
Abstract
Dedifferentiation of cells to multipotential cells is of interest since they have a potential regenerative capacity. Our purpose was to de- and redifferentiate murine RAW 264.7 cells, a committed macrophage cell line of hematopoietic origin, into mesenchymal-like cells such as osteoblasts. RAW 264.7 cells in culture were treated with 5 μM reversine, a purine analog that was shown to dedifferentiate myoblasts in osteoblasts. Treatment with reversine resulted in a significant increase in the expression of the STRO-1 antigen, a marker of mesenchymal stem/progenitor cells: from 0.6%±0.5% cells in untreated RAW cells to 19.0%±8.6% in treated cells, but there was no increase in the expression of SH-2 (CD105), an earlier marker of mesenchymal stem cells. The effects of reversine were significantly curtailed by 67% when cultures were pretreated with the c-Jun N-terminal kinase pathway blocker SP600125. These STRO-1+ cells retained a multipotential status and were capable of redifferentiating into cells with osteogenic and lipogenic characteristics under inductive conditions. We showed that STRO-1+ cells in an osteogenic medium significantly increased expression of the osteoblast marker osteocalcin, and formed mineralized nodules. When seeded on a demineralized scaffold of human bone in vitro, these cells deposited a calcium matrix. Under adipogenic conditions, expression of the adipocyte marker peroxisome proliferator-activated receptor gamma 2 on STRO-1+ cells was elevated, and cultures stained positive with Oil red O. Our results demonstrated that treating a committed hematopoietic cell line with a purine analog can alter cell development and result in cellular reverse transformation into stage-limited multipotential cells. These cells could subsequently be redifferentiated into cells with characteristics of the mesenchymal lineage, such as those of an osteoblast and/or adipocyte, under inductive conditions.
Introduction
Evidence suggests that dedifferentiation of human cells may be possible.11–15 Given that protein kinase pathways may be involved in such processes, libraries of heterocyclic compounds that have been designed around kinase-directed scaffolds 16 have been screened for effective dedifferentiation molecules. These compounds have included substituted purines of which a 2,6-disubstituted purine analogue, 2-(4-morpholinoanilino)-6-cyclohexylaminopurine (reversine), has been shown to be at least partially effective in dedifferentiating myoblasts, 17 fibroblasts, 18 and neurons. 19 Chen et al. 17 used reversine to treat murine C2C12 myoblasts that otherwise differentiate into multinucleated myotubes. The reversine-treated cells became multipotent cells and could be redirected to differentiate into osteoblasts or adipocytes using osteogenic or adipogenic conditions, respectively. 17
To confirm and further test the ability of reversine to direct dedifferentiation, we sought to determine if reversine could dedifferentiate a committed hematopoietic cell line into a multipotent progenitor that could then be redifferentiated into specific cell lineages of mesenchymal cells. For the purposes of our experiments, we used the murine RAW 264.7 peritoneal macrophage-like cell line derived from an Abelson leukemia virus–transformed cell line from BALB/c mice. Given the hematopoietic origin and the committed nature of this cell line, it was our purpose to dedifferentiate these cells into mesenchymal progenitor-like cells by treatment with reversine, determine the possible role of protein kinases in this process, and explore the multipotent status of such cells toward redifferentiation.
Materials and Methods
Cell cultures
RAW 264.7 (RAW) cells (ATCC) were cultured in the Dulbecco's modified Eagle's medium (DMEM; Invitrogen), supplemented with 10% fetal bovine serum (FBS, Sigma-Aldrich) and antibiotics that included 100 μg/mL penicillin G, 50 μg/mL gentamycin sulfate, and 0.3 μg/mL fungizone (complete medium). All experiments were carried out with generations of RAW 264.7 cells between passages 2 to 7. The culture medium was changed every 2 to 3 days, and cells were maintained at 37°C in a humidified atmosphere of 5% CO2 and 95% air.
For reversine (Sigma
Flow cytometry and determination of STRO-1 and SH-2
The STRO-1 and SH-2 (CD105) antigens were selected to define the presence of stage-specific mesenchymal progenitor cells.20,21 STRO-1 and SH-2 determination was assessed by a fluorescence-activated analysis (FACS). The cells (∼1–2.5×105) were released from culture plates using 0.25% trypsin and washed in phosphate-buffered saline (PBS) containing 2% FBS. Cells were then re-suspended in 0.2 mL of FBS-PBS with saturated concentrations (1:200 dilution) of the mouse monoclonal antibody anti-human STRO-1 (R&D Systems), which is trypsin resistant and cross-reacts with mouse bone marrow-derived stromal progenitors, and incubated for 1 h at 4°C. The cells were washed and labeled in 0.2 mL of FBS-PBS containing a secondary antibody (goat anti-mouse IgM, μ-chain specific, Cy5-conjugated, 1:100 dilution; Cedarlane Laboratories, ON, Canada), and incubated for 1 h at 4°C. After washing, the stained cells were analyzed by a FACStar Plus flow cytometer (Becton Dickinson FACS Systems) with the acquisition of at least 10,000 events per test. The following controls were used: omission of both antibodies, omission of the first antibody, and substitution of purified mouse myeloma IgM for the STRO-1 antibody.22,23 Each experiment was repeated at least three times. No reversine controls were analyzed in parallel with test samples at the specified time point (at day 1, in time course studies or after 4 days).
To confirm positive STRO-1 antigen results (and rule-out false-positive binding) by the flow cytometer machine, photographs of the florescence-labeled STRO-1 antibody on reversine-treated RAW cells were taken with a florescent microscope (LMD6000; Leica Inc.) at day 4 of culture for visual confirmation of the presence of the STRO-1 antigen.
We tested for SH-2 antigen, another early marker of mesenchymal cells, 20 with the PE-conjugated rat monoclonal antibody anti-mouse CD105 in an identical fashion to STRO-1, and repeated experiments five times.
Inhibition of reversine
To block the effects of reversine and to determine its pathway of action, three specific inhibitors were tested before reversine treatment. RAW cells (4×105 per 35-mm dish) were treated with PD98059 (inhibitor of ERK/MAPK) 30 μg/mL; SB203580 (p38 MAPK inhibitor) 30 μg/mL; or SP600125 (c-Jun N-terminal kinase [JNK] inhibitor II) 10 μg/mL. Each inhibitor was dissolved in DMSO, and DMSO alone was used as a control. Cells were maintained in an incubator with a humidified atmosphere of 5% CO2 and 95% air. One hour after treatment with one of the inhibitors, the medium was removed, and RAW cell cultures were washed once and then treated with 5 μM of reversine in fresh medium. Cells were maintained for another 4 days and then tested for STRO-1 by flow cytometry. Each inhibitor experiment was repeated three times.
Redifferentiation of STRO-1+ cells with osteogenic and lipogenic conditions
Four hundred STRO-1+ cells sorted by flow cytometry were seeded on 60-mm dishes in 5 mL DMEM complete medium. Cells were first cultured for 5 days for expansion, followed by the addition of an osteogenic or adipogenic inductive culture medium. The following controls were used in the experiments: reversine-treated cells in the absence of inductive culture media and untreated RAW cells in the presence of inductive culture media.
The osteogenic induction medium consisted of a complete medium containing dexamethasone (Dex) 10−8 M and 50 μg/mL ascorbic acid. 24 The medium was changed three times per week, and cells were maintained up to 21 days; 5 mM sodium β-glycerophosphate was added to the medium during the final four days of culture to initiate mineralization of bone nodules. Bone nodule formation was detected using von Kossa staining 25 as follows: the cells were rinsed three times with PBS, fixed with 4% paraformaldehyde/PBS solution, and then treated with 1 mL of silver nitrate per dish and incubated in the dark for 30 min. This was followed by adding 4 mL of carbonate formaldehyde per dish for several minutes and then air-dried. Bone nodules were determined under light microscopy, and only nodules >1 mm were counted. Alizarin red was also used for determination of calcium that was incorporated to the newly formed tissue. Cells were fixed with 70% ice-cold ethanol and rinsed with distilled water; dishes were stained at room temperature for 10 min in a stock solution of 2% alizarin red (Sigma-Aldrich) in distilled water. After washing and air-drying, bone nodules were evaluated under a light microscopy and counted under a dissecting microscope. Nodule formation experiments were repeated three times in independent experiments.
The adipogenic induction medium consisted of a complete medium containing 10−7 M Dex, 0.5 mM isobutyl-1-methyl xanthine, and 50 μM indomethacin (all from Sigma-Aldrich)26,27; STRO-1 cells were maintained in culture for an additional 7 days. The cultures were fixed in 4% paraformaldehyde/PBS solution for at least 15 min and stained with freshly made and filtered Oil red O solution with 0.5% (w/v) Oil red O (Sigma-Aldrich) in isopropanol diluted 6:4 in water at room temperature for 1 h to identify lipid-laden fat cells. The adipogenic culture experiments were repeated three times independently.
Semiquantitative reverse transcriptase–polymerase chain reaction for cell markers
Reverse transcriptase–polymerase chain reaction (RT-PCR) was used to identify markers for osteoclasts (tartrate-resistant acid phosphatase [TRAP] and cathepsin K), osteoblasts (runt-related transcription factor 2/core-binding factor alpha1 [Runx2] and osteocalcin [OCN]), and adipose cells (peroxisome proliferator-activated receptor gamma 2 [PPARγ2]) in untreated RAW cells or RAW cells treated with reversine for 4 days. A TRIzol reagent kit (Invitrogen) was used to isolate total cellular RNA at day 7 and 1, according to the manufacturer's instructions. Cells in culture were lysed in a phenol/guanidine isothiocyanate-containing TRIzol solution, followed by precipitation with isopropanol. Total RNA was resuspended in RNAse-free water and quantified by a spectrophotometer.
An aliquot of 1 μg of RNA was treated with DNAse (Invitrogen) for 15 min at room temperature. Total RNA was reverse transcribed using Moloney murine leukemia virus reverse transcriptase (200 units of SuperScript® II reverse transcriptase) plus 0.5 mM deoxy-NTPs, 500 ng oligo(deoxythymidine)18 primers, 5× 1st Strand Buffer, and 10 mM DTT (all from Invitrogen). The RT reaction mixture was incubated on a T Gradient or UNO II thermal cycler (Whatman Biometra) for 2 h at 42°C, followed by 75°C for 15 min to stop the reaction.
For PCR identification of specific markers, Taq DNA polymerase (recombinant, 5 U/μL; MBI Fermentas) was used in reactions containing 1 μL of cDNA from the RT reactions, 0.2 mM deoxy-NTPs, 200 μM each of forward and reverse primers, and 1.6 mM MgCl2. PCR was performed using a T Gradient or UNO II thermal cycler (Whatman Biometra). Oligonucleotide primers and conditions are listed in Table 1, and GAPDH was used as the internal control gene. Samples and a 100-bp ladder were electrophoresed on a 2% agarose gel in a Tris-borate ethylenediaminetetraacetic acid buffer containing 0.5 μg ethidium bromide/mL, and gels were photographed under UV light. PCRs were repeated at least three times, and relative mRNA levels were determined by computerized densitometry using GeneGenius Bioimaging System and SynGene Tools image analysis software (Synoptics Group). Results for each specific product were expressed as a relative mRNA level over GAPDH (giving a ratio of the gene product/control gene) and then expressed as a percent of the ratio obtained with the respective control in the same RT-PCR.
Scaffold preparation for re-mineralization of reversine-treated RAW cells
Sterile human trabecular bone samples (obtained with consent and ethics board approval from healthy patients, in which bone was harvested for surgical reconstruction) were thoroughly washed and hydrated for 1 h in PBS with antibiotics in a medium and cut into fragments with a diameter and thickness of 2–6 mm in the medium. The bone samples were then decalcified in a solution of 20% sodium citrate and 44% formic acid, which was changed every day. After 6 days, micro-CT (GE eXplore Locus SP specimen scanner; General Electric) imaging confirmed that the scaffolds were fully decalcified. The samples were then rinsed in 70% alcohol and sterile PBS sequentially, and stored in PBS at 4°C until cell seeding. 28 Scaffolds were seeded with 4×107 STRO-1+ cells per scaffold in 150 μL of the culture medium. This was performed by mixing the fragments with the cell suspension in sterile 200-μL tubes that were sealed with 0.22-μm filter membranes. Culture continued in six-well plates with 4 mL of osteogenic media with medium changes as above. 29 A bioreactor was not used. Sorted STRO-1− cells and RAW cells without reversine treatment were set up as negative controls. Since results with fetal rat calvaria cells (which contain osteoblasts and progenitors) are routinely used and are easily identifiable, they were used as a positive control for mineralization (in the absence of reversine). After 4 weeks of culture, scaffolds were assessed by micro-CT imaging weekly to determine if calcification was occurring; these scaffolds were maintained up to 6 weeks. Scanning parameters were set by the protocol as follows: 15-mm tube 8 μm 150 min; X-ray tube voltage 80 KV; X-ray tube current 80 μA; number of views 500; exposure time 3000 ms; detector bin mode 1×1; and effective pixel size 0.008 mm.
Statistical analysis
Statistical analysis was performed using GraphPad Prism v5.00 for Windows (GraphPad Software, www.graphpad.com) by t-test or analysis of variance one-way analysis with the Tukey post-hoc test. Results were considered statistically significant if the p-value ≤0.05.
Results
Three concentrations of reversine were initially tested on RAW cells: 2.5, 5, and 10 μM. The two lower doses had similar effects on increasing the number of STRO-1+ cells and on cell death, resulting in a death rate of 16%±4% for the 2.5-μm dose, and 13%±5% for the 5-μM dose as determined on the fourth day of treatment, but none thereafter. The 10-μM dose of reversine resulted in 82%±17% cell death. Therefore, we used 5 μm reversine for all subsequent experiments. The addition of 5 μM reversine to RAW cell cultures resulted in a dramatic rise in STRO-1+ cells: from 0.63%±0.47% cells in untreated RAW cells to 19.01%±8.64% in reversine-treated cells (p≤0.01; Fig. 1). There was no significant increase in the expression of SH-2 after reversine treatment (Fig. 2).
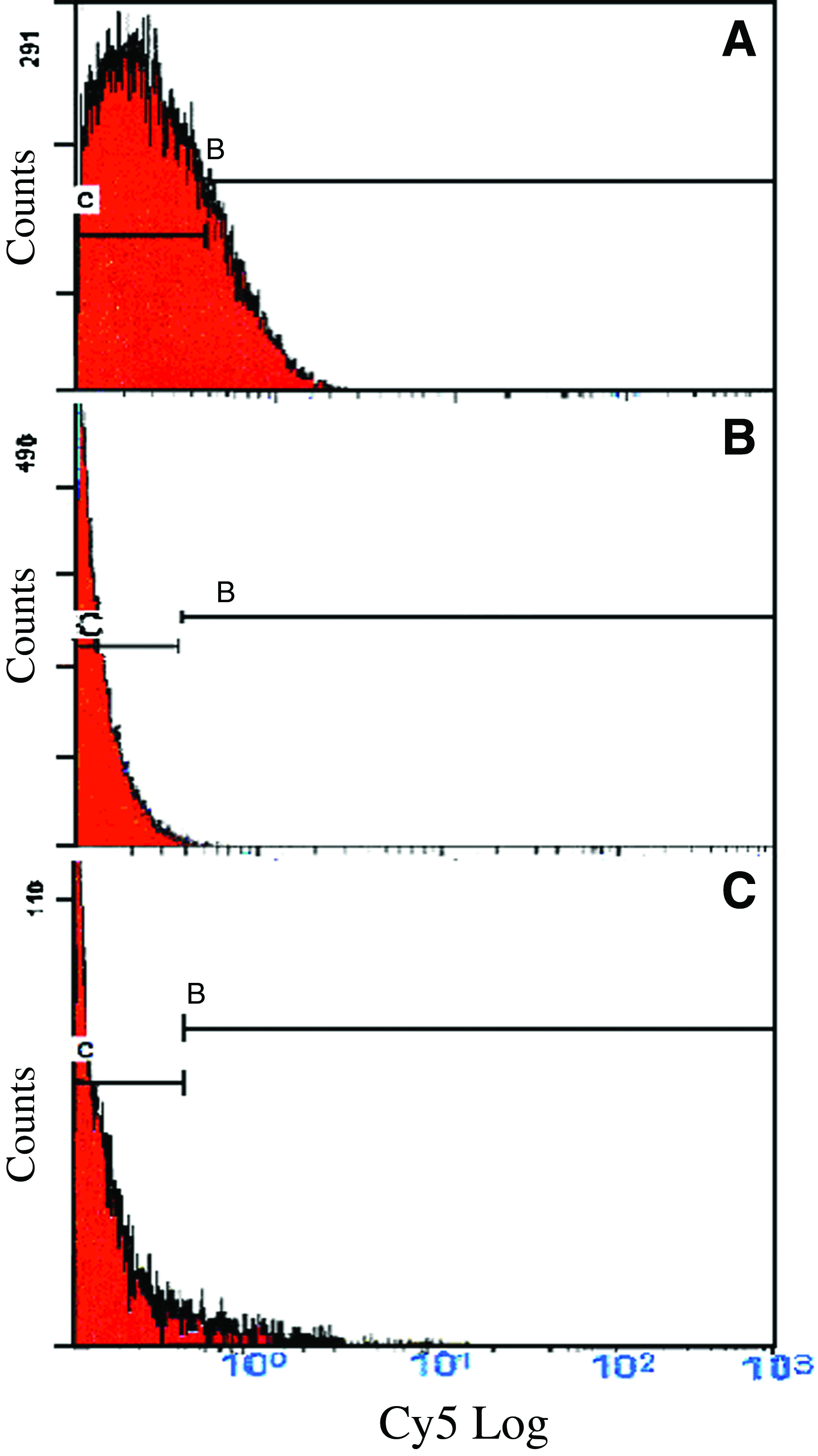
Histograms of the flow cytometric analysis of Cy5-labeled and sorted STRO-1+ RAW 264.7 cells treated with 5 μM reversine.
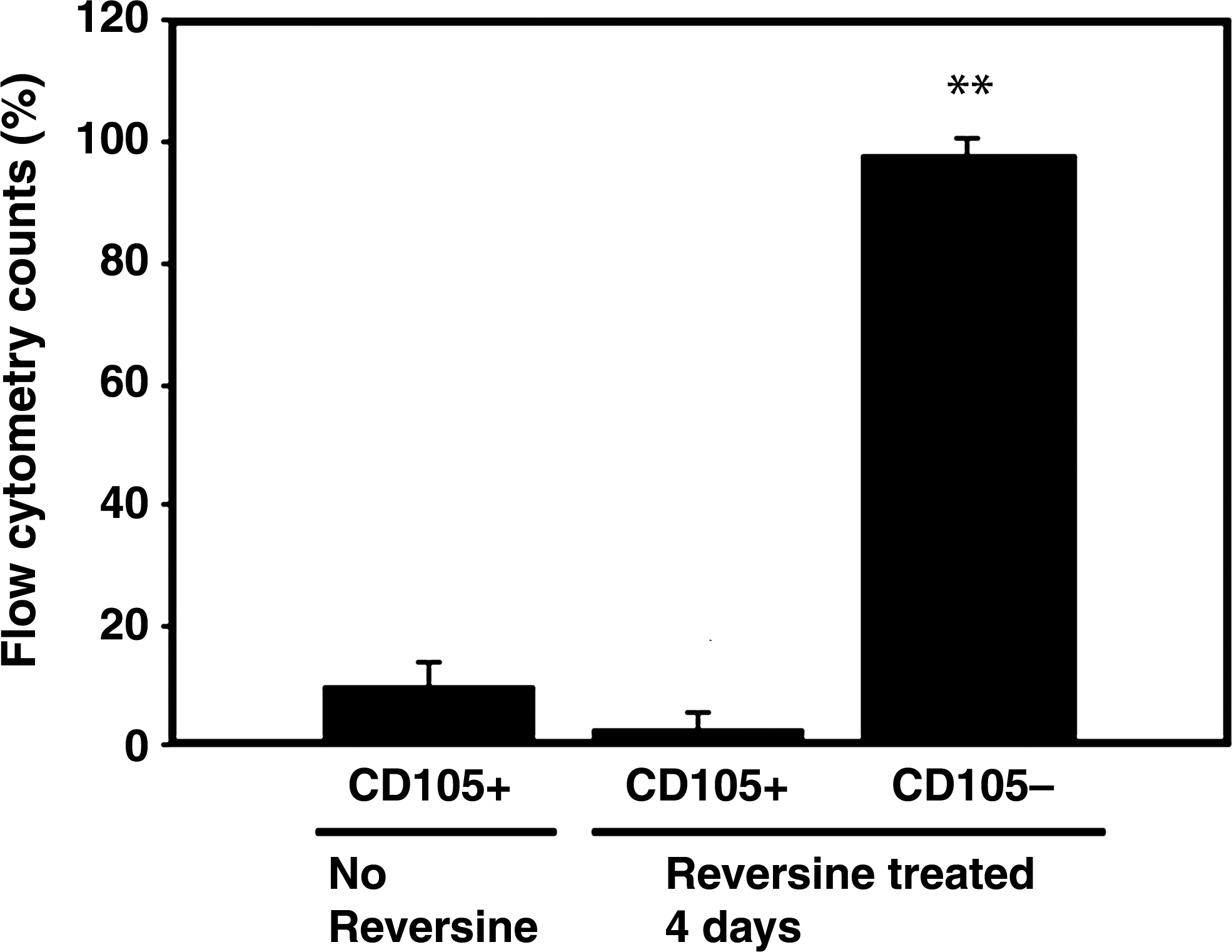
Flow cytometric analysis of SH-2 (CD105)-labeled RAW 264.7 cells after treatment with 5 μM reversine for 4 days. There was no specific increase in SH-2, which represents a progenitor stage that precedes the STRO-1+ stage. Data are from five independent experiments and expressed as flow cytometry counts (% total cells±SD). **p<0.01 versus no reversine control.
Reversine treatment resulted in visual changes in the cellular morphology. Untreated RAW 264.7 cells remained small, round, and uniform in culture (Fig. 3). After treatment with reversine, and over the course of 1 week, cells began to cluster and then formed colonies (Fig. 3). A small subgroup of cells increased in size, and some multinuclear cells were seen with an osteoclast-like appearance; these multinuclear cells did not colonize.
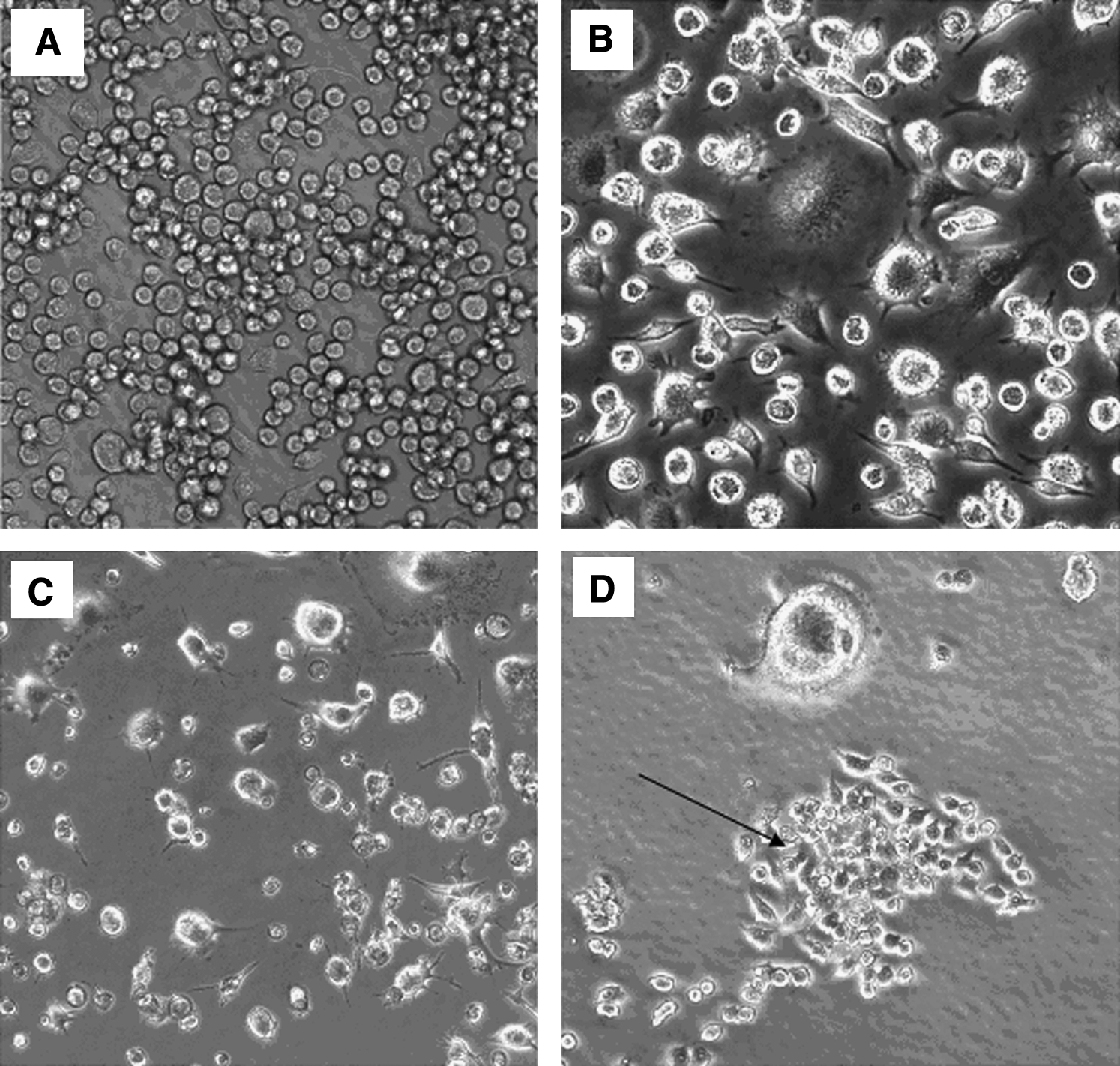
Micrographs of RAW 264.7 cells in culture after treatment with or without reversine.
Upon testing different durations of exposure to reversine, we found that an intermediate treatment had the maximal effect of increasing STRO-1+ cells (Fig. 4). The greatest effect was seen with 4 days of exposure during 7 days of culture, and this time point was therefore used for further experiments. The medium was not changed during the 4 days, or for shorter treatment durations. Longer treatment durations, which did include an additional medium change, resulted in fewer STRO-1+ cells (Fig. 4). STRO-1+ cells were confirmed by fluorescent microscopy of RAW cells cultured in chamber slides simultaneously with the flow cytometer analysis after 4 days treatment with 5 μM reversine; fluorescent images showed typical fluorescence marker positive on the outer surface membrane of the cells compared to that of the cells used as negative controls (Fig. 5).

Percentage of STRO-1+ cells after treatment of RAW 264.7 cells with reversine for different durations. Maximal effect was seen following exposure of cells to 5 μM reversine for 4 days (data from three independent experiments; STRO-1+ cells were determined by flow cytometry and presented as a percentage of total cells counted±SD). No reversine control=vehicle control at day 1. *p<0.05, **p<0.01 versus no reversine control; n=4.
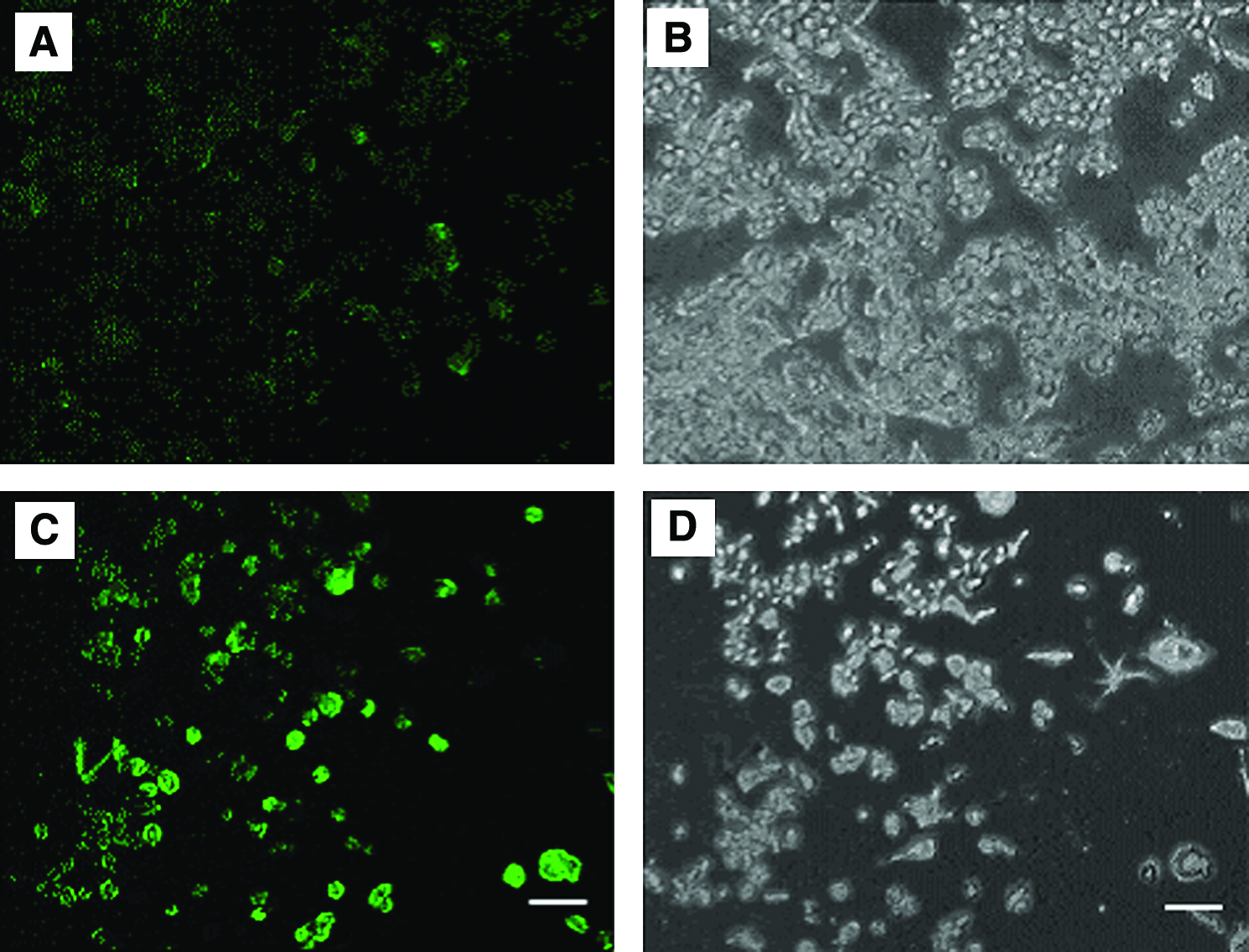
STRO-1+ cells were confirmed by fluorescent microscopy after treatment with reversine.
Each of the cell pathway blockers was added 1 h before reversine treatment and did not result in any appreciable changes in the cell morphology, nor did they result in cell death. The pretreatment of cells with the JNK pathway blocker SP600125 significantly curtailed the inductive effects of reversine by 67% from 19.8%±10.4% STRO-1+ cells (with reversine alone) to 6.5%±2.5% STRO-1+ cells (with blocker and reversine) (p<0.05; Fig. 6); however, the reversine effect was not completely blocked to the level of 0.82%±0.9% seen for untreated cells.
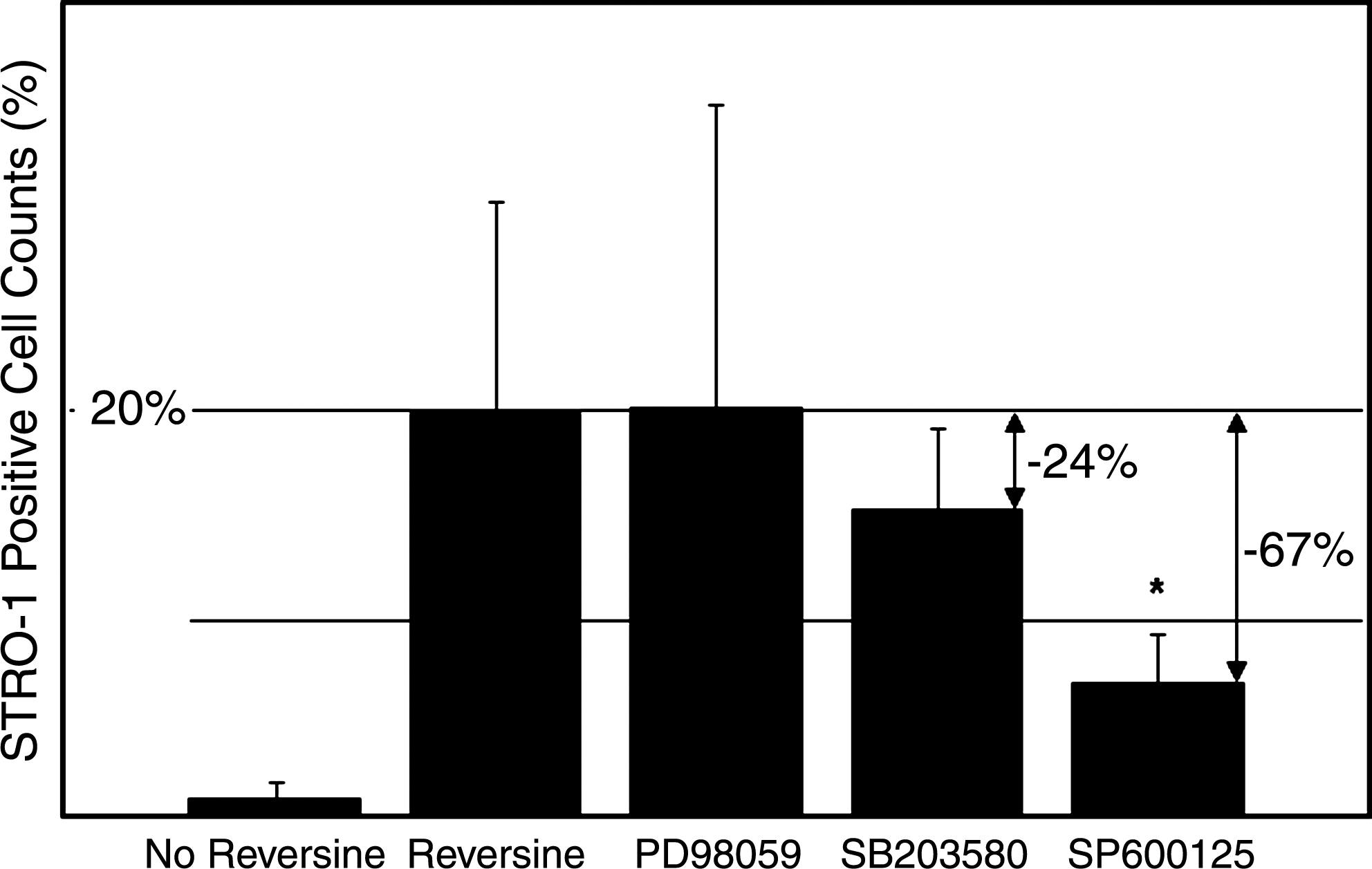
Pretreatment of RAW 264.7 cells with the JNK pathway inhibitor SP600125 resulted in a 67% drop (*p<0.05) in STRO-1+ cells after reversine treatment. Blocking the P38 pathway with SB203580 resulted in a 24% decrease in STRO-1+ cells (NS). Inhibiting the ERK pathway with PD 98059 had no effect on reducing the number of STRO-1+ cells. Data are expressed as % STRO-1+ cells±SD, n=3. JNK, c-Jun N-terminal kinase.
Blocking the P38 pathway with SB203580 resulted in a 24% decrease in STRO-1+ cells from 19.8%±10.4% (with reversine alone) to 15.0%±4.0% after the blocker and reversine (NS). Blocking the ER kinase pathway with PD 98059 did not result in any significant change in STRO-1+ cells (20.0%±15.0%) after reversine treatment (Fig. 6).
After flow cytometry to sort cells, subsequent cell cultures of STRO-1+ cells in the osteogenic medium resulted in mineralized bone nodules (von Kossa-positive or alizarin red-positive) compared to untreated cells (Fig. 7), as well as elevated expressions of osteoblast markers Runx2 (cbfa1) and osteocalcin, and mineralized nodules were detected in vitro. After reversine treatment, Runx2 RNA expression increased by 27% after treatment when compared to untreated cells in the osteogenic medium (NS, Fig. 8A). When maintained in an osteogenic medium, and tested at 7 and 21 days of culture, Runx2 expression was also elevated (23% and 39%, respectively) compared to untreated or RANKL-treated cells (NS). Expression of the specific osteoblast marker osteocalcin significantly increased by 1.63-fold at day 7 and 1.5-fold at day 21 of culture (both p<0.05) when compared to untreated cells (Fig. 8B).
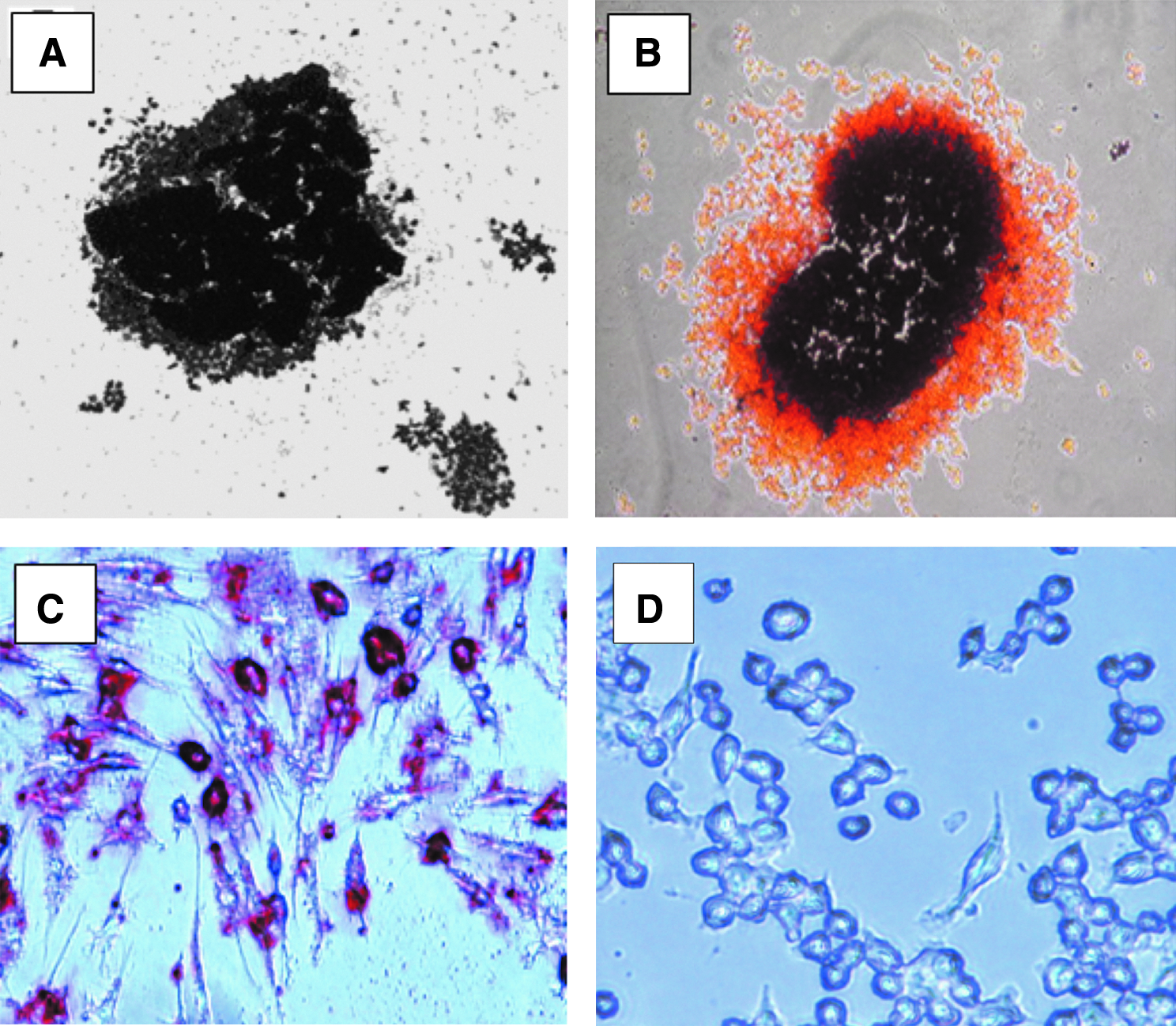
Micrographs of bone nodules and adipogenic cells from RAW 264.7 cells treated with reversine and maintained in an osteogenic or adipogenic medium for 21 days.
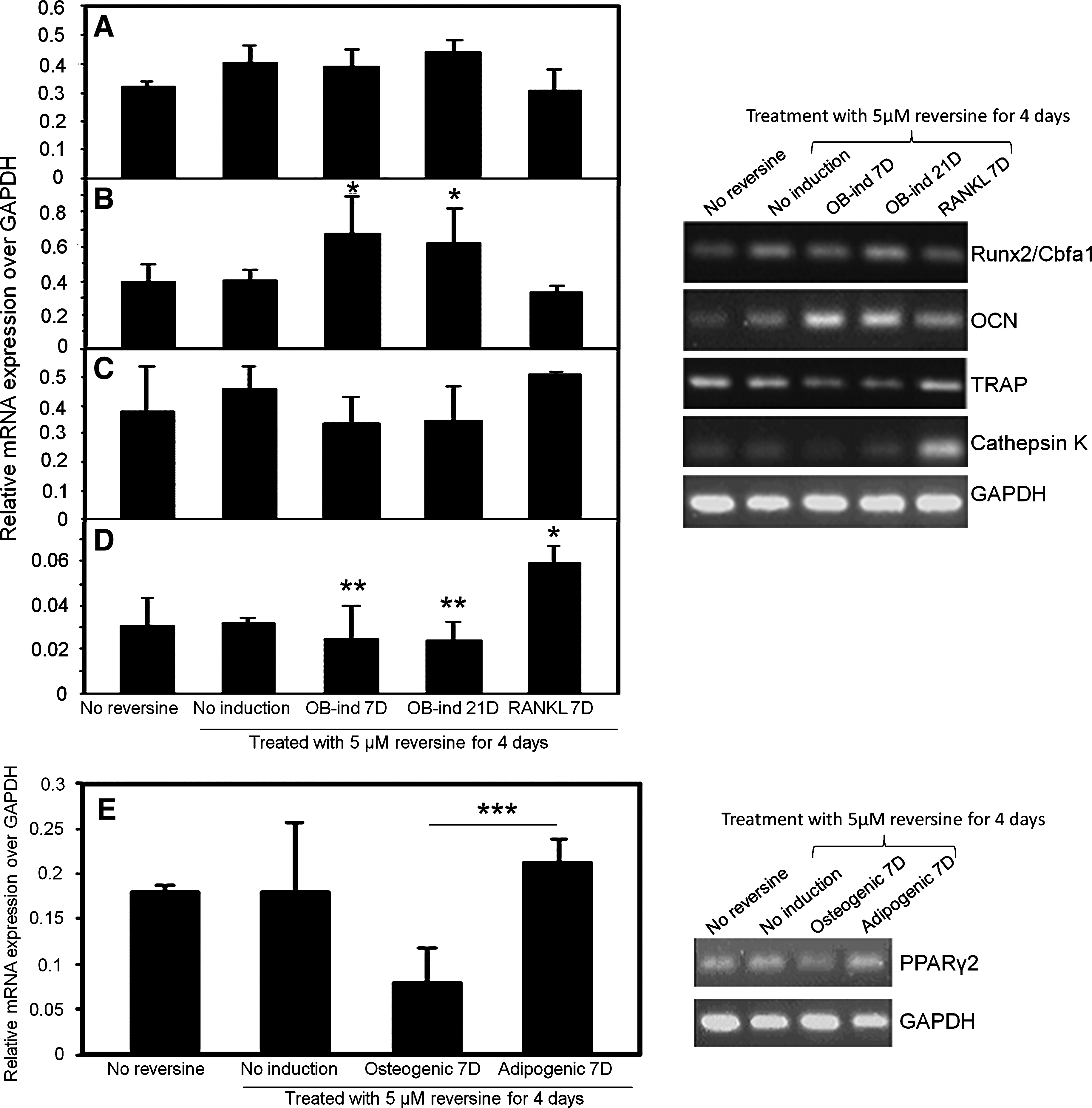
Reverse transcriptase–polymerase chain reaction analysis of selected genes expressed by representative dishes of STRO-1+ cells under osteogenic and lipogenic conditions. Runx2/Cbfa1 was elevated, and osteocalcin (OCN) significantly increased in the osteogenic induction medium.
The STRO-1+ cells maintained in an osteogenic inductive medium showed slight decreases in expression of the osteoclastic markers TRAP and cathepsin K compared to the control cells in a standard medium (NS; Fig. 8C, D). However, there was a significant increase in cathepsin K expression (Fig 8D, p<0.05) when grown in a medium containing RANKL (a stimulator of osteoclast differentiation from RAW cells), but no significant change in TRAP mRNA expression after treatment with reversine (Fig. 8C).
STRO-1+ cells in a lipogenic conditioned medium demonstrated increased Oil-red O staining (Fig. 7C vs. 7D). Those cells also demonstrated a 1.2-fold higher expression of PPARγ2 mRNA after reversine treatment (NS, Fig. 8E) and a 2.3-fold decrease in PPARγ2 mRNA expression in osteogenic media as determined by RT-PCR (p<0.05 vs. adipogenic media).
Reversine-treated RAW cells that were seeded on decalcified human bone scaffolds resulted in production of a calcified matrix within the scaffold (Fig. 9). Calcification could be seen at 4 weeks of culture and continued to increase until the end of the experiments at 6 weeks. Untreated cells did not result in any scaffold calcification. Fetal rat calvarial cells, which include osteoblasts and progenitors, also mineralized the scaffolds and were used as a positive control.
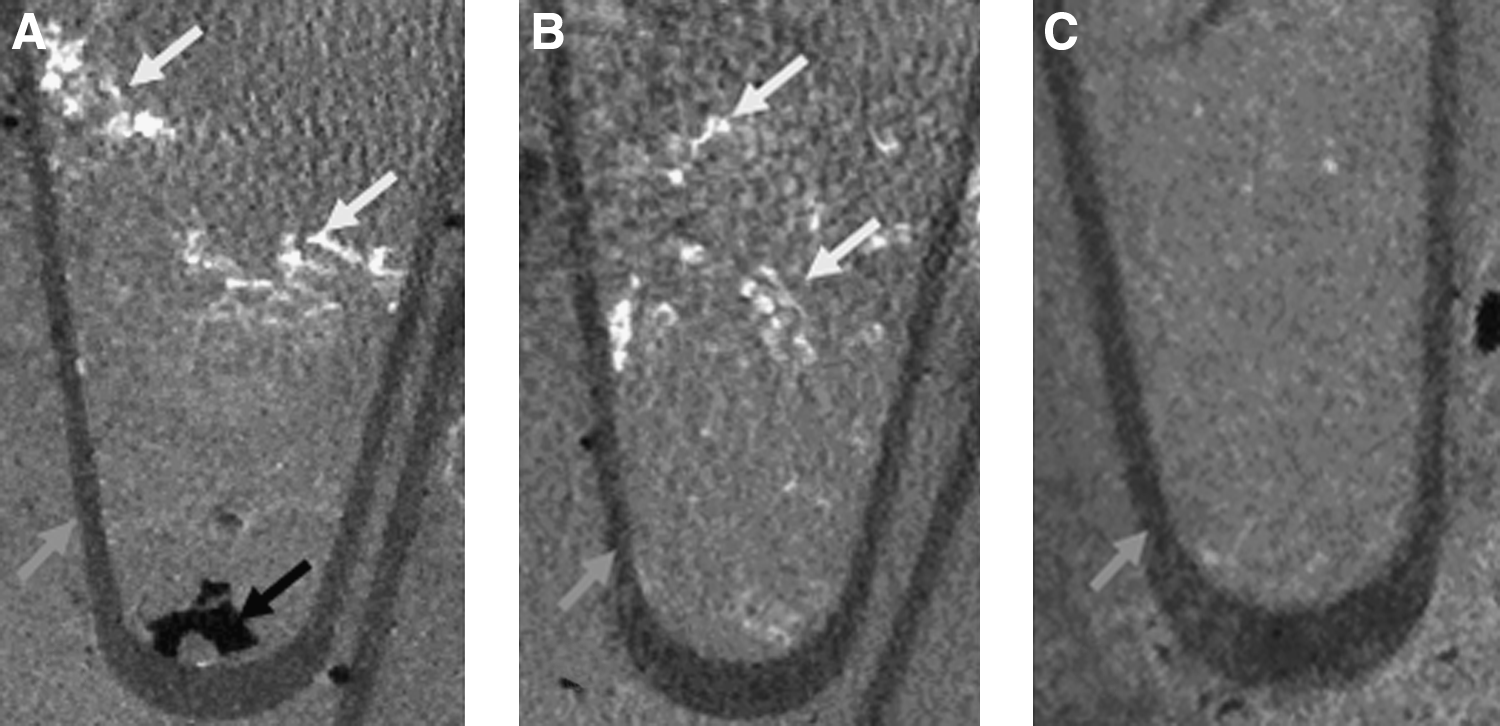
Micro-CT analysis of demineralized human bone seeded with
Discussion
This is the first report of dedifferentiation of the committed murine hematopoietic RAW 264.7 cell line into STRO-1+ mesenchymal-like cells by treatment with the purine analog reversine. Further, this preliminary report demonstrates that these STRO-1+ cells can regain a multipotential function. They could be redifferentiated into cells with osteogenic and lipogenic characteristics after culture under appropriate conditions, and could produce a calcified extracellular matrix when seeded on a demineralized bone scaffold.
Reversine was initially investigated for potential properties as a dedifferentiation agent and has been shown to have such an effect on myoblasts, primary murine and human dermal fibroblasts, 18 and possibly rat hippocampal neurons. 19 Similar to our experiments with RAW cells, Chen et al. showed that human primary skeletal myoblasts and the C2C12 myoblast cell line treated with reversine gained the ability to differentiate into cells with characteristics of osteoblasts and adipocytes,17,24 and reversine has also been shown to directly stimulate adipocyte differentiation of 3T3-L1 cells. 30
The process described by Chen et al. 17 may actually represent transdifferentiation, as opposed to dedifferentiation, because C2C12 cells used in their study have the ability to direct their differentiation to a variety of different cell types upon specific treatments. 31 In our study, the expression of STRO-1 supports the concept that dedifferentiation had occurred back to a very early developmental stage in the RAW cell line. STRO-1 is not known to be expressed in hematopoietic cells,32,33 and its appearance in ∼20% of treated cells represents a specific change in cellular pathways. The expression of STRO-1 is a characteristic of less well-differentiated cells of the osteoblast lineage and is commonly used to define a mesenchymal stem cell progenitor subset that has the potential to develop into variety of cell types.34–36 The appearance of STRO-1 antigen might imply a novel cell change from committed cells backward to an earlier stage of a more immature progenitor as a result of treatment with reversine. The expression of STRO-1 in the dedifferentiation process was dependent on the duration of treatment (Fig. 4), and cells could not be dedifferentiated to the earlier stage that is characterized by SH-2 (CD105) expression 20 (Fig. 2). Since only 20% of RAW cells were capable of dedifferentiation, we speculate that this is a result of the RAW 264.7 cell line being a heterogeneous hematopoietic culture.37,38 This heterogeneity could provide interesting findings with further study into a more specific hematopoietic cell phenotype that is capable of redifferentiation.
The role of reversine in dedifferentiation has been inferred by specific studies using chondrocytes, 39 Sertolli cells, 40 and human schwannoma cells. 41 Recent studies using a variety of methods such as cell sorting have focused on the use of hematopoietic cells to obtain other cell types, including mesenchymal-like cells.3,5–9,42,43 Our data are preliminary evidence that reversine-induced RAW cell-derived STRO-1+ cells access to a stage-limited reversal process. These STRO-1+ cells exhibited mesenchymal progenitor-like features with a multipotent status and appear to be capable of giving rise to two different cell types (osteoblasts and adipocytes) under the defined conditions of inductive differentiation (Fig. 7 and Fig. 9). However, further experiments are required to confirm that they are typical osteoblasts and/or adipocytes. These cells could provide a novel outlet for developing tissue regeneration models in mice for various diseases.
Cellular apoptosis has been reported with glucocorticoid treatment such as with dexamethasone.44–46 RAW264.7 cells exposed to dexamethasone in vitro have reported decreased NO/iNOS and no or little effect on RAW cell apoptosis47,48; elevated NO levels are associated with cell death.49,50 STRO-1+ cells exposed to dexamethasone have not reported elevations in apoptosis in similar types of studies.5–9 However, we cannot rule out that some of the mineralization detected in our cells is a result of dystrophic mineralization and will require further evaluation. Despite this, we believe that mineralization resulting from apoptosis is unlikely, as cells were monitored for cell death throughout the study. The mineralization capabilities of the dedifferentiated RAW cells were demonstrated by von Kossa, alizarin red (Fig. 7), as well as micro-CT (Fig. 9). While reports of dystrophic/nonphysiologic cases of mineralization have been reported and cautioned when studying mineralization,51,52 reports have also suggested that in the absence of a more detailed matrix evaluation methods (Fourier transform infrared spectroscopy [FTIR] and electron microscopic analysis [EM]), using both staining methods employed in this study and testing for enhancement of mineralization markers such as osteocalcin (OCN) (Fig. 8) are sufficient indications of mineralization. 52 Thus, while our studies suggest that the observed mineralization is a real affect, further analysis of the mineral produced is required to confirm that it is consistent with that of typical osteoblasts.
We would have expected the expression of PPARγ in untreated cells to be zero when examining expression of cell markers. We also noted that peak expression of OCN in these cultures was at day 7, which is not typical of osteoblast (OB) cultures (Fig. 8). This could be the result of the cells being derived from an alternate cell source. However, as this is a preliminary study, we would expect further examination of expression profiles over a complete time course and using proper qRT-PCR techniques to provide a more definite understanding of changes and peak marker expressions in this system. In addition, it will highlight differences in this novel dedifferentiated cell line with respect to typical OB cultures from mesenchymal origins.
The concept of physiological dedifferentiation is an area of increasing research interest.53–55 Dedifferentiation is however generally a controversial concept, 56 primarily because lineage commitment and cellular differentiation are typically considered to be unidirectional and irreversible processes. It is also controversial within the context of regenerative medicine, because dedifferentiation occurs during certain types of oncogenesis. The identification of specific factors that can reprogram cells to pluripotential cells13,15 implies that dedifferentiation is physiologically possible, and suggests that such processes are mediated by tightly controlled gene expression and modulation of specific signaling pathways as per any other cell process. It does not necessarily imply the risk of oncogenesis.
Reversine is a moderately potent antagonist for the human A(3) adenosine receptor (AR), 56 a G protein-coupled receptor that is widely expressed in most human cells. Chen et al. found that reversine had a specific mode of function by inhibition of MEK1 and nonmuscle myosin-II heavy chain that resulted in altered cell cycle and changes in the histone acetylation status; reversine also resulted in activation of the PI3K-signaling pathway. 24 Activation of PI3K may in turn lead to activation of JNK. In the present study, there is evidence that the effect of reversine could be significantly reduced (by 67%; Fig. 6) by blocking the JNK pathway with the inhibitor SP600125. The JNK protein, a member of the MAP kinase family, is activated by various cell stimuli and acts as an integration point for multiple biochemical signals and targets specific transcription factors. Activation of JNK results in phosphorylation of nuclear substrates such as c-Jun, a component of the AP-1 transcription factor family, to mediate nuclear events associated with a wide variety of cellular processes, including differentiation, but also proliferation, transcription regulation, development, and cell death.57,58
The role of reversine in cell death appears to be controversial. In the original paper by Chen et al., 17 no cell death was noted. Anastasia et al. showed a 30% cell death rate within the first day of reversine treatment. 18 Our results were under 20% for 2.5- and 5-μM doses of reversine after 4 days of treatment, but cell death of over 80% for 10 μM of reversine clearly showed a toxic effect at high doses. Mathematical calculations of the expected versus actual STRO-1 cells with or without cell death concluded that the increase in STRO-1 cells was 26 times higher than that which could be accounted for by cell selection. Thus, an actual activation of cellular mechanisms toward STRO-1 expression did occur. In addition to the dedifferentiation to STRO-1+ progenitor cells, the mixture of dead cells and multinuclear cells in one culture system implied that reversine has a heterogeneous and multifunctional effect on RAW cells by inducing cells toward different developmental directions.
Although stem cells have a great potential for the regeneration of damaged tissues in countless diseases, problems such as limited availability, control of cell fate, ethical issues, and allograft rejection need to be solved before therapeutic applications may become possible. As a cell source, blood is a highly practical donor tissue, since it is readily available, renewable, and can be easily obtained with minimal morbidity. The generation of multipotential progenitors from differentiated adult human cells by agents such as reversine is a striking alternative to the limited self-renewal of stem cells in vitro.
Footnotes
Acknowledgments
We are grateful to our funding sources: Division of Orthopaedic Surgery and Department of Surgery, University of Toronto, Canada; University Health Network, Toronto, Canada. We thank Dr. Diana Trebec-Reynolds for editing the article.
Disclosure Statement
No competing financial interests exist.