In miniature pigs, articular cartilage lesions in the femoral trochlea were treated by the microfracture technique and either received no further treatment (MFX), or were treated by additional application of recombinant osteogenic protein-1 (MFX+OP-1), recombinant TSP-1 (MFX+TSP-1), or a combination of both proteins (MFX+TSP-1+OP-1). Six and 26 weeks after surgery, the repair tissue and the degree of endochondral ossification were assessed by histochemical and immunohistochemical methods detecting collagen types I, II, X, TSP-1, and CD31. Microfracture treatment merely induced the formation of inferior fibrocartilaginous repair tissue. OP-1 stimulated chondrogenesis, but also induced chondrocyte hypertrophy, characterized by synthesis of collagen type X, and excessive bone formation. Application of TSP-1 inhibited inadvertant endochondral ossification, but failed to induce chondrogenesis. In contrast, the simultaneous application of both TSP-1 and OP-1 induced and maintained a permanent, nonhypertrophic chondrocyte-like phenotype within cartilage repair tissue. The data of this study demonstrate that OP-1 and TSP-1 complement each other in a functional manner. While OP-1 induces chondrogenesis of the ingrowing cells, TSP-1 prevents their further hypertrophic differentiation and prevents excessive endochondral ossification within the lesions.
Introduction
Endochondral ossification is associated with the invasion of blood vessels. Instead, articular cartilage is physiologically an avascular tissue that provides a hypoxic milieu that is typical and probably inevitable for the bradytrophic chondrocytes. 19 Hypoxia is known to be one important factor for maintaining the unique chondrocyte metabolism and phenotype. 20 The resistance of articular cartilage against the invasion of blood vessels may be mediated by the presence of anti-angiogenic proteins, such as thrombospondin-1 (TSP-1), which has been detected in large amounts in articular cartilage.21,22
The role of TSP-1 in articular cartilage has not fully been defined yet. Overexpression of TSP-1 within the joint was shown to inhibit experimental osteoarthritis in a rat model, 23 and we could recently demonstrate that fibrocartilaginous repair tissue lacks in TSP-1, which may allow vascular invasion and secondary endochondral ossification. 21
This study focused on the question if supplementation with TSP-1 could prevent endochondral ossification within cartilage repair tissue that was induced by OP-1.
Materials and Methods
Surgical procedures
Six female adult miniature pigs (Ellegaard) aged 18 months with body weights of 35–40 kg were used in this study.
The animals were anesthetized by intramuscular injection of 30 mg midazolam (Roche) and 300 mg ketamine (Pfizer) followed by ventilation with isoflurane (Baxter Diagnostics) at 2 L/min. In a first surgery, the left knee joint (stifle joint) capsule was opened by a medial parapatellar incision, and the patella was displaced laterally. Each of the six animals was treated by four different approaches that were performed in separate round cartilage defects 5 mm in diameter created in the femoral trochlea using a biopsy punch. All lesions were clearly separated from each other by at least 3 mm of intact cartilage similar to approaches of previous studies.3,21 The cartilage was carefully removed using a curette. Great care was taken to keep the subchondral bone plate provisionally intact. Additionally, five microfractures (“MFX”) (each 1 mm in diameter and a depth of 3 mm) were induced in all lesions to allow ingrowth of mesenchymal cells from the bone marrow.
Afterward, in each of the animals, the microfractured lesions were treated as follows: (1) no further treatment=microfracture control; (2) application of 10 mg type I collagen carrier granules containing 35 μg OP-1 (Osigraft®; Stryker Biotech) (“MFX+OP-1”). The OP-1-containing granules were press-fitted into the microfracture holes and covered with fibrin glue (Beriplast; Aventis). The amount of 10 mg of collagen granules was carefully determined in preliminary experiments to exactly fill the microfracture holes but to exclude any excessive volume of carrier granules that may displace; (3) application of 5 μg of rhTSP-1 (R&D Systems) suspended in 10 μL of sterile PBS and bound to 10 mg of purified bovine type I collagen carrier granules (native component of Osigraft®; Stryker Biotech) (“MFX+TSP”). The TSP-1-containing granules were press-fitted into the microfracture holes and covered with fibrin glue. Five micrograms of rhTSP-1 was the maximal amount to be properly reconstituted in 10 μL PBS and soaked up by 10 mg collagen granules; (4) application of 10 mg of type I collagen granules containing 5 μg TSP-1 and 35 μg OP-1 (“MFX+TSP+OP-1”).
The coverage of all lesions with fibrin glue allowed to provide an immediate clot formation and to avoid diffusion or leakage of the factors applied.
Among the six animals, the different treatment approaches were arranged in an alternating randomized manner within the femoral trochlea. Preceding studies on other miniature pigs confirmed no significant spatial differences with respect to the thickness and morphology of articular cartilage and subchondral bone within the treatment area of the femoral trochlea.3,21
After surgery, the animals were allowed to move freely in their cages. After 20 weeks, the animals were operated on their right knee joints using exactly the same treatment schemes and were sacrificed another 6 weeks later. This treatment scheme allowed two different follow-up periods with 6 weeks for the right knee joint and 26 weeks for the left knee joints. The knee joints were dissected and first assessed macroscopically followed by preparation for histological analysis.
The animal study was approved by the appropriate Institutional and Governmental Review Boards.
Histological and immunohistological assessment
The osteochondral specimens containing the respective repair tissues were cut out of the distal femur using a miniature saw and fixed in 4% paraformaldehyde for 12 h, followed by decalcification in 0.5 M ethylene diamine tetraacetic acid for 3 months. After standard processing, the samples were embedded in paraffin. The specimens were scanned throughout the tissue blocks with serial transverse 5-μm sections and stained with toluidine blue to estimate the proteoglycan content and with Alizarin red to observe calcified tissue and bone structures.
For immunohistochemical detection of TSP-1 and CD31, deparaffinized sections were pretreated either with 0.2% hyaluronidase (Roche) for 15 min and 0.1% pronase (Sigma-Aldrich) for 30 min, or with 10 mM Tris-HCl (pH 10) for 5 min and 0.2% hyaluronidase for 15 min at 37°C, respectively. The sections were then left to react at 4°C over night either with monoclonal mouse anti-human TSP-1 antibodies (Santa Cruz) diluted 1:50, or with monoclonal mouse anti-human CD31 antibodies (Abcam) diluted 1:10, respectively. Both antibodies allow to detect human and porcine antigens. Negative control sections for TSP-1 and CD31 were incubated with isotype normal mouse IgG (Santa Cruz). The sections were incubated with biotinylated anti-mouse secondary antibodies. Bound antibodies were observed using the Super Sensitive Link-Label IHC Detection System with biotinylated anti rabbit Ig, avidin-coupled alkaline phosphatase (Biogenex), and fast red according to the manufacturer's instructions. The sections were counterstained with hematoxylin.
Immunohistochemical detection of type I, type II, and type X collagen was performed as described previously in detail. 24 Briefly, all deparaffinized sections were first pretreated with 0.2% hyaluronidase for 60 min at 37°C. The sections were also treated with 0.2% pronase (Sigma-Aldrich) for 60 min at 37°C for detection of type I and type II collagen, or with protease XXIV (0.02 mg/mL) (Sigma-Aldrich) for 60 min at 37°C for detection of type X collagen. Sections were then exposed overnight at 4°C to anti-human type I collagen antibodies (MP Biomedicals) diluted 1:200, anti-human type II collagen antibodies (MP Biomedicals) diluted 1:500, or to mouse anti-type X collagen IgG. 25 The antibodies against collagen type I, II, and X allow to detect both human and porcine antigens. After incubation with a biotinylated donkey anti-mouse secondary antibody (Dianova), a complex of streptavidine and biotinylated alkaline phosphatase (Vectastain, ABC-AP; Vector Laboratories) was added. The sections were developed with fast red and counterstained with hematoxylin.
The morphological assessment of the different approaches was performed according to the International Cartilage Repair Society (ICRS) Visual Histological Assessment Scale, including analysis of surface, matrix, cell distribution, cell population viability, subchondral bone, and cartilage mineralization. 26 Representative central sections of the defects were analyzed independently by three experts, and the respective values were averaged.
Morphological assessment and quantification of osseous overgrowth
For quantification of excessive bone formation, the relative volume of calcified/bone tissue within the cartilage repair tissue above the virtual line of the former subchondral bone plate was determined by point-counting histomorphometry using a grid in a modified method described by O'Driscoll et al. 27 To determine the percentage of excessive osseous tissue within the repair tissues, five parallel Alizarin red-stained sections with a separation distance of 1 mm were captured using a Leica microscope camera throughout each of the treated lesions and further analyzed at a magnification of 100×by a digital imaging program as described previously. 21 Briefly, a virtual transverse line was created spanning between the intercept points of the cartilage-bone interface of the adjacent intact cartilage and the defect borders. Calcified/bone tissue projecting above this line was distinguished from noncalcified repair tissue and marked by pen-tracing. An overlaid grid was used to count an average of 80 grid boxes per section. A grid box was assigned to noncalcified tissue if more than 50% of the area of the grid box was noncalcified tissue. The relative volume fractions of calcified/osseous tissue in relation to the total area of the repair tissue above the correct level of the original subchondral bone plate were averaged through all sections for each respective lesion and treatment groups.
Tube formation assay
A soluble basement membrane matrix (Geltrex; Invitrogen) was applied into individual wells of 12-well plates and allowed to polymerize at 37°C for at least 30 min followed by plating of 2×104 human dermal microvascular endothelial cells (HDMECs; PromoCell) per well. In a portion of the wells, the culture medium (DMEM-F12, 10% FCS, 50 ng/mL VEGF) was supplemented with 1, 5, or 10 μg/mL rhTSP-1 (R&D Systems), and/or 100 ng/mL OP-1 (R&D Systems), respectively. The endothelial cells were incubated for 48 h to allow the formation of tube-like structures and the extent of tube formation in 10 random fields per well was captured by a digital camera.
Tube formation was quantified as described previously 21 by a modified method according to Sanz et al. 28 The angiogenic index (AI) was determined for each field: (AI=total length of connected tubes/surface of analysis). Representative samples of three independent experiments are shown.
Gene expression analysis in vitro
For quantitative gene expression studies in vitro, human cells were used due to the lack of sequences of the porcine genome. Cells were isolated from macroscopically intact cartilage from the dorsal femoral condyles from 5 patients undergoing total knee replacement at the division of Orthopaedic Rheumatology at the University of Erlangen-Nuernberg and cultured as described previously. 29 The institutional ethics committee approved the study protocol. The cells were grown for 5 days in monolayer in DMEM containing 10% FCS. Subsequently, a portion of the cells was further stimulated by 5 μg/mL TSP-1 and/or 100 ng/mL OP-1 for 6, 24, or 48 h, respectively. Total RNA was isolated from cells for quantitative reverse transcription-polymerase chain reaction (RT-PCR) as described previously. 21 Two potential target genes of TSP-1 involving the cell cycle (p21cip/waf) and chondrocyte hypertrophy (GADD45β) were chosen according to data from the literature or according to data obtained from our own unpublished cDNA array analyses. Further, the mRNA expression of Sox9, a key chondrogenic transcription factor, was determined. The expression levels were quantified by real-time RT-PCR using the ABI Prism 7900 sequence detection system (Applied Biosystems) and Verso One-Step QRT-PCR Rox Kit (Abgene). For each sample, the relative amount of the target mRNA was determined and normalized to human β2m. The following primer and probe sets were used: β2m (forward: 5′-TGACTTTGTCACAGCCCAAGATA-3′, reverse: 5′-AATC CAAATGCGGCATCTTC-3′, FAM/TAMRA probe: 5′-TGAT GCTGCTTACATGTCTCGATCCCA-3′); p21cip1/waf1 (forward: 5′-CAGAGGAAGACCATGTGGAC-3′, reverse: 5′-TTCAGC CTGCTCCCCTGA-3′, FAM/TAMRA probe: 5′-GTCACTG TCTTGTACCCTTGTGCCT-3′); GADD45β (5′-ATTGACGA GGAGGAGGAGGA-3′, reverse: 5′-CGTTGTCACAGCAGA AGGAC-3′, FAM/TAMRA probe: 5′-ATCGCCCTGCAAA TCCACTTCACG-3′). The primer and probe sets for Sox9 were purchased from Applied Biosystems.
Statistical analysis
All data are presented as mean±standard deviation. For the evaluation of the ICRS score nonparametric Kruskal–Wallis tests followed by a post hoc Dunn multiple comparison test were used to determine treatment-specific differences. Excessive bone formation and tube formation were assessed by analysis of variance followed by Tukey–Kramer tests. Quantitative gene expression was analyzed using Student's two-sided t-test. All statistical results were considered significant for p-values of <0.05.
Results
Microfracture treatment (MFX) forms fibrocartilage and leads to excessive endochondral ossification of the repair tissue
Treatment with MFX generated a repair tissue with a typical zone-dependent cellular differentiation pattern. In superficial areas, the ingrowing cells failed to undergo chondrogenesis and only formed fibrous tissue or fibrocartilage with a low content of proteoglycans both at 6 and 26 weeks (Figs. 1a, 2a, 3a, and 4). Fibrocartilage was characterized by positive staining for type I collagen (Figs. 1e and 2e), but only weak staining for the cartilage-specific type II collagen at 6 weeks (Fig. 1i). At 26 weeks, spatial differences within the MFX samples could be observed for type II collagen with no or only weak staining in the superficial zone and distinct staining within the deep zone. In the deepest zone, a layer of cells underwent terminal differentiation characterized by a hypertrophic cellular phenotype (Fig. 3a), which was ascertained using type X collagen assessment (Figs. 1m and 3a). Chondrocyte hypertrophy preceded endochondral ossification that advanced from the deepest zones at 6 weeks toward the upper zones within the following 26 weeks (Fig. 2m). While adjacent healthy cartilage was rich in TSP-1, staining for TSP-1 was less widely distributed within the repair tissue induced by MFX at 6 weeks (Fig. 1q). At 26 weeks, the intensity and distribution of TSP-1 staining within the repair tissue was comparable to the other groups (Fig. 2q).

Repair tissue at 6 weeks after microfracturing (MFX) and supplementation with TSP-1 and/or osteogenic protein-1 (OP-1). The resulting repair tissues were examined by toluidine blue staining and by immunohistochemistry for type I (Col 1), type II (Col 2), and type X collagen (Col 10), or thrombospondin-1 (TSP-1). Microfracture treatment (MFX)
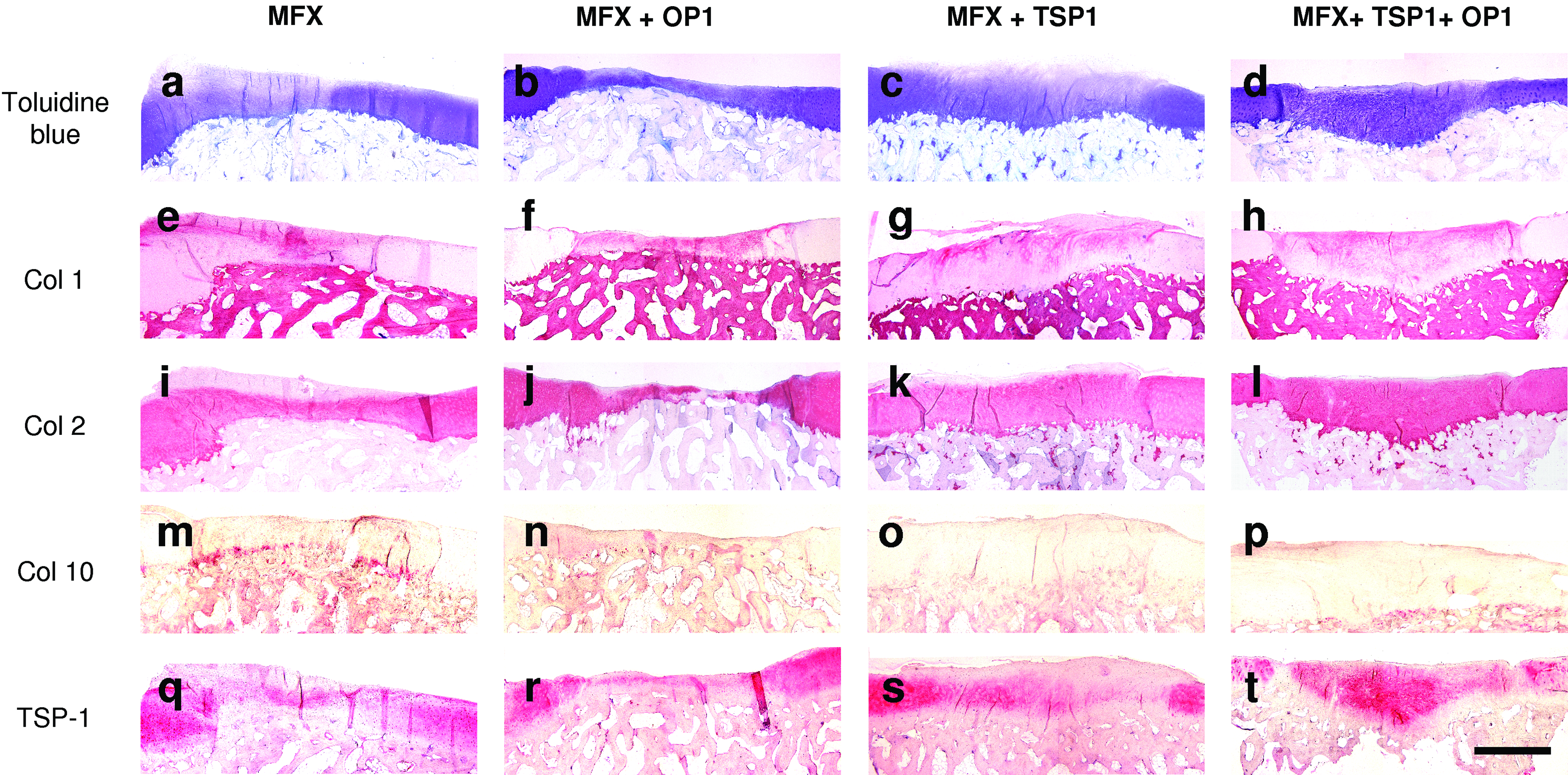
Repair tissue at 26 weeks after microfracturing (MFX) and supplementation with TSP-1 and/or OP-1. The resulting repair tissues were examined by toluidine blue staining and by immunohistochemistry for type I (Col 1), type II (Col 2), and type X collagen (Col 10), or TSP-1. Microfracture treatment (MFX)

High magnifications of superficial and deep zones of the repair tissues at 6 weeks

Overview panel of representative histological sections of the repair tissues of all animals treated (No. 1–6) at 26 weeks. Toluidine blue staining and immunohistochemistry for type II collagen of representative sections. Arrowheads indicate the borders of the defects. Scale bar=1 mm. Color images available online at www.liebertonline.com/tea
OP-1 promotes chondrogenesis but also induces endochondral ossification
The application of OP-1 into microfracture holes strongly induced chondrogenic differentiation of the ingrowing mesenchymal cells within the initial 6 weeks with formation of a proteoglycan-rich matrix that partially stained positive for type II collagen (Fig. 1b, j). However, at 6 weeks, the matrix was still heterogeneously structured with positive staining for type I collagen (Fig. 1f). Strikingly, a considerable portion of the cells underwent terminal chondrocyte differentiation and adopted a hypertrophic phenotype characterized by pericellular staining for type X collagen (Fig. 3a). Interestingly, the spatial pattern for type X collagen immunostaining was apparently completely inverse to that of TSP-1 (Figs. 1n, r and 3b). At 26 weeks, hypertrophic repair cartilage was replaced by osseous tissue which considerably exceeded the original level of the subchondral bone plate (Figs. 2b and 4). The overlying superficial layer of cartilaginous repair tissue stained positive for both type I and type II collagen (Fig. 2f, j). Type X collagen could no longer be detected at 26 weeks indicating a completion of the ossification process (Fig. 2n).
Treatment by TSP-1 inhibits chondrocyte hypertrophy and endochondral ossification
The application of recombinant TSP-1 into the microfracture holes failed to induce chondrogenesis. At 6 weeks, only fibrous tissue was formed (Fig. 1c) with strong staining for type I collagen but no relevant staining for type II collagen (Fig. 1g, k). Terminal differentiation of the repair cells into a hypertrophic phenotype was nearly complete absent in the central regions of the defect, and only the border zones showed some positive staining for type X collagen (Fig. 1o). At 6 weeks, a more pronounced staining for TSP-1 within the repair tissue could be observed in TSP-1-treated defects compared to lesions that did not receive TSP-1 (Fig. 1s). Although the immunostaining does not allow to distinguish between the applied recombinant human TSP-1 and endogenous porcine TSP-1, the differences between the treatment groups detected at 6 weeks imply remainders of the exogeneously applied TSP-1. At 26 weeks, the intensity of TSP-1 staining declined with a more even distribution that was comparable to defects that did not receive TSP-1, an effect that can be ascribed to diffusion or degradation of exogeneously applied TSP-1 (Fig. 2s). The reconstitution of the subchondral bone plate was severely retarded by treatment with recombinant TSP-1. Within 26 weeks, the cells in deeper zones showed partial differentiation into a fibrocartilaginous phenotype with a mixed staining for both type I and type II collagen (Figs. 2c, g, k, and 4). Chondrocyte hypertrophy with synthesis of type X collagen and excessive endochondral ossification was completely absent in this treatment group (Figs. 2o and 3a).
Combined treatment with TSP-1 and OP-1 supports chondrogenesis but prevents terminal chondrocyte differentiation
The combined treatment with TSP-1 and OP-1 significantly stimulated chondrogenesis and generated hyaline-like repair tissue within 6 weeks characterized by strong staining for toluidine blue (Fig. 1d). Staining for type I collagen could be observed at 6 weeks (Fig. 1h) and clearly decreased at 26 weeks (Fig. 2h). Instead, a clear predominance of the cartilage-specific type II collagen throughout the entire depth of the repair tissue could be observed at 26 weeks (Figs. 2l and 4). The repair tissue strongly stained for TSP-1 at both time points (Figs. 1t and 2t). In this treatment group, staining for type X collagen was low and confined to the deepest single cell layer at 6 weeks (Figs. 1p and 3a). Chondrocyte hypertrophy and excessive endochondral ossification was nearly completely absent at 26 weeks (Fig. 2p).
Quantification of excessive osseous tissue within the defect areas
Alizarin red staining served to observe excessive bone outgrowths within the former cartilage defects. In this animal model, the plain microfracture treatment (MFX) was associated with significant outgrowths of subchondral bone even without any additional growth factor treatment. The extent of ossification increased within the observation period and finally amounted to 20.0%±11.1% in relation to the total repair tissue volume at 26 weeks (Fig. 5a, b). Excessive ossification was even more prominent in defects treated additionally by OP-1, in which the percentage of excessive calcified tissue in relation to the total repair tissue volume increased from 7.3%±7.8% at 6 weeks to 26.2%±21.3% at 26 weeks (Fig. 5a, b). In contrast, treatment with TSP-1 completely prevented the formation of osseous outgrowths of subchondral bone (Fig. 5a, b). TSP-1 also prevented excessive ossification when applied simultaneously with OP-1; no outgrowths of subchondral bone could be observed in any of the specimen treated by MFX+TSP-1+OP-1 (Fig. 5a, b). Instead, the opposite effect could be observed in one defect (No. 4 in Fig. 4) treated by MFX+TSP-1+OP-1 with cartilaginous repair tissue persisting in deep zones below the level of the original subchondral bone plate (Figs. 2d, h, l, p, t; 4 No. 4; and 5a).
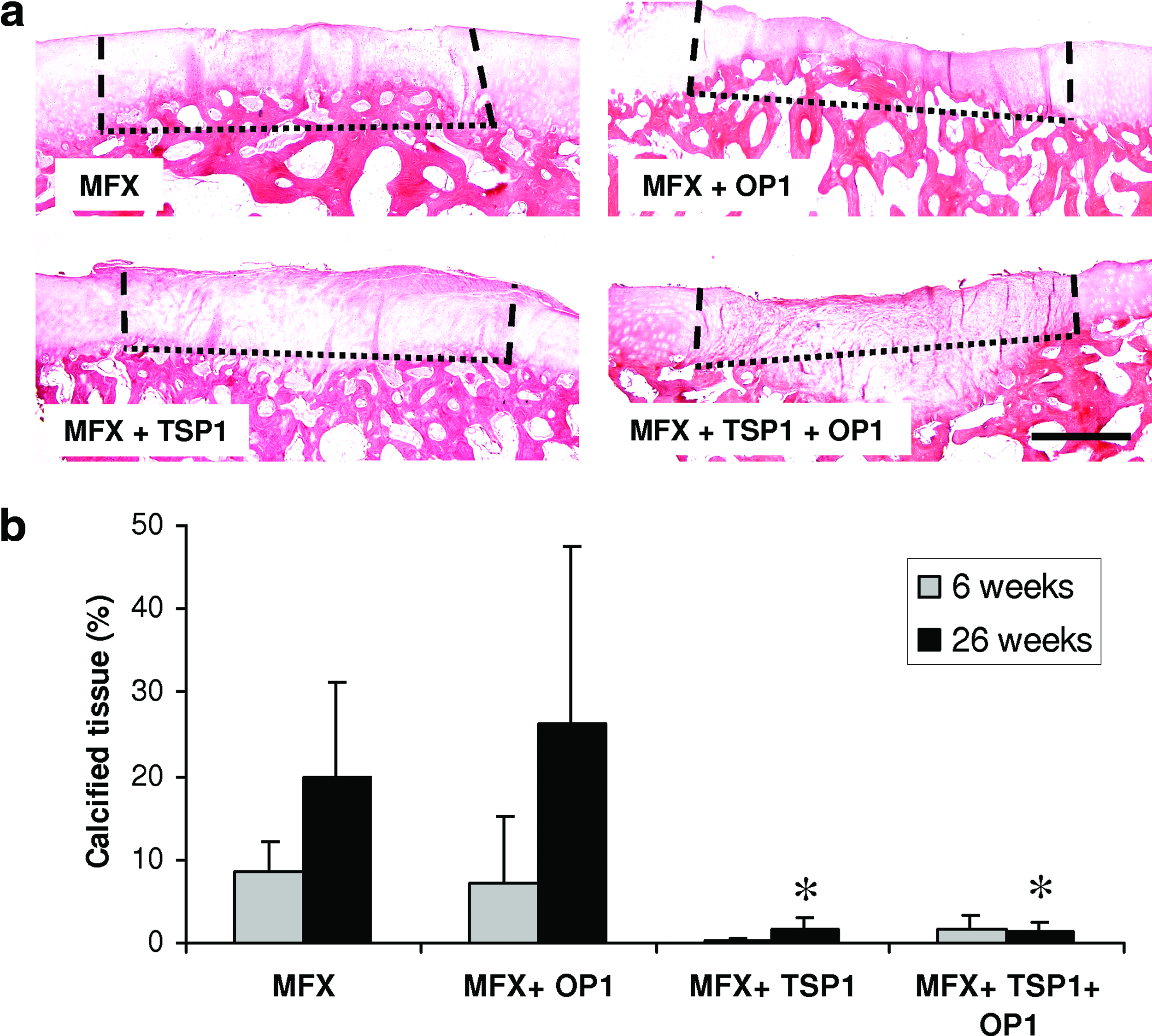
Evaluation of matrix calcification and excessive ossification at 26 weeks: Staining for calcified tissue in lesions treated solely by microfracture (MFX), or microfractured lesions supplemented with OP-1, TSP-1, or a combination of TSP-1 and OP-1
Comparison of the repair tissues of the different treatment groups
The ICRS visual histological assessment scale (Table 1) was used for statistical analysis of the morphological data. Figure 4 shows an overview of representative sections of all repair tissues. The different treatment schemes had no influence on the surface score. An apparent increase of the surface score values from 6 to 26 weeks in all groups did not reach the level of significance. The matrix score was significantly improved by the combined treatment with TSP-1 and OP-1 compared to MFX alone. A physiological remodeling of the matrix with a zone-specific cell distribution pattern was not achieved in any of the different treatment groups within the observation period. The cell population viability within the repair tissue was not affected by the different treatment schemes and relevant signs of apoptosis or necrosis could not be observed. Treatment by MFX or MFX+TSP-1 was associated with poor score values for the subchondral bone at 6 weeks. However, in these two treatment groups, the subchondral bone recovered with significantly improved score values within 26 weeks. Cartilage mineralization was significantly prevented in lesions containing recombinant TSP-1 (MFX+TSP-1 and MFX+TSP-1+OP-1) compared to MFX- and MFX+OP-1-treated lesions.
Data were assessed according to the ICRS visual histological assessment scale and are shown as the mean value±standard deviation. Different indices (comparison between different treatment groups:
MFX, microfracture treatment; OP-1, osteogenic protein-1; TSP-1, thrombospondin-1.
Vascular status of repair tissues
Immunostaining for the endothelial cell marker CD31 was performed to evaluate if excessive endochondral ossification was associated with increased angiogenic activity and vascular invasion. In general, CD31-positive vessel structures could be detected in subchondral bone tissue, but not in healthy articular cartilage. MFX and MFX+OP-1-treated lesions sparsely showed some CD31-positive vessel structures within the repair tissue, whereas CD31-positive vessel structures were completely absent in the repair tissue treated by MFX+TSP-1 or MFX+TSP-1+OP-1 (Fig. 3a). The vascular status in subchondral bone spaces was influenced by neither TSP-1 nor OP-1. We could not observe any relevant invasion of CD31-positive vessel structures into the deep zones of MFX- or MFX+OP-1 treated defects. Thus, chondrocyte hypertrophy was not immediately spatially associated with vascular invasion, since vessels were rather located beneath the zone of hypertrophic chondrocytes in already calcified and ossified tissue.
Potential biological mechanisms of TSP-1
Since TSP-1 is known to be a potent anti-angiogenic protein, its biological effects were investigated by an angiogenesis assay in comparison to OP-1. HDMECs typically form tube-like structures when cultivated on a basement membrane matrix (Fig. 6a). Instead, tube formation was significantly disturbed and the AI was significantly reduced when the culture medium was supplemented with TSP-1 at a concentration of 5 μg/mL or higher (Fig. 6a, b). However, single or additional supplementation with 100 ng/mL OP-1 did not affect tube formation of HDMECs in comparison to the control or TSP-1-treated samples (Fig. 6a, b).
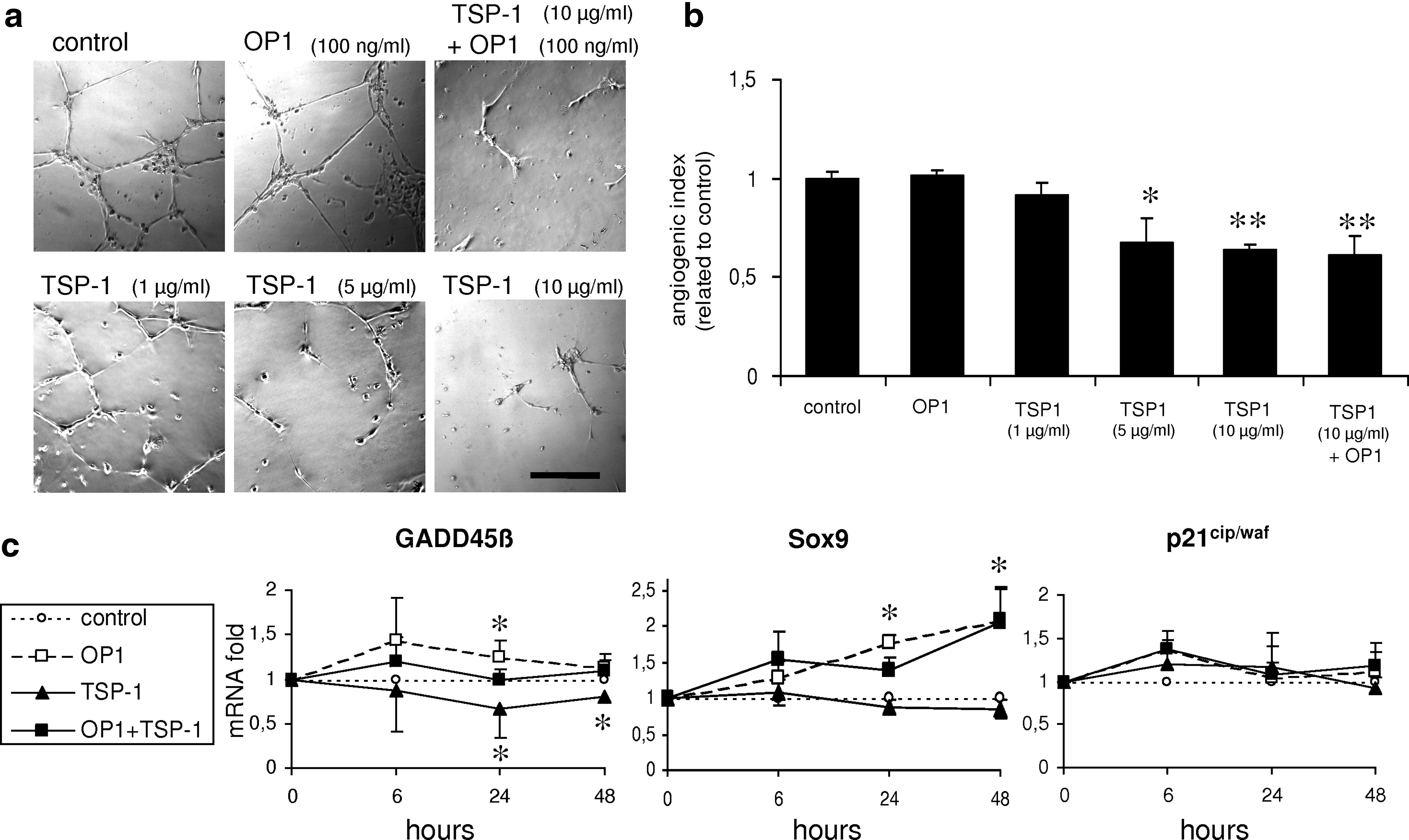
The effect of TSP-1 on the angiogenic activity of human dermal microvascular endothelial cells (HDMECs) was assessed by the tube formation assay. Supplementation with 100 ng/mL OP-1 did not affect the angiogenic activity of HDMECs, whereas the formation of tube-like structures was strongly inhibited by 5 and 10 μg/mL TSP-1
To investigate other potential molecular mechanisms of TSP-1 that may directly mediate its anti-osteogenic and anti-hypertrophic effects on a chondrogenic cell population, the expression of potential target genes, including GADD45β, Sox9, and p21cip/waf, was detected by real-time RT-PCR (Fig. 6c). Stimulation with TSP-1 for 24 or 48 h significantly inhibited the mRNA expression of GADD45β, a factor that was recently shown to be involved in chondrocyte hypertrophy.30,31 On the contrary, single stimulation with OP-1 for 24 h significantly induced the expression of GADD45β, an effect that could be neutralized by simultaneous application of TSP-1. The expression of the chondrogenic key transcription factor Sox9 was significantly increased by OP-1, but was not influenced by TSP-1. In contrast to a previous study using endothelial cells, 32 the expression of the cell cycle regulator p21cip/waf was not significantly increased by TSP-1 in our experiments based on chondrocytes.
Discussion
The present work demonstrated that OP-1 does not only induce chondrogenic differentiation of the invading precursor cells in microfractured lesions, but also strongly promotes terminal chondrocyte differentiation.33,34 Thus, OP-1 exerts both pro-chondrogenic and pro-osteogenic effects, which are, at least in part, mediated by increased expression of the key chondrogenic transcription factor Sox9 on the one hand, 35 and by increased expression of the osteogenic transcription factor Runx2 on the other hand. 36 Further, we could demonstrate that OP-1 induces the expression of GADD45β, a factor that is involved in terminal chondrocyte differentiation.30,31 In this study, these properties of OP-1 were reflected by abundant chondrogenesis in MFX+OP-1-treated defects, but simultaneously a large portion of the repair cells adopted a hypertrophic cellular phenotype, which was underlined by a matrix positive for type X collagen. Thus, OP-1 potentiated the problem of chondrocyte hypertrophy and excessive bone formation that is even spontaneously observed within microfractured lesions without any additional growth factor treatment. Osseous outgrowths of the subchondral bone plate may severely influence biomechanical properties of cartilage repair tissue. Abnormal high indentation stiffness values had been measured for cartilage repair tissues with such osseous outgrowths, which may affect the long-term outcome of the repair procedure. 3
Endochondral ossification is a complex biological process involving a multitude of signaling molecules as well as biophysical and biochemical factors, and that proceeds through several consecutive steps. The transition from nonhypertrophic to hypertrophic cartilage is based on terminal chondrocyte differentiation that is primarily driven and controlled by a network of specific signaling molecules. 30 The subsequent step from hypertrophic cartilage to bone tissue further depends on proteolytic matrix degradation followed by vascular invasion and ingrowth of osteoblasts. 37
The present study demonstrated that excessive bone formation, particuarly within OP-1 treated lesions, is based on endochondral ossification preceded by a zone of hypertrophic chondrocytes. The application of recombinant TSP-1 into microfractured cartilage lesions could significantly inhibit spontaneous or OP-1-induced chondrocyte hypertrophy. The anti-osteogenic effects of TSP-1 are in agreement with data from knockout mice, in which the deletion of TSP-1 or the related TSP-2 led to increased ossification in fracture callus 38 and premature closure of the growth plate. 39
The biological mechanisms by which TSP-1 exerts the observed anti-osteogenic effects are largely unknown and may be based on complex mechanisms since the structure of TSP-1 with multiple domains may involve different pathways. 40 In principle, TSP-1 might interact with three different targets: the surrounding endothelial cells, the chondrogenic cell population, and the extracellular matrix.
Concerning endothelial cells, TSP-1 was shown to exert strong anti-angiogenic effects that can be ascribed to the interaction with CD36, which mediates proapoptotic effects, 41 or to the interference with VEGF signaling. 42 Indeed, we could demonstrate that recombinant TSP-1 inhibited tube formation of endothelial cells in vitro, and the application of TSP-1 also prevented vascular ingrowth into the repair tissue in vivo. In other animal models, the intra-articular overexpression of TSP-1 prevented erosive arthritis or suppressed the progression of osteoarthritis by a reduction of synovial hypertrophy and a reduced hypervascularity of the synovial membrane.23,43 Thus, avascularity and the resulting lower oxygen levels may indirectly stabilize the chondrocyte phenotype since hypoxia is known to support the chondrocyte phenotype.20,44 However, these putative hypoxia-related stabilizing effects on the chondrocyte phenotype were not sufficient to induce chondrogenic differentiation of invading mesenchymal stem cells.
Thus, it can be concluded that TSP-1 also directly targets the chondrogenic cell population within the repair tissue. Indeed, chondrocytes express a number of TSP-1-interacting receptor molecules, including CD36. 45 Binding to CD36 was shown to trigger anti-proliferative activities due to inhibition of G1/S phase transition mediated by accumulation of the cell cycle inhibitor p21cip/waf and Rb dephosphorylation. 46 Indeed, TSP-1 increased the protein levels of p21cip/waf in endothelial cells in a previous study. 32 In our experiments, however, the increase of p21cip/waf mRNA expression by TSP-1 was not significant in chondrocytes. In this respect, endothelial cells may differ from differentiated chondrocytes, since post-mitotic or slowly dividing differentiated chondrocytes are characterized by already high endogenous activity of p21cip/waf47 and, thus, further stimulation may not necessarily lead to significantly increased levels of mRNA expression. In general, a consecutive cell cycle arrest is considered to play an important role in maintaining a stable postmitotic chondrocyte phenotype.47,48
Further, TSP-1 significantly prevented chondrocyte hypertrophy and strongly inhibited matrix mineralization, which has also been described in another study on osteogenic progenitor cells. 49 These antihypertrophic effects of TSP-1 may be mediated by downregulation of GADD45β. This factor plays an essential role during terminal chondrocyte differentiation by inducing MMP13 and type X collagen expression. 30 GADD45β was shown to be an early response gene of BMP-2, 31 and the present study also revealed a significant response to the closely related factor OP-1 (BMP-7). On the contrary, TSP-1 was able to counteract the BMP-/OP-1-induced induction of GADD45β, and thus, TSP-1 may functionally antagonize terminal chondrocyte differentiation that is induced by BMPs.
In addition, as mentioned above, TSP-1 might functionally act on an extracellular level. TSP-1 could inhibit the activation of the preform of MMP9 and reduce the release of bioactivity of VEGF from extracellular stores. 50 TSP-1 was also shown to interact with the low density lipoprotein receptor-related protein (LRP1), which serves as a scavenger receptor to clear complexes of TSP-1 with MMP2, MMP9 or VEGF from the pericellular matrix.40,51 Thus, TSP-1 promotes the clearance or downregulation of growth factors and proteases that are essential for endochondral ossification.
The data of the in vitro tube formation assay demonstrated a dose- or threshold- dependent activity of TSP-1 with a threshold in the range between 1 and 5 μg/mL. This threshold effect could explain the observations in vivo, in which excessive ossification was observed in those defects that did not receive recombinant TSP-1 despite some faint immunostaining for TSP-1. In these cases, the faint positive staining for TSP-1 could be ascribed to endogenous TSP-1, which is known to be present at low levels in several repair tissues and mesenchymes. 52 However, such low endogenous levels of TSP-1, which are just detectable by highly-sensitive immunohistochemical analyses, may fail to reach the bioactive threshold and, thus, are insufficient to exert biological effects.
In conclusion, the inhibition of excessive ossification by TSP-1 may not be ascribed to one single mechanism, but may rather involve multimodal mechanisms addressing the vascular system, cell cycle, chondrocyte differentiation, interaction with other growth factors, and the clearance of matrix degrading enzymes. It is beyond the scope of this study to identify all these different potential mechanisms on a molecular basis, and further studies will have to identify the relevance of each respective different mechanism. Future studies will also have to include the use of high-resolution magnetic resonance imaging, which may allow to monitor the maturation of the repair tissues in vivo without the need for destructive histological analysis. Novel MRI sequences will provide not only morphological data, but also information on the compositional and biochemical properties of the repair tissues. 53
The data of this study suggest that both the induction of chondrogenesis and the prevention of terminal differentiation have essentially to be considered for cartilage repair strategies. It is obvious that the subsequent steps of the chondrocyte differentiation program cannot be controlled by a single therapeutic factor and a spectrum of different factors seems mandatory. The combination of OP-1 and TSP-1 was shown to be one suitable complementary “cocktail” of two different factors that allows both the induction and stabilization of the articular chondrocyte phenotype. In this context, however, it has to be considered that not only excessive bone formation but also persisting cartilaginous tissue below the original subchondral bone plate, as observed in one of the MFX+TSP-1+OP-1-treated defects, may represent a problem and may interfere with the outcome. Thus, for a future clinical use, further studies will have to complete and optimize the ingredients of such a therapeutic cocktail and will also have to address and optimize their respective doses and release kinetics.
Footnotes
Acknowledgments
We thank M. Pfluegner and H. Rohrmueller for excellent technical assistance. This work was supported by the Interdisciplinary Center of Clinical Research (IZKF) at the University Hospital Erlangen (grant A36) and the German Research Foundation (DFG) (grant GE 1975/2-1).
Disclosure Statement
No competing financial interests exist.