
Abstract
Enhancement of in vivo mobilization and homing of endogenous mesenchymal stem cells (MSCs) to an injury site is an innovative strategy for improvement of bone tissue engineering and repair. The present study was designed to determine whether mobilization by AMD3100 and/or local homing by delivery of stromal cell-derived factor-1 (SDF-1) enhances recombinant human bone morphogenetic protein-2 (rhBMP-2) induced ectopic bone formation in an established rat model. Rats received an injection of either saline or AMD3100 treatment 1 h before harvesting of bone marrow for in vitro colony-forming unit-fibroblasts (CFU-F) culture or the in vivo subcutaneous implantation of absorbable collagen sponges (ACSs) loaded with saline, recombinant human bone morphogenetic protein-2 (rhBMP-2), SDF-1, or the combination of SDF-1 and rhBMP-2. AMD3100 treatment resulted in a significant decrease in CFU-F number, compared with saline, which confirmed that a single systemic AMD3100 treatment rapidly mobilized MSCs from the bone marrow. At 28 and 56 days, bone formation in the explanted ACS was assessed by microcomputed tomography (μCT) and histology. At 28 days, AMD3100 and/or SDF-1 had no statistically significant effect on bone volume (BV) or bone mineral content (BMC), but histology revealed more active bone formation with treatment of AMD3100, loading of SDF-1, or the combination of both AMD3100 and SDF-1, compared with saline-treated rhBMP-2 loaded ACS. At 56 days, the addition of AMD3100 treatment, loading of SDF-1, or the combination of both resulted in a statistically significant stimulatory effect on BV and BMC, compared with the saline-treated rhBMP-2 loaded ACS. Histology of the 56-day ACS were consistent with the μCT analysis, exhibiting more mature and mineralized bone formation with AMD3100 treatment, SDF-1 loading, or the combination of both, compared with the saline-treated rhBMP-2 loaded ACS. The present study is the first that provides evidence of the efficacy of AMD3100 and SDF-1 treatment to stimulate trafficking of MSCs to an ectopic implant site, in order to ultimately enhance rhBMP-2 induced long-term bone formation.
Introduction
Trafficking of MSCs to injury sites is initiated by mobilization, which is defined as forced egress of MSCs from the physiological stem cell niche of the bone marrow, the richest and most reliable reservoirs of MSCs, to the circulating peripheral blood.6,7 There is previous evidence that MSCs circulate in peripheral blood, although at extremely low incidence under normal physiological conditions.6,8 A bioactive agent known to affect mobilization and homing of MSCs is AMD3100, a novel bicyclam and a highly specific antagonist to CXCR4, which is expressed by stem and progenitor cells that reside in the bone marrow stem cell niche. 9 Specifically, CXCR4 is highly expressed in hematopoietic stem cells (HSCs)/hematopoietic progenitor cells (HPCs), but it is also expressed in MSCs, albeit at lower levels and primarily intracellularly.2,3,10–12 AMD3100 (also known as Plerixafor) was recently FDA-approved (Mozobil®) as an HSC mobilizer for autologous stem cell transplantation in patients with non-Hodgkin's lymphoma and multiple myeloma. 13 AMD3100 has previously been used by itself to rapidly mobilize HSCs/HPCs,14–20 endothelial progenitor cells (EPCs),18,21 and MSCs, 18 from bone marrow into peripheral blood. It has been reported that peak mobilization of HSCs/HPCs14,19 and EPCs 22 was achieved 1 h postinjection of a single treatment of AMD3100. Given this knowledge, it was expected that MSCs would have similar in vivo mobilization kinetics induced by AMD3100/CXCR4 antagonism as those previously reported for HPCs and EPCs.
After mobilization, homing is known to be the other major coordinated and multistep process involved in the trafficking and recruitment of MSCs to injury sites. 23 Homing can be defined as the arrest of MSCs within the vasculature of a tissue followed by transmigration across the endothelium. 7 Homing of circulating marrow-derived MSCs and osteoprogenitor cells to ectopic sites of induced bone formation is known to involve homing receptor CXCR4, a chemotactic receptor for all isoforms of chemokine ligand stromal cell-derived factor-1 (SDF-1, CXCL12).24,25 SDF-1 is well known to be important for the regulation of migration, survival, and development of multiple cell types, and it is continuously produced by bone marrow stromal cells, immature osteoblasts, and endothelial cells, as well as expressed from various tissues homeostatically.26–28 The importance of the SDF-1/CXCR4 axis is obvious as knock outs are lethal, due to bone marrow, heart, and brain failure. 29 It is known that SDF-1 is a potent chemoattractant expressed in the bone marrow stem cell niche under normal physiological conditions for the retention of HSCs, and in upregulated injured/hypoxic/ischemic tissues, which, in turn, induces CXCR4-expressing cells, including MSCs, to home to high gradients of SDF-1.30,31 A previous study using heterozygous SDF-1 and CXCR4 mice reported that the SDF-1/CXCR4 axis is of critical importance for recruitment of MSCs to a segmental defect site during early phases of endochondral bone fracture healing. 32 Additionally, it has been shown that the local controlled release of SDF-1 from subcutaneously implanted gelatin hydrogels enhances angiogenesis compared with the injection of SDF-1, 33 and continuous delivery of SDF-1 from alginate patches significantly accelerated wound healing and reduced scarring compared with saline controls. 34
In this study, we test the overall hypothesis that systemic pharmacologic mobilization and/or local homing of endogenous MSCs enhances bone formation in an established rat model of recombinant human bone morphogenetic protein-2 (rhBMP-2) induced ectopic bone formation. To the best of our knowledge, the combination of mobilization and homing of MSCs originating from endogenous reserves has previously not been studied in an ectopic bone formation model. Specifically, we sought to determine whether rhBMP-2 induced bone formation in an ectopic model can be enhanced by mobilization of MSCs by AMD3100 treatment or by local delivery of SDF-1 at the ectopic implant site. The primary endpoint at 28 and 56 days was bone formation at the ectopic implant site, as assessed by microcomputed tomography (μCT) and histology.
Materials and Methods
AMD3100
AMD3100 (AMD3100 Octahydrochloride Hydrate) was purchased from Sigma-Aldrich and was reconstituted using sterile water at a concentration of 4 mg/mL (2 mg per 500 μL injection).
In vitro bone marrow-derived colony-forming unit-fibroblasts assay
Eight male Sprague-Dawley rats (∼350 g; 12 weeks old) each received a single intraperitoneal (i.p.) injection of saline (N=3) or AMD3100 (N=5; 2 mg) 1 h before harvesting of femoral bone marrow. The distal and proximal ends of the femur were cut off, and the marrow was flushed out with sterile culture media. A single cell suspension was prepared by mechanical disruption and filtered using a 70 μm cell strainer. 35 The total number of viable nucleated cells was determined by trypan blue stain (Sigma). About 5×105 nucleated viable cells in the marrow suspension were plated in 25 cm2 polystyrene cell culture flasks (BD Falcon), and cultured in α-MEM (Gibco) with 10% fetal bovine serum (GemCell) and penicillin (5000 units/100 mL)/streptomycin sulphate (5 mg/100 mL; Gibco). Cultures were incubated at 37°C in 20% O2 +5% CO2 (normoxia) or 5% O2+10% CO2 (hypoxia) for 14 days, with the media changed every 3 days. Culturing primary MSCs in hypoxia significantly amplifies the number of colony-forming unit-fibroblasts (CFU-F),36–39 thereby resulting in more prominent CFU-F for identification and quantitation. On day 14, the media was removed, and the adherent layer of CFU-F was washed in phosphate-buffered saline (PBS; Sigma). Cells were fixed in 10% formalin (Sigma), stained with 1% Toluidine Blue O (Sigma), and imaged with an Epson Expression 1600 scanner at standard 24 bit color, 720 dpi. Photoshop version 7.0.1 was used to convert the image to grayscale, invert, select the cell culture area from the background of the culture flask, and save as an individual TIFF. CFU-F numbers were manually counted. Image analysis software (MetaMorph; Molecular Devices), set to a grayscale threshold (40–254), was used to measure % area coverage (which represents the area that the colonies cover in each culture flask and was calculated as the ratio of white pixels to the total number of pixels) and integrated intensity (which represents relative signal density of the colony and was calculated as the average intensity of pixels).
Growth factor loading onto absorbable collagen sponges
Helistat® hemostatic absorbable collagen sponges (ACSs) were purchased from Integra Lifesciences Corp. ACSs were cut with a scalpel into pieces with dimensions 12.5 mm length×6.5 mm width×7 mm thickness. ACSs were loaded with solutions of saline, rhBMP-2 (Medtronic; 10 μg in sterile 5 mM glutamate, 5 mM NaCl, 2.5% glycine, 0.5% sucrose, 0.01% Tween-80 at a pH 4.5), SDF-1 (ProSpec–Tany TechnoGene Ltd.; recombinant rat SDF-1α; 200 ng in sterile saline), or both rhBMP-2 and SDF-1, by soaking the ACS pieces with 100 μL of solution, and storing the soaked ACS in six-well plates at 4°C overnight.
AMD3100 treatments for in vivo experiments
12 adult male rats (Sprague Dawley, ∼350 g) each received a single i.p. injection of saline (N=6) or AMD3100 (N=6; 2 mg; Sigma-Aldrich) 1 h before surgical implantation, for the 28-day time point. 6 adult male rats each received i.p. injections of AMD3100 (N=6; 2 mg; Sigma-Aldrich) 1 h before surgical implantation and also three additional injections weekly, until sacrifice at the 28-day time point. Twelve adult male rats each received a single i.p. injection of saline (N=6) or AMD3100 (N=6; 2 mg; Sigma-Aldrich) 1 h before surgical implantation, for the 56-day time point.
Rat surgery model for in vivo ectopic bone formation assay
In an Institutional Animal Care and Use Committee of the Rush University Medical Center (IACUC; Protocol #10–034) approved study, each of the 30 adult male rats (Sprague Dawley, ∼350 g) received the same surgical procedure of dorsal subcutaneous implantation of four ACSs, using aseptic technique and adapting the method previously described. 40 The implantation was performed under anesthesia using ketamine (100 mg/kg, intramuscular) and xylazine (5 mg/kg, intramuscular), and supplemented as necessary by inhalation anesthesia using isoflurane. The animals were placed in a prone position, and the dorsal surface was shaved. A template was placed over the back, and an alcohol-resistant surgical pen was used to mark the four sites of ACS placement to ensure consistent implant placement. The dorsal surface was scrubbed with alcohol, betadine, and two 1.5-cm-long incisions (1 cm from the midline) were made equidistant to the four marked template dots to make the subcutaneous space for ACS. Each animal received four ACS, which were loaded with saline, rhBMP-2 (10 μg), SDF-1 (200 ng), or both rhBMP-2 and SDF-1. An absorbable 3-0 Plain Gut transdermal suture (Ethicon, Inc.) was placed through the subdermal ectopic ACS as an anchor to accurately maintain its location. A non-absorbable 5-0 Monocryl suture (Ethicon, Inc.) using an interrupted stitch was used to approximate the soft tissues at the incision site, and the rat was placed on a heating pad for recovery. A subcutaneous injection of buprenex (0.02 mg/kg delivered with saline) was given once for postoperative pain management.
For the 28-day and 56-day time point groups, the rats were euthanized using carbon dioxide (CO2) inhalation. A large incision and dorsal subcutaneous skin flap was made to allow for in situ implant visualization and harvesting. ACS implant capsules were carefully trimmed from the surrounding soft tissue, fixed in 10% neutral buffered formalin for 2 days at 4°C, and stored in PBS (pH 7.4) at 4°C.
Microcomputed tomography
To calculate the total bone (mineralized tissue) volume in each ACS implant sample, μCT scanning was used. The fixed specimens were securely held in a specimen holder, which was filled with PBS (pH 7.4). All samples were examined by μCT using 70 kV energy (X-ray voltage), and yielding 15-μm isotropic voxel in-plane resolution, with 15-μm thick slices. The manufacturer's software was used to filter noise (sigma=0.8, support=1.0) and segment the data at a global threshold of 200. The bone volume (BV, mm3) and bone mineral density (mg/cm2 of hydroxyapatite [HA], based on a standard calibration phantom provided by the manufacturer) were determined. The total bone mineral content (BMC, mg of HA) was calculated by multiplying BV with mineral density.
Histology
Samples (N=4) were decalcified in 0.5 M ethylenediaminetetraacetic acid for 1 week, embedded in paraffin, sectioned (6 μm), and stained with Toluidine Blue O.
Statistical analyses
All data are reported as mean±standard error of the mean. For all tests, statistical significance was defined by statistical probability of p≤0.05.
A student's t-test was used to compare characterization of in vitro bone marrow-derived CFU-F assay after saline or AMD3100 treatments, and to assess statistical significance.
For in vivo ectopic bone formation assays, at 4 and 8 weeks for rhBMP-2 loaded ACS implants, we used student's or paired t-tests to assess statistical significance of the comparisons of the control implants (SDF-1 negative in animals injected with saline) to SDF-1 negative implants in animals treated with AMD3100 (student's t-tests), SDF-1 positive implants in rats treated with saline (paired t-tests), and SDF-1 positive implants in rats treated with AMD3100 (student's t-tests). For the student's t-tests, Levene's test of equality of the variances was examined, and the relevant levels were used.
Results
Single treatment of AMD3100 mobilizes MSCs from the bone marrow
Fixing, staining, and scanning of CFU-F resulted in images of total CFU-F in each 25 cm2 cell culture flask (Fig. 1). AMD3100 treatment resulted in a statistically significant decrease in CFU-F number,% area coverage, and integrated intensity compared with saline under hypoxic conditions (Fig. 2). The CFU-F assay confirmed that a single systemic AMD3100 treatment rapidly mobilized MSCs from the bone marrow.
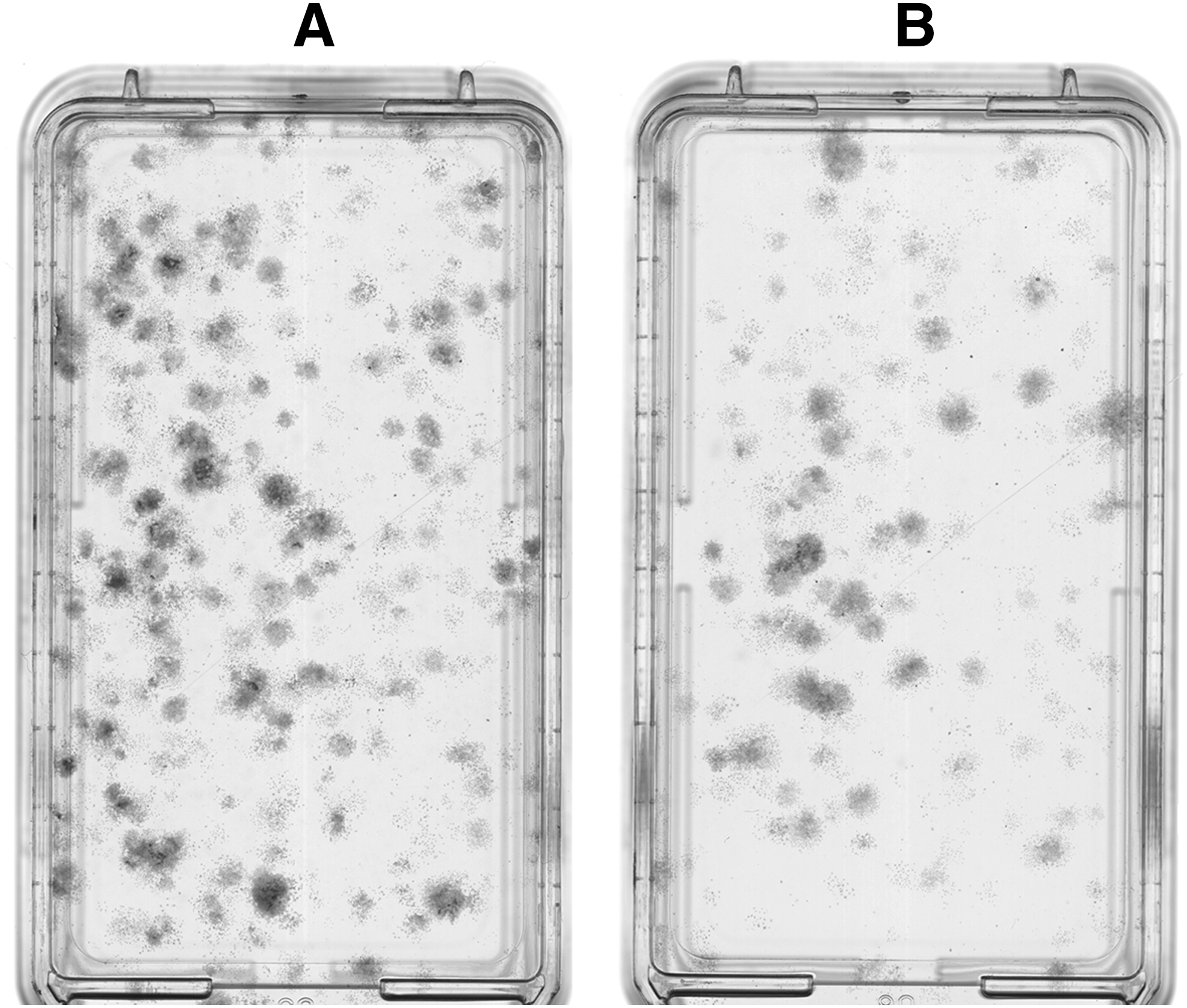
Colony-forming unit-fibroblasts (CFU-F) originating from rat femoral bone marrow-derived mesenchymal stem cells at 14 days culture in vitro on representative plastic culture flasks. Cells were obtained from rats treated with injections of saline
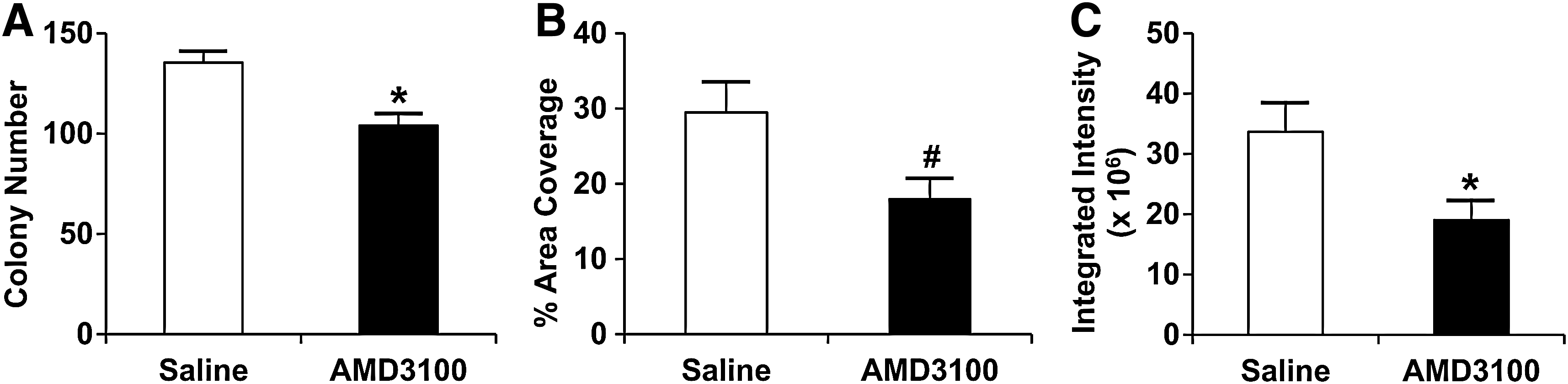
CFU-F characterization at 14 days.
Single AMD3100 treatment and/or SDF-1 loading does not enhance BV or BMC in rhBMP-2 loaded ACS at 28 days
BV and BMC were determined by μCT for ACS implanted for 28 days. There was no statistical difference between BV and BMC when comparing saline or AMD3100-treated ACS loaded with rhBMP-2 or the combination of rhBMP-2 and SDF-1 (Fig. 3). Considering the means of the data, the addition of a single AMD3100 treatment and/or SDF-1 loading resulted in a slight decrease in BV or BMC for rhBMP-2 loaded ACS at 28 days (Figs. 3 and 4). Furthermore, continuous (weekly) AMD3100 treatment for 28 days resulted in a statistically significant decrease in BV (p=0.006) and BMC (p=0.003) for ACS loaded with rhBMP-2 (BV=2.2 mm3; BMC=1.4 mg HA) or the combination of rhBMP-2 and SDF-1 (BV=2.0 mm3; BMC=2.3 mg HA), compared with the saline-treated rhBMP-2 loaded ACS at 28 days. There was a lack of evidence of bone tissue formation in ACS loaded with only saline or SDF-1, as determined by μCT.

Bone volume (BV) and bone mineral content (BMC) for 28-day samples. BV
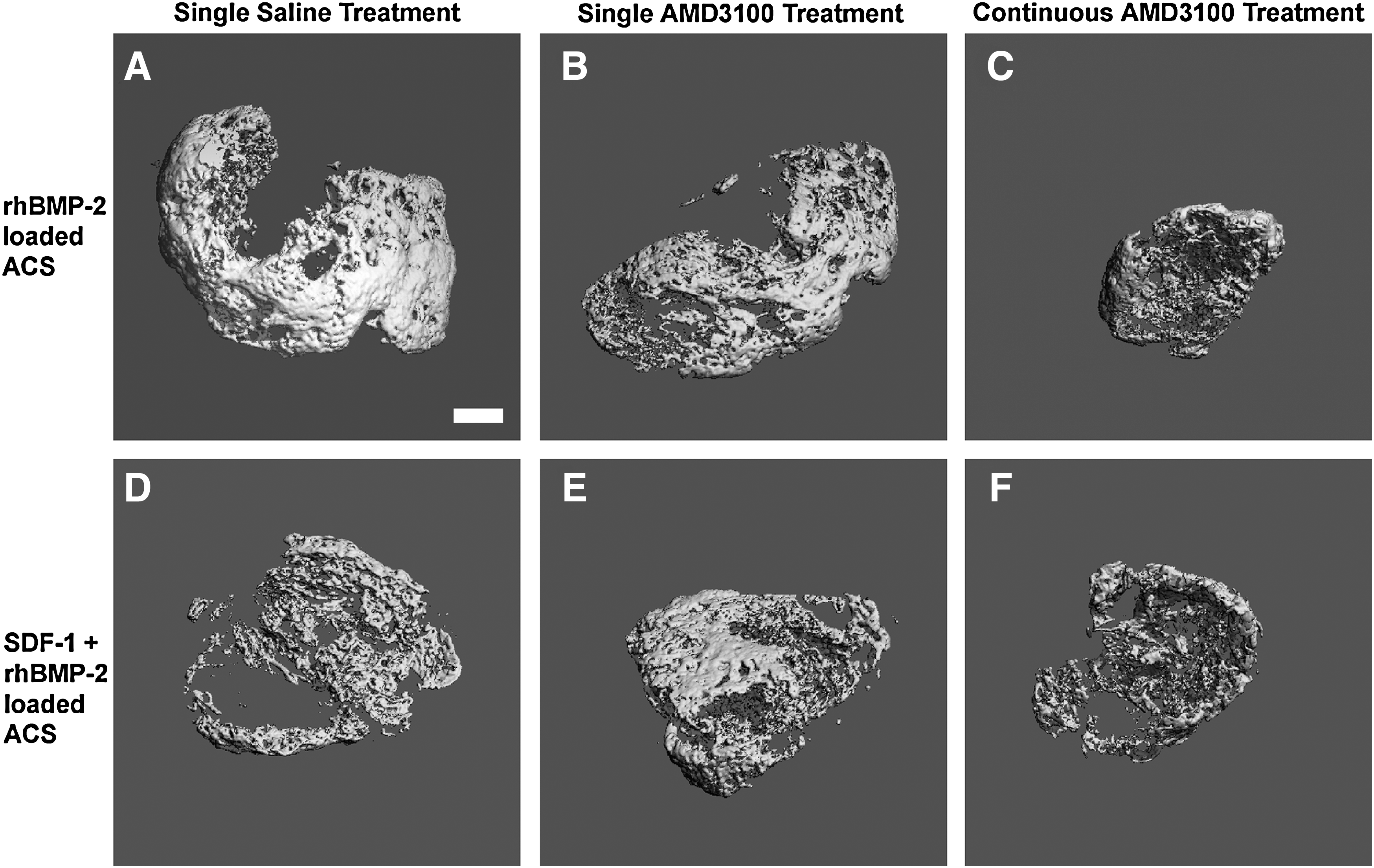
Reconstructed three-dimensional (3D) microcomputed tomography (μCT) images of representative 28-day samples. rhBMP-2-loaded ACS implants at 28 days, which received a single saline treatment
Single AMD3100 treatment and/or SDF-1 loading leads to more active bone formation in rhBMP-2 loaded ACS at 28 days
Histological staining with Toluidine Blue O of 28 day ACS samples revealed markedly more active endochondral bone/early trabecular bone formation (as evident by aligned cuboidal osteoblasts, osteoid, and areas of cartilaginous remnants) in the presence of a single treatment of AMD3100 (Fig. 5B), or SDF-1 loading (Fig. 5D), or the combination of both AMD3100 and SDF-1 (Fig. 5E), compared with more mature/mineralized bone with embedded osteocytes and fatty marrow/adipose-like tissue identified in samples treated with only saline (Fig. 5A). Toludine Blue O staining of ACS that received continuous (weekly) treatment of AMD3100 (Fig. 5C, F) for 28 days exhibited areas of mostly cartilaginous tissue, easily identifiable residual collagen sponge (fibers), and noticeable amounts of engrafted inflammatory-like cells, especially with the addition of SDF-1 loading (Fig. 5F). As supported by the histology, continuous (weekly) AMD3100 treatment for 28 days resulted in distinct impairment of active bone formation in the presence of rhBMP-2 (Fig. 5C) or the combination of SDF-1 and rhBMP-2 (Fig. 5F).
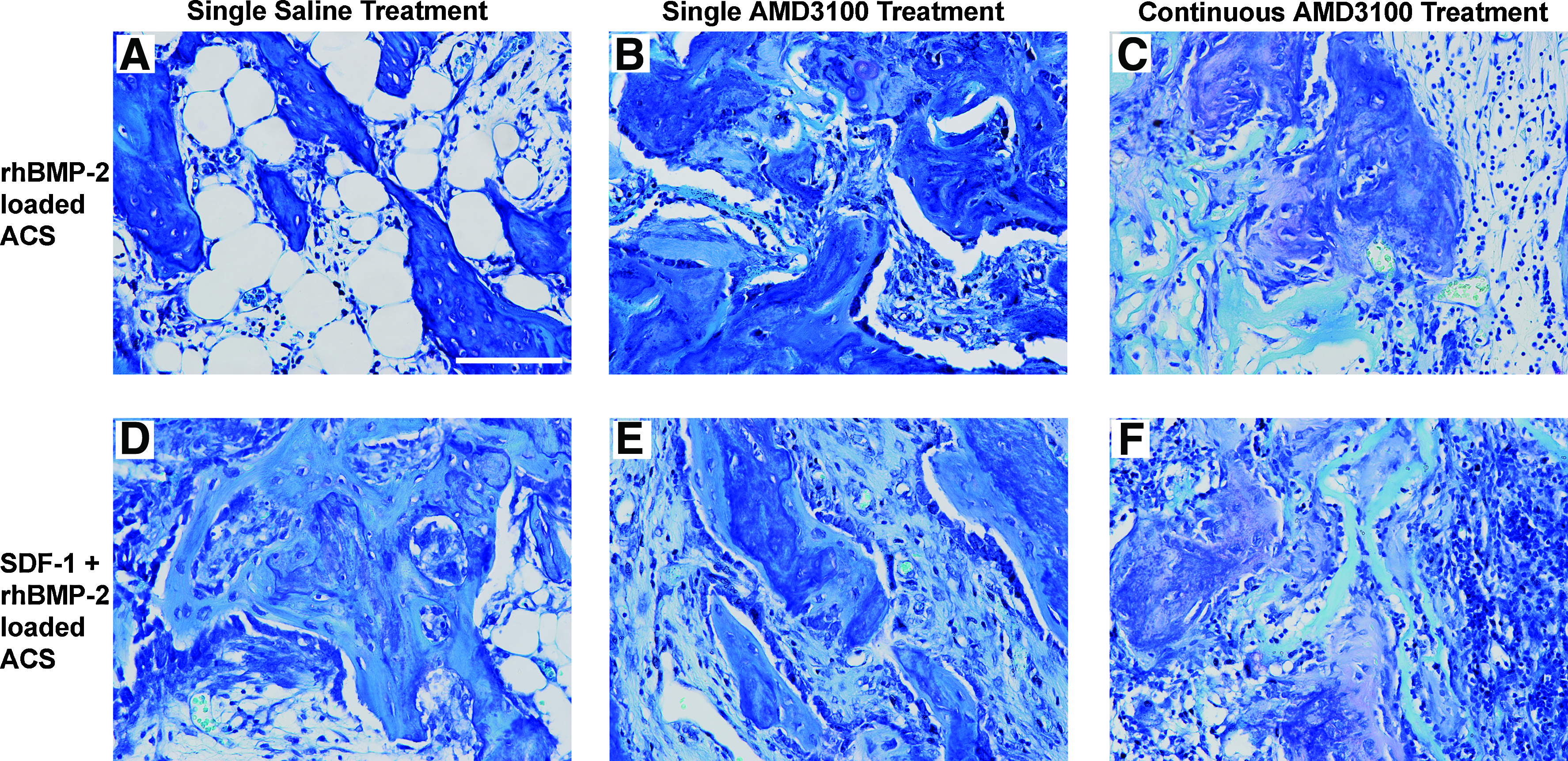
Histology of 28-day samples. Representative views of rhBMP-2-loaded ACS implants at 28 days, which received a single saline treatment
Single AMD3100 treatment and/or SDF-1 loading significantly enhances BV and BMC in rhBMP-2 loaded ACS at 56 days
Compared with saline-treated rhBMP-2 loaded ACS at 56 days, the addition of loading SDF-1 in combination with rhBMP-2 resulted in a statistically significant stimulatory effect on BV and BMC (Figs. 6 and 7). Likewise, the addition of a single injection of AMD3100, with or without the presence of SDF-1 in the rhBMP-2 loaded ACS, also had a stimulatory effect on BV and BMC, which were statistically significant compared with the saline-treated rhBMP-2 loaded ACS at 56 days (Figs. 6 and 7). Therefore, at 56 days, a single treatment of AMD3100 and/or SDF-1 loading of ACS significantly enhanced mineralized bone formation, but the combination of AMD3100 and SDF-1 was not statistically different than using either AMD3100 or SDF-1 alone. There was a lack of evidence of bone tissue formation in ACS loaded with only saline or SDF-1, as determined by μCT.

BV and BMC for 56-day samples. BV
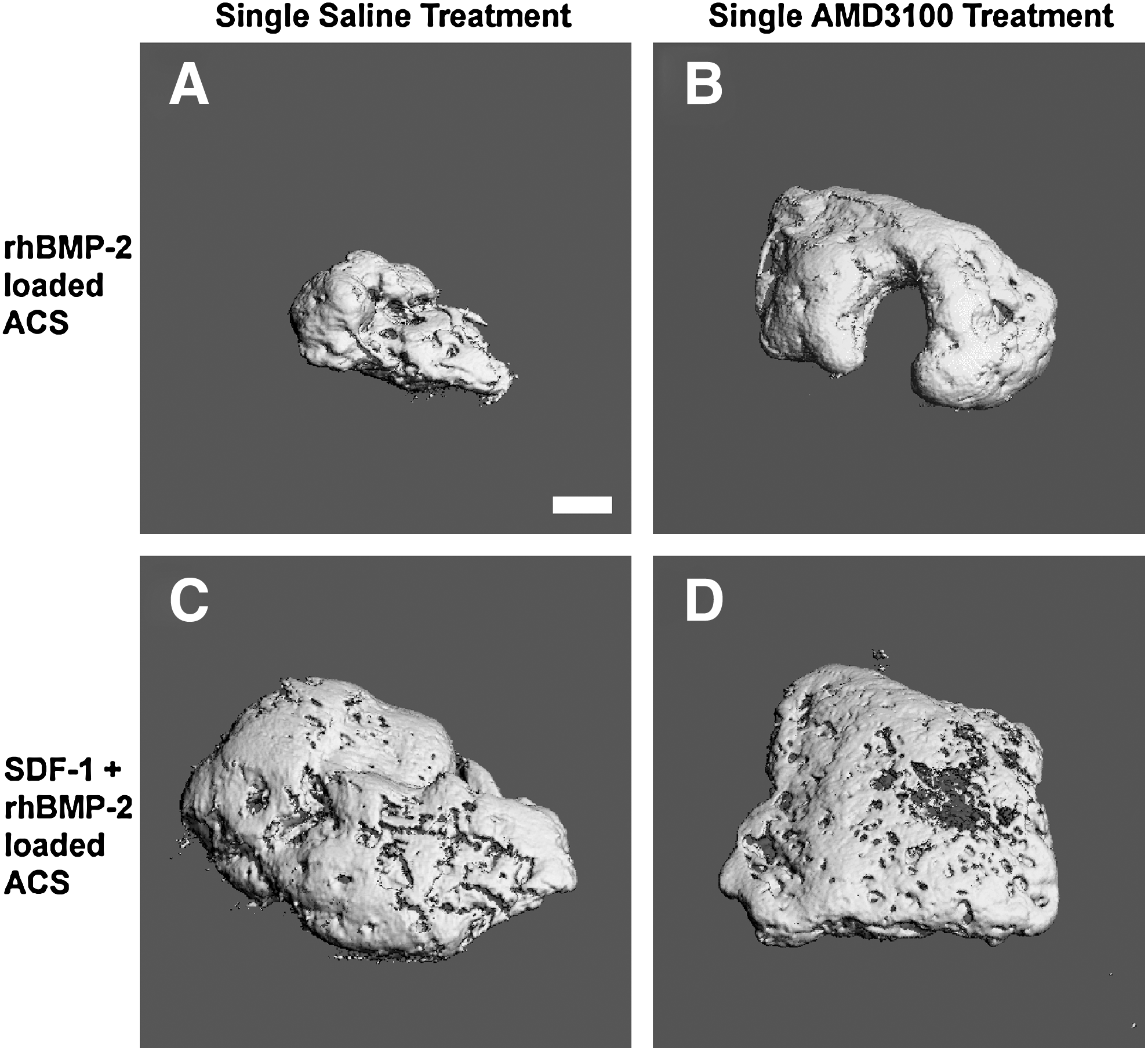
Reconstructed 3D μCT images of representative 56-day samples. rhBMP-2-loaded ACS implants at 56 days, which received a single saline treatment
Single AMD3100 treatment and/or SDF-1 loading leads to more mature/mineralized bone formation in rhBMP-2 loaded ACS at 56 days
Histological staining with Toludine Blue O and imaging of representative views of rhBMP-2 loaded ACS samples at 56 days displayed that the addition of AMD3100 treatment (Fig. 8B), or SDF-1 loading (Fig. 8C), or the combination of AMD3100 and SDF-1 (Fig. 8D), results in the formation of more mature and mineralized bone embedded by osteocytes, compared with ACS loaded with only rhBMP-2 and treated with saline (Fig. 8A). Histology of the saline-treated rhBMP-2 loaded ACS at 56 days primarily exhibited fatty marrow/adipose-like tissue within the majority of the whole sample area, surrounded by mature bone with a lamellar appearance predominantly at the periphery, along with small islands of mature bone (Fig. 8A). These findings for the saline-treated rhBMP-2 loaded ACS at 56 days (BV=4.6 mm3; BMC=3.3 mg HA) are indicative of resorption of the mature bone, which is known to have previously existed more abundantly at 28 days (BV=7.6 mm3; BMC=5.0 mg HA) (Fig. 5A).
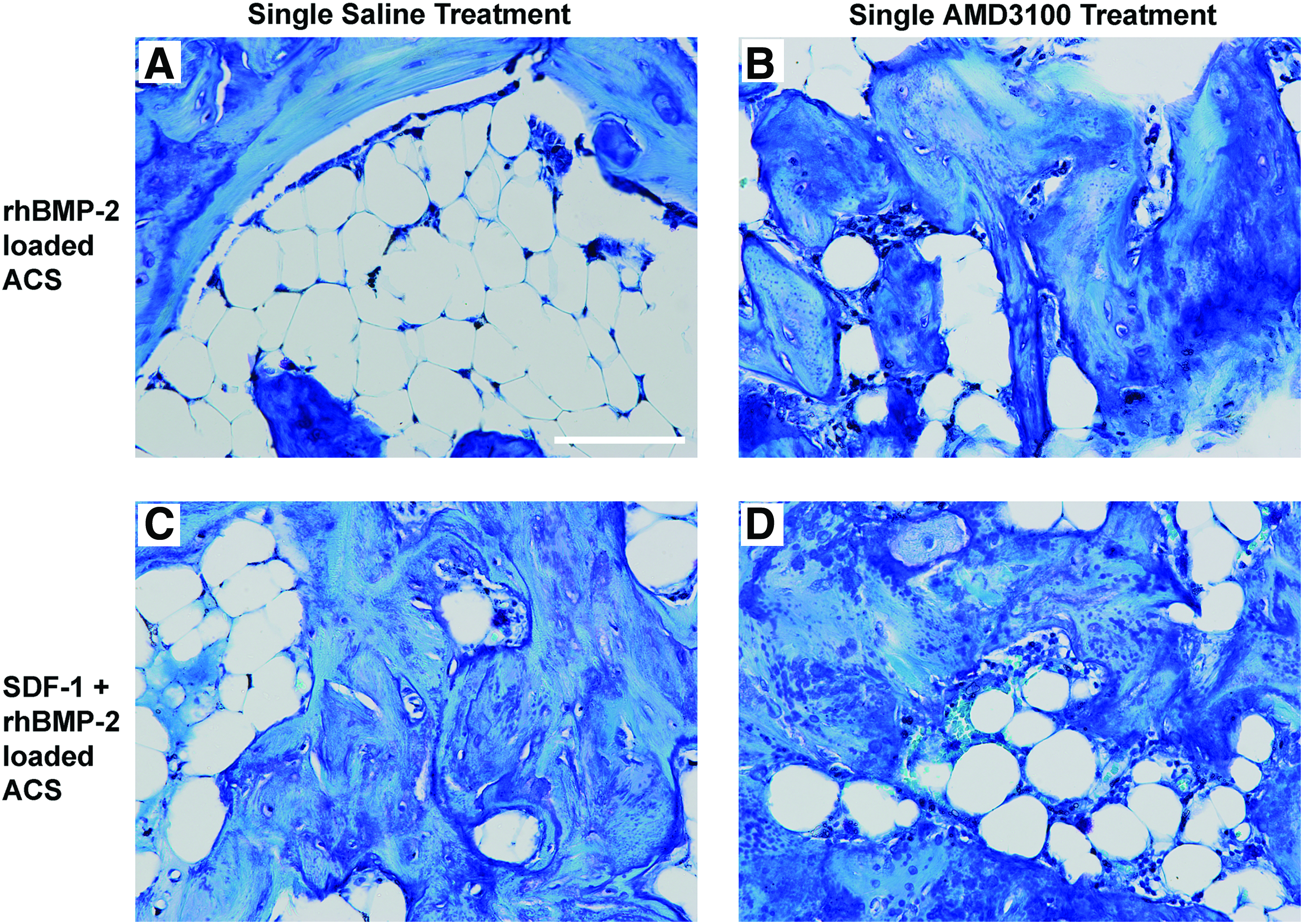
Histology of 56-day samples. Representative views of rhBMP-2-loaded ACS implants at 56 days, which received a single saline treatment
Discussion
The objective of this work was to investigate whether deliberate mobilization and/or homing of MSCs would enhance rhBMP-2 induced ectopic bone formation. Mobilization was induced by single systemic injection treatment of AMD3100, and homing was induced by local delivery of SDF-1 loaded on the ACS, along with local delivery of rhBMP-2 to induce ectopic bone. We found that the amount of bone formation was significantly greater at a later time point. Histology showed evidence of more active endochondral bone formation in animals receiving the CXCR4 antagonist, AMD3100. At 56 days, we found a statistically significant enhancement of rhBMP-2 induced bone formation (BV and BMC) with all three types of treatments: AMD3100 only, SDF-1 only, and the combination of AMD3100 and SDF-1. Histology of the 56-day samples confirmed the μCT results, while also contrasting the histology of the 28-day samples, as greater amounts of mature/mineralized bone were now observed for the AMD3100, SDF-1, or combination treatments compared with the saline-treated rhBMP-2 loaded samples which exhibited lesser amounts of mature bone globally and primarily fatty marrow that occupied the core of the implant. This result of bone resorption observed in the 56-day saline-treated rhBMP-2 loaded samples strongly agrees with other reports of bone resorption in rhBMP-2 (10 μg) loaded ACS implanted subcutaneously for 8 weeks in an ectopic bone formation model in rats.41,42
Additionally, we found that 28-day bone formation was hindered with continuous (weekly) treatments of AMD3100 in the presence or absence of SDF-1, as supported by further decreased BV and BMC, and notably less bone formation and more cartilaginous tissue and ACS remnants were observed by histology. Continuous (weekly) treatment of AMD3100 likely caused a more frequent antagonizing of CXCR4-expressing cells and systemic mobilization of MSCs, thereby disrupting or blocking MSCs engraftment at the ectopic site and, therefore, negatively affecting subsequent opportunities of chondrogenic and osteogenic differentiation at the local site. These findings are consistent with results from another recent study which found that biweekly injections of AMD3100 resulted in a notably disrupted and non-adherent MSC stroma at 28 days in ectopic MSC-coated scaffolds. 43
The subcutaneous ectopic bone formation model in the rat, using rhBMP-2 loaded ACS, has several advantages compared with other models of bone regeneration, but also some drawbacks to consider. For this study, the ectopic bone formation model was not only convenient for surgery and necropsy purposes, but also allowed for utilization of uniquely isolated in vivo ectopic implant sites to study the effects on bone formation of a drug and growth factor known to modulate the mobilization and/or homing of stem or progenitor cells. On the other hand, results from this ectopic bone formation model cannot be directly translated to other models that represent a true bone injury or repair site, such as with fracture repair, critical-sized segmental defects, spinal fusion, or craniofacial defect models. 44 Importantly, though, the ectopic bone formation model does not contain any pre-existing local resident stem cells, vasculature, or bone, and begins with only ACS loaded with rhBMP-2. Therefore, the results of significant enhancement of rhBMP-2 induced bone formation at 56-days by AMD3100 and/or SDF-1 can be considered a direct effect of these MSC mobilizing and/or homing treatments. Considering the requirement of neovascularization to first occur before initiation of rhBMP-2 induced endochondral bone formation, we can expect that neovascularization may have also been enhanced by AMD3100 and/or SDF-1 treatment.
In our study, the concentrations used of 5 mg/kg of AMD3100, 18 200 ng of SDF-1 per ACS implant,45,46 and 10 μg of BMP-247–49 were based on previous publications from other groups. Additionally, one publication described the release kinetics of SDF-1 from collagen scaffolds, reporting that there is a rapid release of 50% of the SDF-1 in the first 24 h, and, therefore, a high gradient, followed by a notably slower release of an additional 10% for at least a week thereafter. 45 Release kinetics of rhBMP-2 from ACS have also been previously described.50,51
In addition to mobilization and homing of endogenous MSCs and osteoprogenitor cells expressing CXCR4,24,25 it is expected that other non-bone forming cells, particularly EPCs and HSCs/HPCs, are mobilized from the bone marrow into circulating peripheral blood by AMD3100, which has been previously demonstrated by Pitchford et al., 18 and/or recruited by SDF-1 to the rhBMP-2 induced ectopic bone formation site. Mobilized and homed EPCs likely contributed to neovascularization in the ectopic bone formation model.21,52 It was recently reported that endothelial-mesenchymal transition of vascular endothelial cells results in the generation of mesenchymal stem-like cells which contribute robustly in stages of heterotopic ossification.53,54 Although this is an interesting new discovery to consider, we have shown in our study that AMD3100 can rapidly release MSCs from bone marrow, as supported by our in vitro CFU-F assay results, and that these mobilized MSCs are strongly expected to have contributed to newly formed ectopic bone. HSCs/HPCs are no doubt being mobilized into circulating peripheral blood by AMD3100 and recruited to the ectopic site by SDF-1 within the first few days, initiating the possible generation of a hematopoietic microenvironment in which HPCs will reside, proliferate, and differentiate along different lineages resulted in the presence of several cell types in the ACS after a month of implantation. 45 Osteoclastogenesis could then be occurring in the ectopically implanted ACS site due to differentiation of HPCs (monocyte-macrophage lineage) to osteoclasts, particularly in the presence of rhBMP-2 and rhBMP-2 induced osteoblastic differentiation of recruited MSCs. 55
Reports by other groups have previously shown evidence that SDF-1 plays a crucial regulatory role in BMP-2 induced osteogenic differentiation of MSCs. Hosogane et al. showed that BMP-2-induced alkaline phosphatase (ALP) activity, osteocalcin (OC) synthesis, and calcium deposition was significantly decreased by blocking the SDF-1/CXCR4 signal axis with AMD3100. 56 More so, they reported that with the addition of exogenous SDF-1 protein to BMP-2 induced osteogenic differentiation of MSCs, ALP, and OC was significantly enhanced, and the calcium deposition was restored to the BMP-2-induced level. 56 Another group found that overexpression of SDF-1 in transduced human bone marrow-derived MSCs induced a significantly greater level of ectopic bone formation in mice by 8 weeks. 57 These reports, regarding regulation of BMP-2-osteoinduction by SDF-1, correlate well with our findings of significantly enhanced ectopic formation of mature/mineralized bone at 56 days in ACS initially soaked with SDF-1 alone, or even in combination with a single initial AMD3100 treatment.
A recent paper by Higashino et al. uses a similar model to ours and has also shown some interesting findings involving stem cell homing using SDF-1 in an in vivo ectopic bone formation rat model. 58 In their study, in vivo implants of collagen pellets containing a dose of 2.5 μg of rhBMP-2 exhibited maximally enhanced ectopic bone at 28 days with the addition of 5 μg of SDF-1, which is an amount 25 times greater than the amount of SDF-1 that we loaded on ACS in our study. It was shown that there was a dose-dependent effect on rhBMP-2-induced ectopic bone formation of adding 0, 0.5, 1, or 5 μg of SDF-1 to a deliberately suboptimal dose of rhBMP-2. Therefore, a dose higher than 200 ng of SDF-1 that was used in our study would have likely further enhanced the formation of bone at the 28 day time point.
An emerging concept is that mechanisms of MSC homing to damaged tissue may be similar to HSC/leukocyte homing, but through a modified set of adhesion molecules with certain differences.7,59,60 Known differences between MSC homing and leukocyte/HSC homing are that the latter employ L- and E-selectin in the initial rolling stage 61 and use platelet/endothelial cell adhesion molecule 1/CD34 in leukocyte transmigration across the endothelium, 62 whereas expression of these molecules is low or absent on the surface of MSCs. Interestingly, it was found that MSCs utilize some comparable mechanisms to leukocytes regarding their first stage of recruitment to areas in need of repair. Specifically, it has been shown that MSCs roll on endothelial cells in a P-selectin manner and that rolling MSCs interact with the very late antigen-4 (VLA-4)/vascular cell adhesion molecule-1 (VCAM-1) axis, which is responsible for mediating firm adhesion of human MSCs to endothelial cells. 61 Furthermore, it was reported that MSCs can transmigrate through non-activated endothelial monolayers via VCAM-1/VLA-4 interactions, in addition to MMP-2 secretion and by other interactions with various integrins and cytokines, and possibly integrate with the endothelial layer as embedded pericytes. 63 Future studies are required to discover more similarities and differences of MSC and HSC homing mechanisms.
In conclusion, this study provides evidence of the efficacy of AMD3100 and SDF-1 treatment to stimulate trafficking of MSCs to an ectopic implant site, in order to ultimately enhance rhBMP-2 induced long-term bone formation. Future studies could utilize AMD3100-induced mobilization and SDF-1-induced homing of MSCs to enhance and accelerate bone repair in more clinically relevant and challenging bone defect surgical models.
Footnotes
Acknowledgments
Funding was provided by NIH Institutional Training Grant (T32 AR052272), the Grainger Foundation, and the Musculoskeletal Transplant Foundation. Julie Brown provided assistance with surgical preparation and necropsy. The MicroCT/Histology Core provided access. David Karwo provided assistance with histology. Dana Glock provided assistance with μCT.
Disclosure Statement
No competing financial interests exist.