
Abstract
Engineering cartilaginous tissue of a scale necessary to treat defects observed clinically is a well-documented challenge in the field of cartilage tissue engineering. The objective of this study was to determine how the composition and mechanical properties of cartilaginous tissues that are engineered by using bone marrow-derived mesenchymal stem cells (MSCs) depend on the scale of the construct. Porcine bone marrow-derived MSCs were encapsulated in agarose hydrogels, and constructs of different cylindrical geometries (Ø4×1.5 mm; Ø5×3 mm; Ø6×4.5 mm; Ø8×4.5 mm) were fabricated and maintained in a chemically defined serum-free medium supplemented with transforming growth factor-β3 for 42 days. Total sulfated glycosaminoglycan (sGAG) accumulation by day 42 increased from 0.14% w/w to 0.88% w/w as the construct geometry increased from Ø4×1.5 to Ø8×4.5 mm, with collagen accumulation increasing from 0.31% w/w to 1.62% w/w. This led to an increase in the dynamic modulus from 90.81 to 327.51 kPa as the engineered tissue increased in scale from Ø4×1.5 to Ø8×4.5 mm. By decreasing the external oxygen tension from 20% to 5%, it was possible to achieve these higher levels of mechanical functionality in the smaller engineered tissues. Constructs were then sectioned into smaller subregions to quantify the spatial accumulation of extracellular matrix components, and a model of oxygen diffusion and consumption was used to predict spatial gradients in oxygen concentration throughout the construct. sGAG accumulation was always highest in regions where oxygen concentration was predicted to be lowest. In addition, as the size of the engineered construct increased, different regions of the construct preferentially supported either sGAG or collagen accumulation, thus suggesting that gradients in regulatory factors other than oxygen were playing a role in determining levels of collagen synthesis. The identification of such factors and the means to control their spatial concentration within developing tissues represents a central challenge in engineering large cartilaginous grafts.
Introduction
There has been increased interest in using mesenchymal stem cells (MSCs) for functional cartilage tissue engineering. 9 The potential advantages of using MSCs as an alternative to chondrocytes for cartilage repair strategies are well documented. 10 Isolating chondrocytes requires retrieving a biopsy of articular cartilage that can produce additional damage to the joint surface. 11 In addition, chondrocytes have demonstrated a decreased capacity for chondrogenesis with extended in vitro passaging and with age.12–14 For these reasons, numerous studies report using progenitor cells isolated from tissues such as bone marrow,15–21 synovium,22–25 infrapatellar fat pad,26–30 and adipose tissue,31–33 among others, for cartilage tissue engineering. Such progenitor cells have been combined with a variety of scaffolds, from relatively inert marine-derived biomaterials such as agarose and alginates,34–39 to silk scaffolds40–42 and electrospun meshes, 43 to more bioactive scaffolds derived from native articular cartilage extracellular matrix.44,45 Whether such approaches can be used to engineer cartilaginous grafts of sufficient scale to treat large defects has yet to be addressed.
A feature of MSCs in regenerative events such as endochondral ossification during fracture healing is their ability to synthesize large volumes of cartilaginous tissue in avascular regions of the regenerating tissue. In a tissue engineering context, a similar low oxygen environment may be created within cell-seeded constructs due to cellular utilization of oxygen in the peripheral regions of the developing tissue. 46 Low oxygen tension has been shown in numerous studies to enhance chondrogenesis of MSCs.21,26,47–50 The objective of this study is to determine how the properties of cartilaginous tissues engineered using bone marrow-derived MSCs will change both spatially and temporally as the scale of the construct increases. It is hypothesized that the functional properties of such tissues will improve as the geometrical volume of the engineered construct is increased due to the development of a lower oxygen environment within larger volume constructs. Computational modeling will be used to predict the oxygen tension within such cell-seeded constructs. It is further hypothesized that the more chondrogenic microenvironment generated in such large MSC-seeded constructs can be recapitulated in smaller construct geometries by lowering the surrounding oxygen tension, thereby enhancing the functional properties of the resulting tissue.
Materials and Methods
Cell isolation and construct assembly
Bone marrow was aseptically harvested from the femoral diaphysis of two 4-month-old pigs (∼50 kg). Marrow was repeatedly aspirated by using a 16-gauge needle to break up large aggregates before centrifugation at 650 g for 5 min. The separated floating adipose layer was discarded, and the cell pellet was resuspended in expansion media consisting of high-glucose Dulbecco's modified Eagle's medium (hgDMEM) GlutaMAX supplemented with 10% fetal bovine serum and 100 U/mL penicillin/streptomycin (all Gibco–Biosciences). Cells were triturated through a 20-gauge needle to create a single-cell suspension and filtered through a 40 μm nylon cell strainer (B.D. Falcon–Unitech). An aliquot of this suspension was treated with 4% acetic acid to lyse the red blood cells, and viable mononuclear cells were counted by using a hemocytometer in the presence of 0.4% trypan blue. Mononuclear cells were initially seeded at a density of 50,000 cells/cm2 in T-75 cm2 flasks. Cells were subcultured at a density of 5000 cells/cm2 after colony formation and expanded to passage two. We have previously demonstrated that porcine bone marrow-derived MSCs expanded in this manner can differentiate down an osteogenic, adipogenic, or chondrogenic pathway. 27
Cells were mixed with 4% agarose (Type VII; Sigma-Aldrich) at a ratio of 1:1 at ∼40°C to yield a final gel concentration of 2% at a density of 15 million cells/mL. The agarose-cell suspension was cast in stainless steel moulds and allowed to set for 20 min before coring with sterile biopsy punches to produce cylindrical constructs of the following dimensions (diameter×height): Ø4×1.5 mm; Ø5×3 mm; Ø6×4.5 mm; and Ø8×4.5 mm. During the fabrication stage, a hatched pattern was molded onto the top surface of constructs to permit tracking of construct orientation for histology purposes and also to ensure that constructs were not turned over during the culture period. Constructs did not float during the culture period, and rested on the bottom surface of wells of each culture plate. Details of media volumes and well plates are provided in Figure 1A.

Constructs were maintained in a chemically defined chondrogenic medium consisting of hgDMEM supplemented with 100 U/mL penicillin/streptomycin (both from Gibco), 100 μg/mL sodium pyruvate, 40 μg/mL L-proline, 50 μg/mL L-ascorbic acid-2-phosphate, 1.5 mg/mL bovine serum albumin, 1×insulin–transferrin–selenium, 100 nM dexamethasone (all Sigma-Aldrich), and 10 ng/mL of transforming growth factor-β3 (ProSpec-Tany TechnoGene, Ltd.). Medium was changed twice per week for 6 weeks. In parallel, constructs of dimensions Ø5×3 mm and Ø6×4.5 mm were cultured at either 20% or 5% oxygen tension at 37°C and 5% CO2. For culturing at 5% oxygen tension, a hypoxia incubator was used (Brunswick Galaxy 48R incubator with 1%–19% O2 control), which is maintained with nitrogen gas to create this low oxygen environment. All samples were assessed at day 42.
Histology and immunohistochemistry
At each time point, two additional samples per group were fixed in 4% paraformaldehyde overnight, dehydrated, and embedded in paraffin. The constructs were sectioned (6 μm thickness) perpendicular to the disk face. Sections were stained with 1% alcian blue 8GX (Sigma-Aldrich) in 0.1 M HCl for sulfated glycosaminoglycans (sGAGs), and picro-sirius red for collagen. The deposition of collagen types I and II were identified through immunohistochemistry. Briefly, sections were rinsed with phosphate-buffered saline (PBS), quenched of peroxidase activity for 20 min, and treated with chondroitinase ABC (Sigma; 0.25 U/mL) in a humidified environment at 37°C for 1 h to enhance permeability of the extracellular matrix by removal of chondroitin sulfate. Slides were again rinsed with PBS and blocked with 10% goat serum for 30 min. Sections were incubated with mouse monoclonal collagen type I diluted 1:400 (Abcam) (concentration 5.4 mg/mL) or mouse monoclonal anti-collagen type II diluted 1:100 (Abcam) (concentration 1 mg/mL) for 1 h at room temperature. After washing in PBS, the secondary antibody for type I and type II collagen (Anti-Mouse IgG Biotin antibody produced in goat) (concentration 1 g/L) binding was applied for 1 h. Color was developed by using the Vectastain ABC reagent (Vectastain ABC kit; Vector Laboratories) for 45 and 5 min exposure to peroxidase DAB substrate kit (Vector Laboratories). Slides were dehydrated through ethanol and xylene and mounted with Vectamount medium (Vector Laboratories). Positive and negative controls (porcine cartilage and ligament) were included in the immunohistochemistry staining protocol for each batch.
Quantitative biochemical analysis
To quantify the spatial accumulation of biochemical constituents, all but the Ø4×1.5 mm constructs were sectioned into specific subregions and analyzed separately (Fig. 1A). Samples were digested in screw cap microtubes with 500 μL of 125 μg/mL papain (Papain from papaya latex, P3125, Batch No. 010M7013) in 0.1 M sodium acetate, 5 mM L-cysteine-HCl, 0.05 M EDTA, and pH 6.0 (all from Sigma-Aldrich) at 60°C under constant rotation (Stuart® Rotator SB3; Lennox Laboratory Supplies) for 18 h. DNA content was quantified by using the Hoechst Bisbenzimide 33258 dye assay as previously described 51 with a calf-thymus DNA standard. sGAG content was quantified by using the dimethylmethylene blue dye-binding assay (Blyscan, Biocolor, Ltd.), with a chondroitin sulfate standard. Total collagen content was determined by measuring the hydroxyproline content. Samples were hydrolyzed at 110°C for 18 h in 38% HCl and assayed by using a chloramine-T assay 52 with a hydroxyproline:collagen ratio of 1:7.69. 53 Each biochemical constituent was normalized to the tissue wet weight and DNA content.
Mechanical analysis
Constructs from each group were mechanically tested in unconfined compression between impermeable platens by using a standard material testing machine with a 5 N load cell (Zwick Z005; Roell) as previously described. 54 Stress relaxation tests were performed, consisting of a ramp and hold cycle with a ramp displacement of 1 μm/s, until 10% strain was obtained and maintained until equilibrium was reached (30 min). Dynamic tests were performed immediately after the stress relaxation cycle. A cyclic strain amplitude of 1% superimposed on the 10% strain was applied for 10 cycles at 1 Hz. The compressive equilibrium modulus and dynamic modulus were determined from these tests.
Statistical analysis
Statistical analysis was performed by using MINITAB 15.1 software package (Minitab Ltd.) with three to four samples analyzed for each group at each time point. Groups were analyzed for significant differences by using a general linear model for analysis of variance with factors, culture time, construct size, construct region, culturing regime (normal or low oxygen), and interactions between these factors examined. Tukey's test for multiple comparisons was used to compare conditions. Significance was accepted at a level of p<0.05. Numerical and graphical results are displayed as mean±standard deviation. The entire experiment was replicated with cells from the second donor, which confirmed the findings presented in this article.
Finite element modeling of oxygen concentration profiles
A numerical method was developed to predict the oxygen gradients within cylindrical constructs by using a commercial finite element package (COMSOL Multiphysics™ v.3.2) in a similar fashion as previously described.55–57
An axisymmetric model was employed with symmetry boundary conditions imposed at the construct centerline. The concentrations at the edge and top surfaces of the construct in contact with the culture medium were assumed to be equivalent. The oxygen concentration Cox as a function of time t within a cylindrical construct can be described as follows:
where oxygen is assumed to diffuse with a constant diffusion coefficient, Dconstruct (m2s−1). The oxygen concentration is denoted by Cox (mol m3). The depth into the construct sample is z (m). The reaction rate, R (mol·m3s−1), is the rate at which oxygen is consumed by the cells within the hydrogel system. The reaction rate R (mol·m3s−1) is dependent on the local number of cells (ncell) and on the maximal rate of consumption of oxygen of the cells Vmax (mol·cell−1·s−1). As oxygen concentration, Cox, becomes depleted, the oxygen consumption rate of the cells alters according to Michaelis–Menten kinetics, with Km (mol·m3) being the oxygen concentration at half maximal oxygen consumption, yielding an equation for R:
The diffusion coefficient Dconstruct was estimated by using the Mackie and Meares relation,58,59 defined as the ratio of diffusivity in the construct (Dconstruct) to diffusivity in free solution (D0), which is a function of the volume fraction of water in the construct:
where D0 is the diffusion coefficient of oxygen in medium (3.0×10−9 m2 s−1) at 37°C, 60 and ϕ w is=0.98 for 2% agarose hydrogel yielding a value for Dconstruct=2.77×10−9 m2 s−1. Based on the average reported oxygen consumption rate for bone marrow MSCs undergoing chondrogenesis in a pellet culture system over 21 days, 61 Vmax was estimated to be 18.7 fmol h−1 cell−1. Since this parameter is known to vary during chondrogenesis, 61 in a second simulation, we predicted how the spatial concentration of oxygen would change after doubling this rate (Vmax=37.4 fmol h−1 cell−1). The Michaelis–Menten constant, Km, was set to 5×10−3 mol m3. 60 The cell density, ncell, was set to 15×106 cells/mL. Results for the oxygen concentrations as a function of depth (centerline) and along the central radial direction were predicted as depicted in Figure 1B.
Results
Model predictions of oxygen concentration within MSC-seeded constructs
Oxygen concentration profiles within the MSC-seeded hydrogels were predicted to depend on construct size (Fig. 2A). The oxygen concentration was predicted to decrease with increasing distance from both the top (Fig. 2C) and side (Fig. 2D) surfaces of the construct. The predicted oxygen concentration within the bottom core of the construct decreased as the size of the construct increased from Ø4×1.5 to Ø8×4.5 mm. Assuming a peripheral oxygen tension of 20%, an oxygen concentration of approximately 10.8% was predicted within the core of the largest Ø8×4.5 mm construct, compared with a predicted concentration of approximately 18.2% in the core of the smallest Ø4×1.5 mm construct. As expected, doubling the oxygen consumption rate decreased the predicted oxygen concentration within constructs (Fig 2B, E, F). For example, the minimum oxygen tension in the core of the largest Ø8×4.5 mm construct was predicted to reduce from 10.8% to 2.6% when Vmax was increased by a factor of two. Reducing the oxygen concentration at the edge and top surfaces from 20% to 5% (Fig. 3) was also predicted to reduce core oxygen concentration values from approximately 16.3% to 1.9% in the Ø5×3 mm constructs, and from approximately 14.3% to 0.7% in the Ø6×4.5 mm constructs (Vmax=18.7 fmol h−1 cell−1).

Predicted oxygen concentration profiles for hydrogel constructs (Ø4×1.5 mm; Ø5×3 mm; Ø6×3 mm; Ø8×4.5 mm) cultured at 20% oxygen tension as illustrated in Figure 1.

Predicted oxygen concentration profiles for hydrogel constructs (Ø5×3 mm; Ø6×4.5 mm) cultured at 5% oxygen tension.
Composition and distribution of extracellular matrix within constructs of different scales
Less intense staining for sGAGs and collagen was observed for smaller-sized constructs (Fig. 4). The bottom core of constructs cultured in a normoxic environment (20% O2) generally stained intensely for type II collagen, with cells appearing to be slightly larger and/or more clustered in this region. This spatial difference in cell shape was particularly evident in intermediate-sized constructs, where cells appeared larger in the core of the construct compared with the periphery (see Fig. 4). In a normoxic environment, a peripheral region staining weakly for sGAGs and type II collagen was also observed, particularly in the smaller and intermediate-sized constructs. A more homogenous staining for sGAGs in the Ø5×3 mm and Ø6×4.5 mm constructs was observed when maintained in a low oxygen environment (5% O2). For larger constructs (Ø8×4.5 mm), it was observed that the very core region stained less intensely for cartilaginous matrix by day 42. For the constructs cultured in a low oxygen environment, the central core regions of the constructs also stained slightly less intensely for collagen compared with the surrounding tissue.
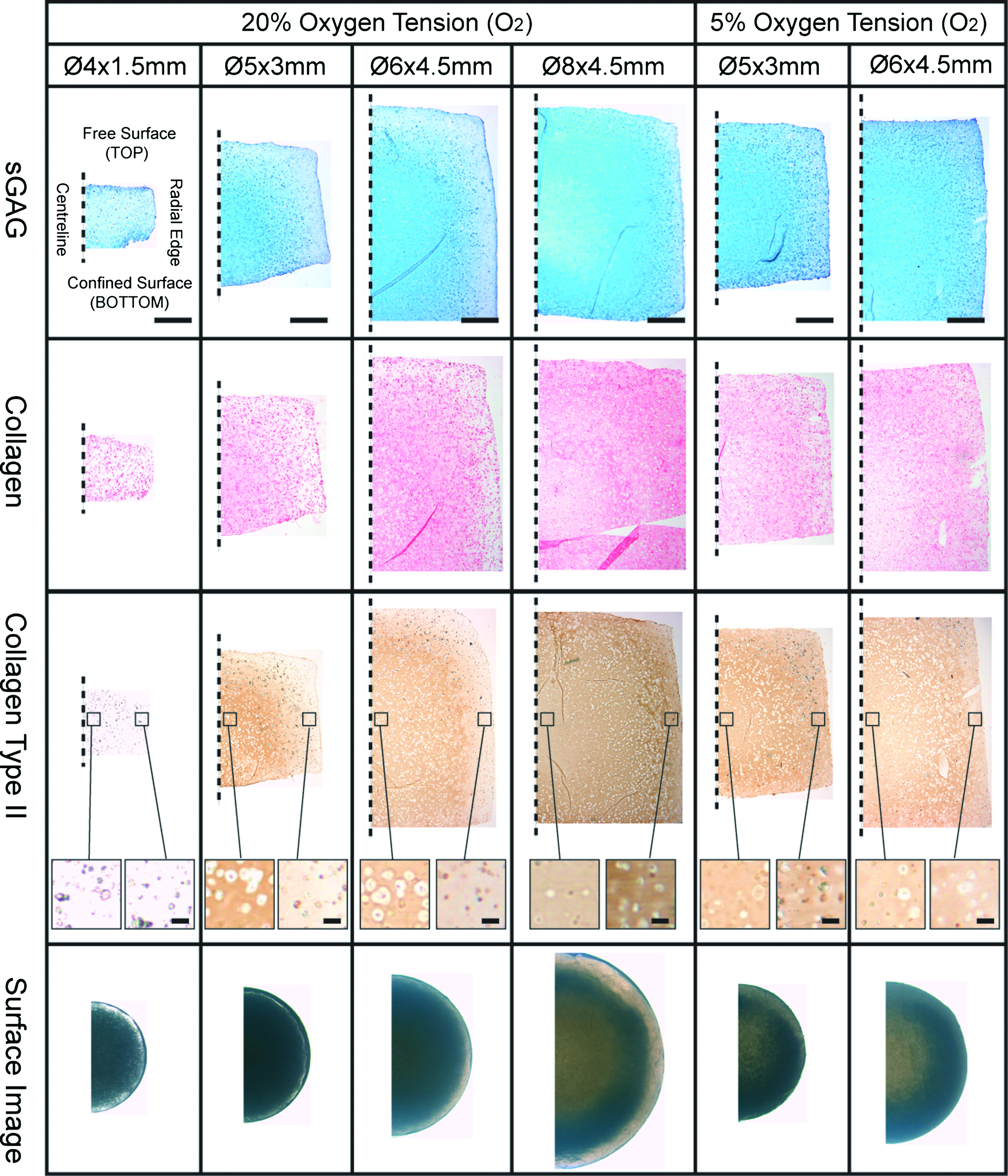
Histological evaluation (sGAG, collagen, collagen type II, and light microscopic surface image) of mesenchymal stem cell-seeded constructs of differing geometries at day 42. Constructs of dimension Ø5×3 mm and Ø6×4.5 mm were also cultured at a lower oxygen tension of 5%. Dashed line indicates the centerline of the engineered tissue. Scale bar for low magnification=1 mm. Higher magnification images in collagen type II stained sections are from core and annulus regions showing the difference in cell shape, scale bar=50 μm. sGAG, sulfated glycosaminoglycan. Color images available online at www.liebertonline.com/tea
A slight decrease in DNA content was observed in the largest constructs (Ø8×4.5 mm) compared with all other construct sizes (Fig. 5A). Total sGAG and collagen accumulation, measured as a percentage of wet weight or normalized to DNA, increased as the construct geometry increased from Ø4×1.5 to Ø8×4.5 mm (Fig. 5B, C). Total sGAG accumulation increased from a value of 0.14%±0.0% w/w for the Ø4×1.5 mm construct to 0.88%±0.04% w/w for the Ø8×4.5 mm construct at day 42. Total collagen accumulation increased from a value of 0.31%±0.01% w/w for the Ø4×1.5 mm construct to 1.62%±0.17% w/w for the Ø8×4.5 mm construct. Reducing the external oxygen tension to 5% O2 resulted in a decrease in DNA content for Ø6×4.5 mm constructs (Fig. 5D). In addition, reducing the external oxygen tension led to an increase in sGAG accumulation in both Ø5×3 mm and Ø6×4.5 mm constructs (Fig. 5E, F). Higher collagen accumulation was also observed in the Ø5×3 mm constructs when cultured at 5% O2, but not in the Ø6×4.5 mm constructs (Fig. 5E, F).
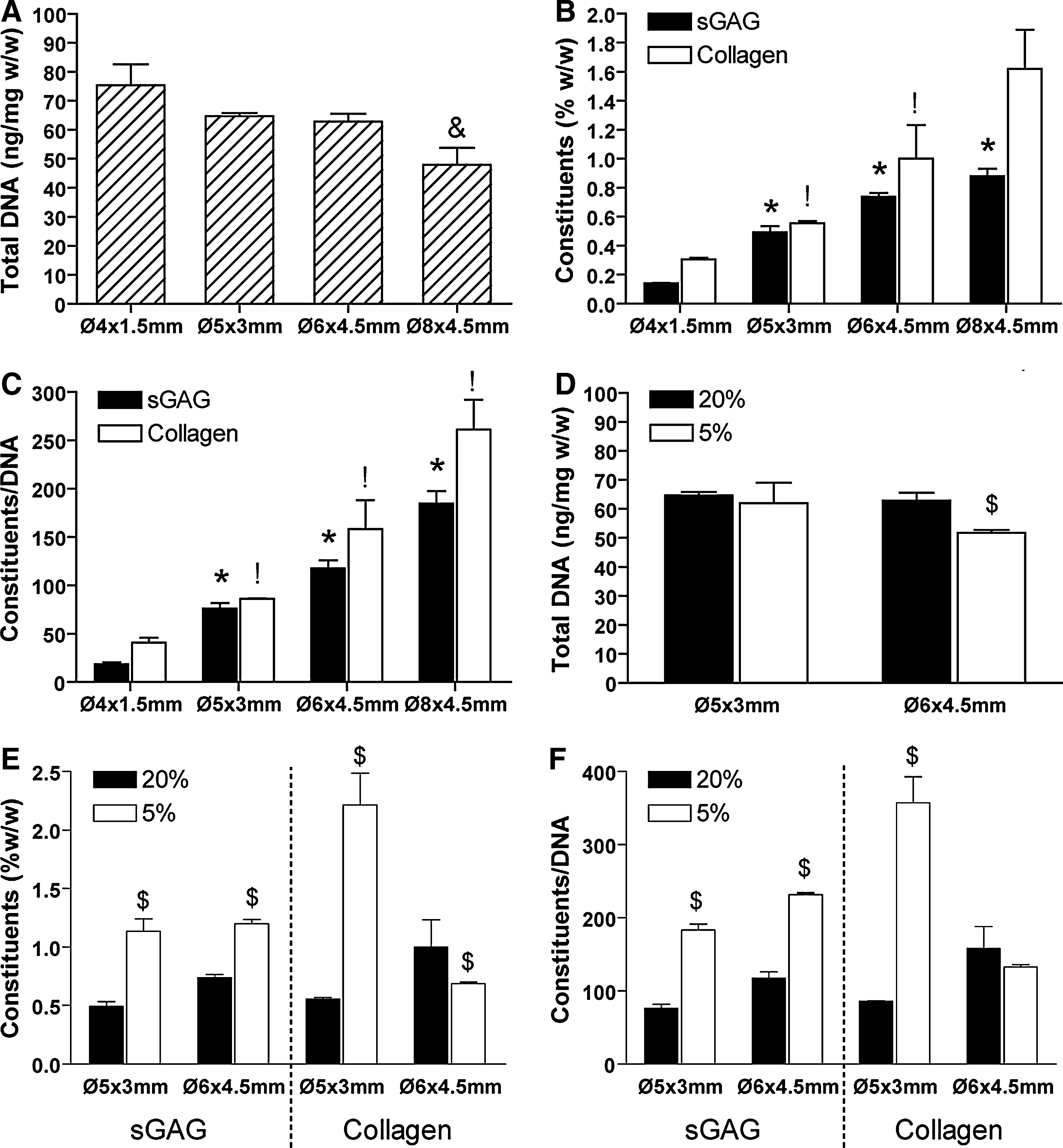
Biochemical accumulation of engineered cartilaginous constructs (Ø4×1.5 mm; Ø5×3 mm; Ø6×3 mm; Ø8×4.5 mm) at day 42 cultured under normoxic (20%) and low oxygen (5%) conditions.
The entire experiment was replicated by using MSCs isolated from a second donor, with identical findings observed in relation to the influence of both construct scale and external oxygen tension, albeit with higher overall levels of matrix accumulation observed in the replicate study (see Supplementary Fig. S1; Supplementary Data are available online at www.liebertonline.com/tea).
All but the smallest Ø4×1.5 mm constructs were sectioned into smaller subregions to quantify the spatial changes in both DNA content and the accumulation of extracellular matrix components (Fig. 6 and Table 1). DNA content was generally lower in the bottom core of all construct sizes, with no evidence of enhanced cell viability observed in constructs maintained at lower oxygen tensions (Fig. 6A and Table 1). sGAG accumulation was generally highest in the bottom core, decreasing toward the construct periphery. These spatial differences were maintained, or even accentuated, when sGAG levels were normalized to DNA content. sGAG accumulation was greater in the bottom core compared with the top core for the Ø5×3 mm and Ø6×4.5 mm constructs (p<0.0001), with near comparable levels in the top and bottom cores of the larger Ø8×4.5 mm constructs (Fig. 6B and Table 1). There was less spatial variation in sGAG accumulation in constructs maintained at 5% O2 compared with 20% O2, although sGAG accumulation was still higher in the core compared with the annulus.
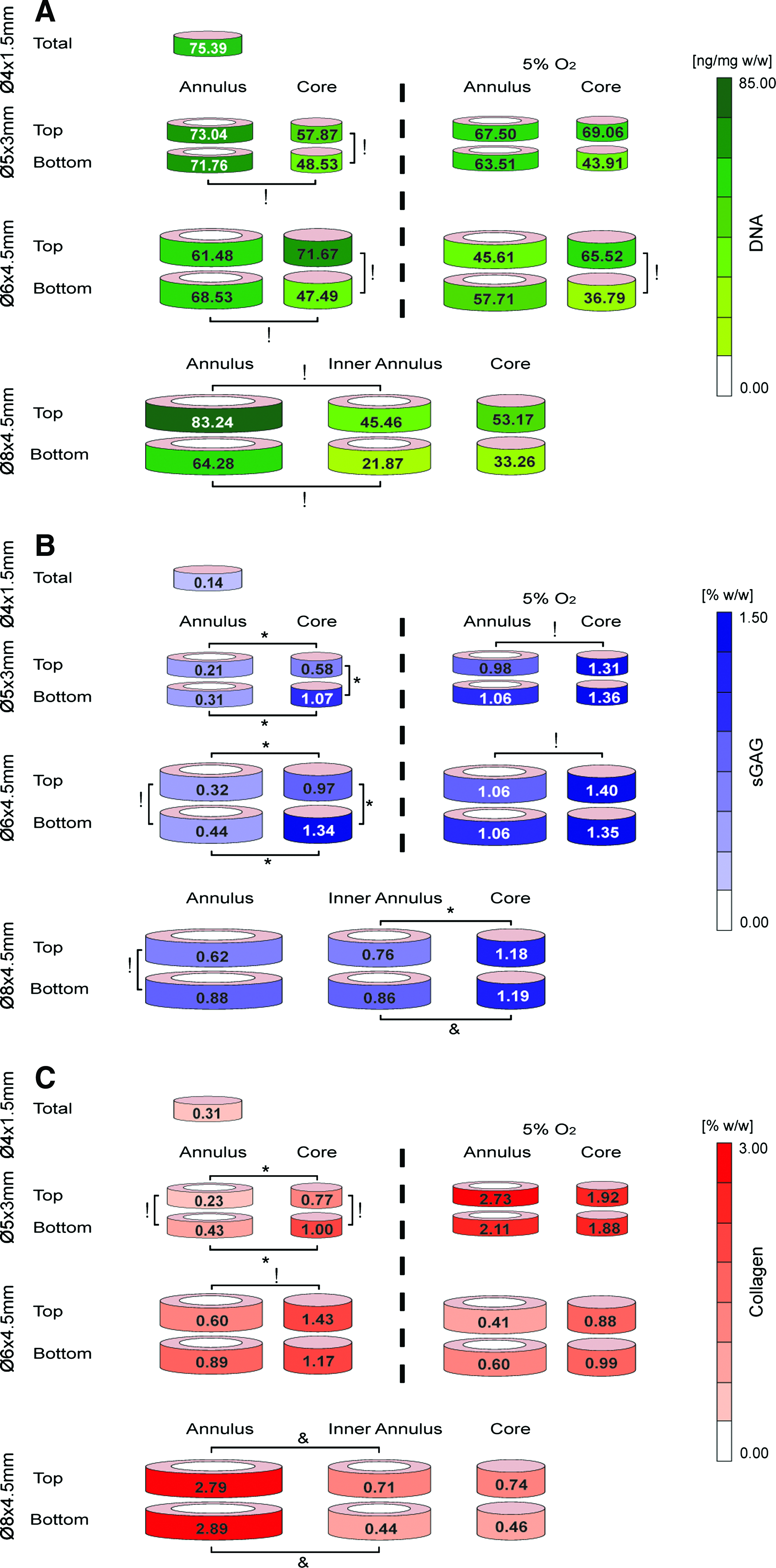
Spatial accumulation of
sGAG, sulfated glycosaminoglycan.
Collagen accumulation was also higher in the core of the intermediate-sized constructs; however, it was lower in the core for the larger Ø8×4.5 mm constructs (Fig. 6C and Table 1). In fact, collagen accumulation was highest in the outer annular regions of the Ø8×4.5 mm constructs compared with both the inner annulus and core regions. These differences were less dramatic, although still present, when collagen levels were normalized to DNA content (Table 1). There was less spatial variation in collagen accumulation in the Ø5×3 mm constructs cultured at 5% O2 compared with 20% O2, with collagen accumulation higher in the annulus compared with the core. Collagen accumulation was reduced in all regions of the Ø6×4.5 mm constructs when cultured at 5% O2.
Structure-function relations
The equilibrium modulus of the Ø8×4.5 mm constructs (36.52±2.12 kPa) was significantly (p<0.05) higher than all other sized constructs (Fig. 7A). The dynamic modulus was observed to increase with increasing construct size, reaching a maximum value of 327.5±30.3 kPa for the Ø8×4.5 mm constructs (Fig. 7A). Culture in a low (5%) oxygen environment was also observed to increase both the equilibrium and dynamic mechanical properties of the engineered constructs (Fig. 7B, C), with a 1.9-fold increase in the dynamic modulus for the Ø5×3 mm constructs when maintained at 5% oxygen tension (Fig. 7C).
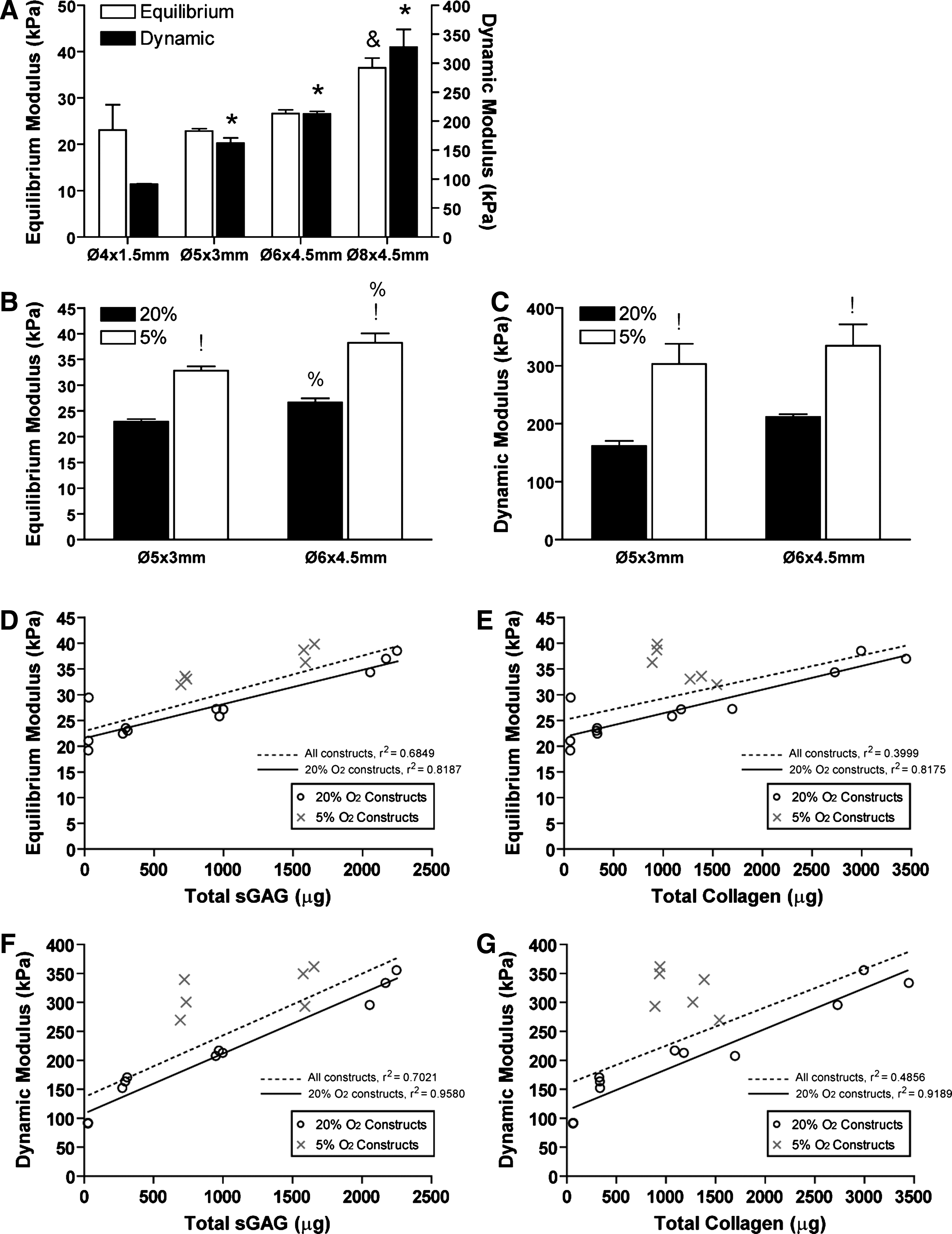
Structure-function plots (Fig. 7D–G) relating both the equilibrium and dynamic modulus of the engineered tissues to biochemical constituents revealed strong correlations. This analysis considered the effects of including all constructs (those maintained at both 20% and 5% O2) or the 20% O2 constructs only in the regression analysis. For equilibrium modulus, when all constructs are considered (20% and 5% O2), a correlation existed for sGAG content (r 2 =0.68) but not collagen content (r 2 =0.40). These correlations became stronger when only constructs maintained at 20% O2 were considered in the analysis (sGAG: r2=0.82, collagen: r2=0.82). This suggests that for the same total amount of matrix components, constructs cultured at 5% O2 appear stiffer. Similar trends were seen for the dynamic modulus correlations.
Discussion
Engineering cartilaginous tissue of a scale necessary to treat defects observed clinically is a significant challenge in the field of cartilage tissue engineering. The results of this study suggest that the inherent capacity of bone marrow-derived MSCs to form large cartilaginous templates in low oxygen environments can be manipulated to engineer tissues of the scale necessary to treat large cartilaginous defects. The mechanical properties of the engineered tissues were observed to increase with increasing construct scale, reaching a maximum value for the Ø8×4.5 mm engineered tissues. Comparable values of equilibrium and dynamic modulus were obtained for smaller constructs maintained at lower oxygen tensions, thus demonstrating that it is possible to recapitulate the chondrogenic oxygen microenvironment which develops within larger constructs in smaller constructs by altering the external oxygen tension.
A simplified model of oxygen diffusion and consumption was used to predict spatial gradients in oxygen concentration throughout the construct. The validity of this model strongly depends on the value of the reaction rate, R, inputted into the model. This term was based on the average reported oxygen consumption rate for bone marrow MSCs undergoing chondrogenesis in a pellet culture system over 21 days; 61 however, this term may depend on multiple factors, including the material in which the cells are encapsulated 62 and the local levels of glucose and lactic acid.63–65 For higher oxygen consumption rates, the model predicts lower oxygen concentrations in the cores of MSC-seeded constructs. For example, if the oxygen consumption rate (Vmax) is increased by a factor of two, then the oxygen concentration in the bottom core of the larger Ø8×4.5 mm constructs reduces to nearly 2.6%; however, for this consumption rate, the oxygen concentration is predicted to reduce to 0.26% in the smaller Ø5×3 mm constructs maintained in low oxygen conditions. Therefore, the model utilized in this study will always predict a lower oxygen tension in the core of the smaller Ø5×3 mm constructs maintained in low oxygen conditions compared with the larger Ø8×4.5 mm constructs maintained in normoxic conditions for reasonable values of R (The potential implications of this are discussed in the next paragraph.). Direct measurements are needed to confirm these predictions, as oxygen consumption rates are known to increase with low glucose availability, 64 which may exist in the core of the larger constructs. A further assumption of the model is that the diffusivity of the construct to oxygen is constant. Clearly, as large amounts of extracellular matrix are laid down, the diffusivity of the tissue to oxygen will reduce, thus further reducing core oxygen levels.
sGAG accumulation was always highest in regions of the construct where oxygen concentration was predicted to be lowest. Reducing the ambient oxygen tension to 5% O2 increased sGAG accumulation in nearly all regions of the Ø5×3 mm and Ø6×4.5 mm constructs. Therefore, sGAG synthesis would appear to be inversely correlated with oxygen tension (see Supplementary Fig. S2), although this may not be the only determinant of synthetic activity. A more complex relationship exists between collagen synthesis and spatial levels of oxygen concentration (see Supplementary Fig. S2). Collagen accumulation is greater in the core of intermediate-sized constructs, where oxygen concentration is clearly lower; however, for larger constructs (Ø8×4.5 mm), collagen accumulation was greater in the annular region. Further, maintaining the Ø6×4.5 mm constructs at 5% O2 was observed to suppress collagen synthesis. The disparity in the spatial accumulation of sGAG and collagen is particularly evident in the larger Ø8×4.5 mm constructs. Although an oxygen tension below 20% O2 is clearly beneficial for collagen synthesis, the fact that a lower oxygen microenvironment in the larger Ø8×4.5 mm enhances sGAG but not collagen accumulation suggests that environmental conditions which promote sGAG synthesis are not necessarily optimal for collagen synthesis, and that gradients in other regulatory factors are playing an equally important role in determining rates of collagen synthesis. For example, collagen synthesis could be more sensitive to nutrient availability than sGAG synthesis, which may explain why collagen accumulation is lower in the core of the larger Ø8×4.5 mm constructs compared with the core of the smaller constructs maintained at 5% O2, where predicted oxygen concentrations are lower but where nutrient availability is presumably greater. An additional factor to consider may be the consumption rates of nutrients such as glucose, which have been shown to increase in low oxygen environments. 65 In addition, it has been shown that cellular oxygen consumption increases at low glucose concentrations,59,63,64 thereby increasing the likelihood of anoxic, nutrient deficient regions developing within the cores of large tissue engineered constructs under certain conditions. Further studies are required to quantify gradients in oxygen and nutrient concentrations in large MSC-seeded constructs. Finally, it should be noted that spatial differences in collagen accumulation are not as dramatic when normalized to DNA content, thus suggesting that the lower levels of collagen within the cores of larger constructs can be partially attributed to lower cell viability in these regions.
As expected, both the equilibrium and dynamic modulus of the engineered tissues were strongly correlated to the sGAG content. Interestingly, for comparable levels of extracellular matrix accumulation, constructs maintained at 5% O2 appeared stiffer than those maintained at 20% O2. This can be potentially explained by the fact that matrix is more homogenously accumulated in constructs maintained at 5% O2, whereas a greater proportion of sGAG accumulates in the core when maintained at 20% O2. Inhomogeneous accumulation of extracellular matrix can negatively impact the magnitude of material constants derived from mechanical tests that assume a homogenous tissue. 66 Alternatively, a lower oxygen tension could potentially alter the ratios or amounts of different types of collagen being generated by MSCs, or collagen cross-linking, all of which are known to impact the mechanical properties of tissue engineered cartilage.67,68
There are numerous studies reported in the literature comparing the chondrogenic capacity of stem cells isolated from different tissue sources22,24,25,69–72 and/or comparing stem cells to chondrocytes for cartilage tissue engineering.19,26,27,73,74 We have previously shown that stem cells and chondrocytes seeded into agarose hydrogels respond differentially to variations in oxygen tension. 26 Coupled with the results of this study, this suggests that comparisons between different cell types should be made with caution, as for a given scaffold type and cell seeding density, chondrogenesis will be dependent on the geometry of the construct. For example, it has been shown that the functional properties of cartilaginous tissues engineered using chondrocyte-seeded hydrogels or scaffolds are superior to those generated using MSCs for small constructs,19,38 but whether this result would translate to larger constructs is unclear. Ideally, comparisons would be made by using a range of scaffold geometries.
The oxygen microenvironment within MSC-seeded constructs will depend, among other factors, on the scaffold geometry and architecture, the local concentrations of glucose and lactic acid, cell-seeding density, and the external oxygen tension. In this study, we have demonstrated that cartilaginous extracellular matrix accumulation within MSC-seeded agarose hydrogels increases with increasing construct size, and we have provided indirect support for the hypothesis that this is due to a reduced oxygen concentration within these larger engineered tissues. This led to increases in the mechanical functionality of the engineered tissue, with both the equilibrium and dynamic modulus shown to be strongly correlated with the sGAG content within the engineered tissue. As the size of the engineered construct increased, different regions of the construct preferentially supported either sGAG or collagen accumulation, thus suggesting that gradients in regulatory factors other than oxygen were also playing a role in determining levels of matrix synthesis. The results of this study demonstrate that all these factors should be tailored to the geometry of the tissue being engineered to enhance chondrogenesis within the developing construct.
Footnotes
Acknowledgments
This study was supported by a President of Ireland Young Researcher Award from Science Foundation Ireland (08/YI5/B1336) and by an IRCSET EMPOWER scholarship.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.