
Abstract
To evaluate effects of motor neurons prostheses (MN-prosthesis) fabricated with MNs differentiated from neural stem cells of fetal spinal cord seeded in self-assembling peptides (sapeptides) scaffolds on functional recovery after spinal cord injury (SCI). Spinal cord hemisection was performed in adult rats. The MN-prosthesis was implanted into the lesion of injured spinal cord. TUNEL staining showed that the MN-prosthesis could decrease the number of apoptotic cells at 2, 4, and 7 days after transplantation. In the MN-prosthesis group, some cholinergic neurons survived within and around the implant, more neural fibers went through the implant from rostral side to caudal side. On day 84, Basso, Beattie and Bresnahan (BBB) score, the peak latencies and amplitudes of N1 wave in both motor evoked potential (MEP) and cortical somatosensory evoked potential (CSEP) on the ipsilateral hind limb in the MN-prosthesis group were also significantly greater than those in the other groups (p<0.05). Thus, implantation of MN-prosthesis increases survival of the damaged cells, promotes formation of connection of neural fibers between the regenerative axons and the host tissue, and improves motor and electrophysiological functions. These findings demonstrate that MN-prosthesis serves as a potential tool for restoring neurologic function after SCI.
Introduction
However, axons can regrow in unscarred regions and traverse the injured area.6,7 Further, functional recovery can be significantly enhanced even if a small amount of normal tissue survives. 8 Although blocking inhibitory molecules and antagonizing secondary injury, 9 delivering small molecules and growth factors, 10 replacing myelin by stem cells, 11 and implanting fetal tissues 12 and scaffolds 13 have been shown to promote axonal regrowth and improve functional recovery, 14 currently, no effective treatment is available for SCI patients.
Rapid progress has been made in the field of repairing injured spinal cords using transplantation of neural stem cells (NSCs)15,16 and tissue engineering technology. 9 Because NSCs have the ability to proliferate and self-renew and have multipotentiality in both mammal embryonic neural tissue and the adult CNS, 17 transplantation of NSCs to the injured spinal cord has been applied to replace lost neurons and create a beneficial environment for neural regeneration. However, some articles showed that only little transplanted NSCs were differentiated into neurons.18,19 There were some contrary articles in which authors reported substantial neuronal differentiation of transplanted NSCs.20,21 Moreover, the proliferation of glial cells derived from NSCs (host or graft) forms a glial scar, which not only could isolate the foreign cells to protect the tissue but also might become a physical barrier for axonal growth and secrete inhibitory factors,22–25 thereby preventing axonal regeneration. Thus, it is difficult to determine how to promote the differentiation of transplanted NSCs into neurons in the injured spinal cord, to inhibit the proliferation of the glial scar, and to build bridges for neuronal regeneration.
Previous studies have shown that motor neurons (MNs) can extend their axons and form constructional conjunction with their muscle targets to contribute to improved functional recovery.26,27 Therefore, in this study, we chose the differentiated MNs as seeding cells instead of NSCs to avoid the disadvantages of NSCs.
The extracellular matrix is a crucial structural component for supporting cells and provides a permissive microenvironment that influences the function of cells. Numerous short sequences of proteins located in the extracellular matrix recently have been identified as playing important roles in nerve regeneration, survival, attachment, proliferation, migration, and differentiation of neural stem/progenitor cells.28–30 These short sequences of proteins can form hierarchical supramolecular structures in special conditions. 31 Further, they can undergo spontaneous assembly into nanofibers and scaffolds that are 10 nm in diameter with pores ranging from 5 to 200 nm and contain 99% water.32,33 Self-assembling peptides (sapeptide) have been considered for application in SCI treatment because they create a permissive environment for axonal extension by means of a synthetic biological nanofiber network of material that connects the proximal and distal stumps of the lesion. This connection allows the migration of cells and processes into the scaffold while simultaneously prevents the scar formation that normally occurs in early stages of CNS injury,29,34 and it also promotes rapid and selective differentiation of neural stem/progenitor cells into neurons. 30 Sapeptide has extraordinary biocompatibility, porousness, and mechanical properties.28,35,36 Thus, in the present study, we hypothesized that implantation of a degradable and synthetic scaffold of a specified sapeptide seeded with MNs to the damaged spinal cord in adult rats after spinal cord hemisection would improve axonal regeneration and functional recovery.
Materials and Methods
Scaffold fabrication
Powdered sapeptide (RAD16-II, AcN-RARADADARARADADA-CNH2, purity through HPLC >99%; GL Biochem) was dissolved in deionized water to obtain a saturated solution as previously described. 35 The scaffolds were fabricated by blending equal volumes of sapeptide solution and 0.9% (w/v) sodium chloride solution until they reached the size of 4×4×2 mm. The scaffolds were removed, trimmed, assembled, and stored in a vacuum desiccator until use.
Differentiation of embryonic spinal cord stem/progenitor cells and identification of spinal cord MNs
All animals used in this study were handled according to the Guidelines of Chongqing Association for Animal Care and Use (SCXK Yu20020003) and the Research Institute of Surgery for the management of laboratory animals. Animal Ethic for this study was approved by the Third Military Medical University. Fetal Wistar rats (embryonic 14 days) were used in the experiment. The single-cell suspension of the spinal cords was collected. The collected cells were dissolved with neurobasal medium (Gibco) and plated at the density of 2.5×103/cm2 in culture medium with N2 supplement and basic fibroblast growth factor (bFGF, 20 ng/mL; Gibco). The third passage cells were plated on circular coverslips coated with polylysine (Sigma) at the density of 2×105/cm2 and cultured with neurobasal medium (Gibco) with N2 supplement and inductive factors, including retinoid acid (RA, 2000 nM; Sigma) and sonic hedgehog (Shh, 300 nM; R&D). Half of the medium was replaced every other day. Fetal bovine serum (5%) was added to all cultures 12 days after treatment. Eighteen days after culture, differentiated cells were identified with antibody of neurofilament 200 (NF-200) and vesicular acetylcholine transporter (VAChT) and harvested with 0.125% trypsin (Gibco) from coverslips for seeding. The number of cells was counted after DAPI staining (Sigma). Images were taken using a TCS SP-2 laser scanning confocal microscope (Leica). The number of positive cells in every field (measurement frame of 500×500 μm) was calculated. The software (Analysis; Soft Imaging System GMBH) was processed to facilitate cell identification and to reduce counting errors. The undifferentiated and differentiated NSCs were used for real-time reverse transcription–polymerase chain reaction (RT-PCR).
Real-time RT-PCR
The procedure described by Hou et al. 37 was used for real-time RT-PCR. Total RNA was extracted from undifferentiated NSCs and differentiated NSCs (MNs) with Trizol (Invitrogen). The separation and precipitation of RNA was accomplished with chloroform and isopropyl alcohol. Total RNA was isolated and purified by an Rneasy minikit (Qiagen) with the addition of Rnasefree DNase I (Qiagen). The two-step SYBR® ExScript™ RT-PCR kit (Perfect Real Time; TaKaRa) for real-time RT-PCR was used to analyze the expression of the following trophic factors gene, brain-derived neurophic factor (BDNF), and mitogen gene, bFGF. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the housekeeping gene. Quantification was achieved using standard curves derived from gene expression relative to the level of GAPDH gene expression. The primers used are listed in Table 1.
bFGF, basic fibroblast growth factor; BDNF, brain-derived neurophic factor; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Cellular prosthesis formation
The undifferentiated and differentiated NSCs were labeled with DAPI (50 mg/L) for 1 h at 37°C, respectively. Next, scaffolds were soaked in phosphate-buffered saline (PBS) solution for 24 h to form the cuboid with a size of 4×4×2 mm, and 5×105 cells/mL were seeded using an orbital shaker at 37°C in the incubator. The serum-containing medium was changed the next day, and the implants were incubated for 4 days before implantation.
Spinal cord hemisection
Adult Wistar rats (200–220 g, 10–12 weeks of age) were anesthetized by intraperitoneal injection of pentobarbital sodium (40 mg/kg; Shanghai Pharmaceutical Factory). The model of spinal cord hemisection was prepared according to the description in the article. 38 The spinal cords were exposed by laminectomy at the T12–L1 level of the spine. First, there is a posterior spinal cord vein locating along the midline of the spinal cord, which is determined as a landmark. Second, the spinal dura mater was cut through by a knee No. 11 knife. Third, a homemade knife was used to determine the size of cutting. Last, a No. 11 surgical blade was repeatedly scraped along the ventral surface of the vertebral canal to ensure complete resection, and then suction was used to remove any residual fibers at the lesion site. Ultimately, the injured area on the right side of the lumbar spinal cord was 4 mm (long) ×4 mm (depth)×2 mm (width), which was confirmed by an independent blinded observer. This size of the lesion area was confirmed across all experimental groups and animals.
Experimental groups and animal care
Seventy Wistar rats were randomly divided into five groups as follows: (1) Group A: hemisection alone, n=14; (2) Group B: transplantation of MNs alone, 10 μL of 107 cells/mL suspension were injected into the cavity after hemisection, n=14; (3) Group C: scaffold implant without any cells, n=14; (4) Group D: NSC-prosthesis group, scaffold implant with embryonic NSCs, n=14; (5) Group E: MN-prosthesis group, n=14. The MN-prostheses were made as described above.
Surgeries were performed after a randomized block design. The surgeries for the implantation plus controls were performed on the same day to minimize variation among groups, and the order for the surgery was varied to reduce surgical bias. After making spinal cord lesion and transplantation, the musculature was sutured and the skin was closed. Animals recovered in a clean cage on sterile wound filings. Ringer's lactate solution (10 mL) was given daily for 7 days postoperation. The bladders were evacuated twice daily until reflex function of the bladder was established.
Behavioral assessment
Functional recovery was determined using the locomotor rating scale of Basso, Beattie and Bresnahan (BBB), in which zero indicated complete paralysis and 21 indicated normal mobility. 39 Behavioral tests were performed at the same time of each day and were graded by the same blinded observers. All animals survived and exhibited no signs of autophagia throughout the course of the study.
Evoked potential examination
The left posterior part of the head was shaved and then the skin was sterilized with iodine and 75% alcohol. The skin was incised and exposed, the periosteum of the posterior skull was dissected, and the foramen coecum (3 mm) was drilled using a dentistry drill from 3 mm posterior to the bregma and 1 mm lateral to the midline on the left side of the skull. The right sciatic nerve was exposed as well. The MEP (motion evoked potential) and CSEP (cortical somatosensory evoked potential) were tested using electrical instruments (Powerlab/16SP AD Instruments), and the stimulus parameters were as follows: frequency 4 Hz; duration 0.2 ms; intensity 4.5 mA with a single pulse wave; the signal was magnified 100,000 times with a preamplifer; and the average times was 256. When recording CSEP, the stimulating electrode was placed on the right sciatic nerve, the recording electrode on the surface of the left cortical somatosensory cortex area of the brain, the negative electrode on the lateral scalp, and the reference electrode in the paravertebral muscle. When recording MEP, the stimulus electrode and recording electrode were exchanged, and the stimulus cathode (silver disc) was placed below the hard palate.
Retrograde and anterograde neural tracing
Rats were anesthetized 12 weeks postoperation, and tracing experiment was performed by injecting 30% horseradish peroxidase (HRP) (dissolved in 0.01 M PBS; Sigma) into the right sciatic nerves and 10 mg/mL Dil (dissolved in DMSO; Sigma) into the right cervical spinal cord. Three days (HRP) or 7 days (Dil) after injection, the animals were transcardially perfused with isotonic saline followed by 4% paraformaldehyde in 0.1 M acetate buffer (pH 6.5, 0.8 mL of perfusion solution per gram of body weight). The rostral and caudal areas of the spinal cord and the implant were removed and immersed in 30% sucrose for 48 h. Transverse sections (40 μm for HRP or 20 μm for Dil) through the entire spinal cord and implants were cut on a cryostat. The sections were processed to detect HRP using the tetramethylbenzidine method, 40 and the red Dil-positive neurons were visualized using laser scanning confocal microscopy under excitation of 565 nm.
Using light microscopy at ×100 magnification, the number of HRP-positive neurons in the rostral spinal cord was calculated (n=5) and analyzed. Pictures at high resolution (2776×2074 pixels) were taken with a digital image microscope system (Olympus DP50; Olympus) and were processed using the Soft Imaging System to facilitate cell identification and to reduce counting errors. Every stained cell profile with an identifiable nucleus or with two or more dendrites was counted in every section processed for HRP. The cell count was performed in one field (0.45×0.60 mm) per section, and five sections per animal (n=5 animals) were chosen for quantitative analysis.
Histology, immunocytochemistry, and TUNEL
After perfusion, the spinal cords were carefully dissected, postfixed overnight in 4% paraformaldehyde, and dehydrated at room temperature with graded 50%–100% alcohol and dimethyl benzene. Sections were longitudinally cut in the position of the coronal plane for hematoxylin/eosin (HE) and Holmes staining as described by Wu et al. 41
Immunocytochemistry was performed on 10 μm spinal cord sections incubated with the following antibodies: nestin (1:500, mouse anti-rat monoclonal antibody; Pharmigen); NF-200 (1:1000, mouse anti-rat monoclonal antibody; Sigma); glial fibrillary acidic protein (GFAP) (1:400, rabbit anti-rat monoclonal antibody; Sigma); galactocerebroside (Gc, 1:200, rabbit anti-rat monoclonal antibody, Chemicon); VAChT (1:1500, goat anti-rat polyclonal antibody; Chemicon); and GAP-43 (1:100, goat anti-rat polyclonal antibody; Santa Cruz). All secondary antibodies (Alexa Fluor 594- and FITC-labeled secondary antibodies from rabbit or goat) were purchased from SouthernBiotech. All images were taken with a TCS SP-2 laser scanning confocal microscope.
TUNEL was used to identify the cells exhibiting DNA fragmentation as previously described by Gavrieli et al. 42 The TUNEL assay was performed using the ChromaTide™ Alexa Fluor® 488-5-dUTP (Molecular Probes) and TdT terminal transferase kit (Roche) according to the operation manual. The number of positive cells was counted from sections rostral and caudal to the lesion site and from the contralateral side of the lumbar spinal cord. Five fields per section, five sections per animal, and two animals per group were used for analysis. The measurement frame of each field was 200×200 μm, and then the fields with the random five numbers of positive cells were chosen for analysis, and the average of them was defined as per section. Soft Imaging System software was used to process the data to facilitate cell identification and to reduce counting errors.
Statistical analysis
For statistical analysis real-time RT-PCR data was natural log (LN) transformed to obtain normally distributed data. The paired t-test was used to compare data between MNs and NSCs with the SPSS version 10.0 software package. The Kolmogorov–Smirnov goodness-of-fit test was used to determine the normality (Gaussian-shaped distribution) of the data. The BBB data showed no significant departure from a normal distribution for all time points beyond day 1; therefore, parametric methods were used, including repeated-measures analysis of variance. Bonferroni post hoc analysis was used for multiple group comparisons to determine the statistical significance of the results. Descriptive values were presented as the mean±standard deviation.
Results
Differentiation of MNs from the spinal cord stem/progenitor cells by RA and Shh
Cells were initially cultured in neurobasal medium with N2 supplement and bFGF. We found that these cells proliferated and formed neurospheres 3 days after incubation (Fig. 1), indicating that these cells were proliferative and met one criteria (i.e., proliferation) of stem/progenitor cells. Seven days after differentiation, all of the neurospheres were positive for nestin. Meanwhile, the cells from differentiated neurospheres also expressed NF-200, GFAP, and Gc (Fig. 1), respectively.
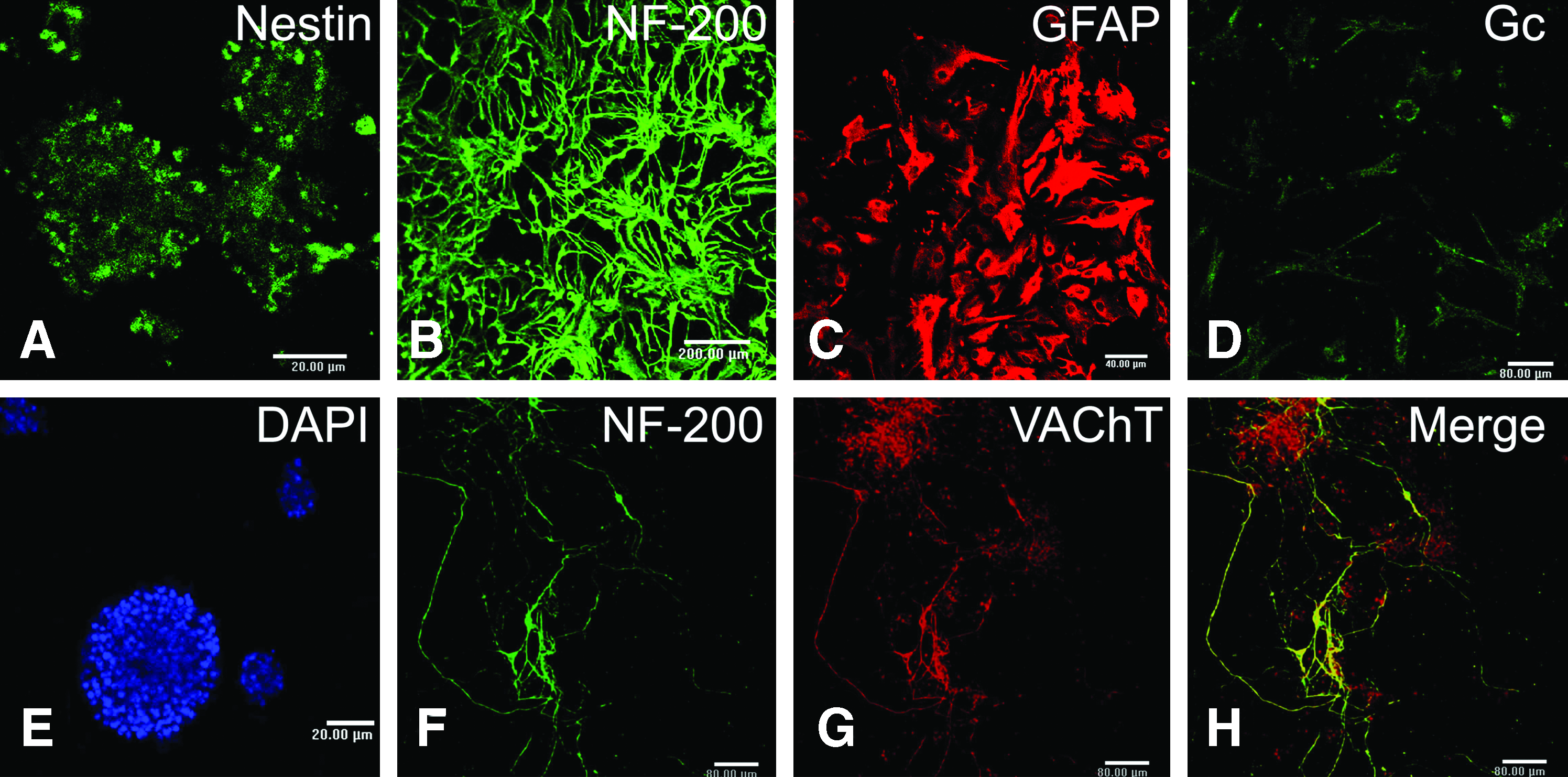
Representative confocal images showing neurospheres from embryonic stage 14 day rat spinal cord and its differentiation into multiple cell types. A representative neurosphere was shown to be positive for nestin
We changed the neurosphere culture medium after three passages with bFGF-free neurobasal medium supplemented with RA, Shh, and N2. On the second day, long and slender neurites extended from the cells in medium with RA and Shh. Immunostaining on the 18th day showed that most differentiated cells (>90%) were double positive for NF-200 and VAChT. This result showed that the differentiated neurons were mostly MNs. The detailed results were shown in Table 2.
RA, retinoic acid; Shh, sonic hedgehog.
The expression of mitogen (bFGF) and trophic factors (BDNF) were detected in NSCs and differentiated NSCs (MNs) (Fig. 2). The level of trophic factors in NSCs was higher than that in MNs. There was a significant difference in expression of bFGF between NSCs and MNs (p<0.01).
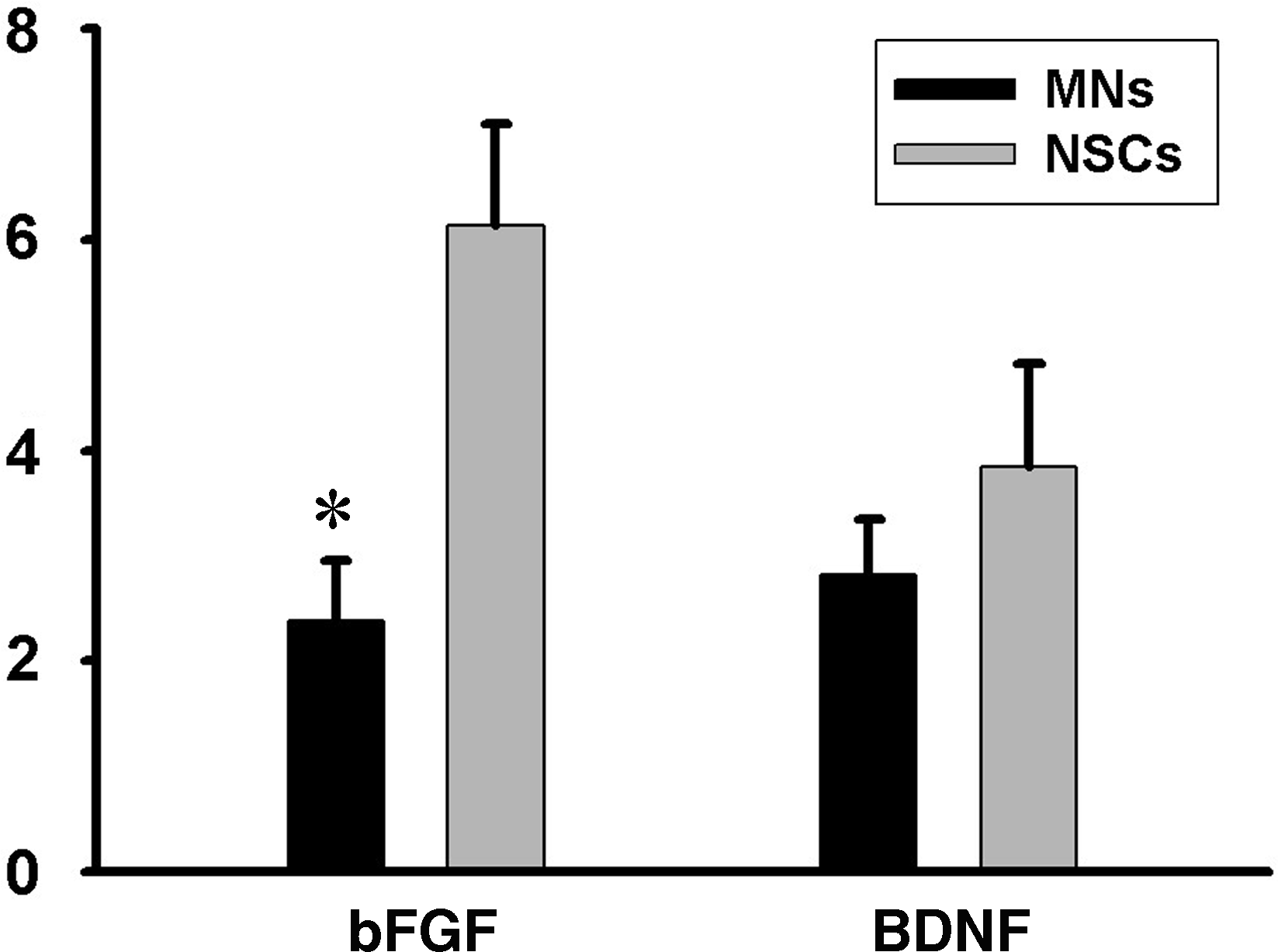
The expression of basic fibroblast growth factor (bFGF) and brain-derived neurophic factor (BDNF) were detected in neural stem cells (NSCs) and differentiated NSCs (MNs) by real-time reverse transcription–polymerase chain reaction. NSCs were cultured with neurobasal medium with N2 supplement, retinoic acid (2000 nM), and Shh (300 nM) for 12 days. Then, fetal bovine serum (5%) was added for another 6-days culture. Finally, the differentiated NSCs (MNs) were harvested. The level of bFGF and BDNF in NSCs was higher than that in MNs. *There was a significant difference in expression of bFGF between NSCs and MNs (p<0.01).
Fabrication of prostheses
The macroscopic scaffolds were fabricated from the RAD16-II in the presence of monovalent salt solution. The cells were seeded on the scaffolds, and the prostheses made of MN and the sapeptide scaffold was observed under inverted and scanning electron microscopes (Fig. 3). We found that MNs extended their neurites in the scaffolds (Fig. 3A, C) and extensively expressed GAP-43 (Fig. 3D). The scanning electron microscope showed that a considerable number of cells were distributed within the prosthesis (Fig. 3B).
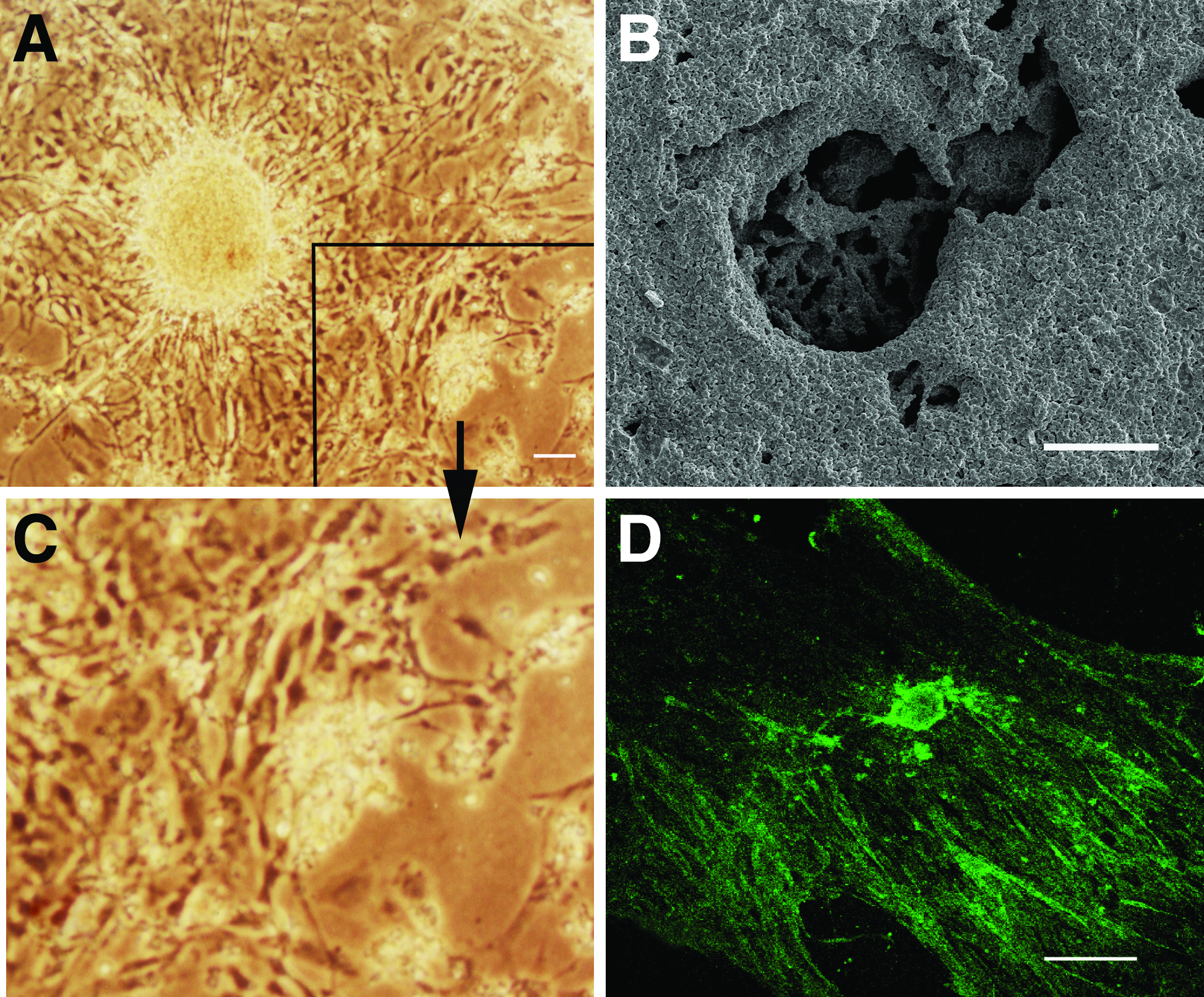
Differentiated MNs were seeded on the sepaptide scaffold. Under the inverted microscope, a cell with processes was observed
MN-prosthesis implantation reduced apoptosis and improved survival of the seeding cells in the host
To assess effects of the transplant on apoptosis in injured spinal cords, the number of apoptotic host cells in rostral and caudal sections and in sections contralateral to the lesion were calculated based on TUNEL-positive cells on days 2, 4, and 7 postoperation (Fig. 4). The number of TUNEL-positive cells in Group E (MN-prosthesis group) was markedly fewer than that in the other groups in the three regions. In addition, there were fewer positive cells in Groups B–E than in Group A (Fig. 4, p<0.05), except that there was no significant difference between Groups B and A in caudal tissue for 2 days postoperation and on the contralateral side tissue for 7 days postoperation (p>0.05).

Apoptosis of host cells was assessed by TUNEL
The expression of NF-200 and VAChT was detected in the rostral, caudal, and contralateral side of the injured spinal cord 7 days postoperation (Fig. 5). Cells colabeled for NF-200 and VAChT were observed in all groups, however, the number of colabeled cells in Group E was far greater than that in other groups.
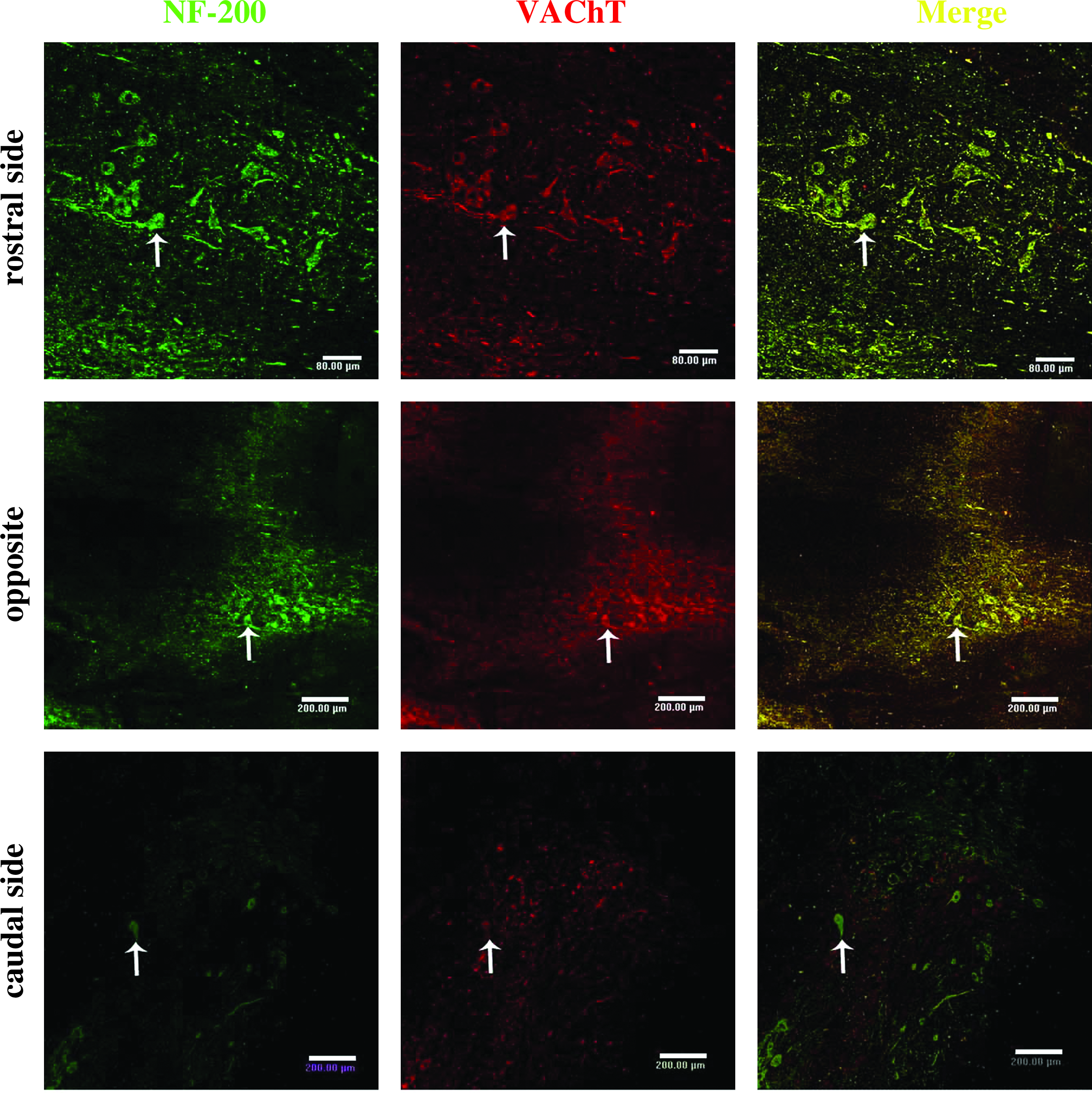
The expression of NF-200 and VAChT were detected in the rostral, caudal, and contralateral sides of the lumber spinal cord in Group E on day 7 postoperation. The double positive cells of NF-200 and VAChT (arrow) were observed in all groups, although the double labeled cells in Group E were significantly more abundant than in other groups. Scale bar: 200 μm.
MN-prosthesis transplantation promoted axonal regeneration after spinal cord hemisection
After spinal cord hemisection, degeneration occurred in the injured spinal cord and cavitation appeared in the injury site. The bigger cavity was formed in the injured group (Fig. 6A). In the MN alone group, some small cavities were present in the operative region of the contralateral side, and the arrangement of regenerative fibers appeared to be disrupted (Fig. 6B). In the scaffold group, there was not remarkable destruction on the contralateral side, and the regenerative fibers were fewer but they were ordered (Fig. 6C). In the NSC-prosthesis group, the tissue on the contralateral side was almost normal, and a number of regenerative fibers were arranged in order on the ipsilateral side of the spinal cord (Fig. 6D). In the MN-prosthesis group, the tissue of the contralateral side was entirely normal, many regenerative fibers were arranged in order on the ipsilateral side, and they connected with the host tissue between the rostral and caudal sides of the ipsilateral spinal cord (Fig. 6E).
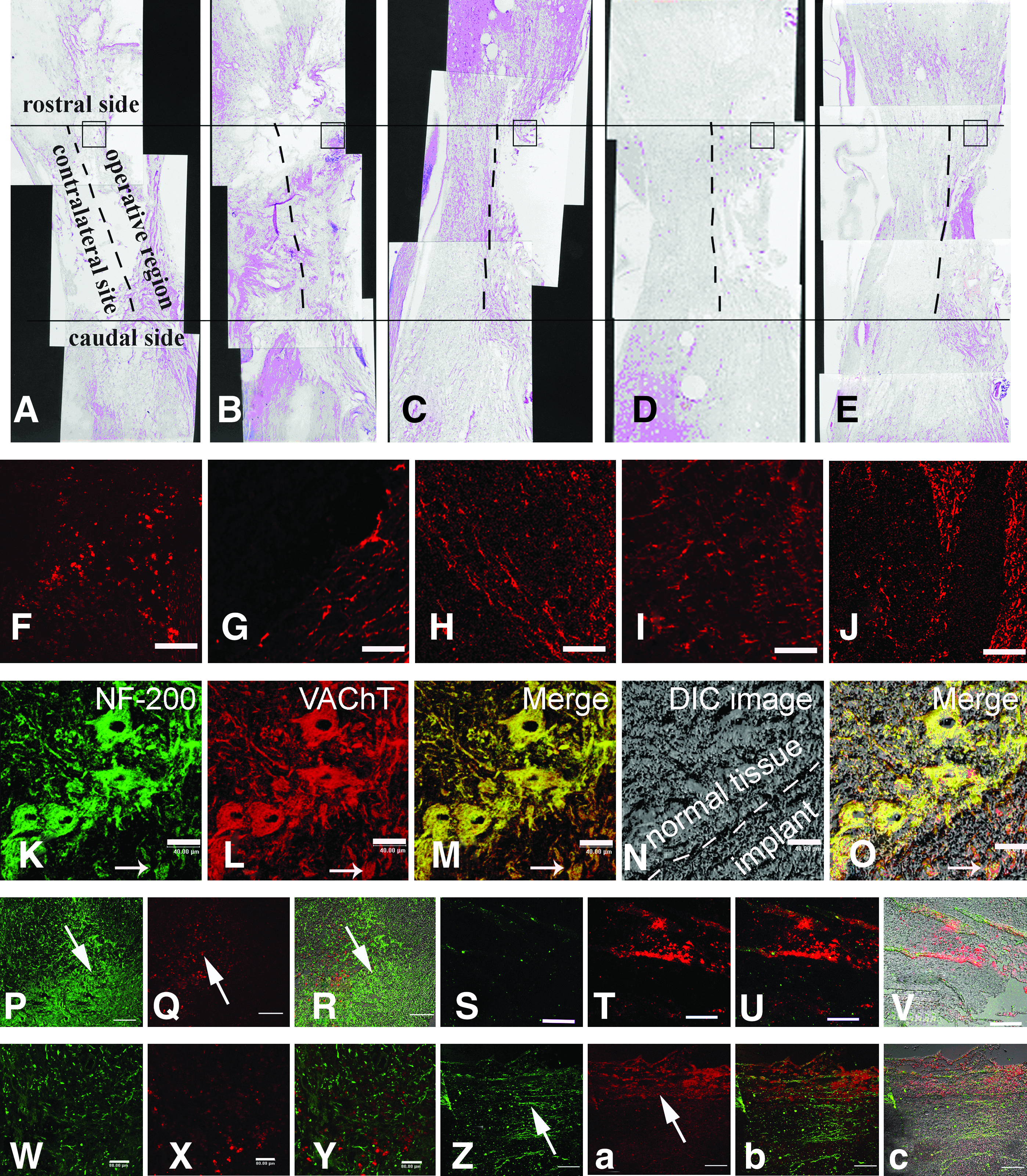
Effect of MN-prostheses implant on axonal regeneration 12 weeks after spinal cord transection. The hemisection model of lumbar spinal cord was made in adult rats with a lesion size of 4 mm (long) ×4 mm (depth) ×2 mm (width). Five different treatments were used in this study.
The nerve fibers expressing GAP-43 were fewer and more disordered in Group A than in other the groups. However, the nerve fibers were organized in order like beams in Group E (Fig. 5J). To observe the morphology of the surviving or regenerated cells in Group E, the expression of NF-200 and VAChT was observed (Fig. 6K–O). We found that the cells that were double labeled for NF-200 and VAChT were present in normal tissue close to the lesion site and the implant (Fig. 6O). Additionally, the glial scar was observed in the graft of Group D (Fig. 6P, R).There was no significant glial scar in Group E (Fig. 6W, Y). VAChT-positive nerve fibers were found in the graft of Group E (Fig. 6Z, b, c). But there was few VAChT positive neurofibra (Fig. 6 S, U, V) in Group D.
To further examine effects of the transplant on axonal regeneration, HRP tracing was performed at 12 weeks postoperation. The results of HRP tracing showed that the labeled neurons could be observed on the rostral (Fig. 7B) and caudal sides of the spinal cord. The number of HRP-positive neurons in Group A was fewer (6.20±1.92) than that in Group B and gradually increased up to 12.40±1.14. The number of HRP-positive neurons in Groups C and D increased remarkably (17.20±2.28 and 20.40±1.14, respectively). In Group E, the number was significantly greater than that in the other groups (p<0.05), reaching up to 25.80±1.30 (Fig. 7D). The long extensive nerve fibers of HRP-positive neurons were clearly visible, and some of them extended into the implants.
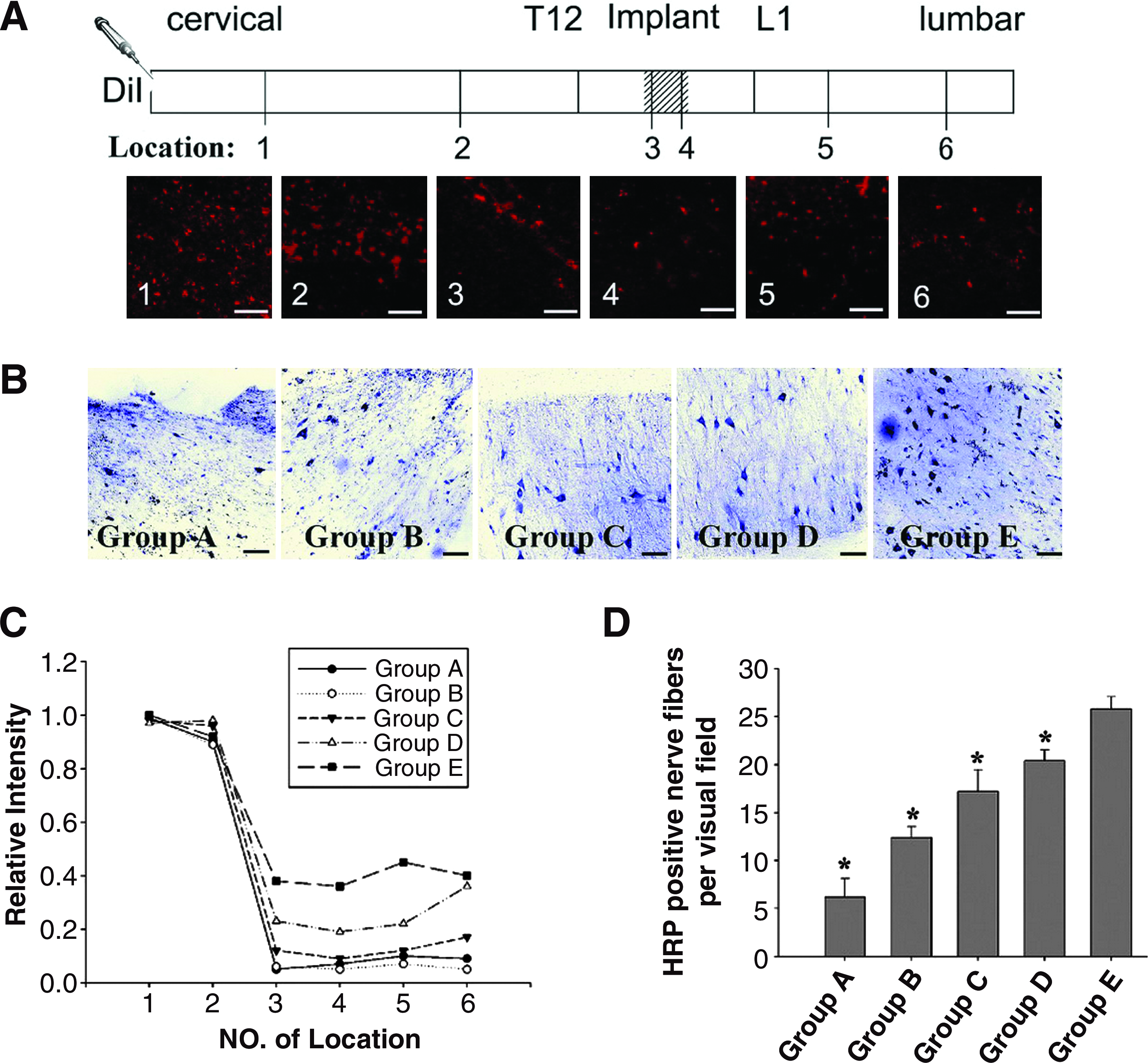
Effect of a MN-prosthesis on regenerative fibers communication after spinal cord hemisection. The tracing experiment was performed by the injection of 10 mg/mL Dil into the ipsilateral cervical spinal cord
In addition to HRP tracing, the application of Dil to the cervical spinal cord resulted in the expected labeling of fibers that extended throughout the implant and ended in the lumbar region (Fig. 7A). Dil-labeled fibers appeared in the right rostral and caudal regions of the spinal cord and within the implant. An abrupt decrease in the number of Dil-labeled fibers originating from the rostral area of the spinal cord was noted in the implant and around the caudal part of the spinal cord. As expected, few axonal projections originating from the proximal uninjured spinal cord were observed in the newly formed tissue in the area of the implant. However, the numerous Dil-labeled fibers originating from the center of the regenerated tissue in the implant extended to the caudal region of the spinal cord (Fig. 6Q, T). There was no significant difference in the number of Dil-positive fibers among the different groups in the right rostral region of the spinal cord close to the injection site of Dil (Location of 1 and 2 in Fig. 7C). However, in the right caudal region of the spinal cord and in the implant (Location of 3–6 in Fig. 7C), the number of Dil-labeled fibers in Group E was greater than that in the other groups (p<0.05).
MN-prosthesis transplantation improved evoked potentials after spinal cord hemisection
Figure 7 showed the peak latency and amplitude of N1 in both MEP and CSEP at 4, 8, and 12 weeks postoperation. The latency of MEP (Fig. 8A) and CSEP (Fig. 8C) gradually decreased with time, however, the amplitude of MEP (Fig. 8B) and CSEP (Fig. 8D) gradually increased. In every group, the recovery of latency and amplitude of N1 was more remarkable before 8 weeks postoperation, and it became relatively slow from 8 to 12 weeks postoperation. The amplitude and latency of MEP/CSEP of Groups E, D, and C surpassed those of Group A and B, as shown by higher amplitude and shorter latency (Fig. 8E). At 12 weeks postoperation, Group E was superior to those of other groups in the amplitude and latency of MEP (p<0.05).
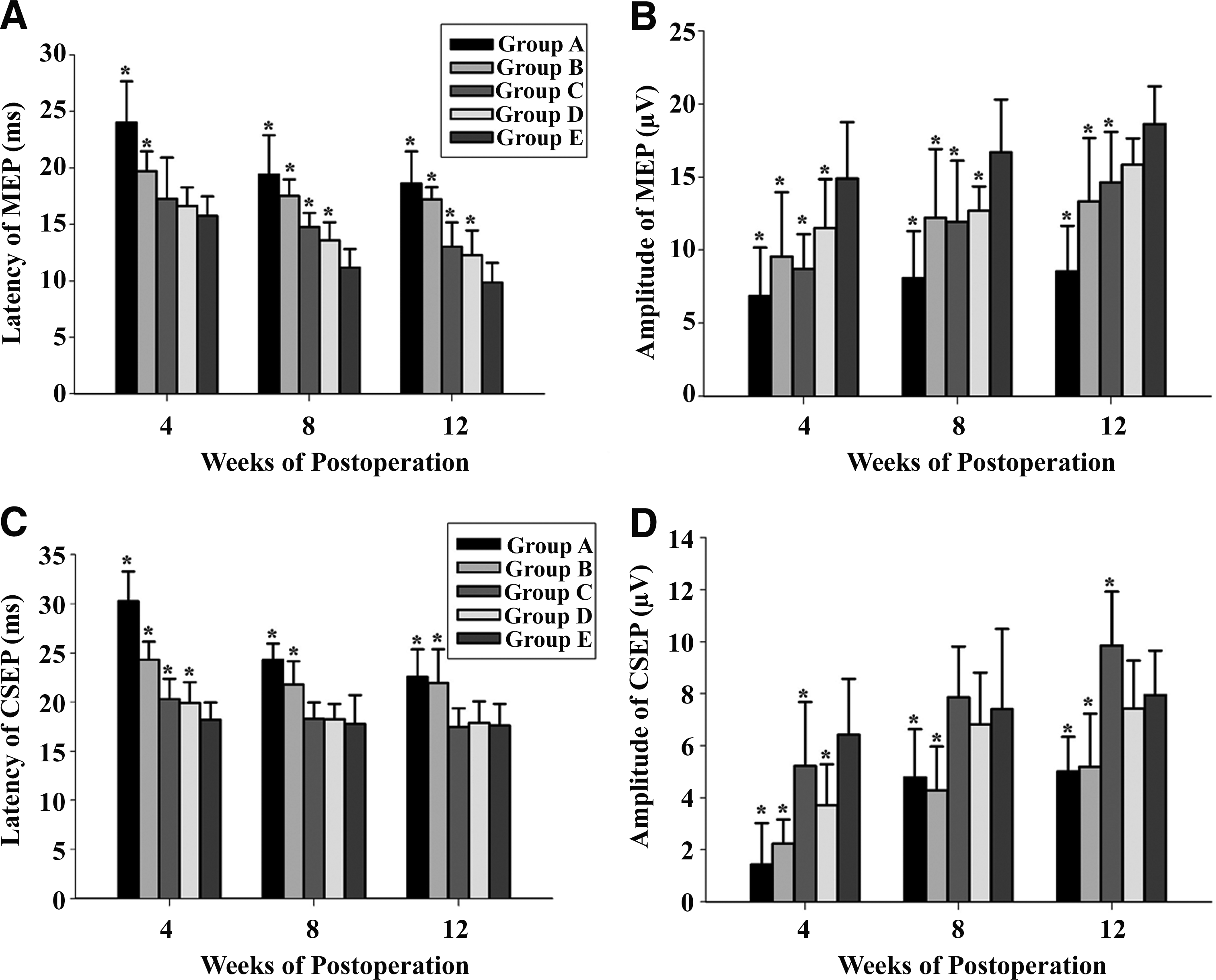
Quantitative analysis of peak latency and amplitude of N1 in both MEP and CSEP at 4, 8, and 12 weeks post-treatment
MN-prosthesis transplantation enhances motor functional recovery after spinal cord hemisection
On the ipsilateral side (Fig. 9), the functional recovery of rats in Groups A and B was extremely limited within 7 days postoperation. At 84 days postoperation, the BBB score was ∼3 and 5 in Groups A and B, respectively. In contrast, the BBB scores from Groups C to E increased significantly from 4 days postoperation. The tendency for rapid increase lasted until 28 days postoperation. After 28 days, the increase in BBB scores became slow. At 84 days postoperation, the score of Group E was the highest (∼12), and a significant difference existed between Group E and the other groups (p<0.05). For the control groups, Group D had the highest mean BBB score (9.70±1.25).
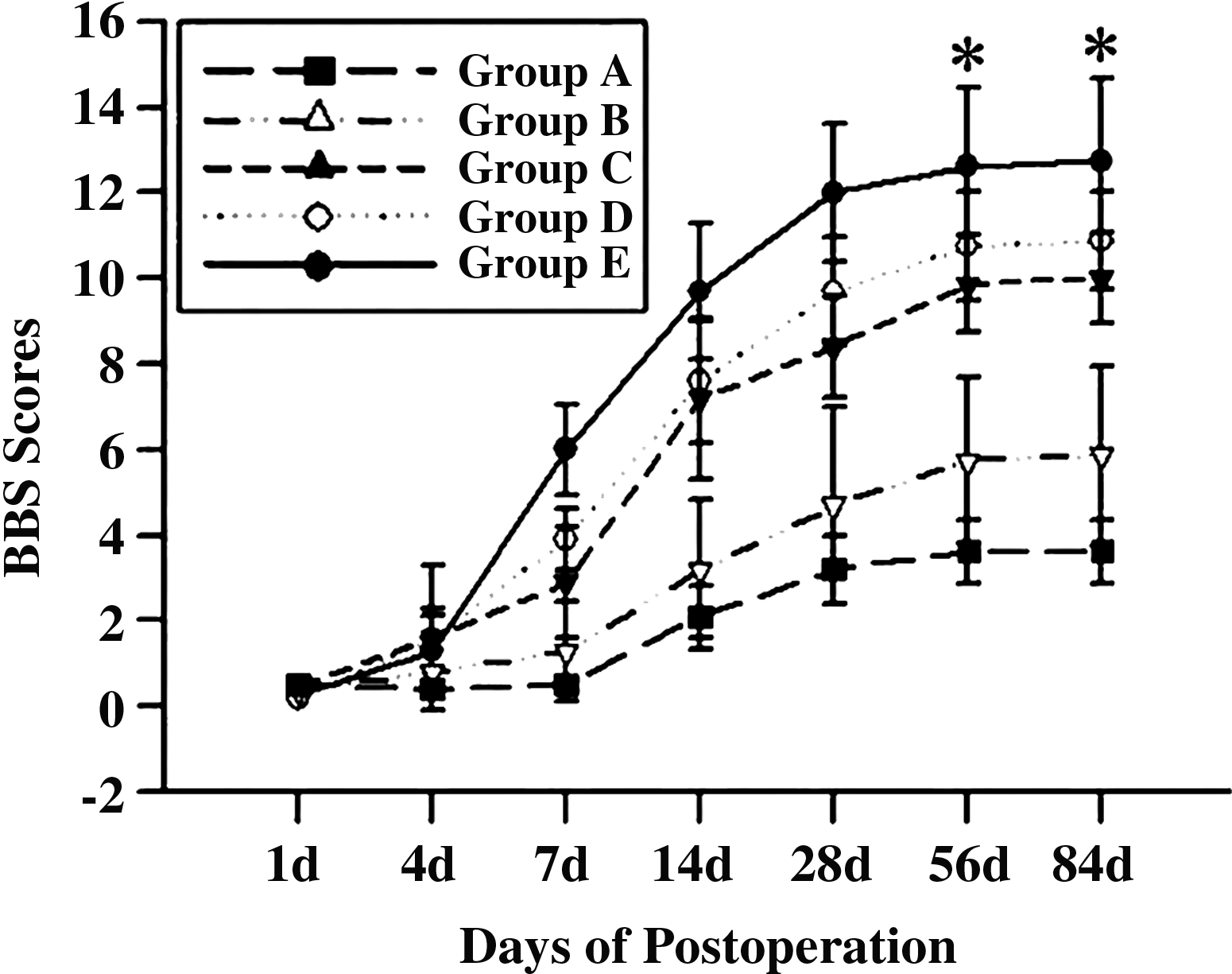
Effect of a MN-prosthesis on functional recovery as assessed by Basso, Beattie and Bresnahan (BBB) scores. At 4 days postoperation, the scores of Group E and Group D were remarkably increased compared with other groups. From day 7 to 84, the scores from the MN-prosthesis group were significantly higher than those from the other controlled groups. The greatest difference in scores existed between the MN-prosthesis group and other groups at 56 and 84 days postoperation (*p<0.05). There was no significant difference between Group D and Group C at all time points tested.
Discussion
In this study, we showed that the lesioned spinal cord of rats could be partially repaired using tissue-engineered MNs from fetal rat spinal cords. We used a cellular prosthesis composed of MNs suspended in a Sapeptide scaffold. We found that the MN-prosthesis occupied the damaged spinal cord, decreased the number of apoptotic cells in the surrounding tissues of the cavity, and promoted axonal regeneration after spinal cord hemisection. More importantly, the rats with an implanted MN-prosthesis exhibited improved functional recovery, as shown by the coordinated gait in the hind limbs and the close to normal motor and evoked potentials.
SCI induces a series of endogenous biochemical changes that lead to secondary damage, such as apoptosis. 43 Apoptosis of transplanted cells in implants has been extensively studied.39,44 Many studies have shown that only a very small amount of the transplanted cells (∼1%–2%) could survive beyond the initial 10 days if implanted immediately after SCI. 44 Conversely, delayed transplantation could improve the survival of transplanted cells in the lesioned spinal cord.39,44 In this study, we used immediate transplantation after SCI to observe the effect of the implants on the host cells; however, the apoptotic rate of transplanted cells in this study was similar to those of previous studies (data not shown). Compared with the spinal cord hemisection alone group, all other groups had fewer apoptotic cells and the difference in the number of apoptotic cells among them was significant in most phases (p<0.05), suggesting that MNs, sapeptide scaffolds, NSC prostheses, and MN-prostheses could decrease apoptosis. However, further comparison showed that sapeptide scaffolds, NSC prostheses, and MN-prostheses exhibited better capability for anti-apoptosis than the transplantation of MNs alone, indicating the importance of three-dimensional scaffolds in repairing spinal cord lesions, especially column-defect lesions.19,45,46 One possible reason for this finding is that scaffolds serve as a bridge to prevent the collapse of adjacent tissue and secondary degeneration caused by spinal cord hemisection. The prostheses surpassed the scaffolds in anti-apoptosis ability, and the cells in the prostheses possibly produced a marked effect.47,48 On day 7 postoperation, cells colabeled for NF-200 and VAChT were observed in the rostral, caudal, and contralateral side of the lumbar spinal cord injured region in all groups. However, there were more MNs in transplanted region in Group E, which might be the reason of promoting recovery after SCI.
Spinal cord hemisection induced considerable degeneration and necrosis on the contralateral side of the lumbar spinal cord. Nevertheless, we found limited spontaneous axonal regeneration in the injured spinal cord, as shown by HE staining and the expression of GAP-43. This result is consistent with previous studies.49,50 Although severe degeneration and necrosis occurred in the lumbar region of the MN group, greater regeneration was observed in the transplanted region, suggesting that transplanted MN could promote axonal regeneration. However, when compared with the scaffold and prosthesis groups, severe degeneration and necrosis occurred on the contralateral side of the spinal cord in the MN group, and the regenerated tissue was disordered, which might be due to lack of scaffold support. In contrast, in the injured spinal cord implanted with scaffolds and prostheses, the contralateral spinal cord seemed normal in the lumbar region. Moreover, regeneration was more robust and organized. These findings demonstrated that cellular prostheses fabricated by MNs and scaffolds exhibited a better repairing effect than scaffolds alone or MNs alone after spinal cord hemisection, suggesting that MN-prostheses might be suitable and effective for repairing injured spinal cord tissue.
The functional recovery in all groups was estimated using evoked potentials and BBB scores. Evaluation of evoked potentials and BBB scores showed the limited functional recovery in Group A. The tracing experiments showed that spontaneous regeneration occurred in the lumbar region of rats in the blank control group. The functional recovery of rats in Group B (MNs group) was better than that in Group A, however, the improvement was limited and was possibly caused by MNs. Similarly, many studies have shown that cell implantation alone fails to generate functional recovery.11,18,22,23,51 Possibly, implanted cells are unable to receive essential extracellular cues due to insufficient intercellular contacts, and the nonhomogeneous distribution of implanted cells lacks three-dimensional scaffold support. 45 In contrast, three-dimensional scaffolds may provide anchorage sites for cell attachment and cues for tissue maturation that are essential to functional recovery.19,45 The functional recovery of the rats in Groups C (scaffold group), D (NSC prosthesis), and E (MN-prosthesis) was conspicuous, in addition to the supportive role of the scaffold as mentioned above, possibly due to the promotion of nerve regeneration by the sapeptide scaffold,28,29,35,36 in which can supply a compatible environment for the regeneration of nerve tissue by inhibiting glial scar formation and promoting axon elongation. Among Groups C–E, the prostheses showed better locomotion recovery, especially the MN-prosthesis, which suggested that cellular application in tissue-engineered spinal cord could promote the recovery of locomotion of rats. We further observed that more MNs (colabeled for NF-200 and VAChT) appeared in the transplanted region, surrounding the tissue near where the MN-prosthesis was implanted. Based on the fact that previous studies have shown that transplanted MNs could promote functional recovery in some spinal cord diseases,52,53 we presume that the application of MN-prostheses might provide benefit for the treatment of SCI. Possible mechanisms underlying better functional recovery after MN-prosthesis treatment than that by NSC prosthesis treatment in terms of promoting repair include the following: Embryonic stem cells involved in NSC prostheses might differentiate into the glial cells that could form a glial scar,22–24 which had been observed in this study and would occupy the pores in scaffolds and block axon endings, thereby obstructing axonal regeneration. Meanwhile, a glial scar could secrete a considerable amount of neurite outgrowth inhibitors that would inhibit axonal regeneration. In addition, the VAChT-positive neurofibra has been found in the graft of MN-prosthesis, which indicated that VAChT-positive neurofibra might be derived from MN-prosthesis or the host. Deshpande et al. 54 reported that the transplanted MNs could promote functional recovery of paralytic rats. However, they emphasized that the speed of functional recovery through MNs transplantation was slower than that through stem cells transplantation that supplied tropic support. The axonal elongation from the grafted MN progenitors is a long process which takes up to 6 months. The longest time in our study was 12 weeks, which might be insufficient to achieve specific replacement of impaired MNs. All of these suggested that MN-prosthesis might provide trophic support for host MNs. 20 However, our study in vitro showed that the level of trophic factors (BDNF) and mitogen (bFGF) in MNs was lower than that in NSCs. It was possible that some undetected factors, that would promote recovery of motion, were secreted by MNs in graft. Alilain et al. 55 reported that peripheral nerve graft was used as a bridge for promoting recovery of diaphragmatic function in C2-hemisected rat. In report, chondroitinase ABC was also important, and it could inhibit expression of chondroitin sulfate proteoglycans (CSPGs), CSPGs was a key component of glial scars and they powerfully inhibit neuronal plasticity. For the same reason, the tissue-engineered MNs prosthese also acted as a bridge for spread of axon, and we also found that there were no significant glial scars in group MNs prosthese, and there were some nerve fibers between two hemisected ends of spinal cord. On the contrary, there were some visible glial scars in NSCs prosthese graft, and few nerve fibers between two hemisected ends of the spinal cord. The possible mechanism will be explored in our further studies.
Conclusions
Neural repair after SCI could be accomplished by the application of multipotent spinal cord-derived cells associated with a synthetic, biocompatible, and biodegradable scaffolding material. This treatment could partly restore the lost function caused by SCI. Herein we demonstrated that the MN-prosthesis fabricated from MNs and a sapeptide scaffold can promote functional recovery after spinal cord hemisection. Our results suggest that the MN-prosthesis is a potential tool for developing tissue-engineered spinal cord to treat human SCI.
Footnotes
Acknowledgments
We wish to thank Jun Yan (Department of Physiology and Biophysics, University of Calgary, Calgary, Albert, Canada) for his assistance in preparing and amending the article. This work was partly supported by the National Natural Science Foundation of China (No. 30171068 and No. 30772299) and the Natural Science Foundation for Academician in Chongqing (No. 7671) and Natural Science Foundation Project of CQ CSTC (CSTC, 2007BB5083).
Disclosure Statement
No competing financial interests exist.