
Abstract
Neural tissue engineering focuses on development of biomaterials that could support regeneration of neurons after trauma as well as injury caused by degenerative diseases. In this work we describe novel soft alginate hydrogels, which provide an adhesive matrix for rat and human neurons and facilitate neurite outgrowth. Only soft hydrogels, prepared with sub-stoichiometric concentrations of Ca2+, Ba2+, and Sr2+ cations by cross-linking with no >10% of all potentially available gelation sites in alginate, facilitated rapid and abundant neurite outgrowth in primary neuronal monolayer cultures, neural spheroids, and neurons derived from rat and human neural stem cells. To support neurite growth, hydrogels did not require modification by any extracellular matrix components and were prepared from high as well as low viscous alginates of different origin. In addition, neurons cultured on soft hydrogels were resistant to oxidative stress injury induced by hydrogen peroxide. These findings, which apply both to rat and human neurons, go beyond the well-described role of alginates as inert materials for cell encapsulation. Such soft alginate hydrogels may be useful for the preparation of pharmaceutical compositions for prophylaxis and treatment of neurodegenerative disorders, for promoting neuronal regeneration in the peripheral and central nervous system and for neural tissue engineering applications.
Introduction
Biomaterials for tissue scaffolds include natural and synthetic materials as well as their combinations.3–6 Hydrogels, which are cross-linked networks of polymers that swell with water, became very attractive as scaffolds for soft tissue engineering because of their porous structure, tissue-like mechanical properties, and potential to attach growth-promoting molecules.7–9 Polymeric biomaterials for hydrogel formation can be either hydrophilic or hydrophobic.10,11 Polymeric networks are classified according to the mechanism of gelation: cross-linking through covalent bonds or via physical association. Physical association is achieved through (1) physical entanglements between different polymer chains, (2) lipophilic interactions between segments on polymer chains, and (3) ionic cross-linking of opposite charges of the polymers and added multivalent cations or anions.
Polysaccharides, which are present in the extracellular matrix, have been found to influence cell migration, axonal guidance, synapse development, and function.12–14 Therefore, polysaccharide-based scaffolds or scaffolds modified with polysaccharides have been tested in neural tissue engineering. Heparin-functionalized scaffolds were tested for providing topographical and chemical cues to growing neurons.15–17 Hyaluronic acid hydrogels supported differentiation of neural precursors18–20 and promoted outgrowth of neurites in vitro, 21 xyloglucan, and functionalized chitosan hydrogels supported the differentiation and growth of primary cortical neurons,22,23 and chitosan-blended agarose supported neural attachment. 24
Alginate is a natural polysaccharide, which is mainly known as an inert material applied for three-dimensional cell-encapsulation including neural stem cells.25–27 It consists of covalently linked D-mannuronic acid (M) and its C5-epimer, L-guluronic acid (G) monomers. Alginate can be categorized as natural hydrophilic, negatively charged polymer. It forms hydrogels by physical association of chains via ionic cross-linking by multivalent cations (e.g., Ca2+, Ba2+, Sr2+, Cu2+, Ni2+, Zn2+, or Fe3+), which interact with G-sequences located on different chains. 28 The proportion of M and G and their sequential compositions influence hydrogel properties and vary widely depending on alginate origin.29–31 Alginate network density within a hydrogel correlates with stiffness, which is directly influenced by the concentration of the cross-linking cation. Studies demonstrated that increase in hydrogel stiffness leads to decrease in hydrogel permeability and subsequent decrease of viability and proliferation of encapsulated neural stem cells. 26
Alginate hydrogels were shown to be nonadhesive for cultured neurons, unless functionalized with signaling molecules, such as fibronectin or polypeptides.32,33 Peptides and proteins used for alginate hydrogel functionalization are derived from the extracellular matrix and have been shown to promote neuronal adhesion and neurite outgrowth. 34 As an example, the extracellular matrix protein fibronectin is present in the developing mammalian central nervous system at areas of axon extension and promotes neuronal adhesion and neurite outgrowth in vitro and in the adult white matter. 35
Alginate scaffolds containing polyglycolic acid or heparin were able to promote peripheral nerve regeneration in vivo and supported regenerating axons in the transected rat spinal cord.17,34–37 Functional reinnervation of motor and sensory nerves occurred 13 weeks after implantation of alginate gels in transected peripheral nerves in the cat, as demonstrated by recovery of compound muscle action potentials and somatosensory evoked potentials. 36 Within the transected rat spinal cord, numerous descending regenerating fibers were found at 2 and 4 weeks after alginate implantation.37–39 At 21 weeks after injury, regenerating fibers were seen to traverse the alginate filled gap and reenter the distal stump of the cord forming functional synapses. 38
In the process of exploring conditions that promote growth of axons through alginate scaffolds, we analyzed how variable hydrogels may influence neurite growth. We found that nonfunctionalized soft alginate hydrogels, formed by gelation of alginates of distinct origin, physical state, and viscosity with solutions containing as little as 2 mM of a cross-linking cation, were able to support rapid and abundant neurite growth. Given biocompatibility of alginates, we suggest that treatments utilizing such hydrogels may open novel perspectives for neural tissue engineering applications, which may lead to new practical modalities for the therapies of acute and chronic neurodegenerative diseases.
Materials and Methods
Ultrapure sodium alginates with trade name Pronova were purchased from Novamatrix, now part of FMC Biopolymer, as guluronic acid rich alginate (LVG) or mannuronic acid rich alginates (LVM, SLM20 and SLM100), containing 68% or 43% of guluronic acid residues (G), respectively. Abbreviations used here refer to companies trade name extensions (LV low viscosity, SL sterile lyophilized, M or G main monomer unit of specific alginate). Alginate with unknown G content was purchased from Sigma (71238-50G). Reagents for cell culture, including Neurobasal cell culture medium and B-27 supplement, were purchased from Invitrogen. CaCl2, BaCl2, SrCl2, Poly-L-lysine (pll) (W 70.000–150.000), DNase I, hydrogen peroxide (H2O2) (35% stabilized solution), and Hoechst 33342, originated from Sigma. AlamarBlue® reagent (AbD Serotec) and Cytotoxicity Detection Kit (LDH) (Roche Applied Science) were used according to the manufacturer's instructions.
Preparation of alginate hydrogels for in vitro cell culture experiments
Alginate hydrogel was formed as a thin layer (∼1 mm in thickness) which covered the bottom of cell culture wells or glass coverslips. Alginate powder was dissolved in water to obtain 1% or 0.2% alginate sol, which was sterilized by filtering (0.45 μm filter). Dry alginate films were formed after drying an aliquot of aqueous alginate sol at room temperature for 12 h. Alginate hydrogel was made by cross-linking 200 μL aqueous sol or dry film derived from 200 μL aqueous sol with solutions containing CaCl2, BaCl2, or SrCl2 in water or with Neurobasal cell culture medium (1.8 mM CaCl2, 0.814 mM MgCl2, 5.33 mM KCl, 26.19 mM NaHCO3, and 0.906 mM NaH2PO4-H2O) at room temperature for 60 min. Excess of cross-linking solution was completely removed; hydrogel was washed 4 times with sterile water and once with Neurobasal cell culture medium. Finally, a fresh aliquot of complete Neurobasal cell culture medium was added and incubated for 60 min at 37°C and 5% CO2 prior to seeding of primary neural cells.
The abbreviation describing a hydrogel denominates alginate concentration in the sol (before cross-linking) and type of alginate followed by the formation route in subscript as well as the concentration and type of the cross-linking solution separated by a slash. For example, 0.2% LVMdry film/2 mM CaCl2 encodes a hydrogel formed by cross-linking of dry alginate film (formed by drying 0.2% aqueous LVM sol) with 2 mM CaCl2 solution. LVMthermo and LVGthermo encode alginates sterilized by autoclaving, LVMdry film and LVGdry film encode dry alginate film, LVMaqueous, and LVGaqueous encode aqueous alginate sol.
Preparation of neuronal cell cultures
Neuronal cell cultures were prepared from E20 Wistar rat fetuses. Cells were isolated from cortices as previously described. 40 Cell culture medium for culturing of neural cells consisted of 2% B-27 supplemented Neurobasal medium, 0.5 mM L-glutamine and 1% ampicillin, and streptomycin solution. Cells were sieved through a 40 μm cell strainer (Falcon, Becton Dickinson Labware), counted, and plated at a cell density in the range from 0.1×106 to 1×106 cells/well into six-well plates, or 0.5×105 cells/well into 24-well plates, which were coated with pll or contained alginate hydrogel. Neurite length was quantified in cell cultures with low primary cell plating density of 0.4×106 cells per ∼94.2×103 μm2 surface. Plastic plates were incubated with pll solution (10 μg per mL) for 1 h at 37°C. Unbound pll was washed out with DPBS, and plates were used on the same day.
Neural spheroid cultures were used to better evaluate neurite outgrowth and to omit cleavage of cell surface molecules with trypsin that constantly occurred during primary cell isolation from the brain. To prepare neural spheroids, single cell suspension was seeded into six-well plates, either noncoated or containing 1% LVGdry film/1000 mM CaCl2 hydrogel. The plating cell concentration was 0.5×106 viable cells per 1 mL of cell culture medium. Incubation was carried out at 37°C in a humidified 95% air and 5% CO2 atmosphere. The culture medium was refreshed every third day by replacing 20% of it.
Neural stem cells were isolated from P1 rat hippocampi. Tissue was collected and placed in ice-cold 0.6% glucose in PBS. Cells were dissociated in StemPro Accutase (Invitrogen A11105) for 20 min in a 37°C water bath. The tissue settles during this treatment allowing easy removal of the Accutase. DNAase (diluted in 0.6% glucose-PBS to 3360 U/mg) was added and the tissue was returned to 37°C for 10 more min. The tissue was allowed to settle again during this treatment and the DNase was subsequently removed. The hippocampus tissue was re-suspended in 1 mL Starting media, was triturated 50 times using a P1000 and distributed to 6-8 T-25 (Nunc) flasks containing starting media Dulbecco's modified Eagle's medium (DMEM)/F-12 (70%/30%) (DMEM 11965-092: F-12 11765-054; Invitrogen), 2% B27 Supplement, epidermal growth factor (EGF) (20 ng/mL final concentration [E9644; Sigma]), Fibroblast growth factor (FGF-2; 20 ng/mL final concentration [100-18B; Peprotech]), Heparin (H-3149-25K; 5 μg/mL final concentration; Sigma) and 1% antibiotic–antimycotic liquid (Invitrogen 15240-062). 40 Flasks were supplemented daily with FGF-2 (20 ng/mL) and EGF (20 ng/mL) and fed every 3 or 4 days by allowing the neurospheres to settle in each flask, removing half of the conditioned media and replacing with fresh media. As the neurospheres grew to an average diameter size of about 500 μm they are chopped using a McIlwain tissue chopper as described previously. 40
Human neural stem cells (hnpcM031), originally obtained from human fetal brain tissue, were a gift from Dr. Bhattacharyya, Waisman Center. These cells are also chopped using a McIlwain tissue chopper when they grew to an average size of about 500 μm. Cells were grown in Maintenance Media (DMEM/F12(70/30), 1% N2 (Invitrogen 17502-048), EGF (20 ng/mL), Leukemia inhibitory factor (LIF, 10 ng/mL final concentration; LIF1005; Millipore), and 1% PSA and fed every 3–4 days as described above. After several passages, cells were plated on Poly-Ornithine (Sigma P4638)/laminin (Sigma L2020)-coated coverslips (1943-10012; Bellco) (30,000 cells/coverslip), which had been placed in 24-well plates (Nunc). The cells were allowed to attach for 2–3 h and then overlaid with media (DMEM/F12 2% B27 Supplement with 1% heat-inactivated fetal bovine serum (Invitrogen 10082-147) containing alginate and allowed to differentiate for 1–10 days. The alginate (Sigma 71238-50G) was dissolved in MilliQ water, and then diluted in media to the appropriate concentration. Neural stem cells were exposed to alginate concentrations of 0.05% or 0.1% in differentiating media. Neuronal identity of analyzed cells derived from neural stem cells was confirmed by immunostaining for MAP2 or β-tubulin III markers.
Induction of oxidative stress to neuronal cell cultures
Primary neurons were seeded into 24-well plates with 1 mL of complete Neurobasal cell culture medium (phenol red free), cultured for 14 days either in pll-coated wells or containing 0.2% LVMdry film/Neurobasal medium hydrogel. H2O2 solution was applied into cell culture medium to a final concentration of 50, 100, or 150 μM and neurons cultured in the presence of H2O2 for additional 16 h. Untreated control cells were incubated in fresh aliquot of complete cell culture medium, or in the presence of 1% Triton X-100 in medium. At least six repeats were done for every condition; five independent dissections for preparation of primary neuronal cultures were performed. Neuronal viability or damage was measured with AlamarBlue reagent or LDH Cytotoxicity Detection Kit.
Cytotoxicity assay (lactate dehydrogenase) and cell viability assay AlamarBlue
Lactate dehydrogenase (LDH) is a stable cytoplasmic enzyme present in all cell types and is released from damaged cells. LDH activity was determined in medium and absorbance was measured at 492 nm in ELISA reader (Tecan). AlamarBlue is a dye which changes its color in response to chemical reduction of growth medium resulting from living cell metabolism; neurons were cultured in the presence of AlamarBlue for 5 h and absorbance was measured at 570 nm in ELISA reader (Tecan).
Immunocytochemistry
Primary monoclonal mouse anti-β-tubulin III (R&D Systems, Minneapolis) and mouse anti-MAP2 (Chemicon/Millipore) were used; secondary antibodies were Alexa-488 labeled chicken anti-mouse (Molecular Probes/Invitrogen). Neurons, which were cultured on pll-coated glass coverslips or on alginate hydrogels, were fixed with acetone/methanol mixture (1:1 volume) for 10 min at−20°C and then labeled with antibody according to a standard procedure. Hoechst 33342 (10 μg/mL, in TBS) was used to label nuclei. Samples were covered with Aqua PolyMount (Polysciences, Inc.), and examined under a fluorescence microscope (Axiovert 40, Zeiss).
Rheological characterization of alginates and hydrogels
Rheological measurements were carried out using Anton-Paar Physica MCR301 instrument. The viscosity values of aqueous alginate solutions were recorded under continuously logarithmically increasing shear in a double slit chamber DG26.7 at 20°C. An ideal viscous behavior was assumed for shear rates in the domain of 0.1–100/s and the constant viscosity was interpolated. High values of the intrinsic viscosity indicate high molar mass of a polymer. Subsequently, 1% alginate solutions of LVG, LVGthermo, LVM, and LVMthermo were tested for their viscosity values.
Stress sweep at a constant frequency of 1 Hz was performed to obtain values of elastic (G’) modus of alginate hydrogels; viscosity values were recorded in parallel-plate geometry measuring cell PP25 at 37°C under logarithmically increasing amplitude (0.001–100 Pa). Alginate solutions were poured on top of the bottom plate followed by the addition of CaCl2 solution, hydrogel was cut in the shape of a disk, excess of the cross-linking solution was removed, and samples were equilibrated at 37°C for 20 min before measurements.
Measurements of alginate hydrogel swelling
A preset volume (600 μL) of 1% aqueous alginate sol was distributed over a glass coverslip (24×24 mm) located in a petri dish (Ø 5 cm) and then overlayed with 15 mL of a cross-linking solution, containing either 2, 10, or 100 mM of CaCl2 in water. Cross-linking reaction was performed at room temperature for 24 h and hydrogel was formed on a glass surface as a single slice. To preserve hydrogel integrity, the weight of a glass coverslip together with the hydrogel was measured, subsequently the average weight of a glass coverslip (0.22 g) was subtracted. Three independent hydrogels were prepared for every test condition.
Alginate sol consists of a liquid phase and randomly distributed negatively charged alginate fibers. Within a hydrogel, fibers are cross-linked and the liquid phase contributes 95% of its mass. Hydrogel mass either decreases (distance between fibers becomes smaller), increases (equally charged fibers are pushed apart; meanwhile, some become connected via ion bridges), or remain unchanged in comparison to noncross-linked sol. The rate of swelling was estimated after comparison of the hydrogel mass with the mass of the alginate sol (taken as 100%).
Quantification of neurite length and statistical analysis
Neurite length was quantified on live phase contrast or fluorescence images using NeuronJ (www.imagescience.org/meijering/software/neuronj). Average neurite length (μm) on 1000 μm2 surface was measured on at least five independent images, on at least 2×106 μm2. Student's t-test for unpaired observations was applied for statistical analysis. Differences between samples with p-value<0.05 were accepted as statistically significant.
Results
Ca2+-cross-linked alginates form a matrix accessible for neurites
Alginate hydrogels were prepared using 1000 mM CaCl2 and 1% aqueous sol. Primary cortical neurons did not attach to the surface of 1% LVMaqueous/1000 mM CaCl2 hydrogel (Fig. 1a), but re-aggregated shortly after seeding onto the hydrogels forming spheroids, whereas cells attached to pll-coated plates forming cell monolayers (Fig. 1d).
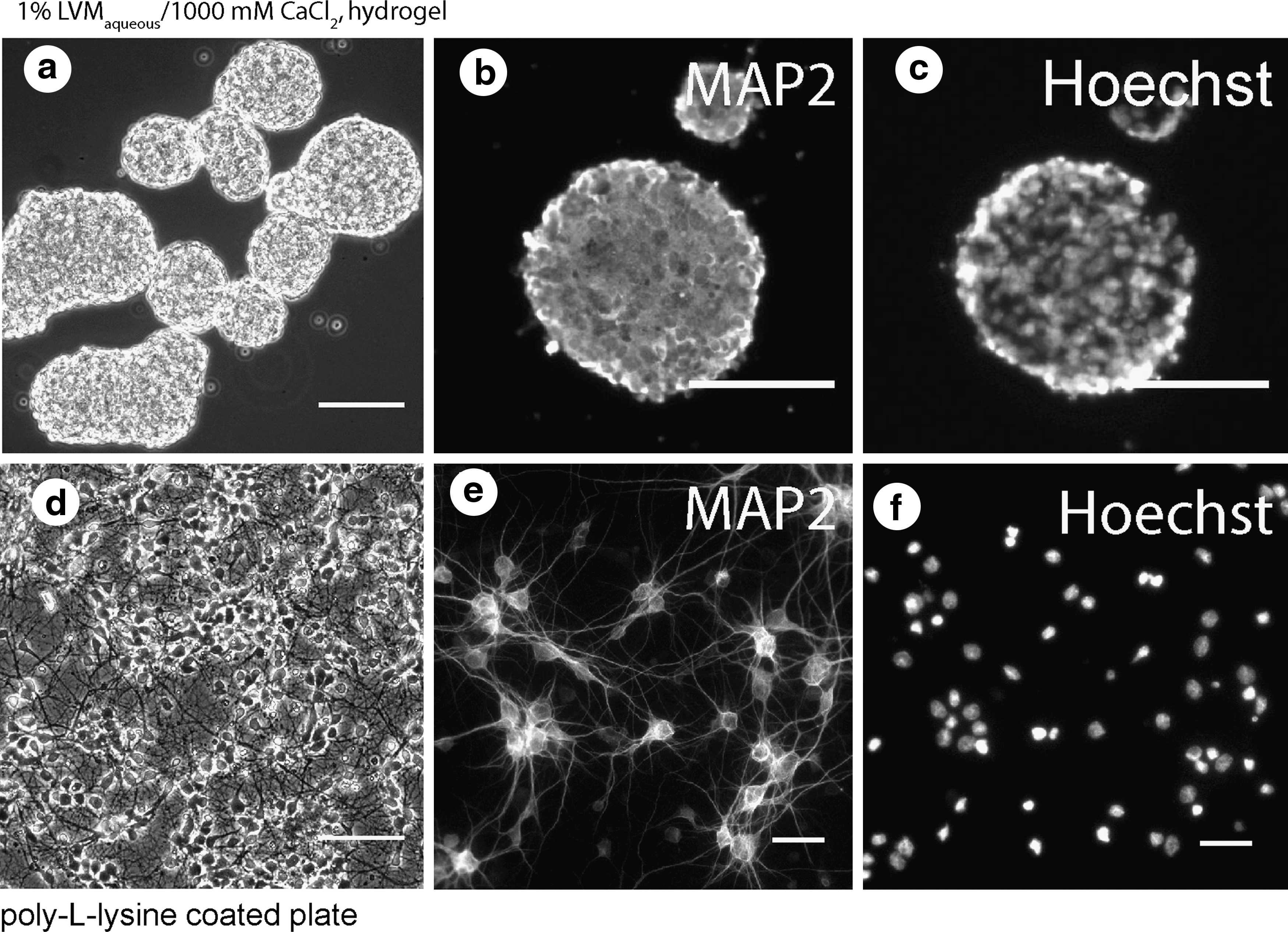
Live
Despite the nonaccessibility of 1% LVMaqueous/1000 mM CaCl2 for neurites, we continued to explore alginate potency in providing a matrix for neuronal growth. Variable hydrogels were prepared by decreasing CaCl2 in the cross-linking solution until the solution was able to form a stable, not rapidly disintegrating hydrogel; CaCl2 concentrations ranged from 1000 to 2 mM; alginates of distinct types, viscosity, and physical state (sol and dry film) were cross-linked. There was no detectable growth of neurites on 1% LVM/100 mM CaCl2 (Fig. 2c, f), neither on 1% LVM/10 mM CaCl2 hydrogels regardless of alginate type (data with SL20, SL100, Sigma-alginate are not shown) and physical state (sol and dry film). On these hydrogels, plated neurons were unable to form adherent monolayer cultures but formed spheroids instead (Fig. 2c, f).
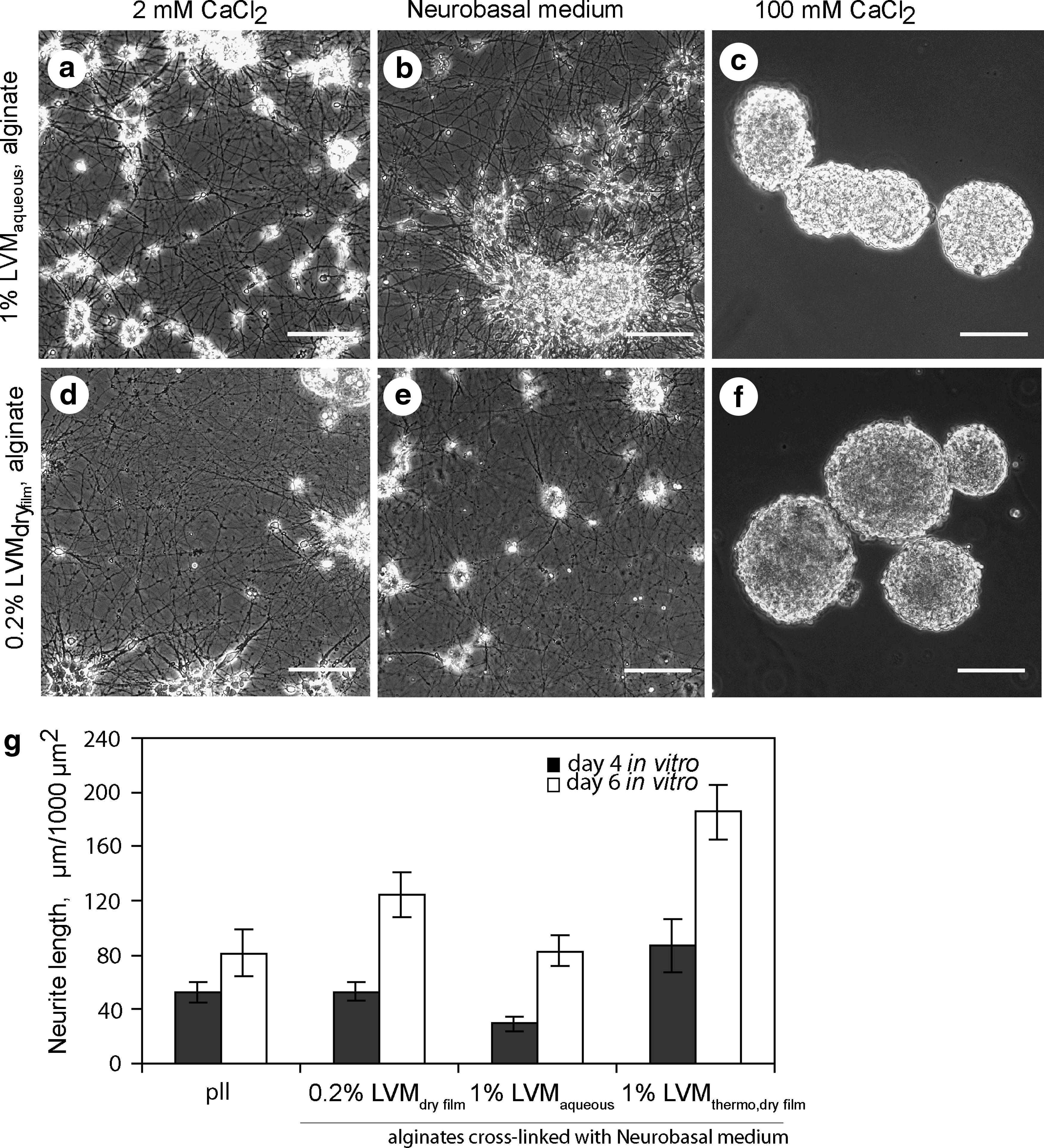
Live images illustrate that neurons form a neurite meshwork (day 4 in vitro) on 1% LVMaqueous or 0.2% LVMdry film alginate hydrogels, prepared with 2 mM CaCl2
To our surprise, there was massive neurite growth noted on 1% LVMaqueous/2 mM CaCl2, 1% LVMaqueous/Neurobasal medium, 0.2% LVMdry film/2 mM CaCl2, 0.2% LVMdry film/Neurobasal medium hydrogels (Fig. 2a, b, d, e). Neurites spread on all hydrogels derived from aqueous sol or dry film of distinct alginate types (as listed in Materials and Methods), which were formed with addition of 2 mM CaCl2 solution or Neurobasal medium (alginate concentration in aqueous sol was 0.2% or 1%). Similar to neurons on pll-plates (Fig. 1d), neurons on the hydrogels, exhibited typical neuronal morphology, that is, cell bodies surrounded by a neurite meshwork, and expressed molecules such as MAP2 and β-tubulin III. Neurites penetrated the entire hydrogel volume (up to 1000 μm), creating a three-dimensional meshwork. Neurite abundance varied between the hydrogels and depended upon the type of alginate used. Average neurite length within an area of 1000 μm2 measured in representative cell cultures corresponded to 50±12, 29±10, 46±17, 42±12, and 39±15 μm on LVM, LVG, SL20, SL100 and Sigma-alginate hydrogels (from 1% aqueous sols), to 53±9, 36±9 μm on LVM and LVG hydrogels (from dry films), and to 53±12 μm on pll-coated plates, respectively. Most prominent neurite growth was consistently detected on LVM-based hydrogels. In addition, rate of neurite extension was higher on LVM-based hydrogels in comparison to pll-coated plates; for example, an increase in average neurite length, measured between day 4 and 6 in vitro, was 155% on pll-coated plate but 235% on 0.2% LVMdry film, 283% on 1% LVMaqueous, and 212% on 1% LVMthermo, dry film alginate hydrogels (Fig. 2g).
Alginate viscosity is proportional to polysaccharide chain length; high viscous alginates form mechanically more stable hydrogels than low viscous, predominantly because of higher probability for cross-linking of the polysaccharide chains by multivalent cations and physical entanglement. 41 Alginate viscosity decreases during polysaccharide chain cleavage, for instance during autoclaving. 42 Although autoclaving caused a decrease in alginate viscosity from 145 to 27 mPas for LVG, and from 21 to 6 mPas for LVM respectively, 1% LVGthermo,dry film/Neurobasal medium and 1% LVMthermo,dry film/Neurobasal medium supported abundant neurite growth, similar to hydrogels prepared from their native counterparts (Figs. 2g and 4c). Thus, low alginate viscosity does not affect hydrogels' ability to support neurite growth.
Neural cultures with different primary cell seeding densities (from 0.1×106 to 1×106 cells per 94.2×103 μm2) were studied on pll-covered plates, on neurite-supportive 1% LVMdry film/Neurobasal medium and 1% LVGdry film/Neurobasal as well as on neurite nonpermissive 1% LVMdry film/100 mM CaCl2 and 1% LVGdry film/100 mM CaCl2 hydrogels. Cell seeding density directly influenced density of the resulting neurite meshwork (evaluated on day 7 in vitro) but not the principal ability of the respective hydrogel to support or inhibit neurite elongation.
Ba2+-, Sr2+-cross-linked alginates support neurite outgrowth
Alginate hydrogels could be formed in the presence of cations other than Ca2+, that is, Ba2+ and Sr2+. Cross-linking of LVM or LVG alginates with solutions containing 10 or 100 mM of BaCl2 or SrCl2 formed hydrogels that were nonaccessible for neurites (Fig. 3). Similar to Ca2+-alginate hydrogels, alginates cross-linked with 2 mM solution of either BaCl2 or SrCl2 supported abundant neurite growth. Subsequently, average neurite length measured on day 4 in vitro within an area of 1000 μm2 was 145±30 μm on Ca2+-LVM, 107±20 μm on Ba2+-LVM and 109±21 μm on Sr2+-LVM hydrogels. These hydrogels were formed from 1% LVM alginate sol crosslinked with 2 mM solution of the respective salt (CaCl2, BaCl2 or SrCl2). LVG based hydrogels were much less permissive for neurites. Average neurite length on day 4 in vitro within an area of 1000 μm2 was 19±9 on Ca2+-LVG, 51±13 on Ba2+-LVG, and 45±15 on Sr2+-LVG, respectively. These hydrogels were formed from 1% LVG alginate sol crosslinked with 2 mM solution of the respective salt (CaCl2, BaCl2, or SrCl2). Thus, the cation that connects alginate chains forming an alginate network does not affect the hydrogels' ability to provide an adhesive matrix for neurons and support neurite growth.

Live images depict neurons form neurite meshwork (day 4 in vitro) on hydrogels prepared with 2 mM solution
Neural spheroids rapidly extend neurites upon contact with the hydrogel
Spheroids formed on the top of hydrogels which were gelled by concentrations of cations above 10 mM. These hydrogels did not support neurite growth. Spheroids were comprised of viable neural cells. Vast majority of cells inside these spheroids, similar to cells cultured on pll-plates in monolayer neural cultures, excluded trypan blue in a dye exclusion test, contained nonfragmented nuclei and expressed the neural marker MAP2 (Fig. 1a–c). Spheroids could be maintained for many weeks in vitro, they gradually enlarged in size (mainly due to fusion of multiple spheroids with each other) as their cross-section area increased from 15±8×103 μm2 (day 7) to up 220×103 μm2 (day 21). Notably, neural spheroids exhibit properties that are remarkably similar to their in vivo counterparts. 43
We questioned whether neurite outgrowth from such spheroids could be induced and facilitated by contact with the appropriate alginate hydrogels. Therefore, spheroids formed on 1% LVMaqueous/100 mM CaCl2 were plated onto 1% LVMaqueous/2 mM CaCl2 or 1% LVMdried film/2 mM CaCl2, or 1% LVMdried film, thermo/Neurobasal medium hydrogels on day 7 in vitro and examined 24 h later. Already 3 h after plating onto the hydrogels, multiple neurites with clearly defined growth cones extended in all directions and away from spheroids, reaching up to 600 μm in length (Fig. 4b, c). Neurites predominantly extended from the bottom of the spheroids, which is the area of the spheroid that is in direct contact with the underlying hydrogel. In contrast to the alginate hydrogels, only few neurites grew from spheroids placed on pll-coated plates (Fig. 4a). Spheroid-derived neurites expressed the neuronal markers MAP2 and β-tubulin III (Fig. 4d, e). Dynamic behavior of growth cones influences neurite morphology; growth cone migration elongates a neurite, while growth cone splitting creates a branch point. We observed that neurite elongation prevailed over neurite splitting on alginate hydrogels. Majority of neurites which extended on alginate hydrogels from spheroids or from neurons in adherent cell cultures did not branch; <1% of all neurites contained a single branching point. In contrast, neurons cultured on pll extended up to 10 neurites, every neurite contained up to three branching points.

Live images show neural spheroids cultured for 24 h on pll-coated plates
We also explored neuritogenic potential of alginates in rat and human neurons which differentiated from neural stem cells in vitro. Differentiated neurons cultured in the presence of 0.05% or 0.1% alginate in the culture medium extended longer neurites within 24 h compared to those grown in differentiating media alone (Fig. 5b, d, e, f). In rat and human neural stem cell cultures, there was a significant effect of alginate on neurite growth.
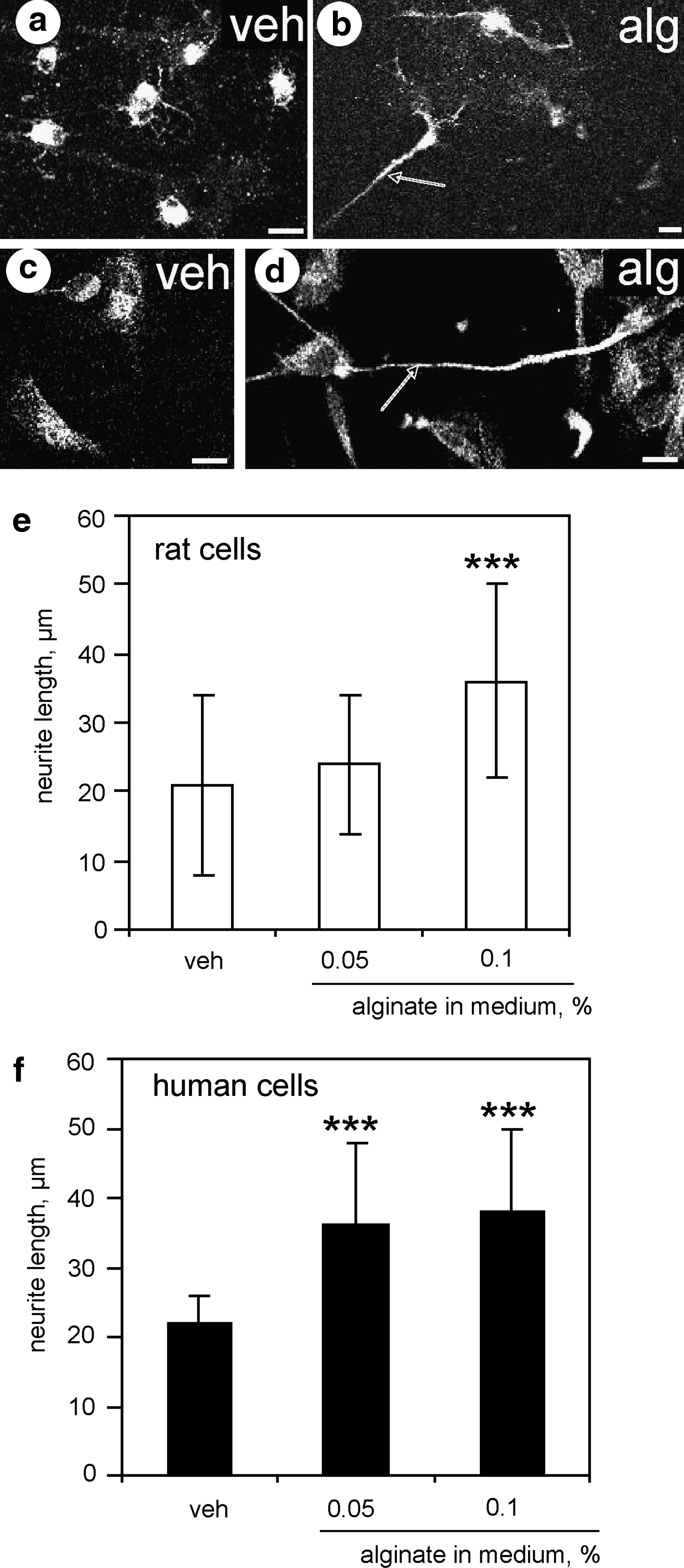
Fluorescence confocal images of rat and human neural stem cells grown on laminin coated plates in differentiating medium in the presence of vehicle
Of note is that astroglial cells were not detected in primary neuronal cultures grown on alginate hydrogels, as revealed by the absence of glial fibrillary acidic protein immunopositive cells on day 7 in vitro (data not shown). In the cultures derived from rat or human neural stem cells, 60%–70% of the cells were immunopositive for the specific neuronal markers MAP2 or β-tubulin III. It is unlikely that neurite growth in the presence of alginate was promoted by astrocytes or astrocyte-derived molecules.
Stiffness and alginate network density within hydrogels, swelling
Next, we measured stiffness of 1% LVGaqueous/1000 mM CaCl2 and 1% LVGaqueous/2 mM CaCl2 hydrogels by carrying out oscillatory shear measurements in a rheometer. The elastic modulus of hydrogels formed using 1% aqueous alginate solution decreased with decreasing CaCl2 concentrations in the cross-linking solution and was 20.8 kPa for stiff 1% LVGaqueous/1000 mM CaCl2 and only ∼0.64 kPa for soft 1% LVGaqueous/2 mM CaCl2. This dramatic change in mechanical behavior could be attributed to a higher cross-linking density in stiff 1% LVGaqueous/1000 mM CaCl2 alginates. Interestingly, the elastic modus of soft, neurite-supportive alginate hydrogels was close to that of brain tissue (0.1–1 kPa).44,45
Two G-residues in an alginate sol, which are connected by a divalent cation (for example Ca2+), form a gelation site. The more gelation sites are occupied, the denser the alginate network within a hydrogel will be. LVM and LVG alginates contain 43% and 68% of G-residues in their primary structure, respectively; thus, in 1% alginate solution, 0.22% and 0.34% solution of CaCl2 will approximately saturate all potentially available gelation sites in LVM and LVG, respectively. Subsequently, in hydrogels formed in the presence of 1000 mM CaCl2, almost all potential gelation sites are saturated. However, in 1% LVMaqueous/2 mM CaCl2 and in 1% LVGaqueous/2 mM CaCl2 hydrogels only ∼10% and ∼7% of potential gelation sites are occupied.
The process of liquid absorption by a polymer is called swelling, and could be estimated by measuring of hydrogel mass. We found that mass of hyrogels that are nonpermissive for neurites increased with decreasing CaCl2 concentrations (p<0.05, 10 vs. 100 mM CaCl2). For example, average weight of 1% LVGaqueous/100 mM CaCl2 and 1% LVGaqueous/10 mM CaCl2 was 0.48 and 0.51 g, respectively, 1% LVMaqueous/100 mM CaCl2 and 1% LVMaqueous/10 mM CaCl2 was 0.52 and 0.55 g, respectively; corresponding degrees of swelling were 80%, 84%, 87%, 92%. Mass of neurite-permissive hydrogels, that is, 1% LVGaqueous/2 mM CaCl2 and 1% LVMaqueous/2 mM CaCl2, was 1.56 and 2.2 g, respectively; corresponding degrees of swelling were 261% and 366%. These data show that neurite-permissive hydrogels swell very strongly in comparison to nonpermissive hydrogels. Hydrogels can change their volume by loosing or gaining a liquid phase or undergo disintegration after sequestration of cross-linking cations. Aginate hydrogels used for culturing of cells in this study were not disintegrated in cell culture medium.
Alginate protects neurons from H2O2 induced oxidative stress
In the course of our experiments, we constantly observed that neurons survived longer on the alginate hydrogels (up to 4 months) than equivalent neuronal cultures grown on pll-coated plates, which were able to survive for a maximum of 4 weeks. Given the observed neuritogenic properties of nonfunctionalized alginate hydrogels, we questioned whether these same hydrogels might also exert trophic effects on neurons and influence cellular vulnerability to oxidative stress. Oxidative insult in vitro was induced by application of H2O2 to neurons at concentrations ranging from 50 to 150 μM. H2O2 caused a concentration-dependent release of LDH from exposed injured neurons (Fig. 6a, dark columns). Neurons cultured on 0.2% LVMdry film/Neurobasal medium hydrogels released significantly less LDH into the culture medium in comparison to neurons grown on pll when exposed to the same concentrations of H2O2 (Fig. 6a, white columns). This indicates that neurons grown on alginate hydrogels were less likely to suffer injury when exposed to oxidative stress induced by H2O2. Triton X-100 caused maximal release of LDH activity which was above 600% in comparison to controls (medium alone) for both cell culture conditions; this result demonstrates that detection of LDH activity by the used assay was not altered in alginate hydrogels as compared to control conditions. Further, neuronal viability was quantified using Alamar Blue. In accordance with the LDH release results, neurons cultured on alginate hydrogels (Fig. 6b, white columns) demonstrated higher viability after exposure to H2O2 than neurons grown on pll-coated plates (Fig. 6b, dark columns).

Neurons cultured on 0.2% LVMdry film/Neurobasal medium are less vulnerable to hydrogen peroxide (H2O2)-induced oxidative stress.
Neurons cultured on pll-plates released 215%±25% LDH in the presence of H2O2 solution (100 μM) which had been preincubated with alginate and 237%±35% of LDH activity in the presence of H2O2 solution (100 μM) without alginate preincubation. These results demonstrate that alginate hydrogel does not decrease neurotoxicity of H2O2 solution but rather diminishes neuronal vulnerability to oxidative stress.
Discussion
In the context of this study we explored how alginate hydrogels of variable stiffness could support outgrowth of neurites in vitro. We came across a novel observation that nonfunctionalized, soft alginate hydrogels, formed by cross-linking with Ca2+, Ba2+, or Sr2+, supported rapid and abundant neurite growth from neurons in primary rat neuronal cultures. Neurons derived from rat and human neural stem cells extended longer neurites in the presence of soluble alginate in the cell culture medium. In addition, we report that these soft hydrogels, which promote neurite outgrowth, also exhibit neuroprotective properties against oxidative stress induced by H2O2.
The novelty of our findings lies in the discovery that a soft, nonfunctionalized alginate hydrogel has the ability to support neural adhesion, rapid and abundant neurite outgrowth on its surface as well as within the hydrogel. To our knowledge, this is the first report that mechanically stable and soft alginate hydrogel does not require modifications with cell adhesion promoting molecules or with pll to provide an adhesive surface to neurites. Given biocompatibility of alginates, these observations open up possibilities for the use of nonfunctionalized alginate hydrogels for neural tissue repair/regeneration but also neuroprotection.
In agreement with previously published reports describing nonfunctionalized alginate hydrogels as nonaccessible for neurites, we observed that neurons neither attach nor spread neurites on top of stiff alginate hydrogels, but form spheroids instead. It is known that alginate network density within a hydrogel is regulated by the strength of cross-linking. Indeed, hydrogel accessibility to neurites depended strongly upon the concentration of the cross-linking cation used and minimally upon cation type (Ca2+, Ba2+, Sr2+), alginate type, viscosity, concentration, physical state or the type of neural cell culture (monolayer or spheroids).
Almost a decade ago pioneering work by Pelham and Wang demonstrated that fibroblast spreading and migration are greater on stiffer compared to more compliant polyacrylamide hydrogels. 46 Since then, physical cues, such as substrate stiffness, are recognizable modifiers of cell behavior. Reports demonstrated that the mechanical properties of the substrate may direct the formation of neurites and drive neuronal differentiation from neural stem cells.19,44 In this work, we demonstrated that soft hydrogels were favored by neurons, whose extended neurites were penetrating the entire hydrogel volume.
Neurite growth might be partly driven by unique physicochemical properties of underlying soft alginate hydrogels. Stiffness of neurite-supportive and nonpermissive alginate hydrogel was 0.64 and 20.8 kPa, respectively. Therefore, we conclude that neurite extension was specific only to soft but not stiff alginate hydrogels. Our results are in agreement with previously published reports which demonstrated that neurons favor soft rather than stiff substrates. Mice spinal cord neurons formed three times more branches on hydrogels with elastic moduli ranging from 0.05 to 0.3 kPa than with 0.55 kPa (dyn/cm2 were converted into kPa). 44 The rate of chick dorsal root ganglion neurite extension was inversely related to mechanical stiffness. 47 Primary neural stem cells differentiate into neurons on soft methacrylamide chitosan hydrogel with stiffness<1 kPa, 48 on polyacrylamide and polyethylene glycol hydrogels with stiffness ranging from 0.1 to 0.5 kPa. 49 Proliferation of encapsulated neural stem cells decreases with increasing alginate hydrogel stiffness from 0.1 to 10 kPa. 26 Published data and our results suggest that, within the same material, neurons prefer soft rather than stiff surfaces, but the exact mechanism for this behavior is not yet known.
In soft hydrogels, with gelation of no >10% of gelation sites, multiple polysaccharide sequences remain uninvolved in the cross-linking reaction and can potentially interact with cell surface molecules. Subsequently, digestion of cell surface molecules would attenuate alginate-neuron interaction. Of note is that we constantly detected a delay (48 h) in the initiation of neurite growth from neurons which had been treated with trypsin, but rapid (3 h) neurite extension from neural spheroids and neural stem cells not treated with trypsin. Thus, alginate polysaccharide sequences might mimic functional groups within the extracellular matrix of the brain, which were found to modulate signal transduction cascades that guide cell migration and neurite growth. 50
Alginates conferred protection to neurons against oxidative stress induced by H2O2. Oxidative stress occurs as a result of an imbalance between the production of reactive oxygen species and the cell's capacity to neutralize them through intrinsic antioxidant defenses. 51 Recently, alginate added to cell culture medium was shown to protect an NT2 cell line against H2O2-induced toxicity. 52 Thus, alginate microenvironment might enhance neuronal resistance to oxidative stress via stimulation of cellular intrinsic antioxidant defenses.
We wish to stress that we cultured neurons under serum-free conditions and in the absence of common extracellular matrix components. Under these conditions, soft alginate hydrogels exert neuritogenic and neuroprotective effects. There is a limited number of other natural and synthetic biocompatible polymers for nerve repair10,11 and all these biomaterials were either (1) applied to solely provide inert mechanical support while neurite extension was driven by externally introduced signaling molecules including laminin, fibronectin, collagen, pll, or gelatin, 53 or (2) used as carriers for neurotrophic factors, encapsulated stem cells or Schwann cells.10,11 Since growth factors, serum 47 or extracellular matrix components were always used when culturing neurons on polysaccharides-based hydrogels, it remains unknown from the existing literature what mechanism could mediate the trophic effects of alginates observed in our studies.
Identification of the molecular mechanisms facilitating neural growth on nonmodified soft alginate-based matrices could provide valuable information for biomaterial sciences and for regenerative medicine.
Footnotes
Acknowledgments
The authors thank the German Research Foundation (DFG) for funding of this study (project numbers IK 2/7-2 and GE 1133/4-2). The authors wish to acknowledge the assistance by rheological measurements of Fabian Luthardt from The Fraunhofer Institute for Ceramic Technologies and Systems IKTS, Dresden, Germany.
Disclosure Statement
No competing financial interests exist.