
Abstract
Using a complete spinal cord transection model, the present study employed a combinatorial strategy comprising rat bone marrow stem cells (rBMSCs) and polymer scaffolds to regenerate neurological function after spinal cord injury (SCI) of different lengths. SCI models with completely transected lesions were prepared by surgical removal of 1 mm (SC1) or 3 mm (SC3) lengths of spinal cord in the eighth-to-ninth spinal vertebrae, a procedure that resulted in bilateral hindlimb paralysis. A cylindrical poly(D,L-lactide-co-glycolide)/small intestinal submucosa scaffold 1 or 3 mm in length with or without rBMSCs was fitted into the completely transected lesion. Rats in SC1 and SC3 groups implanted with rBMSC-containing scaffolds received Basso–Beattie–Bresnahan scores for hindlimb locomotion of 15 and 8, respectively, compared with ∼3 for control rats in SC1-C and SC3-C groups implanted with scaffolds lacking rBMSCs. The amplitude of motor-evoked potentials recorded in the hindlimb area of the sensorimotor cortex after stimulation of the injured spinal cord averaged ∼100 μV in SC1-C and 10–50 μV in SC3-C groups at 4 weeks, and then declined to nearly zero at 8 weeks. In contrast, the amplitude of motor-evoked potentials increased from ∼300 to 350 μV between 4 and 8 weeks in SC1 rats and from ∼200 to ∼250 μV in SC3 rats. These results demonstrate functional recovery in rBMSC-transplanted rats, especially those with smaller defects. Immunohistochemically stained sections of the injury site showed clear evidence for axonal regeneration only in rBMSC-transplanted SC1 and SC3 models. In addition, rBMSCs were detected at the implanted site 4 and 8 weeks after transplantation, indicating cell survival in SCI. Collectively, our results indicate that therapeutic rBMSCs in a poly(D,L-lactide-co-glycolide)/small intestinal submucosa scaffold induced nerve regeneration in a complete spinal cord transection model and showed that functional recovery further depended on defect length.
Introduction
Recent progress in stem cell engineering offers potential alternative treatment strategies for SCI patients.5,6 Stem cells implanted within the injured site may act as a source of regenerating neurons as well as a substrate that promotes and supports axonal growth toward the damaged neural tissue. A variety of therapeutic cell sources, including embryonic stem cells, adult stem cells, neural stem cells, Schwann cells, and olfactory-ensheathing cells, have been shown to produce axon regeneration after implantation and to improve locomotor functional in animal models after SCI. 7 However, only small numbers of implanted therapeutic cells survive in the injured site for a long period after transplantation in SCI models.
To extend the survival period, researchers have employed various biocompatible polymer scaffolds in SCI repair strategies.8–12 Recent studies demonstrated that infusion of neurotrophic factors, including human brain-derived neurotropic factor (BDNF), plays a significant role in enhancing neuronal survival and axonal growth in the SCI models.13,14 Thus, implantation of a combination of a polymer scaffold, BDNF, and a therapeutic cell source can bridge the injury site, guide axonal growth, and function as a therapeutic cell delivery mechanism. Such combinatorial strategies have proven effective in animal SCI models.
In selecting the appropriate SCI repair model, it is important to differentiate between the recovery of locomotion through genuine axonal regeneration and that due to self-recovery.15,16 Several types of spinal cord transection models have been used to compare the effects of treatments designed to promote axon regeneration. Very significant insights into the mechanisms underlying the recovery of locomotion after SCI have been obtained using the partial and incomplete spinal cord transection model. 17 However, animal models of incomplete spinal cord transection cannot predict true axonal regeneration due to high rates of self-recovery. Moreover, little attention has been paid to the length of the defect in SCI models, despite an awareness that SCI length is a critical determinant of functional recovery after a complete injury. 18 As an alternative approach, an anatomically complete transection injury model lends itself to studying lesions of different lengths, and may more clearly demonstrate whether the proposed treatment strategies induce true axonal regeneration.
Recently, we reported the potentially favorable properties of poly(D,L-lactide-co-glycolide)/small intestinal submucosa (PLGA/SIS)–based scaffold in tissue-engineering applications.19–23 The overall aim of the current study was to advance a treatment strategy for spinal cord regeneration using rat bone marrow stem cells (rBMSCs) and PLGA/SIS scaffold in the presence of BDNF. To more accurately gauge axonal regeneration and test the effects of degree of damage on recovery, we used a complete spinal cord transection model with different length of injured spinal cord. Functional improvement was evaluated using behavioral tests and electrophysiological measurements. The results obtained using this complete spinal cord transection model suggest the feasibility of therapeutic cell delivery using a three-dimensional PLGA/SIS scaffold.
Experimental
Preparation and characterization of PLGA/SIS scaffold
Ice particles were prepared by spraying deionized water into liquid nitrogen, and particles of 180–250 μm were selected by sieving. PLGA (molecular weight 90,000 g/mole, 75:25 by mole ratio of lactide to glycolide; Resomer RG756, Boehringer Ingelheim) and SIS powder 23 (20 wt% for PLGA) (PLGA/SIS) was prepared by dissolution in methylene chloride. The solution was cooled to −20°C, the above-prepared ice particles were added (90 wt% for PLGA and SIS), and the dispersion was gently vortexed and poured into a precooled silicon mold (3 mm diameter×5 cm length). The mold containing the dispersion was frozen at −78°C for 2 days and then freeze-dried at −80°C to remove the methylene chloride and ice particles, leaving the desired porous scaffolds behind. The PLGA/SIS was sterilized using 30 kGy gamma irradiation (Atomic Energy Research Institute). Scanning electron microscopy (S-2250N; Hitachi) was used to examine the morphology of the generated scaffolds. Each scaffold was mounted on a metal stub and coated with a thin layer of platinum using a plasma-sputtering apparatus (K575; Emitech) under an argon atmosphere. A porosimeter (Auto Pore IV 9520 V 1.03; Instruments Co.) was used to examine the porosity and mean pore diameter of the PLGA/SIS scaffold.
rBMSC isolation
rBMSCs were harvested from the femurs and tibias of 5-week-old female Fischer rats. Briefly, the rats were subjected to ether euthanasia and the bones of the hind limbs were aseptically excised. The soft tissues were removed, and the femurs and tibias were placed in 50 mL Dulbecco's modified Eagle's medium (DMEM). The proximal end of each femur and the distal end of each tibia were removed using sterile scissors. A hole was then created in the knee joint end of each bone using a 26-gauge needle, and phosphate-buffered saline (PBS; pH 7.4) was used to flush the marrow from the shaft. The flushed marrow was fully suspended in PBS, and the cell suspensions from all bones were combined and centrifuged at 400 g for 5 min. The resulting pellet was resuspended in fresh DMEM supplemented with 10% fetal bovine serum (Gibco Invitrogen), 100 U/mL penicillin, and 100 μg/mL streptomycin, and seeded into tissue culture flasks at 1×105 cells/flask. After 2 days of expansion, the cultures were rinsed three times with PBS to remove nonadherent cells. The medium was exchanged every 2 days throughout the studies. For characterization, rBMSCs were washed with PBS and incubated with fluorescent-conjugated antibodies for CD29, CD44, CD34, and CD45 (BD PharMingen) in PBS (supplemented with 1% bovine serum albumin [BSA] in pH 7.5 PBS) for 30 min on ice, then washed with PBS, and fixed in 1% paraformaldehyde (R & D system) solution. Cells were analyzed using a FACScan (Becton Dickinson) flow cytometry system. Cell analysis was performed using at least 10,000 events per sample. Data acquisition and analysis were then performed by Becton Dickinson cell quest software. The rBMSCs showed positive expression of mesenchymal stem cell markers CD44 (98.4%) and negative expression of hematopoietic stem cell markers CD45 (0.7%). The rBMSCs showed positive immunostaining in CD 29 and CD44, and negative in CD34 and CD45 (Supplementary Fig. S1; Supplementary Data are available online at www.liebertonline.com/tea).
For use in implant experiments, adherent cells were rinsed thoroughly with PBS and then detached by trypsinization. To track grafted cells in vivo, rBMSCs were labeled before transplantation using the PKH67 Fluorescent Cell Linker Kit (Sigma) according to the manufacturer's instructions. In brief, cultured rBMSCs were washed with serum-free media and centrifuged for 5 min at 400 g. The provided diluent C (1 mL) was added to 2×106 rBMSCs and immediately mixed with 1 mL of PKH67 stock solution (4×10–6 M) in diluent C. After incubation for 5 min at room temperature with gentle tapping, 2 mL of fetal bovine serum was added and samples were incubated for 1 min to stop the labeling reaction. Finally, rBMSCs were centrifuged for 5 min at 400 g, transferred to a fresh tube, and washed three times with complete DMEM.
For SCI experiments, PLGA/SIS scaffolds with the specified dimensions (10×5×1 mm and 10×5×3 mm) were prepared, placed individually into 60×15 mm dishes (BD Bioscience), and prewetted with 200 μL PBS for 1 h. Then, 5 μL PBS, 2×106 labeled rBMSCs, and 200 ng BDNF (R & D Systems, Inc.) were seeded into a PLGA/SIS scaffold and incubated for 24 h (Supplementary Fig. S2).
Surgical procedures for SCI experiment
All adult female Fischer rats were housed in sterilized cages with sterile food and water and filtered air, and were handled in a laminar flow hood using aseptic techniques. All animal treatment and surgical procedures followed protocols approved by the Korea Research Institute of Chemical Technology's Council on Animal Care Guidelines. Rats were anesthetized using ketamine and rompun (1:1 ratio, 1.5 mL/kg), and were artificially ventilated with inhaled oxygen during surgery. Surgery was carried out on a 37°C heating pad. The skin and muscle were opened on the back, and micro surgical scissors were used to remove the bone and expose the eighth-to-ninth (T8–T9) spinal vertebrae. A laminectomy was performed under a dissecting microscope by complete transection of T8–T9 after creating a 1- or 3-mm-long longitudinal cut from the ninth thoracic cord with a micro-surgical blade. After surgeries, the lesions were confirmed to be similar across all experimental groups.
Rats were divided into four experimental groups: SC1-C, 1 mm scaffold only (n=7); SC3-C, 3 mm scaffold only (n=7); SC1, 1 mm scaffold+rBMSC+BDNF (n=8); and SC3, 3 mm scaffold+rBMSC+BDNF (n=8). The scaffold was implanted into the cavity immediately after SCI surgery, after which the musculature was sutured and the skin was closed with wound clips. The animal was then allowed to recover in a clean cage on a heating pad. Antibiotics (gentamicin, 100 μg/kg, i.m.) were given daily for 7 days postinjury, and bladders were evacuated twice daily until reflex bladder function was established.
Behavioral studies
The functional recovery of hind limbs was measured by behavioral tests administered at weekly intervals for 8 weeks after SCI. Open-field locomotion in a rat activity cage (380 [W]×280 [L]×380 [H] mm; Jeungdo B&P) was monitored for 3 min by three observers blinded to the treatments. Each observer scored the rat locomotory behavior using the Basso–Beattie–Bresnahan (BBB) rating scale, which rates contact righting reflex, spinal cord-mediated hindlimb withdrawal to pain, and ability to maintain body position on a flat plane.
Electrophysiology
For electrophysiology experiments, rats were anesthetized using ketamine and rompun (1:1 ratio, 1.5 mL/kg), artificially ventilated with inhaled oxygen, and then transferred to a heating blanket maintained at 37°C. The analog signals of motor-evoked potentials (MEP) were determined using a Medelec Synergy system (Oxford Instruments) attached to an IBM PC running Medelec Synergy system software. For MEP recordings, one myoject needle was positioned in the cerebral cortex and a second myoject needle was set for stimulation at the L1 spinal cord. The exact position of the recording electrode was determined by positioning a recording myoject needle at the left tibial forelimb nerve. A single square pulse (0.1 ms duration) of electrical stimulus at a current of 6 mA was delivered by a stimulus isolator. After analyzing the electrically stimulated forelimb MEPs in terms of latency (∼1 ms) and amplitude (∼10 mV), a recording myoject needle was re-positioned at the left tibial hindlimb nerve. We observed two types of ascending evoked potential waveforms: positive peaks (P1) and negative peaks (N1). The MEP latency reflected the time of the onset in the positive peaks. The MEP amplitude reflected the area of phase for the first positive peak. This latency period was compared with the initial P1 peak of the different groups for each recording time.
Histological analysis
At 4 and 8 weeks postinjury, the animals were anesthetized with ketamine and rompun (1:1 ratio, 1.5 mL/kg) and transcardially perfused with 4% formaldehyde. The spinal cords from T7 to T10 were then retrieved and stored in 4% formaldehyde. The spinal cords were immediately fixed with 10% formalin and embedded in paraffin. The embedded specimens were sectioned (4 μm) along the longitudinal or cross-sectional axis of the implant, and the sections were stained with hematoxylin and eosin, DAPI (6-diamino-2-phenylindoadihydrochloride; Sigma), neuron-specific enolase (NSE; AbD Serotec), Tuj1 (Tubulin beta III Isoform; Chemicon), and neurofilament (NF; Scytek).
The staining procedures for DAPI, NSE, Tuj1, and NF were as follows: The slides were washed with PBS-T (0.05% Tween-20 in PBS), and blocked with 5% (BSA; Roche) and 5% horse serum (Gibco Invitrogen) in PBS for 1 h at 37°C. The sections were incubated overnight at 4°C with primary antibodies (Tuj1, 1:300; NSE, 1:300; NF, 1:20) in PBS containing 1% BSA, washed with PBS-T, and then incubated with the secondary antibody (Alexa Fluor 594–conjugated goat anti-mouse; Invitrogen) for 3 h at room temperature in the dark. The slides were washed again with PBS-T, counterstained with DAPI, and then mounted with fluorescent mounting solution (DAKO). Immunofluorescent images were observed under an Olympus IX81 fluorescence microscope (Olympus) equipped with Meta Image Series software (MetaMorph, Molecular Devices Corporation).
Statistical Analysis
MEP measurements were carried out in independent experiments with SC1 (n=5), SC1-C (n=4), SC3 (n=5), and SC3-C (n=4), and the counted NF and Tuj1 images were carried out in independent experiments with n=3 images for each SCI model, with data given as the mean and standard deviation. The results were analyzed with one-way ANOVA, using the Prism 3.0 software package (GraphPad Software Inc.).
Results
Preparation of scaffolds
In previous studies, we prepared porous three-dimensional PLGA/SIS scaffolds using the ice particle leaching method.15–22 In these studies, in which rBMSCs were cultured in vitro directly on PLGA with different SIS concentrations, we found significant cell attachment and proliferation on PLGA containing 20 wt% SIS. On the basis of these previous results, we chose PLGA/SIS containing 20 wt% SIS as a suitable candidate scaffold for the current experiments. The PLGA/SIS scaffold exhibited an interconnected structure, with porosity >90% and pore diameters of 100–200 μm (data not shown).
Surgery
Surgeries to completely remove a 1- or 3-mm segment of the spinal cord were performed as described in the Experimental section (Fig. 1A–E). To effectively implant scaffolds, we tailored the PLGA/SIS scaffold to a cylindrical form with specified dimensions to fit into the lesion. The cylindrical PLGA/SIS scaffold, with or without seeded rBMSCs, was implanted into the lesion immediately after injury (Fig. 1F). This implantation procedure was well tolerated in all models (SC1 and SC3, with and without rBMSCs).
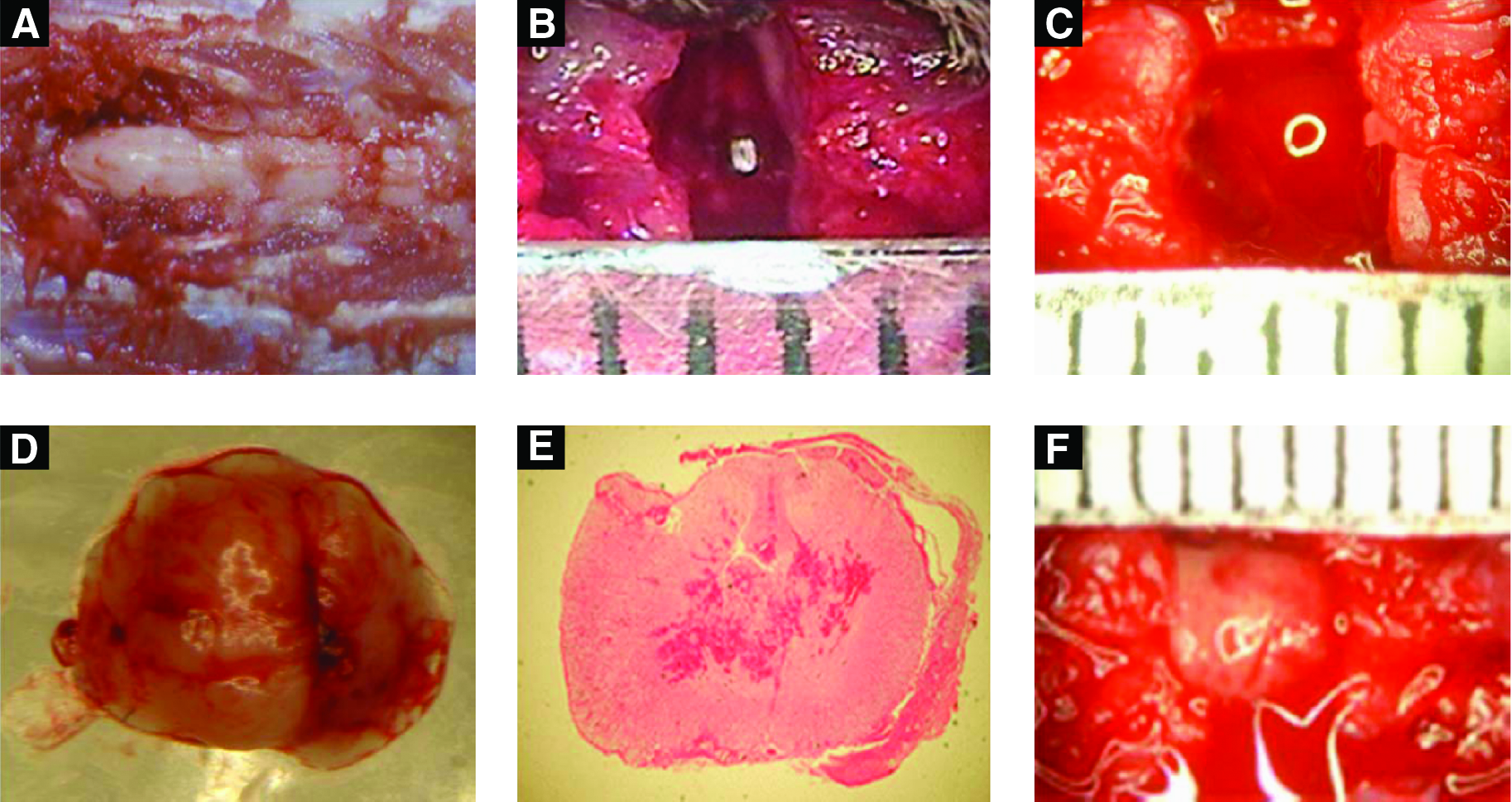
The anatomy image of
Functional recovery: Behavior
After surgery, rats were assigned to their respective groups and treated accordingly. Throughout the duration of the study, the rats remained in relatively good health. Sectioning of the T8–T9 region resulted in bilateral hindlimb paralysis. BBB scores for hindlimb locomotion of all SCI models were determined weekly for 8 weeks after surgery (Fig. 2). All models received BBB scores of 0 immediately after surgery. SC1-C and SC3-C animals seldom moved their hind limbs and received a BBB score of 1 three weeks after injury, and showed gradual improvement, ultimately achieving BBB scores of 3.7±0.6 at 8 weeks after SCI. However, the animals in these groups (SC1-C and SC3-C) never supported their body weight on their hind legs. 19 Animals in rBMSC-implanted SC1 and SC3 groups showed significant functional improvement compared to those in SC1-C and SC3-C groups. Even at the second week after implantation, SC1 and SC3 groups already showed a tendency toward improved BBB scores. The most rapid recovery was observed in the SC1 group, which showed significant improvement after only 4 weeks and reached a final BBB score of 15.8±0.7. Although recovery was slower in the SC3 group, these animals showed significant improvement 8 weeks after SCI and achieved a final BBB score of 8.4±1.5. These results demonstrate the efficacy of rBMSC implantation and highlight the importance of defect length for functional recovery.
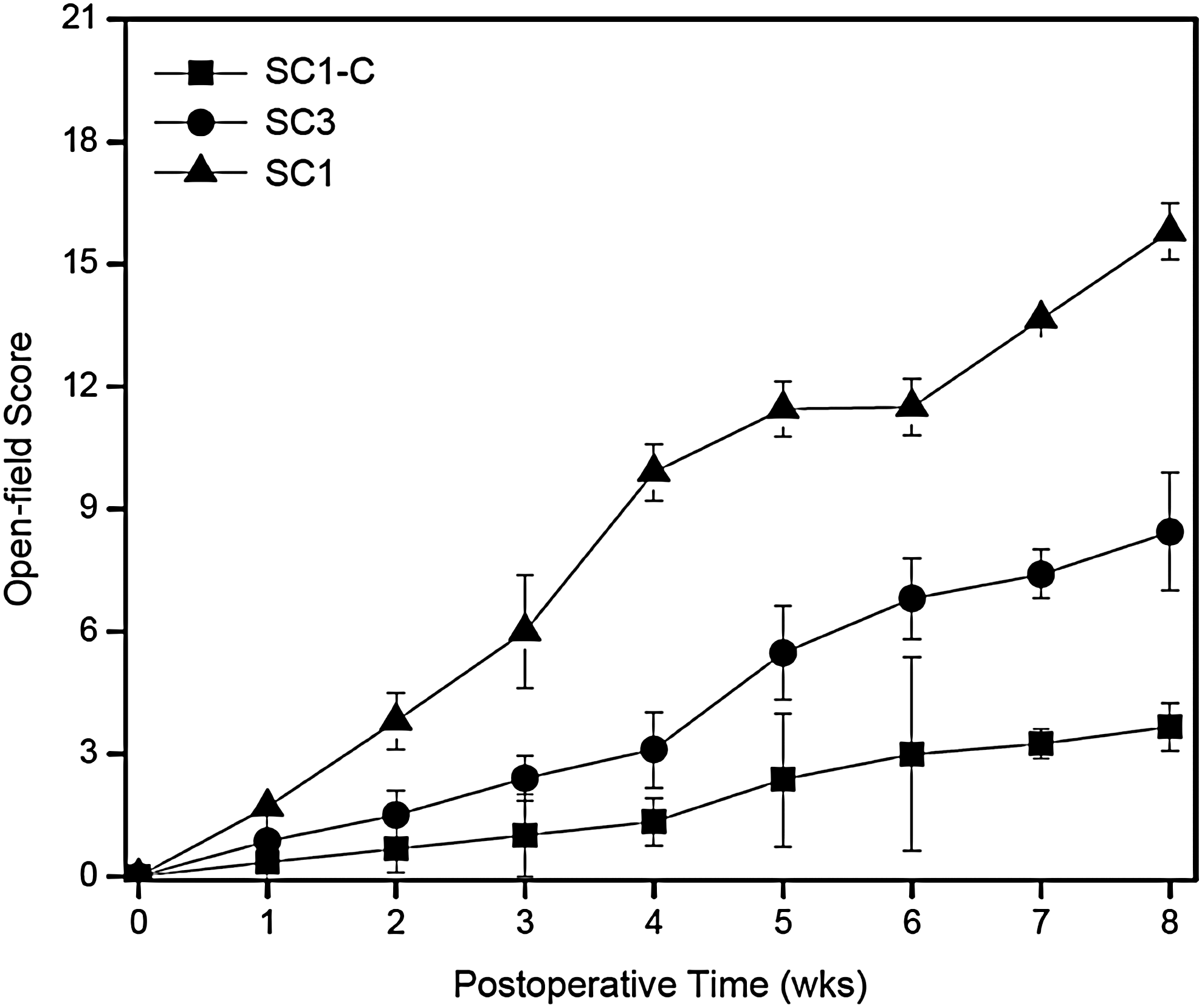
Weekly mean Basso–Beattie–Bresnahan open-field walking scores of SC1 (▴), SC1-C (▪), and SC3 (●) at 4 and 8 weeks after surgery.
Functional recovery: Electrophysiology
The MEP of the injured spinal cord was recorded in the hindlimb area of the sensorimotor cortex using a bipolar disk electrode after stimulation of the LI spinal cord. The waveform of normal animals showed a positive–negative deflection pattern. MEP latency reflected the time to the onset of a positive peak, and amplitude corresponded to the magnitude of the phase of the positive peak. The typical latencies and amplitudes for normal, SC1-C, SC3-C, SC1, and SC3 groups are presented in Figure 3. Normal animals exhibited high-amplitude action potentials, averaging 1780±400 μV at 4 weeks and 2040±450 μV at 8 weeks. A change in the representative waveform became evident for the SCI models. One day after surgery, stimulation elicited no discernable amplitude or latency in any SCI model animals. As shown in Figure 4, at 4 weeks, the peak amplitude of the waveform in SC1-C and SC3-C animals averaged 100±4 μV and 10–50 μV, respectively; after 8 weeks, the amplitude was nearly zero in these control groups. In rBMSC-implanted groups, the amplitude increased from 300±4 μV to 350±4 μV between 4 and 8 weeks for SC1 and from 200±4 μV to 250±4 μV for SC3. The amplitude in SC1 group animals was significantly greater than that in SC3 animals. In both groups, there was a similar linear relationship between MEP amplitude and time, with the amplitudes of SC1 and SC3 increasing with increasing time after surgery. Differences in amplitude between SCI models with and without rBMSCs were significant. The latency was also shorter at 8 weeks than at 4 weeks, indicating faster conduction velocities. However, the change in the position of latency was not significantly different from that of normal animals throughout the experiment (p<0.05 or p<0.001).
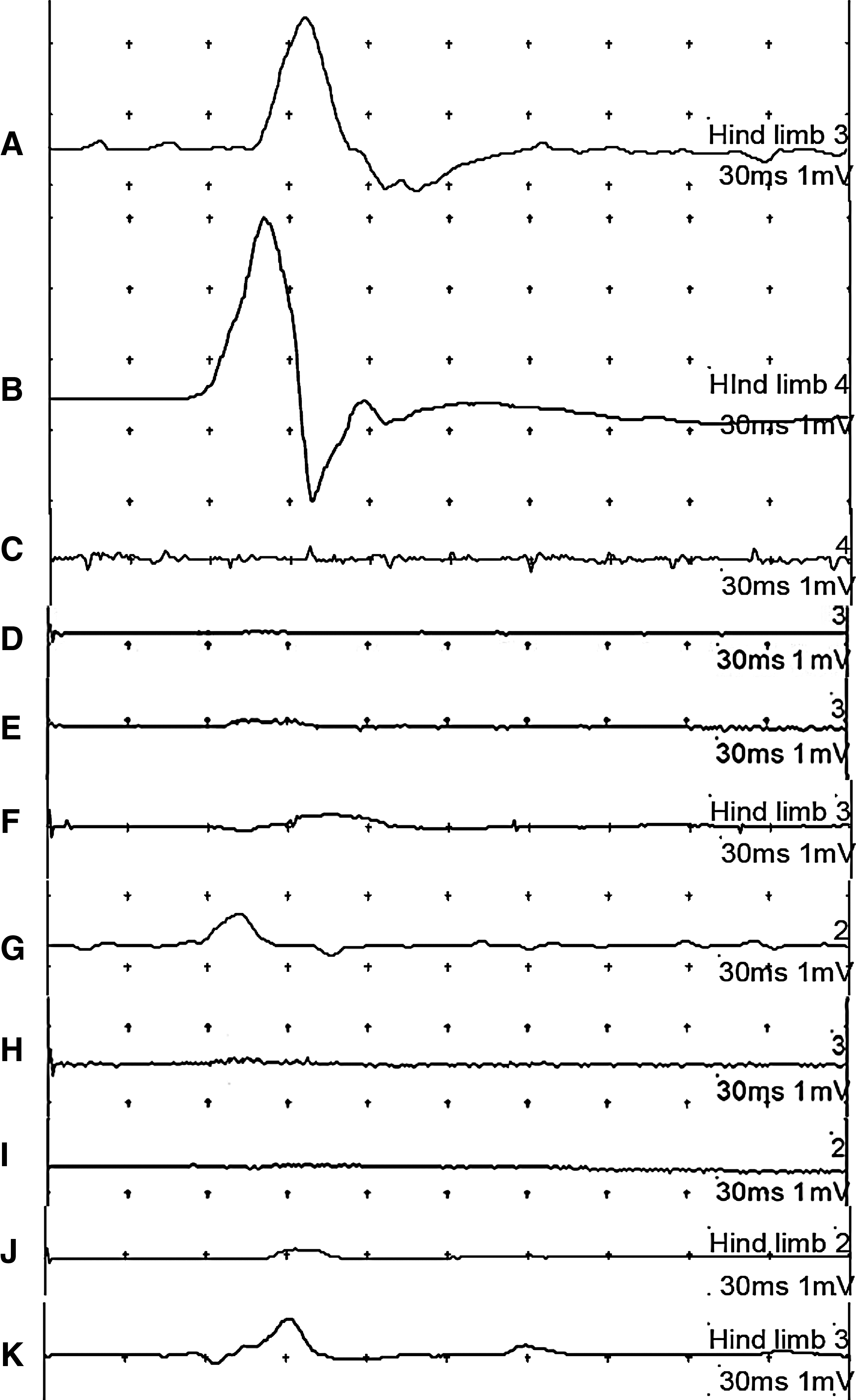
Motor-evoked potentials graphs of normal at
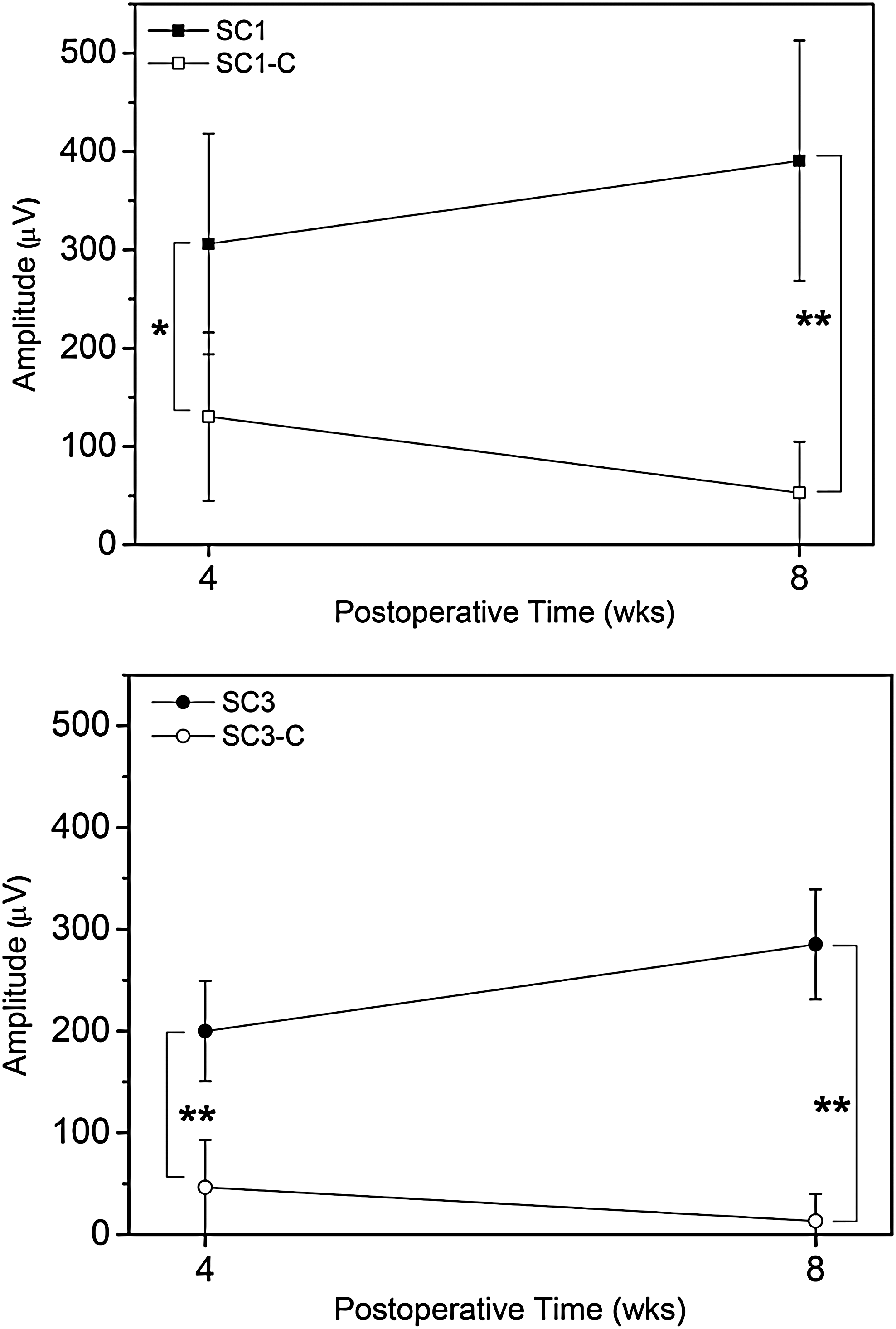
The aveavgae amount of amplititude after motor-evoked potentials measurement of SC1 (n=5), SC1-C (n=4), SC3 (n=5), and SC3-C (n=4) at 4 and 8 weeks after surgery. Statistical analysis was performed using one-way ANOVA with Bonferroni's multiple comparison (*p<0.05, **p<0.001).
Histology
The injured spinal cords were removed 4 and 8 weeks after surgery and grossly imaged to examine changes in shape and histology that occurred in vivo (Fig. 5). In SC1-C and SC3-C animals, there were disorganized arrays at the gaps between the injured site and the original spinal cord. In contrast, rBMSC-implanted SCI models exhibited opaque tissues between the injury site and the original spinal cord, providing probable evidence of an organized array of new tissue.
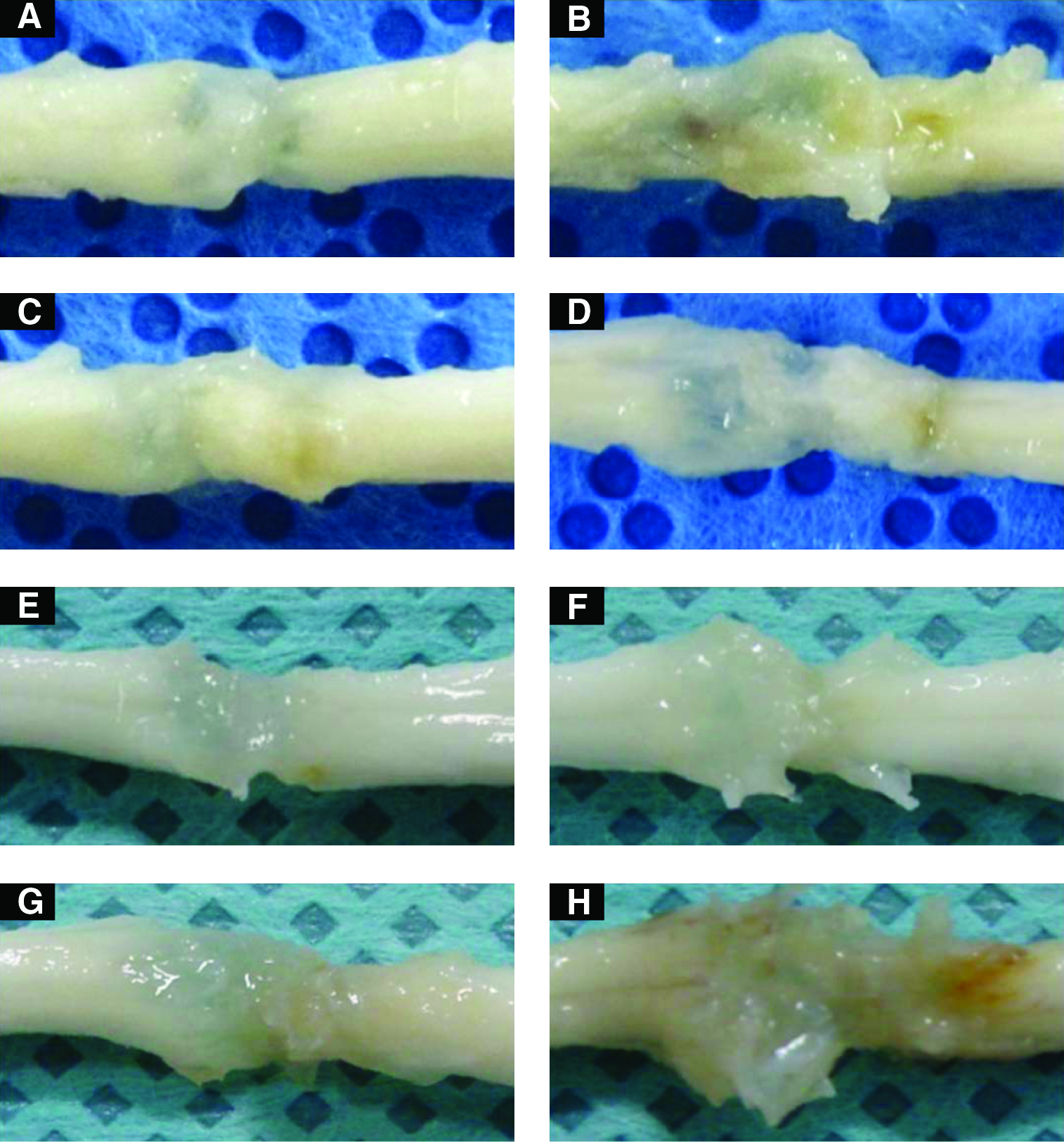
The images of spinal cord
An examination of immunohistochemically stained sections (cross and longitudinal) from the injury sites of SC1-C, SC3-C, SC1, and SC3 animals showed almost no NF or NSE staining in SC1-C or SC3-C animals (Fig. 6). NF staining of longitudinal sections through the injury site showed clear evidence that axonal regeneration had crossed the lesion only in the rBMSC-implanted SC1 and SC3 groups (Fig. 7). NF staining of cross sections also provided an indication of the extent of axonal regeneration. BrdU-labeled rBMSCs used to monitor implanted rBMSCs were visible for only 4 weeks after SCI (data not shown), suggesting that BrdU may not be adequate for longer observation (e.g., 8 weeks). To enable more reliable tracking of cells in vivo, we used PKH-labeling to monitor rBMSCs. The green fluorescence emitted by PKH-labeled rBMSCs was detected at both 4 and 8 weeks after transplantation, indicating cell survival in the implantation site. NF- and Tuj1-positive cells were counted and normalized to the total stained tissue area in order to determine the extent of de novo nerve formation (Fig. 8). Significantly more NF- and Tuj1-positive cells were observed in the SC1 group than in the SC3 group (p<0.05 or p<0.001).
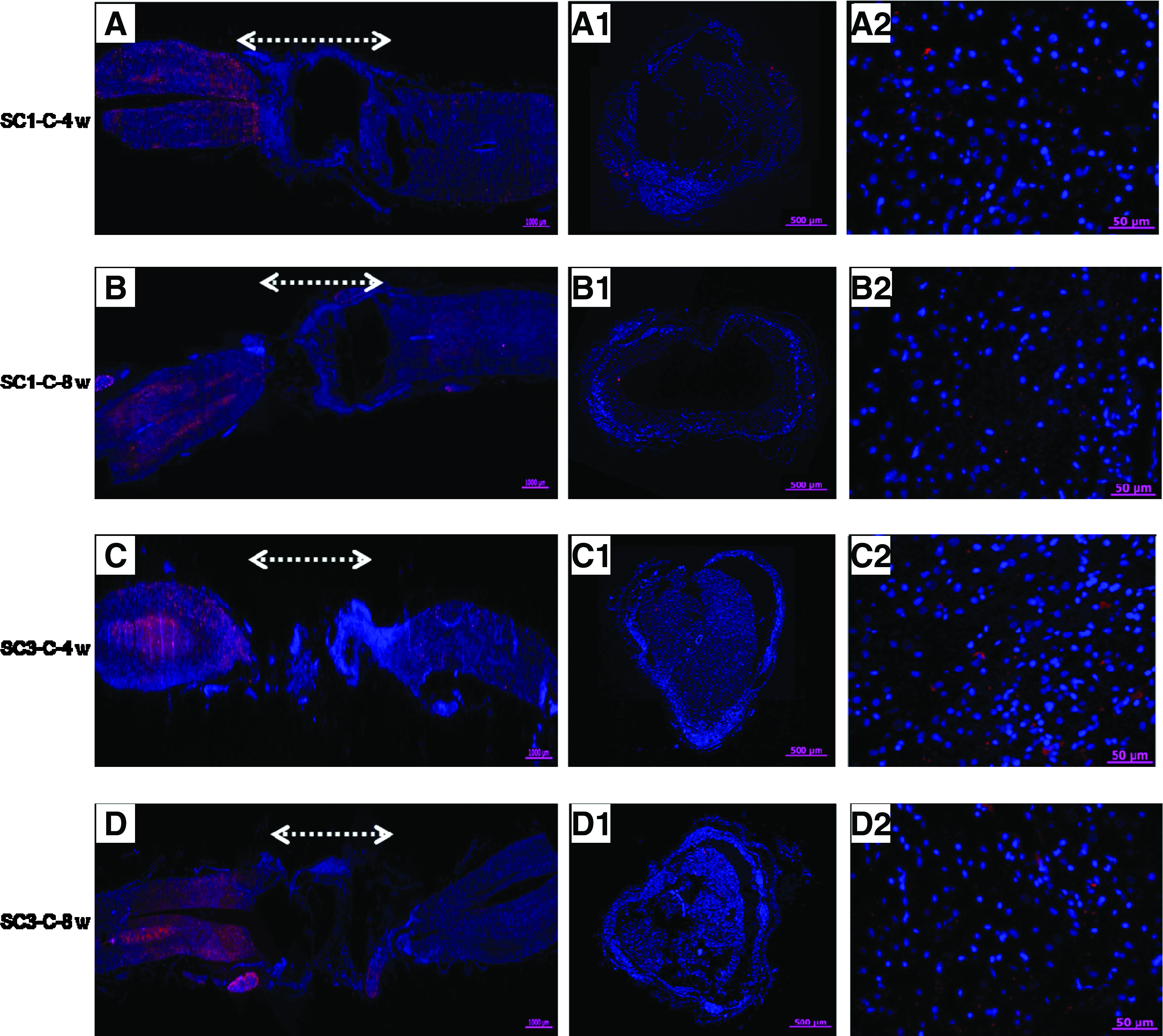
NF (red) and DAPI (blue) merged immunofluorescent staining
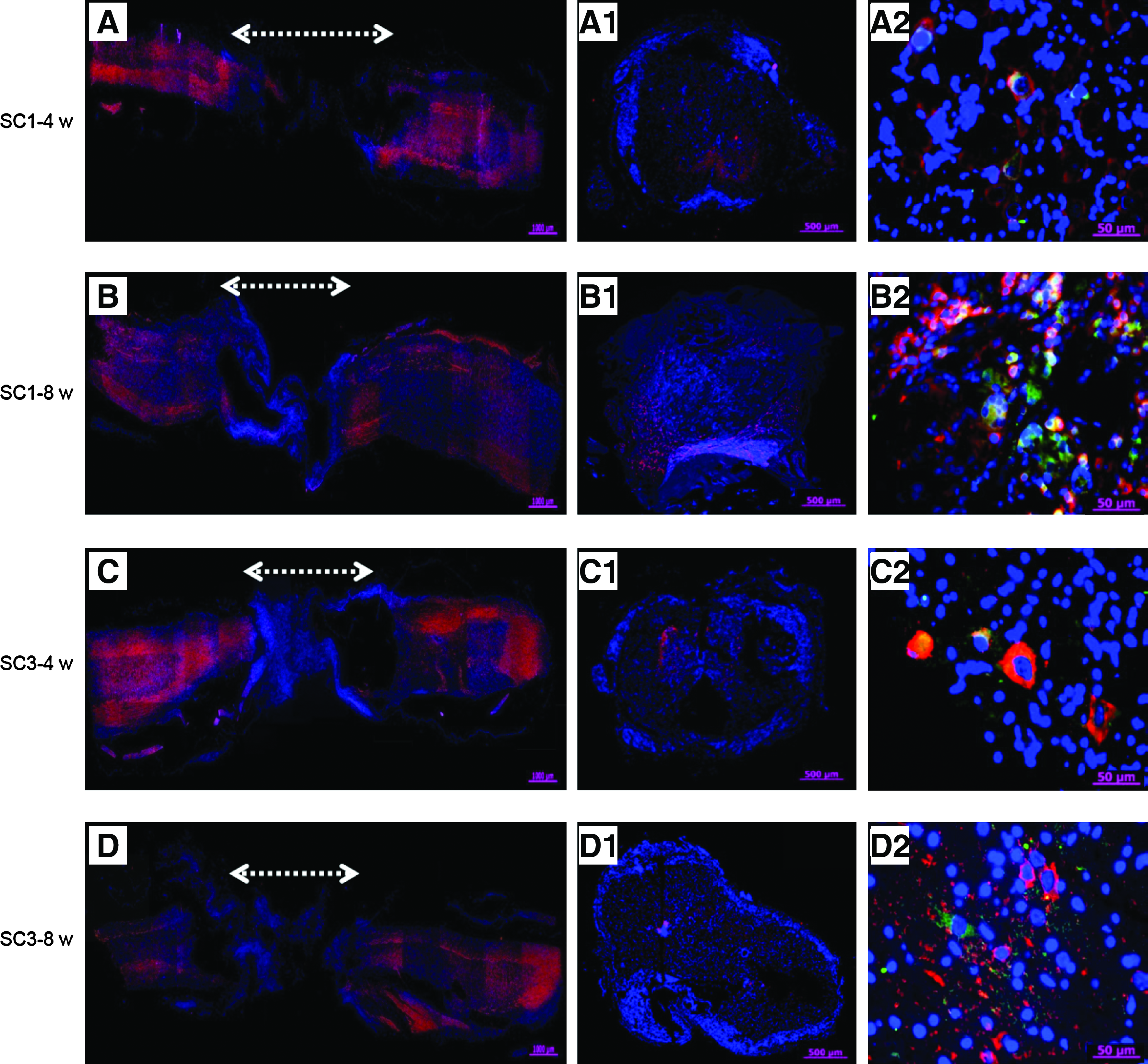
NF (red) and DAPI (blue) merged immunofluorescent staining
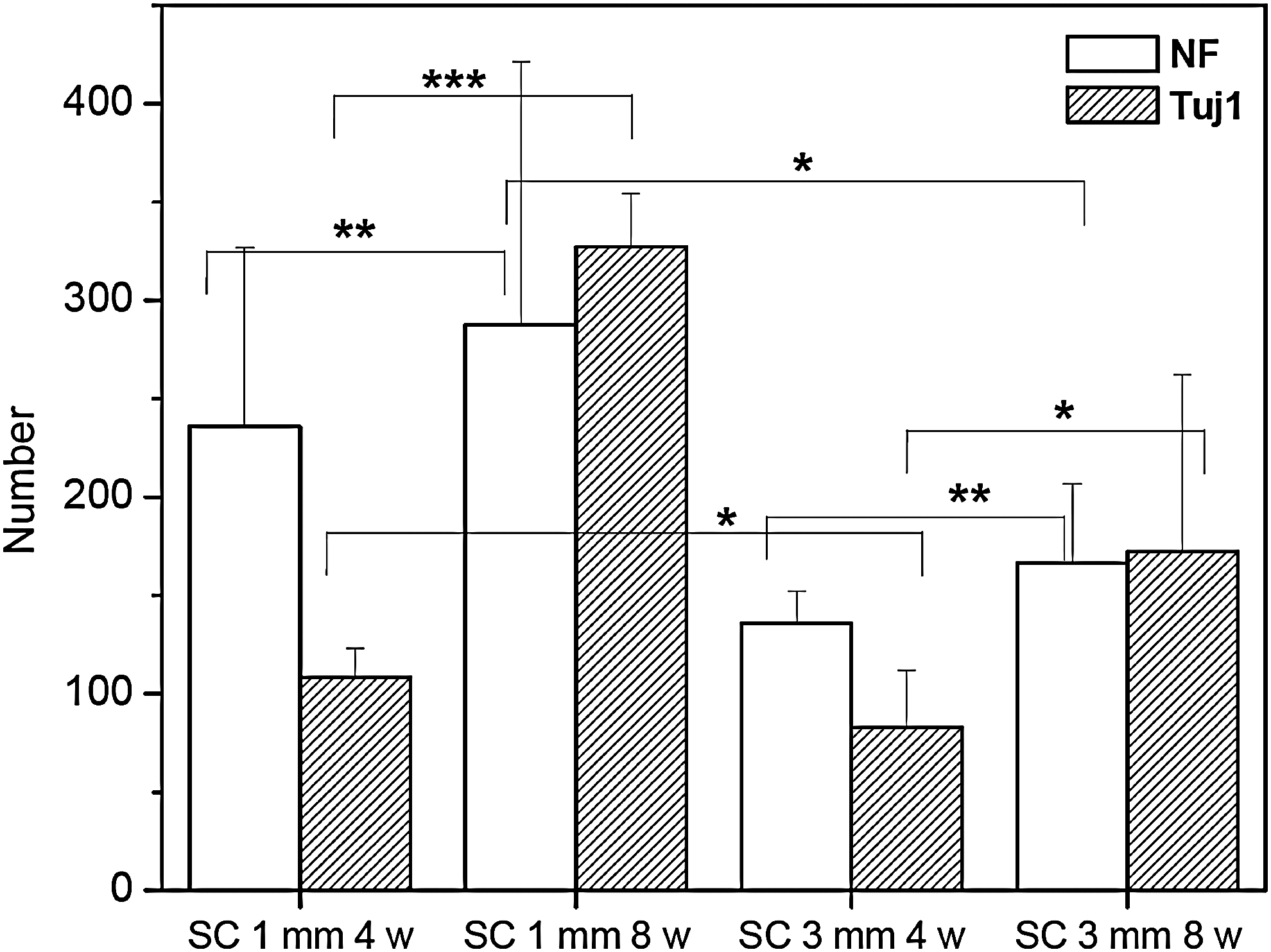
The counted NF and Tuj1 number in immunofluorescent staining of cross-sectioned image of SC1 and SC3 at 4 and 8 weeks. Statistical analysis was performed using one-way ANOVA with Bonferroni's multiple comparison (*p<0.05, **p>0.05, ***p<0.001).
Discussion
There have been many reports on the use of stem cells for cell therapies in various animal SCI models.5,6,24 BMSCs can be readily obtained from bone marrow and expanded in culture, and notably have the capacity to differentiate into cells with neuronal phenotypes. 25 In addition, BMSC can secrete a variety of growth factors and cytokines that could contribute to the repair of SCI26,27 and can trigger the endogenous survival signaling pathway Erk1/2 and Akt phosphorylation in neurons. 28 Thus, BMSCs may be a very useful source of cells for functional reconstitution and repair of tissues in the lesion site of SCI models. However, previous studies have shown that BMSC numbers are high only in the initial period after transplantation in SCI models, and these cells disappear with increasing time after transplantation.29–31
The utility of polymer scaffolds may lie in their ability to extend the survival period of cells. The present work tested the feasibility of therapeutic cell delivery using a scaffold in an animal SCI model. The porous PLGA/SIS scaffold supported seeded rBMSCs and permitted in-growth of implanted cells in vivo; it also facilitates nutrient transport to extend the survival period of rBMSCs in the PLGA/SIS scaffold. This study does not provide exact data for survival ratio of the seeded rBMSCs; however, PKH-labeled rBMSCs seeded within the PLGA/SIS scaffold in this experiment were found to survive for at least 8 weeks after surgery.
Several studies have shown that partial or contusion SCI models at the T8–T9 section causes an initial paralysis of the hind limbs, eventually followed by considerable self-recovery of motor function under normal circumstances.32,33 Thus, it is difficult to determine whether regeneration in spinal cord models is directly correlated with other parameters, like self-recovery. Recent studies have employed the more challenging strategy of using the complete spinal cord transection model rather than partial or contusion SCI models. The completely transected SCI model applied in this study can be used to exclude self-recovery after spinal injury. Even though complete transection injuries do not recapitulate typical human SCIs, the complete spinal cord transection model can allow precise evaluation of spinal cord regeneration by rBMSC-seeded PLGA/SIS scaffold. In this work, we observed that implantation of rBMSC-seeded PLGA/SIS scaffolds into the complete spinal cord transection model promoted significant functional improvement, as measured by behavioral and electrophysiological tests.
Differences in the length of spinal cord that is completely transected may present a critical issue in designing therapeutic strategies to mitigate SCI. Despite this, there are no reports relating the dependence of functional recovery on injury length. Here, we found that completely transected SCI model animals transplanted with rBMSC-seeded PLGA/SIS scaffolds exhibited significantly improved BBB outcomes that varied according to the length of the injury, showing that behavioral performance was greater in animals with small-length defects than in those with larger defects.
MEP measurements are also important because this parameter can assess the electrical function of motor neurons in the spinal cord. 34 For the SCI model, MEPs generally record evoked potentials for hind limbs, and as such provide an assessment of the functional integrity of the spinal cord. This approach may be used to examine the therapeutic efficacy of our SCI model. Complete injuries of the spinal cord destroy the entire neural network at and around the spinal cord. In this work, all SCI animals exhibited prolonged latency immediately after the injury. Importantly, we found that SC1 model animals transplanted with rBMSC-seeded PLGA/SIS scaffolds showed significantly reduced MEP latency at 8 weeks. Our observation of reduced latency of MEP responses in recovering animals indicates that transplanted rBMSC-seeded PLGA/SIS scaffolds may be effective even for completely severed SCIs.
At 8 weeks postsurgery, we found that the tissue defect was bridged by the implanted PLGA/SIS scaffolds, which seemed to be well apposed to the completely transected spinal cord. To evaluate the degree of a nerve formation between transverse sections and further address the feasibility of rBMSC-seeded PLGA/SIS scaffolds, we reconstructed histological images of removed spinal cord tissue. Positive immunofluorescent staining for tracked rBMSCs clearly showed that implanted rBMSCs survived in rBMSC-seeded PLGA/SIS scaffold and induced de novo nerve formation. The merged images were immunoreactive for NF, implying that rBMSCs seeded within the PLGA/SIS scaffolds differentiated into nerve cells. Collectively, these results suggest that new nerves could be conducted across the injured spinal cord on the PLGA/SIS scaffold, connecting proximal and distal spinal cord segments.
Conclusion
We found that implantation of rBMSC-seeded PLGA/SIS scaffolds into a complete spinal cord transection model promotes significant improvement in functional outcome as measured by behavioral and electrophysiological tests. Moreover, axonal regeneration was activated to a greater degree with rBMSC-seeded PLGA/SIS scaffolds than with control PLGA/SIS scaffolds. The present study thus shows that the complete transection SCI model employing different defect lengths is a valuable tool for testing therapeutic strategies for nerve regeneration. As such, it should have a significant impact on evaluating the feasibility of therapeutic cell delivery using three-dimensional scaffolds, especially in the context of complete spinal cord transection. Moreover, the present study establishes the basis for subsequent trials of the complete transection SCI model in larger experimental animals.
Footnotes
Acknowledgments
The initial study of this work started at KRICT, the former affiliation of corresponding author, and then finished at Ajou University. Thus, the corresponding author thanks KRICT for providing equipment and materials to conduct initial experimental of this work. We would like to thank the KMOHW (Grant no. A050082) and MKE (Grant no. 10038665) and Priority Research Centers Program through NRF funded by the Ministry of Education, Science and Technology (2010-0028294) for providing financial support to this study.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.