
Abstract
Several challenges currently exist for rational design of functional tissue engineering constructs within the host, which include appropriate cellular integration, avoidance of bacterial infections, and low inflammatory stimulation. This work describes a novel class of biodegradable, amphiphilic polyanhydrides with many desirable protein-material and cell-material attributes capable of confronting these challenges. The biocompatible amphiphilic polymer films were shown to release laminin in a stable and controlled manner, promote neural cell adhesion and differentiation, and evade inflammatory responses of the immune system. Using high-throughput approaches, it was shown that polymer chemistry plays an integral role in controlling cell–film interactions, which suggests that these polyanhydrides can be tailored to achieve the desired cell adhesion and differentiation while minimizing immune recognition. These findings have important implications for development of engineered constructs to regulate differentiation and target the growth of transplanted cells in stem cell-based therapies to treat nervous system disorders.
Introduction
Many current polymer systems lack the ability to provide controlled release of biologically active molecules, and some fragile proteins may lose their function when encapsulated in biodegradable polymers such as poly(lactide-co-glycolide) (PLGA).4–6 In addition, with bulk eroding polymers such as PLGA, the structural integrity of the device may be compromised because once the molar mass is low enough, there is a rapid loss of mechanical properties.2,7,8 In contrast, surface eroding polymers are characterized by a slow loss of mechanical stability over time, which would reduce the possibility of scaffold collapse before new tissue formation and integration with the host. 2 In addition to mechanical integrity, it is important for the polymer construct to evade clearance by the immune system and avoid an inflammatory response. Inflammation and immune activation can lead to the buildup of fibroblasts, which inhibit tissue integration and implant acceptance by the host.1,2 Stimulation of the innate immune system can occur through mechanisms such as the binding of pathogen-associated molecular patterns to pattern recognition receptors on the surface of immune cells and/or complement activation. Thus, it would be desirable to design surface eroding polymeric constructs for tissue engineering that are capable of modulating the inflammatory response of the immune system to provide a controlled environment conducive to cellular growth and regeneration.
Polyanhydrides have been studied extensively as vehicles for protein and vaccine delivery over the past decade9–27 ; however, little work to date has investigated their use in tissue engineering applications. These surface eroding polymers are biologically inert, nontoxic, and nonmutagenic and are capable of providing sustained release kinetics of encapsulated proteins.11,13,14,17,19,20,24 Additionally, polyanhydrides have been shown to stabilize a wide range of biologically active molecules, such as F1-V (vaccine antigen for pneumonic plague), PA (vaccine antigen for anthrax), lipocalin-2, ovalbumin, lysozyme, and tetanus toxoid (vaccine antigen for tetanus).11,13–15,17,19,21,24,28 Specifically, polyanhydrides based upon 1,6-bis(p-carboxyphenoxy)hexane (CPH) and 1,8-bis(p-carboxyphenoxy)-3,6-dioxaoctane (CPTEG) have been studied extensively because of their amphiphilic properties, controlled protein release and protein stabilization capabilities, immune modulation, and cellular compatibility.9,22,23,25–27 These polymers can also be functionalized to target-specific cellular receptors. 12 Upon degradation, CPTEG:CPH copolymers result in small changes in pH (7.6–7.1) unlike other systems (i.e., PLGA), which result in much greater decreases in pH to as low as 2.6.19,29–32 These properties make this polymer system a promising candidate for tissue engineering applications combined with targeted delivery and/or surface functionalization.
The focus of this work was to investigate amphiphilic CPTEG:CPH polyanhydride copolymer films for their ability to control protein release and stability while providing a nontoxic, immune-stealthy, and cell conducive environment for neural stem cell growth and adhesion. The findings indicate that CPTEG:CPH copolymer films provide an ideal nonfouling substrate for cellular exclusion, but with the incorporation of extracellular matrix proteins, cellular adhesion is restored. Additionally, by altering the chemistry of the polymer, this polymer system was capable of modulating immune stimulation, protein release kinetics, and cellular differentiation. These studies were performed using a high-throughput, combinatorial approach, which enabled the rapid assessment of protein–polymer and cell–polymer interactions.
Materials and Methods
Polymer library synthesis and characterization
CPH and CPTEG monomers were synthesized as described previously.33,34 CPTEG:CPH copolymer film libraries were synthesized via a melt polycondensation reaction in multiwell substrates utilizing a robotic deposition apparatus, as reported previously.20,21,23,34 Copolymer chemistry and molecular weight were determined by proton nuclear magnetic resonance ( 1 H NMR) spectroscopy using a Varian VXR 300 MHz spectrometer (Varian, Inc.). Gel permeation chromatography (GPC) was also used to measure the polymer molecular weight with a Waters GPC chromatograph.
Fabrication of protein encapsulated and blank (no protein) film libraries
Blank and laminin-loaded combinatorial film libraries were fabricated from the polymer libraries as described previously.20,23 Briefly, laminin and methylene chloride were added to the polymer film library and sonicated, and the protein/polymer solution in each well was separately dispensed onto glass coverslips (1.5 mg polymer/coverslip at 1% laminin loading [for a total of 15 μg of laminin]) and allowed to dry resulting in a protein-encapsulated film library.
High-throughput protein release
After encapsulation of laminin into the polymer films, the films were placed in a 24-well polystyrene plate, and 2 mL PBS buffer (0.1 mM, pH 7.6) added to each well. The plate was sealed to prevent evaporation and incubated in a horizontal shaker at 37°C and 100 rpm for the duration of the experiment. At incremental time points, samples of the supernatant were removed and replaced with fresh PBS buffer to maintain constant sink conditions. Laminin concentration in each sample was determined with the microbicinchoninic acid assay (Thermo Scientific), which was carried out in triplicate. The mass of protein released at each time point was normalized by the total mass of protein encapsulated into the film, which is presented as cumulative mass fraction of released laminin. 20
Culture and stimulation of dendritic cells
All procedures involving animals were conducted in accordance with procedures that were approved by the Iowa State University Institutional Animal Care and Use Committee. Bone marrow-derived dendritic cells (BMDCs) were isolated from C57BL/6 mice (ISU Laboratory Animal Resource Facility) and cultured using a previously developed method.12,22,23,26 BMDCs were grown in DC culture medium (RPMI containing 1% L-glutamine, 1% penicillin–streptomycin solution, 2% HEPES, 0.5% gentamycin, 0.1% β-mercaptoethanol, and 10% heat inactivated fetal bovine serum) supplemented with granulocyte macrophage colony-stimulating factor (10 ng/mL). After 9 days of culture, DCs were incubated for 48 h with the different treatments: polymer films [poly(CPTEG), 75:25 CPTEG:CPH, 50:50 CPTEG:CPH, 25:75 CPTEG:CPH, and poly(CPH), at a concentration of 1.5 mg/coverslip], no polymer (NP; negative control), and lipopolysaccharide (LPS; 200 ng/mL, positive control).
Cell surface marker expression
The expression of cell surface markers, including CD11c, CD86, CD40, MHC I, and MHC II, was assessed with flow cytometry as described elsewhere.23,25 The samples were run on a Becton-Dickinson FACSCanto flow cytometer and analyzed with FlowJo (TreeStar, Inc.).
Cytokine production
After incubation with the treatments, supernatants from DC cultures were collected and assayed for cytokines, TNF-α, IL-6, and IL-12, using the Luminex® Multiplex assay.23,25
MTT cellular toxicity assay
The CellTiter 96® Non-Radioactive Cell Proliferation Assay MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) (Promega) was used to determine the cellular viability of DCs incubated with the polymer films for 48 h. The DCs were incubated with the MTT assay for 3 h and the cell viability was quantified by measuring the optical density.
Culture and stimulation of human neural progenitor cells
Human neural stem/progenitor cells (hNPCs), isolated from 16- to 18-week fetal cortex, were purchased from Lonza. 35 The hNPCs were cultured as neurospheres in maintenance medium (a proprietary medium, Bulletkit; Lonza) further supplemented with 10 ng/mL human recombinant basic fibroblast growth factor (Promega, Madison, WI) and 20 ng/mL epidermal growth factor (Gibco). 35 This maintenance medium is referred to as MM. At day 0 the cells were plated at ∼10 neurospheres per well (each well containing a 12-mm glass coverslip coated with the different polymer film treatment groups) in a 24-well plate in 500 μL of differentiation medium (a proprietary medium, Bulletkit; Lonza) further supplemented with 20 ng/mL recombinant human brain-derived neurotrophic factor (BDNF; Sigma-Aldrich). The differentiation medium is referred to as DM. The polymer film treatment groups included poly(CPTEG), 75:25 CPTEG:CPH, 50:50 CPTEG:CPH, and 25:75 CPTEG:CPH (blank or 1% laminin loaded in the polymer film) at a polymer coating of 1.5 mg/coverslip. The control treatment groups included laminin-coated coverslips incubated either in DM with BDNF as a positive control (laminin DM) or in MM as a negative control (laminin MM). Cultures with the different treatment groups were maintained for 7 days. Culture media were replenished with 350 μL of fresh media at days 3 and 5.
Immunocytochemistry on hNPCs
After the 7-day incubation period, the hNPCs were immunolabeled. The coverslips containing the cells were rinsed in 0.1 M phosphate (PO4) buffer and then fixed with 4% paraformaldehyde in 0.1 M PO4 buffer for 30 min. After fixation, the cells were washed in PBS and then incubated with the primary TUJ1 antibody (mouse monoclonal IgG; R&D Systems) diluted (1:200) in blocking solution (0.4% BSA [Sigma-Aldrich], 0.2% Triton X-100 [Fisher Scientific], and 2.5% normal goat serum [Jackson ImmunoResearch, Inc.]) overnight at 4°C. The next day the cells were rinsed in PBS and incubated with a goat anti-mouse secondary antibody (Cy5-conjugated; Jackson ImmunoResearch, Inc.) diluted (1:500) in the same blocking solution for 2 h. Cells on the coverslips were rinsed in PBS and then incubated with a Alexa Fluor 488 (AF488)–conjugated primary microtubule-associated protein 2B (MAP2B) antibody (mouse monoclonal IgG; BD Pharmingen) and a Cy3-conjugated primary glial fibrillary acidic protein (GFAP) antibody (mouse monoclonal IgG; Sigma-Aldrich) diluted in the blocking solution (1:18 and 1:400, respectively) overnight at 4°C. The next day, cells on the coverslips were rinsed in PBS. Then, nuclei were counterstained with 1 μM 4′,6-diamidino-2-phenylindole diactate (DAPI) and the preparations were mounted on microscope slides with Gel Mount (Fisher Scientific) for fluorescence microscopy analysis.
Microscopy, image acquisition, and data analysis
Epifluorescent microscopy was carried out with an upright fluorescence microscope (Nikon Microphot FXA; Nikon, Inc.) equipped with a Retiga 2000R digital camera controlled by QCapture software (QImaging). A minimum of eight images with an average of 222 cells per image was taken per fluorochrome per treatment group using a 20× objective. ImageJ software (NIH, Bethesda, MD) was used to analyze the microscopy images. To calculate the percentage of immunoreactive cells, the number of cells immunoreactive for each antibody was divided by the total number of cells (DAPI-stained nuclei).
Statistical analysis
JMP software (SAS Institute) was used to make comparisons between multiple different treatments using a model ANOVA with Tukey's HSD as indicated by letters above each bar. Differing letters represent difference between treatments in Figures 2, 3, and 5 and Supplementary Figures S1 and S2 (Supplementary Data are available online at www.liebertonline.com/tea). In Figure 2A comparisons between each treatment and the control were made using the Student's t-test and indicated by an asterisk above the bars.
Results
Polymer characterization and protein release
The NMR and GPC characterization indicated that polymer molecular weight, structure, and composition were as intended and in agreement with previous studies.24,34,36 High-throughput laminin release studies from CPTEG:CPH copolymer films (Fig. 1) were carried out because laminin is an adhesion protein for cellular binding to substrates.37–39 Figure 1 shows that laminin was released in a controlled manner over time as a function of polymer chemistry. By increasing the content of the less hydrophobic CPTEG in the copolymer, protein release rates were higher.
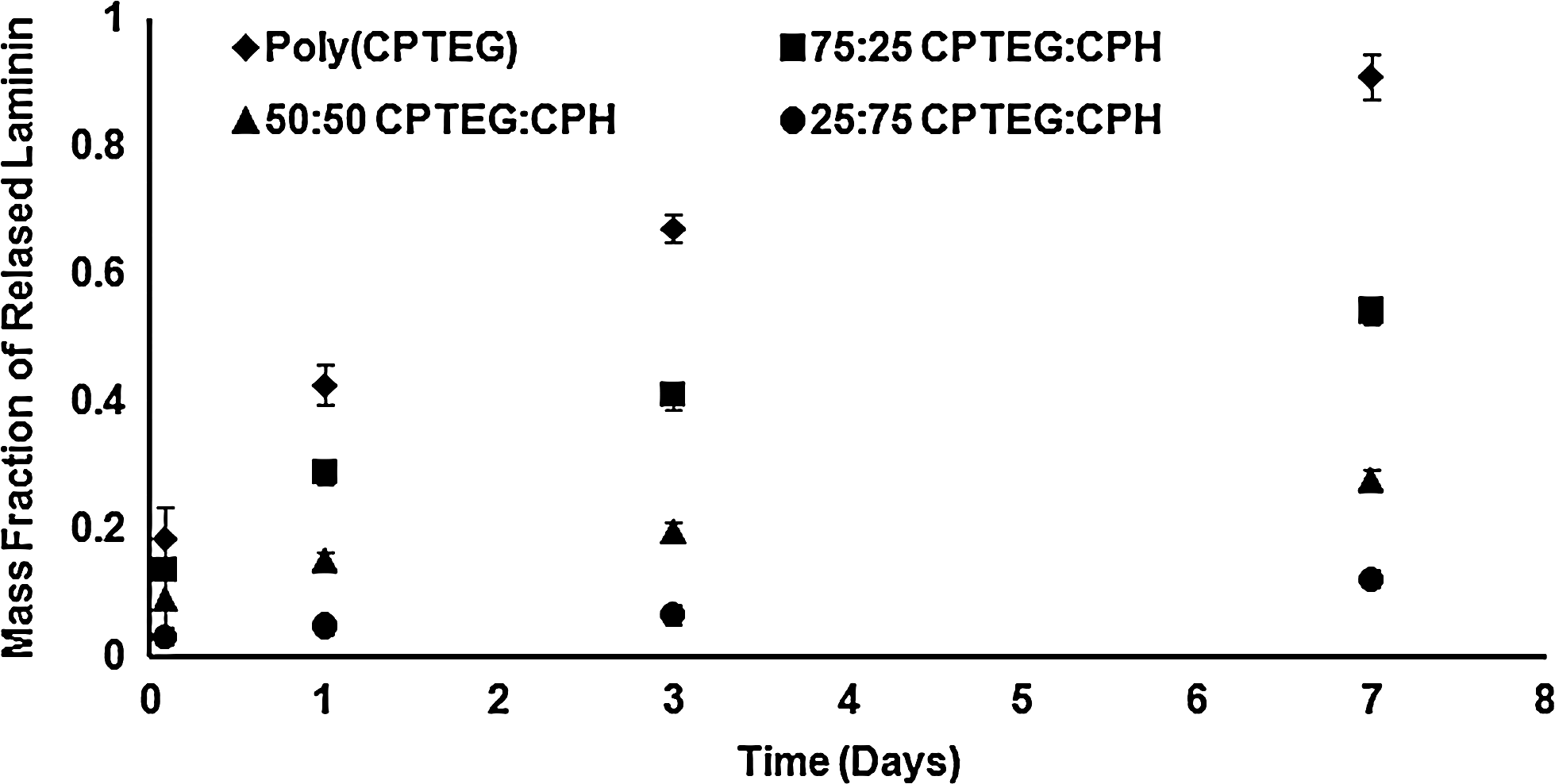
Cumulative mass fraction release of laminin from CPTEG:CPH copolymer film libraries. The CPTEG-rich films released laminin most rapidly, whereas CPH-rich films released laminin the slowest. Error bars represent standard deviation and six independent experiments were carried out. CPH, 1,6-bis(p-carboxyphenoxy)hexane; CPTEG, 1,8-bis(p-carboxyphenoxy)-3,6-dioxaoctane.
Polyanhydride libraries are biocompatible and induce low levels of immune response
To test the toxicity of polyanhydride film libraries to primary cells, dendritic cells (DCs), which are professional antigen presenting cells of the immune system, were incubated with varying concentrations of 50:50 CPTEG:CPH copolymer films and cell viability was examined by the MTT assay (Fig. 2A). All polymer concentrations below 16.9 mg/mL resulted in no statistical difference for the cell viability compared to the NP-positive control. This result signifies that a wide range of polymer coating or scaffold thicknesses could be utilized with this system.
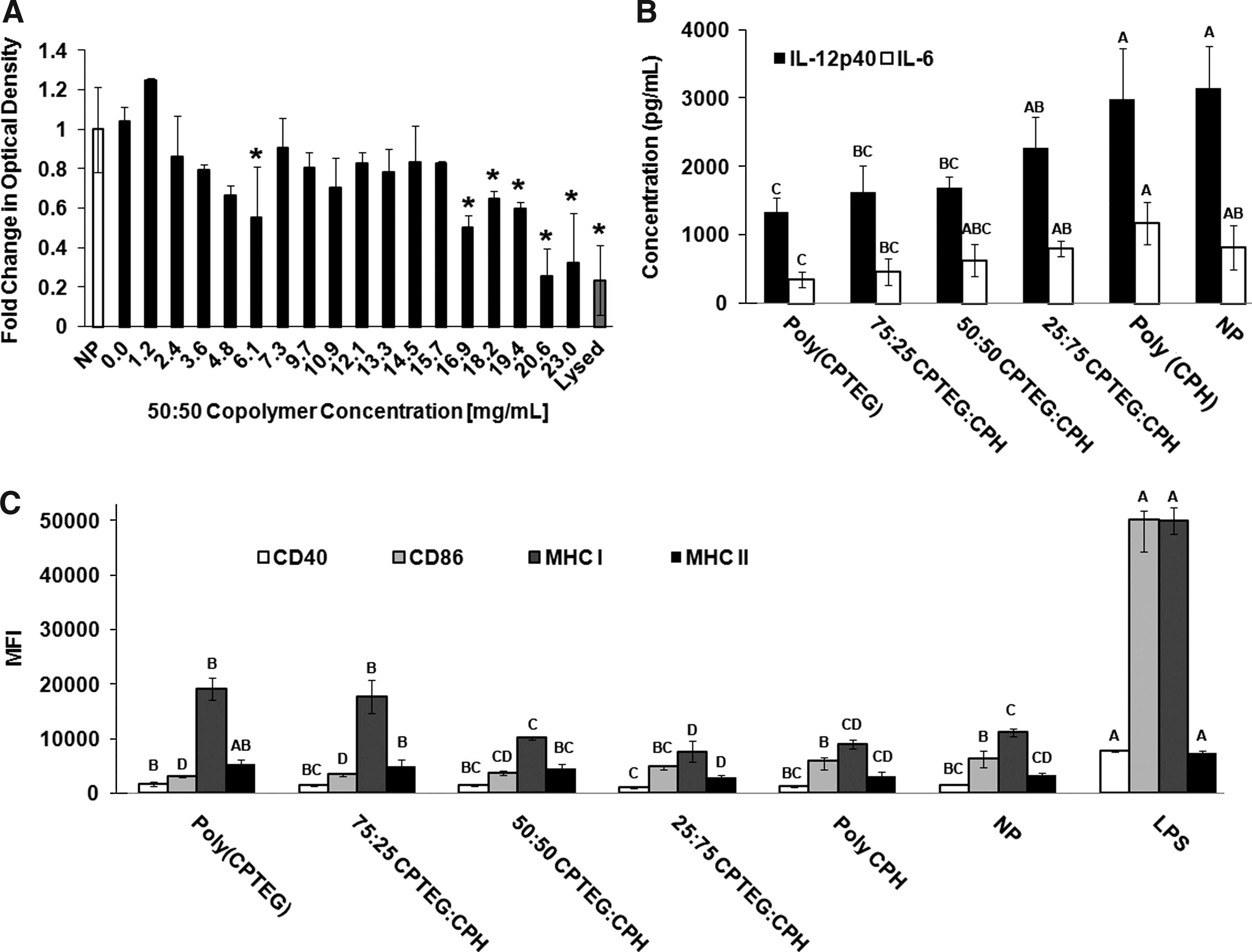
Biocompatibility and immune stimulation capabilities of CPTEG:CPH polymer films.
To test the effects of the film libraries on induction of an immune response, DCs were incubated on the polymer films for 2 days. Supernatants were analyzed for secreted cytokines, TNF-α, IL-6, and IL-12p40, and cells were stained for expression of cell surface markers, MHC I, MHC II, CD40, and CD86, and analyzed by flow cytometry. Secretion of these cytokines and expression of these cell surface markers are indicators of immune activation and DC maturation. 40 The results showed that the CPTEG:CPH polymer films cause DCs to downregulate cytokine secretion (IL-12p40 and IL-6) compared to the NP control (Fig. 2B). TNF-α was below detection limits for all treatments except LPS. Additionally, low levels of cell surface marker expression were observed (Fig. 2C). Both cell marker expression and cytokine production appeared to be modulated in a chemistry-dependent manner. These findings suggest that there may be an optimal copolymer composition, such as 50:50 CPTEG:CPH, that is capable of achieving the lowest level of immune stimulation.
Adhesion of hNPCs to the polymer library is controlled by polymer chemistry
hNPCs were incubated for 7 days with the polymer film library and investigated for their ability to adhere and differentiate. Initial studies evaluated the adhesion of hNPCs on blank or laminin-encapsulated polymer film libraries, including poly(CPTEG), 75:25 CPTEG:CPH, 50:50 CPTEG:CPH, and 25:75 CPTEG:CPH. The results indicated that blank polymer films provided a nonfouling background preventing cellular adhesion with all polymer chemistries (data not shown). However, when laminin was encapsulated into the polymer films, cellular adhesion was restored in for CPH-rich polymer film chemistries (data not shown). These results indicate that the biological functionality of laminin was preserved upon encapsulation into the polymer films. In addition, CPTEG-rich polymer chemistries resulted in minimal cellular adhesion (even with laminin encapsulated into the polymer) (data not shown), which is likely a result of the ethylene glycol1,41 incorporated into the polymer backbone.
Differentiation of hNPCs is controlled by polymer chemistry
The three polymer film chemistries with adequate cellular adhesion (75:25 CPTEG:CPH, 50:50 CPTEG:CPH, and 25:75 CPTEG:CPH) were studied for their influence on hNPC differentiation. Figure 3 suggests that film chemistry controlled the differentiation of hNPCs with 50:50 CPETG:CPH resulting in the lowest percentage of hNPCs immunoreactive for MAP2B, a marker of immature neurons. The other two chemistries resulted in significantly higher percentages of MAP2B-immunoreactive cells. The percentages of cells immunoreactive for beta III tubulin (TUJ1, marker of developing neurons) or GFAP (astrocyte marker) were increased in the presence of CPTEG-rich chemistries. The data in Figure 3 suggest that 75:25 CPTEG:CPH polymer films enhance the most hNPC differentiation into both astrocytes and neurons compared with other film treatments and control treatments (positive control: laminin-coated glass coverslip with NP film in differentiation media [laminin DM]; negative control: laminin-coated glass coverslip with NP film in maintenance media [laminin MM]).
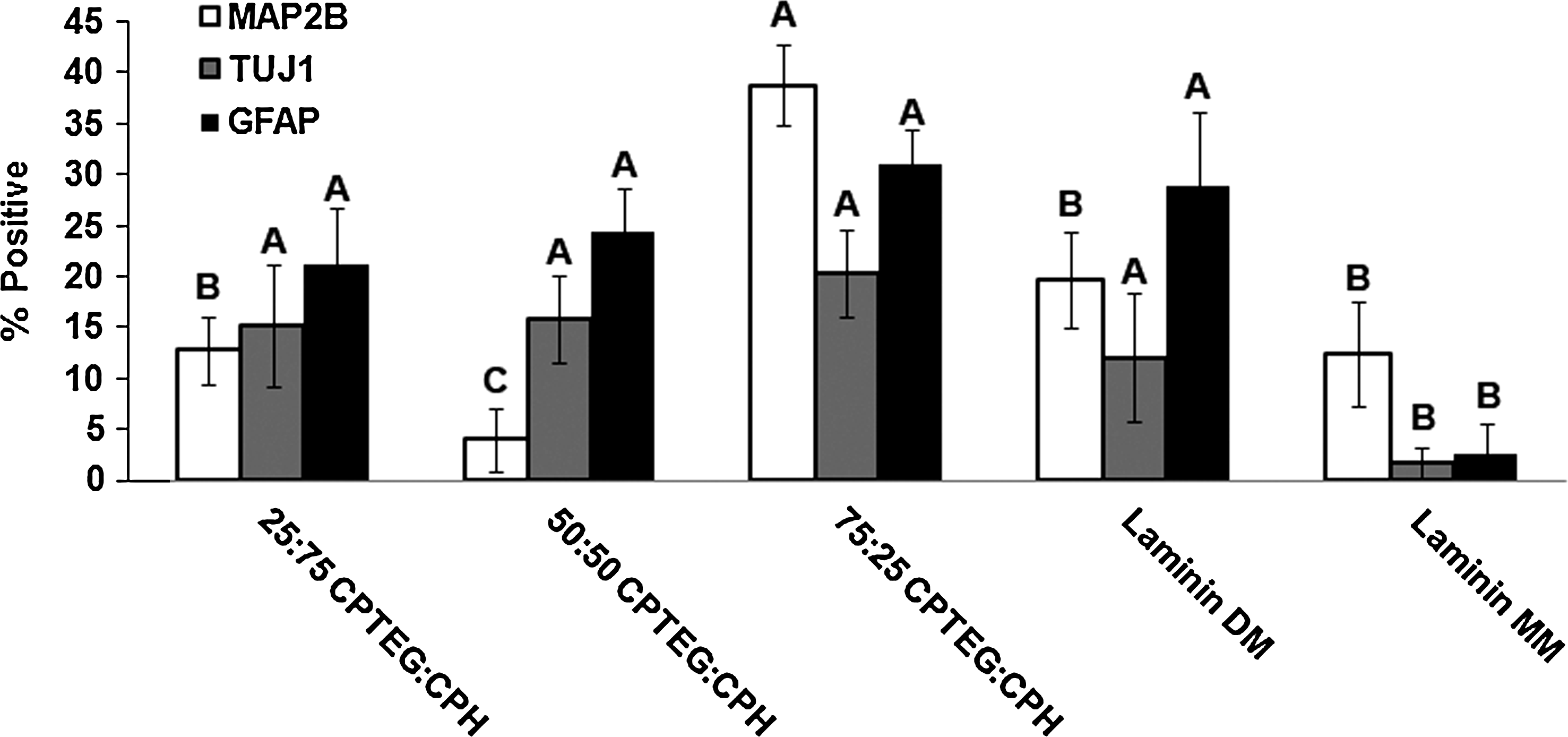
Percentage of hNPCs displaying neuronal (MAP2B and TUJ1) or glial (GFAP) differentiation. The CPTEG-rich films best enhanced hNPC differentiation. Letters indicate statistical significance between treatments for each cell marker, eight different images were analyzed with an average of 222 cells per image, and error bars represent standard deviation. GFAP, glial fibrillary acidic protein; hNPCs, human neural stem/progenitor cells; MAP2B, microtubule-associated protein 2B.
Supplementary Figure S1 presents the percentages of hNPCs that were immunostained for MAP2B (Supplementary Fig. S1A) and TUJ1 (Supplementary Fig. S1B), respectively. The data in Supplementary Figure S1A suggest that higher fraction of hNPCs were undergoing neuronal differentiation when they were incubated on CPTEG-rich polymer films (75:25 and 50:50 CPTEG:CPH) than on 25:75 CPTEG:CPH films. In addition, the percentage of cells immunoreactive for MAP2B in both cell body and neurites was higher when the cells were cultured on 75:25 CPTEG:CPH films than CPH-rich polymer films. Supplementary Figure S1B also demonstrates increasing trends for the percentages of TUJ1-positive hNPCs (immunolabeled in both cell body and neurites) when incubated with increasing CPTEG content in the polymer film.
Images of each treatment group and marker can be observed in Figure 4 and corroborate the data presented in Figure 3. By quadruple labeling the hNPCs (DAPI-nucleus, AF488-MAP2B, Cy5-TUJ1, and Cy3-GFAP), double-positive cell populations for the differentiation markers were assessed as shown in Figure 5. Figure 5A indicates that cells immunoreactive for MAP2B are also largely positive for TUJ1 or GFAP, especially when incubated on CPH-rich polymer films. However, cells immunoreactive for TUJ1 or GFAP are less positive for each other, but follow the same chemistry-dependent trend (Fig. 5B, C). The hNPC populations positive for MAP2B appeared to be more influenced by polymer film chemistry in their immunoreactivities for TUJ1 or GFAP than populations positive for TUJ1 or GFAP, which only resulted in chemistry-dependent trends for colabeling with GFAP or TUJ1, respectively. When examining the entire hNPC population (based upon DAPI count) (Supplementary Fig. S2), the data indicate an opposite chemistry trend with CPTEG-rich polymer films influencing the most double-positive expression of cells for TUJ1 and GFAP, and GFAP and MAP2B. In contrast, double-positive cells for both neuronal makers, TUJ1 and MAP2B, trended toward CPH-rich chemistries.

Representative epifluorescent images of hNPCs stained for DAPI (blue, column 1), MAP2B (AF488 green, column 2), TUJ1 (Cy5 red, column 3), and GFAP (Cy3 yellow, column 4), after incubation with 25:75 CPTEG:CPH, 50:50 CPTEG:CPH, 75:25 CPTEG:CPH, laminin-coated coverslip in DM (positive control, Laminin DM), and laminin-coated coverslip in MM (negative control, Laminin MM). The hNPCs readily adhered to CPH-rich polymer films and strongly expressed differentiation markers. DAPI, 4′,6-diamidino-2-phenylindole diactate; DM, differentiation medium; MM, maintenance medium. Color images available online at www.liebertonline.com/tea
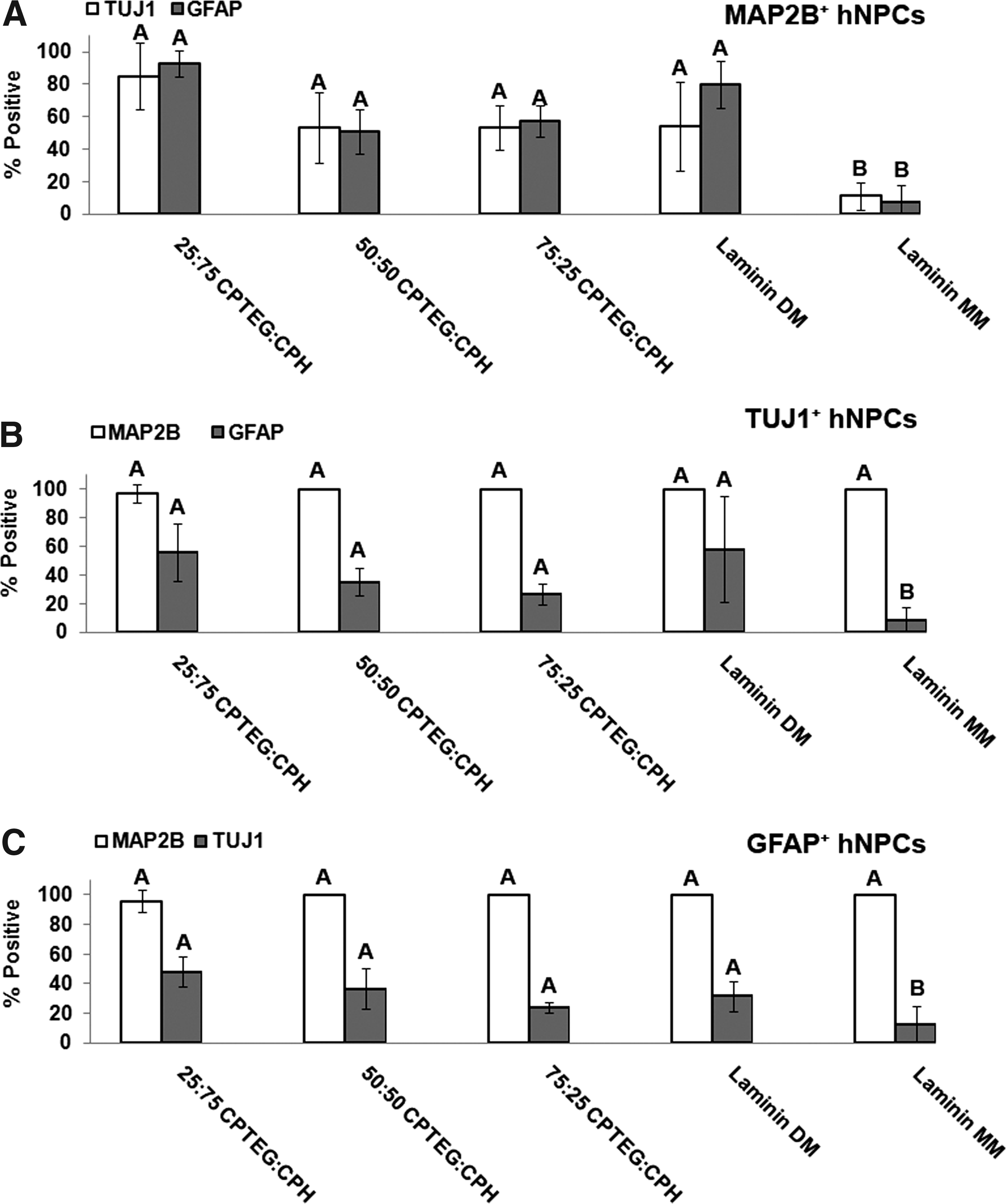
CPH-rich chemistries resulted in the most double-positive hNPC populations for MAP2B+TUJ1, MAP2B+GFAP, TUJ1+GFAP, and GFAP+TUJ1.
Discussion
Many current approaches to tissue engineering scaffolds or implant coatings have focused on the use of surface functionalization or controlled release of biological signals to promote cellular adhesion, growth, vascularization, differentiation, and so on, and/or to inhibit the growth and viability of infectious microorganisms.1,2 In this regard, CPTEG:CPH copolymers represent a biodegradable delivery system capable of both controlled release and surface functionalization. In this work these copolymers were demonstrated to release laminin in a chemistry-dependent manner, which was a strong function of the hydrophobicity of the polymer film (Fig. 1). All the polyanhydride chemistries tested in this work demonstrated the ability to preserve the functionality of laminin. This is consistent with other work from our laboratory on the protein stabilization capabilities of this amphiphilic polymer system.11,17,19,21,24 In those studies, the stabilization of fragile recombinant proteins as well as more robust globular proteins was shown to be strongly dependent upon polymer chemistry, with the amphiphilic chemistries providing the best environment for preservation of protein activity (and structure). Recently, Carrillo-Conde et al. demonstrated that CPTEG:CPH nanoparticles can be functionalized with carbohydrates with an eye toward targeting DCs for immune activation. 12 That approach could be applied to functionalize the polymer films in this work for promoting cell-specific adhesion and growth and/or for reducing bacterial adhesion and growth. Bactericidal activity of a tissue engineering scaffold or implant coating would be a key advancement for reducing implant infections, especially intracellular bacteria, which cannot be cleared by antibiotics and pose a threat to implant viability. 1
Biocompatibility and immune acceptance, specifically low inflammatory responses, of the implant by the host often dictate acceptance of the implant and in vivo viability. Materials capable of inducing low immune recognition would be ideal candidates for tissue engineering applications as coatings to noninflammatory implants or implants themselves. DCs were used to study the effect of CPTEG:CPH polymer films on cellular toxicity, expression of surface markers responsible for DC maturation and antigen presentation (MHC II, MHC I, CD86, and CD40), and secretion of both inflammatory and immune activating cytokines (TNF-α, IL-6, and IL-12p40). This work has demonstrated that the CPTEG:CPH polymer film system modulated the immune response of DCs in a chemistry-dependent manner (Fig. 2). Moderate to low CPTEG-containing copolymer compositions demonstrated simulation levels no higher than the negative (NP) control group. This indicates that these films could be stealth-like to the immune system. Further, all the chemistries studied maintained background levels or decreased production of the pro-inflammatory cytokines TNF-α and IL-6 (Fig. 2B). These data suggest that these films may elicit weak inflammatory responses in vivo. In other related work from our laboratories, when nanoparticles made of these polymers were administered intranasally or subcutaneously, very low levels of inflammation were observed. 42 Since nanoparticles can be phagocytosed, it is reasonable to expect that the same chemistry in nonphagocytic geometries (i.e., film, coating, and scaffold) would elicit even lower levels of inflammation similar to what was observed in vitro. 23
Stealth-like behavior enabling protection from immune activation and clearance by the immune system are important characteristics for implant or scaffold acceptance and tissue integration. However, it is also important that cell survival, adhesion, and differentiation within the polymer are conducive for cellular integration and tissue regeneration. As shown in Figure 3A, the CPTEG:CPH films were biocompatible at concentrations 30-fold higher than those administered in vivo.16,43 Adhesion and differentiation of hNPCs was also enabled by this system in a chemistry-dependent manner. Adhesion was not favored for the CPTEG-rich chemistries due to the ethylene glycol content in the backbone of poly(CPTEG). This is consistent with other studies in which low protein adsorption was observed on the surface of microparticles composed of CPTEG-rich chemistries. 10
Cellular differentiation is also influenced by chemistry with CPTEG-rich compositions influencing the most overall expression of cells positive for neuronal markers (Figs. 3 and 4). This suggests that 75:25 CPTEG:CPH has the ability to enhance differentiation of hNPCs into neurons and glial cells. In addition, 50:50 CPTEG:CPH may also be able to promote neural differentiation as indicated by the higher fraction of hNPC cell bodies immunoreactive for MAP2B (Supplementary Fig. S1A). Further, the double-positive phenotype of the hNPCs suggests that all treatments, except Laminin MM, result in a TUJ1-positive cell population that is largely MAP2B positive (∼100%) (Fig. 5); however, the MAP2B-positive population reveals a smaller population positive for TUJ1 (50–80%). It has been reported that TUJ1 is an axonal marker for immunolabeling maturing neurons44,45 and mammalian MAPs are specifically localized in dendrites and play a role in microtubule-stabilization and possibly supporting dendritic elongation.46,47 As such, immunolabeling with these two antibodies allows identification of different compartments within neurons (TUJ1: soma and developing axons; MAP2B: soma and developing dendrites). Thus, the variation observed between these two double-positive populations suggests differential maturation of the neuronal processes (e.g., dendrites vs. axons). Additional phenotypes of hNPC populations double positive for both neural (TUJ1 or MAP2B) and glial (GFAP) phenotypes can be identified in Figure 5 as well. During the differentiation process, it is possible that exogenous factors may have induced an “indeterminate” differentiation phenotype characterized by simultaneous expression of glial and neuronal markers (MAP2B and GFAP or TUJ1 and GFAP). 48
This work demonstrates the capabilities of CPTEG:CPH polymer films to control cellular adhesion and differentiation in a chemistry-dependent manner while providing a system capable of releasing biologically relevant proteins for further controlling cellular behavior. By employing a combinatorial platform approach, these protein–material and cell–material interactions were rapidly screened, which will facilitate the rational design of tissue engineering systems composed of biodegradable polyanhydrides.
Conclusions
Amphiphilic polyanhydride films provide a biologically tunable environment capable of overcoming many of the challenges associated with tissue engineering constructs. They were capable of preserving protein structure while controlling protein release, which is integral for dictating cellular behavior (adhesion, differentiation, etc.). The polymer films also demonstrated immune stealth characteristics indicated by background level stimulation of DCs and low inflammatory signaling. These attributes are important for implant host acceptance, whereas other properties promoting tissue-specific cell interactions are vital for implant integration into the host. This amphiphilic polymer film system also demonstrated the capability to promote and control hNPC adhesion and differentiation. A copolymer containing 50% CPTEG and 50% CPH may present optimal properties for neural tissue engineering with hNPCs by limiting immune recognition while providing a conducible environment for cellular adhesion and differentiation. These amphiphilic polymers may provide an excellent platform technology for tissue engineering constructs.
Footnotes
Acknowledgments
The authors would like to acknowledge financial support from the ONR-MURI Award (NN00014-06-1-1176). The authors would also like to thank Shawn Rigby for his expertise in flow cytometry and Christian Tormos for his help with data analysis.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.