
Abstract
The selection of a suitable scaffold material is important for dentin tissue regeneration, as the characteristics of biomaterials can potentially influence cell proliferation and differentiation. We compared the effects of different scaffolds on dentin regeneration based on dental pulp stem cells (DPSCs) and investigated the regulatory mechanisms of odontogenic differentiation of DPSCs by these scaffolds. Five different scaffolds were tested: demineralized dentin matrix (DDM), ceramic bovine bone (CBB), small intestinal submucosa (SIS), poly-L-lactate-co-glycolate, and collagen–chondroitin sulfate–hyaluronic acid. DPSCs cultured on DDM and CBB exhibited higher levels of alkaline phosphatase (ALP) activity and mRNA expression of bone sialoprotein, osteocalcin, dentin sialophosphoprotein (DSPP), and dentin matrix protein-1 (DMP-1) than those cultured on the other three scaffolds. Further, the phosphorylation levels of mitogen-activated protein kinase (MAPK) ERK1/2 and p38 in DPSCs cultured on DDM and CBB were also significantly enhanced compared with the other three scaffolds, and their inhibitors significantly inhibited odontogenic differentiation as assessed by ALP activity and mRNA expression of DSPP and DMP-1. The implantation experiment confirmed these results and showed a large amount of regular-shaped dentin-pulp complex tissues, including dentin, predentin, and odontoblasts only in the DDM and CBB groups. The results indicated that natural mineralized scaffolds (DDM and CBB) have potential as attractive scaffolds for dentin tissue-engineering-promoted odontogenic differentiation of DPSCs through the MAPK signaling pathway.
Introduction
Equally as important as the scaffold in the classic tissue-engineering paradigm is the cell source. Dental pulp stem cells (DPSCs) have the ability to self-renew and to differentiate into particular odontoblast-like cells under specific cultural conditions in vitro and can give rise to regular-shaped dentin-pulp complex-like structures in vivo.5,7,8 Moreover, DPSCs are derived from a very accessible tissue resource and relatively easy to expand in vitro to provide enough cells for potential clinical applications. All these studies confirm that DPSCs are one of the most promising candidates for dentin tissue repair or regeneration after seeding on tissue engineering implants.
In addition, it is well known that cell behavior on biomaterials is crucial to the formation of new tissues. However, in dentin tissue engineering, the molecular mechanisms underlying the DPSC responses to scaffold signals have not been clarified. The mitogen-activated protein kinase (MAPK) signaling pathway conveys information about the extracellular environment to the nucleus and plays a critical role in the regulation of cell growth, differentiation, and apoptosis.9–11 There are three central elements of MAPK cascades: extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and the p38 kinases. 12 Recent studies have found that MAPK signaling is involved during dental pulp cell differentiation and tooth development.13–15 However, whether the MAPK signaling pathways play an important role in the dentinogenic capacity of DPSCs and whether scaffolds could affect the MAPK signaling pathways of DPSCs have not been assessed.
The aim of this study was to compare the suitability of five different biomaterials as scaffolds for dentin regeneration based on DPSCs and to investigate the molecular mechanisms by which these scaffolds modulate the biological response of DPSCs.
Materials and Methods
Cell culture and analysis
All experimental procedures including the use of animals were performed in accordance with the committee guidelines of the Fourth Military Medical University, which met the NIH guidelines for the care and use of laboratory animals. The DPSCs were isolated as described elsewhere.16,17 The dental pulps were extracted from the incisors of twenty 2-week-old Sprague-Dawley (SD; FMMU Medical Laboratory Animal Center) rats and physically separated from the enamel organs and apical buds. After being washed in sterile phosphate-buffered saline (PBS; Gibco-BRL), the isolated dental pulps were cut into small pieces and digested with type I collagenase (0.6 mg/mL; Sigma) at 37°C for 1 h. Next, the solution was filtered through a 70 μm strainer, centrifuged, resuspended in Dulbecco's modified Eagle's medium (DMEM; Gibco-BRL) supplemented with 10% fetal bovine serum (FBS; Gibco-BRL), 0.292 mg/mL glutamine (Invitrogen), 100 units/mL penicillin G, 100 μg/mL streptomycin, and 50 μg/mL ascorbic acid (Sigma) and incubated at 37°C in 5% carbon dioxide. The DPSCs were enriched by collecting multiple colonies, and the cells at the third passage were used in each experiment.
Colony-forming assay
Single-cell suspensions of DPSCs (1×103 cells) in DMEM containing 15% FBS were seeded into a culture plate (Corning) and then incubated at 37°C in 5% carbon dioxide. After 10 days of cultivation, DPSCs were observed and photographed under a phase-contrast inverted microscope (Olympus Optical).
Analysis of differentiation ability of DPSCs
DPSCs were analyzed for their capacity to differentiate toward adipogenic or osteogenic lineages. A total of 2×104 cells were seeded into each well of a six-well plate. At 80% confluence, the DPSCs were cultured in adipogenic and osteogenic medium, respectively. Adipogenic medium was DMEM supplemented with 10% FBS, 0.5 mM isobutyl methylxanthine (Sigma), 2 μM insulin (Sigma), and 10 nM dexamethasone (Sigma). Osteogenic medium was DMEM supplemented with 10% FBS, 100 nM dexamethasone, 5 mM β-glycerophosphate (Sigma), and 50 μg/mL l-ascorbic acid (Sigma). The control group was cultured in DMEM plus 10% FBS. After 14 days of cultivation, the samples were fixed with 4% polyoxymethylene for 30 min. The adipogenic and osteogenic differentiation was assessed using Oil Red O (Sigma) staining and Alizarin Red S (Sigma) staining, respectively.
Reverse transcription–polymerase chain reaction
DPSCs incubated with osteogenic medium were harvested after 7 days. Total RNA was extracted by using the TRIZOL® Reagent (Invitrogen Life Technology), and first-strand cDNA synthesis was performed according to the manufacturer's protocol. Reverse transcription–polymerase chain reaction (RT-PCR) was carried out including primers to the following genes: dentin sialophosphoprotein (DSPP; GenBank accession no. NM 012790), dentin matrix protein-1 (DMP-1; GenBank accession no. NM 203493), and β-actin (GenBank accession no. NM 031144). The primer sequences are as follows: (1) DSPP-sense, 5′-AGA CAC GGG TTC TGG TGA TGG T-3′, and DSPP-antisense, 5′-GTC AGA CTC CCC TTG CTT TGG G-3′; (2) DMP-1-sense, 5′-TCG CTG TCA CCT TGC TCC TC-3′, and DMP-1-antisense, 5′-GAC AAG ACC CCA GCA GTG AG-3′; (3) β-actin-sense, 5′-TGG AAT CCT GTG GCA TCC ATG AAA C-3′, and β-actin-antisense, 5′-TAA AAC GCA GCT CAG TAA CAG TCC G-3′. β-actin was used as an internal control.
Materials preparation
Ceramic bovine bone
CBB was derived from calcinated fresh bovine bone as previously described. 18 Briefly, cancellous bone was cut from the middle of the femoral heads and cut into small cubes of approximately 1 cm3. The obtained shafts were deproteinized with NaOH solution and, after washing, calcinated at 850°C for 4 h.
Demineralized dentin matrix
DDM was prepared from extracted incisors of twenty 8-week-old SD rats as previously described. 4 Briefly, the outer enamel, inner dental pulp, and predentin tissues were removed from each tooth by using a diamond bur until the dentin was exposed all over the surface. All specimens were mechanically prepared into 200–280 μm granules and washed with distilled water. The granules were exposed to 17% EDTA (Sigma), pH 8.0 at 4°C for 10 min to demineralize and then washed in distilled water for 12 h to remove the remaining EDTA.
Small intestinal submucosa
SIS was prepared as previously described.19,20 Briefly, sections of fresh porcine jejunum were obtained from a local slaughterhouse. The fat was first removed from porcine jejunum, followed by carefully washing with water. Next, the SIS was obtained by mechanical removal of the tunica mucosa, the serosa, and tunica muscularis. The obtained SIS was cut into lengths of approximately 10 cm and thoroughly rinsed in a saline solution to remove the resident cells. Finally, SIS was lyophilized and stored at 4°C for future use.
Poly-L-lactate-co-glycolate
The 85:15 molar ratio PLGA (Sigma) synthetic scaffold was fabricated as previously described. 21 Briefly, 1 g PLGA was dissolved in 10 mL chloroform, and then 9 g NaCl particles sieved to diameter 100–200 μm was added. The suspension was cast in a 10 cm aluminum foil dish. Subsequently, the samples were lyophilized for 48 h to remove the chloroform solvent. The PLGA/salt mixture was placed in distilled water for 24 h with ultrasonic vibrations to dissolve the salt crystals. Finally, the obtained PLGA scaffolds were lyophilized and stored at 4°C for future use.
Collagen–chondroitin sulfate–hyaluronic acid
Co–CS–HA was prepared as previously described. 20 Bovine tendon Co I, CS, HA, 2-(N-morpholino)ethane-sulfonicacid (MES), 1-ethyl-3-3-dimethylaminopropylcarbodiimide hydrochloride (EDC) and N-hydroxysuccinimide (NHS) were all purchased from Sigma Chemical (Badlapur). Co I was dissolved at 4°C at a concentration of 12.5 mg/mL in a solution of 0.05 M acetic acid, and the pH was adjusted to 7.4. CS and HA were dissolved at 4°C at a concentration of 12.5 mg/mL in a solution of double-distilled water, respectively. Next, CS was added in the Co I solution before HA was added. The ratio of the three elements (V/V/V) was 9:1:1. After being well mixed, the slurry was poured into a 6-well plate and was lyophilized for 48 h. These meshes were subsequently cross-linked for 24 h at room temperature using 40% ethanol-water (pH 5.5) solution supplemented with 50 mM MES, 5 mM EDC, and 5 mM NHS. The cross-linked membranes were then rinsed twice with 0.1 M disodium phosphate for 1 h, twice with 1 M sodium chloride for 2 h, six times with 2 M sodium chloride for 24 h, and ten times with distilled water to remove the residual EDC. Finally, the membranes were lyophilized again and stored at 4°C for future use.
For experiments, CBB, SIS, PLGA, and Co–CS–HA scaffolds were pulverized using a freezer mill (6700; SPEX, Inc.) to yield scaffold granules of 200–280 μm in size. Subsequently, all samples were sterilized with ethylene oxide gas.
Cell cytotoxicity analysis
Conditioned cell culture medium was prepared from various scaffolds according to the “Biological Evaluation of Medical Devices” test for in vitro cytotoxicity (GB/T16886.5-2003) as previously described. 22 Briefly, 2 cm3 of each of the five different scaffolds were soaked in 10 mL serum-free DMEM and incubated in a humidified 37°C, 5% CO2 incubator for 24 h. The mixture was then centrifuged for 10 min at 2000 g, and the supernatant was sterilized through a 0.22 mm filter and supplemented with 10% FBS and antibiotics before use.
For the detection of cell cytotoxicity, a gradient of the extract dilutions was used (to 1/2, 1/4 and 1/8). Approximately 2×104 DPSCs were seeded into each well of a 96-well plate. After 12 h of cultivation, the culture medium was replaced by the growth medium supplemented with or without various concentrations of scaffold extracts and then cultured for 3 and 7 days. The MTS assay (CellTiter96®AQueous; Promega) was performed. The optical density values were determined at a wavelength of 490 nm.
Cell proliferation analysis
The growth behavior of the cells on the scaffolds was studied in stationary culture conditions as follows. Various volumes of the scaffolds (10, 20, and 40 mm3) were placed into the 96-well plates. After rehydration of the scaffolds, DPSCs were seeded into the 96-well plates at 2×104 cells/well and cultured for 3 and 7 days. The MTS assay was performed.
Alkaline phosphatase activity analysis
Approximately 2×104 DPSCs were seeded on various volumes of five scaffolds in the 96-well plates. After 3 and 7 days of cultivation in osteogenic medium, the alkaline phosphatase (ALP) activity of DPSCs was detected using an ALP assay kit (JianCheng Co.).
Flow cytometry analysis
Approximately 2×104 DPSCs were seeded on 20 mm3 scaffolds in the 24-well plates and cultured for 5 days. After treatment with 0.25% trypsin (Sigma), single-cell suspensions of DPSCs were harvested through a 70 μm strainer as previously described. 23 After fixing in ice-cold 75% ethanol at 4°C overnight, the cells were washed twice with PBS, resuspended in an RNase solution (0.02 mg/mL; Sigma) containing propidium iodide (PI, 0.02 mg/mL; Sigma) for 30 min, and subjected to Elite ESP flow cytometry (Beckman Coulter) for cell cycle analysis. For apoptosis analysis, single-cell suspensions of DPSCs were washed twice with PBS, incubated for 15 min with fluorescein-conjugated annexin V and PI, and analyzed by using the same flow cytometer.
Scanning electron microscopy
Approximately 2×104 DPSCs were seeded on 20 mm3 scaffolds in a 24-well plate and cultured for 24 h. Cell-scaffold complexes were fixed in 3% glutaraldehyde for 24 h and dehydrated through a series of grade ethanol from 70% to 100%. The specimens were placed in a vacuum oven overnight at room temperature before being coated with gold for examination by scanning electron microscopy (SEM; Hitachi S-3400N). Scaffolds without cells were used as controls.
Real-time quantitative PCR
DPSCs on scaffolds incubated with osteogenic medium were harvested after 7 days. Real-time quantitative PCR (real-time PCR) was performed with a 7500 Fast Real-Time PCR System (Applied Biosystems). Primer sequences for bone sialoprotein (BSP; GenBank accession no. NM 012587), osteocalcin (OCN; GenBank accession no. NM 013414), DSPP, DMP-1, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH; GenBank accession no. NM 017008) were as follows: (1) BSP-sense, 5′-TGT GGA ATG GTG CTA CGG TCT C-3′, and BSP-antisense, 5′-GAT CAA CAG CCC TGA TTT ACG ATG-3′; (2) OCN-sense, 5′-GGT GCA GAC CTA GCA GAC ACC A-3′, and OCN-antisense, 5′-AGG TAG CGC CGG AGT CTA TTC-3′; (3) DSPP-sense, 5′-CAG CCA GGC AGA AGC ATG TC-3′, and DSPP-antisense, 5′-CCA TTC CAT GGT GTC CGT TG-3′; (4) DMP-1-sense, 5′-CAT GGC TGA GGT GTG ACC AGA-3′, and DMP-1-antisense, 5′-TCT CCA GGC TTT GCT ACG TGT G-3′; (5) GAPDH-sense, 5′-GAC AAC TTT GGC ATC GTG GA-3′, and GAPDH-antisense, 5′-ATG CAG GGA TGA TGT TCT GG-3′. The GAPDH gene was used as an internal control. Relative quantization was done by using the ΔCt method by taking the difference (ΔCt) between the ΔCt of GAPDH and ΔCt of each transcript and computing ΔCt. 17 The sequences of primers for BSP, OCN, DSPP, and DMP-1 were designed by Takara (Bio Co Ltd.). Triplicate reaction, and three separate experiments were performed.
Western blot analyses
The DPSCs cultured on the scaffolds for 7 days were analyzed by Western blotting.
After treatment with 0.25% trypsin, single-cell suspensions of DPSCs were harvested through a 70 μm strainer. After being washed twice with PBS, the cells were lysed for 30 min in ice-cold lysis buffer containing 20 mM Tris, pH 7.5, 150 mM NaCl, 1 mM EDTA, 10% glycerol, 1% Triton X-100, 1 mM NaF, 1 mM Na3VO4, and Protease Inhibitor Cocktail (Sigma). Equal aliquots of protein (20 μg) were separated by SDS-PAGE on 10% polyacrylamide gels and transferred onto PVDF membranes. The membranes were blocked with 5% nonfat milk in PBST buffer and incubated overnight at 4°C with primary rabbit antibodies against ERK1/2, JNK, p38, P-ERK1/2, P-JNK, P-p38, and β-catenin at 1:1000 (Abcam). After washing, the blots were incubated with anti-rabbit IgG-HRP conjugated secondary antibodies (Abcam) at a dilution of 1:5000 for 2 h. Finally, the protein bands were visualized with chemiluminescent reagents (WesternBreeze; Invitrogen) in Alpha Innotech Digital Imaging System.
MAPK inhibition studies
The role of MAPK pathways in the odontogenic differentiation of DPSCs on scaffolds were assessed by analyzing ALP activity and real-time PCR after blocking the different pathways. DPSCs seeded on scaffolds were cultured in the growth medium. After 24 h of cultivation, the growth medium was replaced with osteogenic medium in the presence or absence of PD98059 (ERK-specific inhibitor, 20 μM; Abcam), SP600125 (JNK-specific inhibitor, 20 μM; Abcam), or SB203580 (p38-specific inhibitor, 20 μM; Abcam). At day 7, the ALP activity and real-time PCR assays were performed.
In vivo analysis
In vivo transplantation
To further investigate the in vivo biocompatibility and the differentiation of DPSC supported by the five kinds of scaffolds, an in vivo transplantation experiment was performed. After a 7-day pretreatment with osteogenic medium, 2.0×106 DPSCs were seeded on 20 mm3 scaffold granules. Cell-scaffold complexes were implanted in the renal capsule of 8-week-old allogenic SD rat hosts for 6 weeks. The treatments were the same with the control groups (scaffold without cells) as the test group. Sixteen samples for each scaffold (eight samples of scaffold with cells and eight samples of scaffolds without cells) were planted in vivo. Forty 8-week-old rats were used.
Light microscopic assessment
Six weeks after implantation, the transplant samples were harvested. After fixation with 4% paraformaldehyde overnight at 4°C, four samples of each group were decalcified with 10% EDTA (pH 8.0), embedded in paraffin, and 5-μm sections were prepared. For histological analysis, sections were stained with hematoxylin and eosin (H&E) (Baso Diagnostic, Inc.).
Image analysis was performed with an image analysis system (Image-Pro Plus 5.0; Media Cybernetics) to evaluate the level of novel dentin-like tissue formation in different specimens as previously described. 24 Three tissue sections per implant were subjected to measurement.
Real-time PCR
The remaining four samples of each group were used for real-time PCR analysis as previously described. 3 Fibrous tissue capsules and other surrounding tissues were removed immediately after the retrieval of the samples. The scaffolds together with the interstitial tissues were cut into small pieces and transferred to liquid nitrogen immediately. All the following procedures were performed as just described. The odontogenic differentiation of cells was monitored by analyzing the specific markers DSPP and DMP-1.
Statistical analysis
All data were expressed as mean±standard deviation and analyzed by one-way ANOVA with corrections for multiple comparisons using SPSS software (version 12.0; SPSS). p-values <0.05 were considered statistically significant. All experiments were repeated at least thrice.
Results
Isolation and characterization of DPSCs
In the present study, using the limiting dilution technique, we isolated and purified rat DPSCs based on their ability to form adherent clonogenic cell clusters. The isolated DPSCs were capable of forming adherent colonies, in which the majority of the cells presented fibroblastic spindle shapes, and a few were cuboidal or polygonal in appearance (Fig. 1A, B).
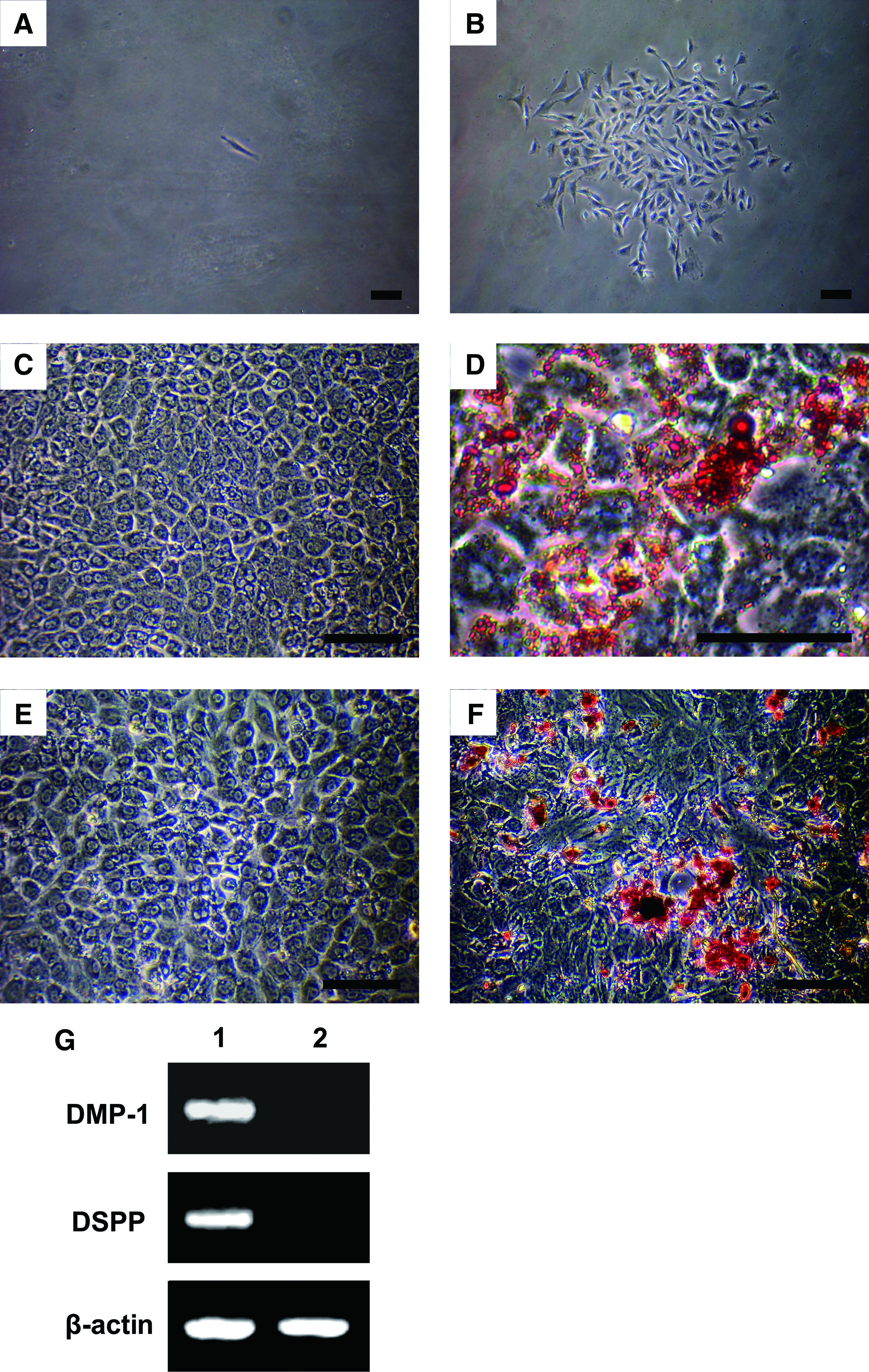
Characterization of rat DPSCs.
To investigate the differentiation potential of DPSCs, multiple colony-derived DPSCs were cultured in adipogenic medium and osteogenic medium, respectively. After 14 days of induction, the cells were able to form extensive amounts of Oil Red O-positive lipid clusters, exhibiting the adipogenic phenotype and Alizarin Red-positive mineral deposits (Fig. 1C–F). Further confirmation was provided by mRNA expression of DSPP and DMP-1 for odontogenic phenotype using RT-PCR (Fig. 1G).
Cell cytotoxicity assay
None of the scaffolds showed significant cytotoxicity to DPSCs, and there were significant differences among the five extracts (Fig. 2A, B). The proliferation was significantly greater when the DPSCs were cultured in lower concentrations of scaffold extracts (to 1/8) than those at higher concentrations. In addition, at the same concentration of extracts, the DPSCs cultured in DDM and SIS extracts proliferated faster than those cultured in CBB extracts on days 3 and 7, whereas DPSCs cultured in CBB extracts proliferated faster than those cultured in PLGA and Co–CS–HA extracts.

The proliferation of DPSCs cultured in growth medium supplemented with different concentration extracts of the scaffolds for 3
Effects of the scaffolds on proliferation of DPSCs
The proliferation of DPSCs cultured on various scaffolds was analyzed by an MTS assay (Fig. 3A, B). The results were generally similar to that of the cell cytotoxicity assay. DPSCs seeded on 20 mm3 scaffolds displayed higher proliferation after 3 and 7 days of cultivation. By contrast, DPSCs seeded on 40 mm3 scaffolds displayed lower proliferation. Under the same volume condition, the proliferation of DPSCs cultured on DDM and SIS showed significantly higher levels than those cultured on CBB on days 3 and 7 after cell seeding, whereas DPSCs cultured on CBB showed significantly higher levels compared with PLGA and Co–CS–HA. The results showed that all these five kinds of scaffolds could enhance cellular attachment and proliferation, whereas the DDM and SIS scaffold would offer a more favorable environment for cellular migration and proliferation. Further evidence was obtained with cell cycle analysis (Fig. 3C–H). DPSCs seeded on the five kinds of scaffolds presented a higher percentage of cells in “S” and “G2M” phases compared with cells seeded on the plates (control), thus suggesting that each of the scaffolds may stimulate DPSCs to shift from “G0G1” phase to “G2M”or “S” phases. Among the five different scaffolds, DPSCs seeded on DDM and SIS presented a higher percentage of cells in “S” and “G2M” phases compared with cells seeded on CBB, whereas DPSCs seeded on CBB presented a higher percentage of cells in “S” and “G2M” phases compared with cells seeded on PLGA and Co–CS–HA. In addition, the apoptosis of DPSCs seeded on the five kinds of scaffolds were not significantly different from each other and, in fact, similar to those seeded on a plastic culture plate, thus suggesting that none of the scaffolds induced significant cell apoptosis (Fig. 3I–N).

Effects of the scaffolds on proliferation and apoptosis of DPSCs.
Effects of the scaffolds on the morphology of DPSCs
SEM observation of all scaffolds showed the internal porous structures (Fig. 4A–E). In the experimental groups, the DPSCs adhered well and gradually spread on the scaffold surfaces after the 24 h cultivation (Fig. 4F–J). When cultured on DDM, CBB, PLGA, and SIS, the cells displayed a flat and fibroblast-like phenotype. Among them, cells seeded on DDM displayed the best adhesion and spread. By contrast, Co–CS–HA caused the DPSCs to show a thin and long morphology. These results indicate that the characteristics of a scaffold can definitively predict the cell morphology and behavior.

Scanning electron micrographs.
Effects of the scaffolds on odontogenic differentiation of DPSCs
The ALP activities of DPSCs cultured on the scaffolds are shown in Figure 5A and B. DPSCs cultured on DDM and CBB displayed higher ALP activities than that of DPSCs cultured on SIS throughout the experiment, whereas DPSCs cultured on SIS displayed higher ALP activities than that of DPSCs cultured on PLGA and Co–CS–HA. In addition, the ALP activities of DPSCs in the groups with 20 mm3 scaffolds were significantly enhanced at all time points, compared with the other two groups. Combined with the results of the cell proliferation assay, a 20 mm3 scaffold volume was chosen as the appropriate amount for further cell culture and in vivo implantation experiments.
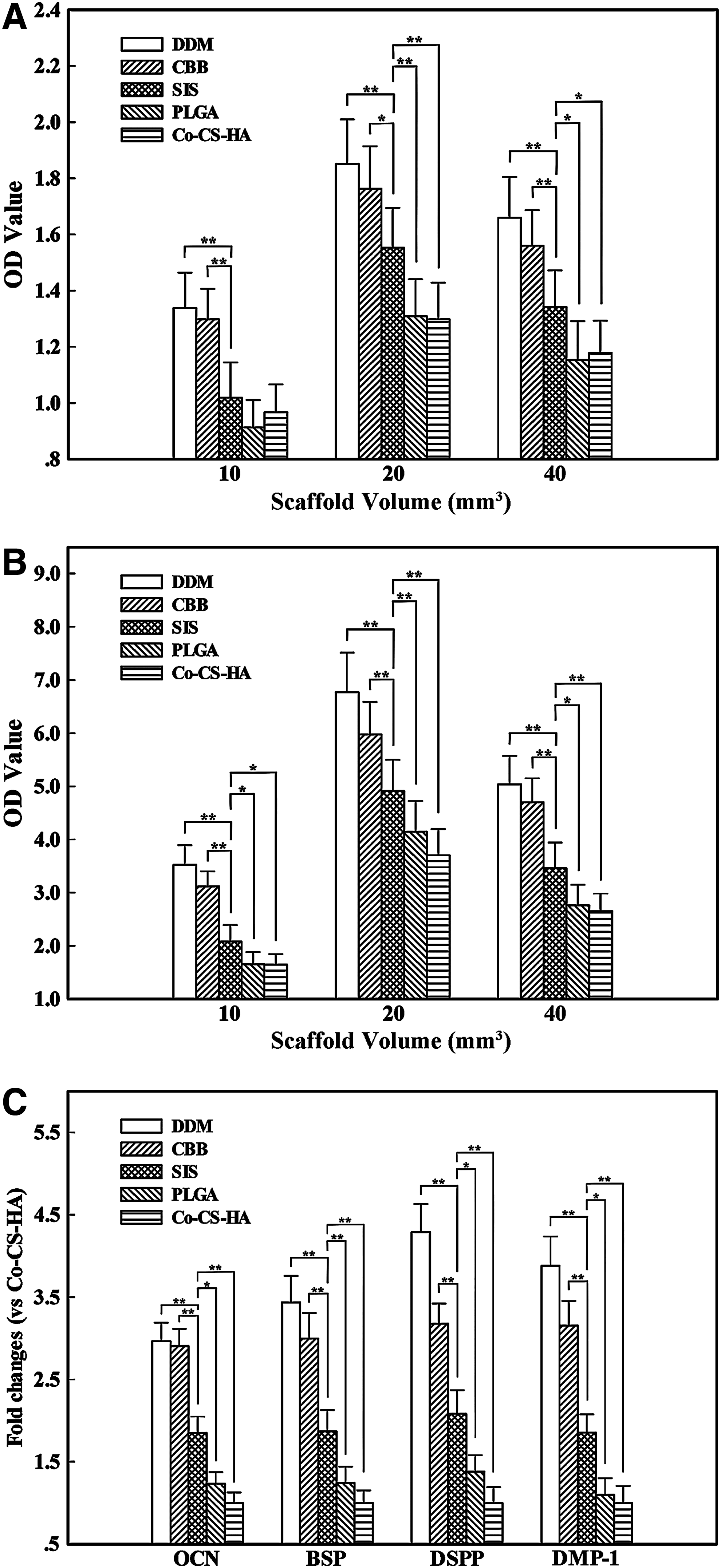
Effects of the scaffolds on odontogenic differentiation of DPSCs.
We next examined relative mRNA expression compared with GAPDH expression of corresponding genes. After 7 days of cultivation in osteogenic medium, the DPSCs seeded on the five kinds of scaffolds expressed BSP, OCN, DSPP, and DMP-1 mRNA (Fig. 5C). The relative quantity showed that the mRNA expression levels of BSP, OCN, DSPP, and DMP-1 were significantly enhanced in DPSCs cultured on DDM and CBB, compared with those cultured on the other three scaffolds.
Effects of MAPK signaling pathways on the differentiation of DPSCs cultured on the scaffolds
To investigate whether the MAPK signaling pathways are involved in the regulation of odontogenic differentiation of DPSCs cultured on the scaffolds, the activities of ERK1/2, p38 and JNK MAPKs were qualitatively measured by Western blotting on day 7 after cell seeding. As shown in Figure 6, DPSCs cultured on the five kinds of scaffolds displayed different levels of ERK1/2 and p38 activities. The phosphorylation of ERK1/2 and p38 were higher in DPSCs cultured on DDM and CBB than in those cultured on SIS, whereas ERK1/2 and p38 phosphorylation levels were higher in DPSCs cultured on SIS than in those cultured on PLGA and Co–CS–HA (Fig. 6A, B), which showed that DDM and CBB could promote ERK1/2 and p38 activation more efficiently compared with the other three scaffolds. In contrast to MAPK/ERK and p38, the scaffolds did not affect the phosphorylation of JNK (Fig. 6C).
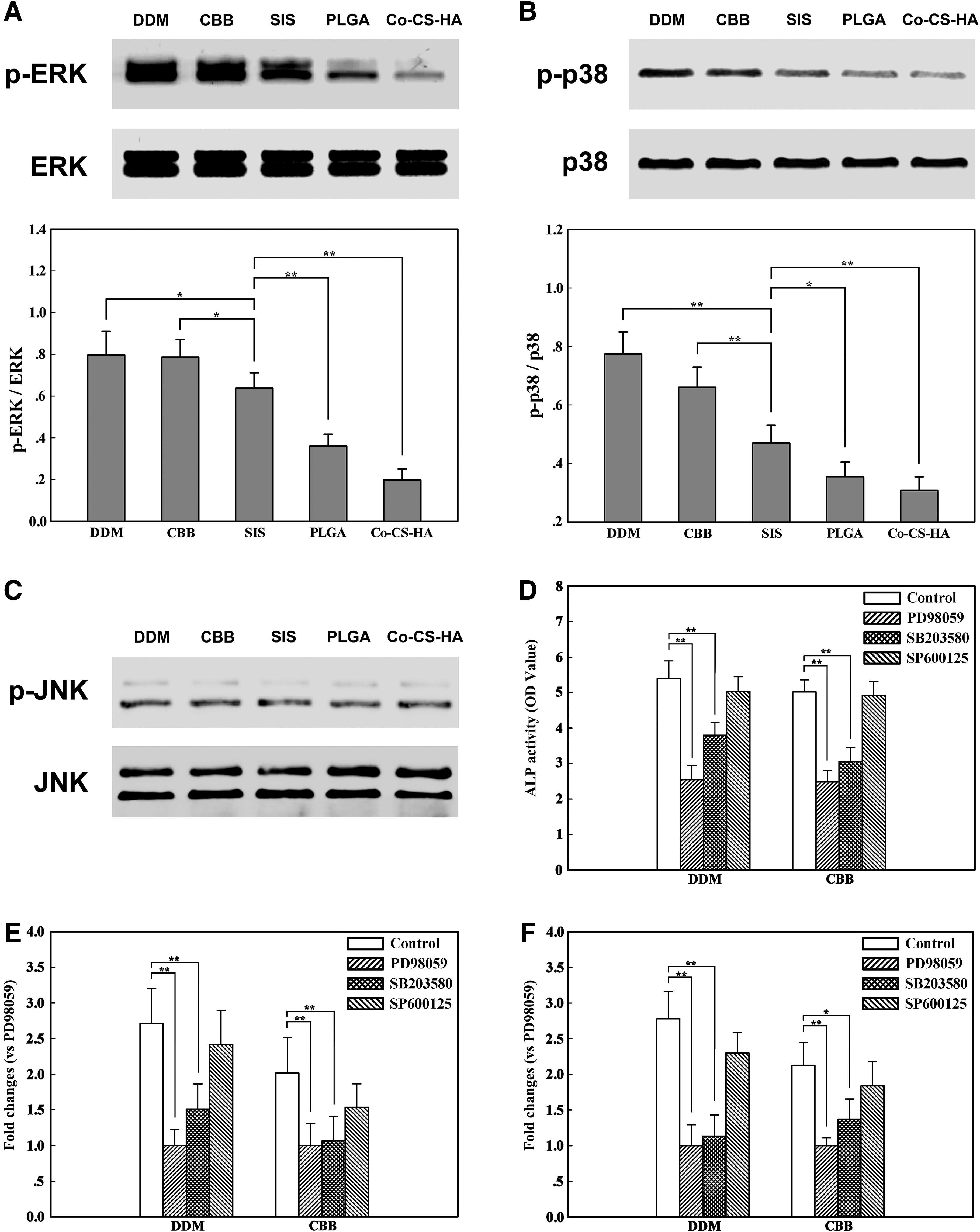
Effects of MAPK signaling pathways on the differentiation of DPSCs cultured on the scaffolds.
We next examined odontogenic differentiation of DPSCs on the scaffolds in the presence or absence of MAPK inhibitors (Fig. 6D–F). Inhibition of the ERK1/2 and p38 pathways resulted in significantly lower levels of ALP activities and mRNA expression of DSPP and DMP-1 of cells cultured on the five kinds of scaffolds at the 7-day time point compared with the untreated controls. However, blocking the JNK pathway by a specific inhibitor, SP600125, did not result in changes in ALP activities and mRNA expression of DSPP and DMP-1 of cells cultured on any of the five kinds of scaffolds. Therefore, these data further indicated that the ERK1/2 and p38 pathways may play critical roles in the scaffold-induced odontogenic differentiation of DPSCs (The SIS, PLGA, and Co–CS–HA groups showed similar profiles to DDM and CBB groups. Data were not shown.).
In vivo analysis
All rats appeared to be in good health throughout the test period. There were no symptoms of severe inflammation at the implantation sites. Six weeks after the implantation, all implants were retrieved with the surrounding soft tissues.
Light microscopic evaluation. Histological observations indicated that all implants after the 6-week implantation period were encapsulated with a thin fibrous tissue capsule. In the control groups, ingrowth of host cells was found in all the scaffold implants unloaded with cells, and the pores among of the scaffold granules were filled with fibrous tissue. No obvious mineralized ECM and dentin-like tissue formation was detected in all scaffold implants without cells (Fig. 7A–E). In the test groups, H&E staining showed that obvious dentin-like tissue formation had occurred in each of implants loaded with cells (Fig. 7F–J). At higher magnification, the dentin-pulp complexes with a regular outline containing dentin, predentin, and pulp cavity, including palisade-like parallel arrangement odontoblast-like cells, were formed in the DDM and CBB groups (Fig. 7K, L), but not in the SIS, PLGA, and CHS groups. However, the amount of newly formed dentin-like tissue was very different among the five kinds of scaffolds. The quantitative analysis showed that the dentin-like tissue areas in the DDM and CBB groups were significantly higher than those in the other three groups (Fig. 7N). Among the SIS, PLGA, and Co–CS–HA groups, the SIS group showed significantly more dentin-like tissue formation than the PLGA and Co–CS–HA groups.
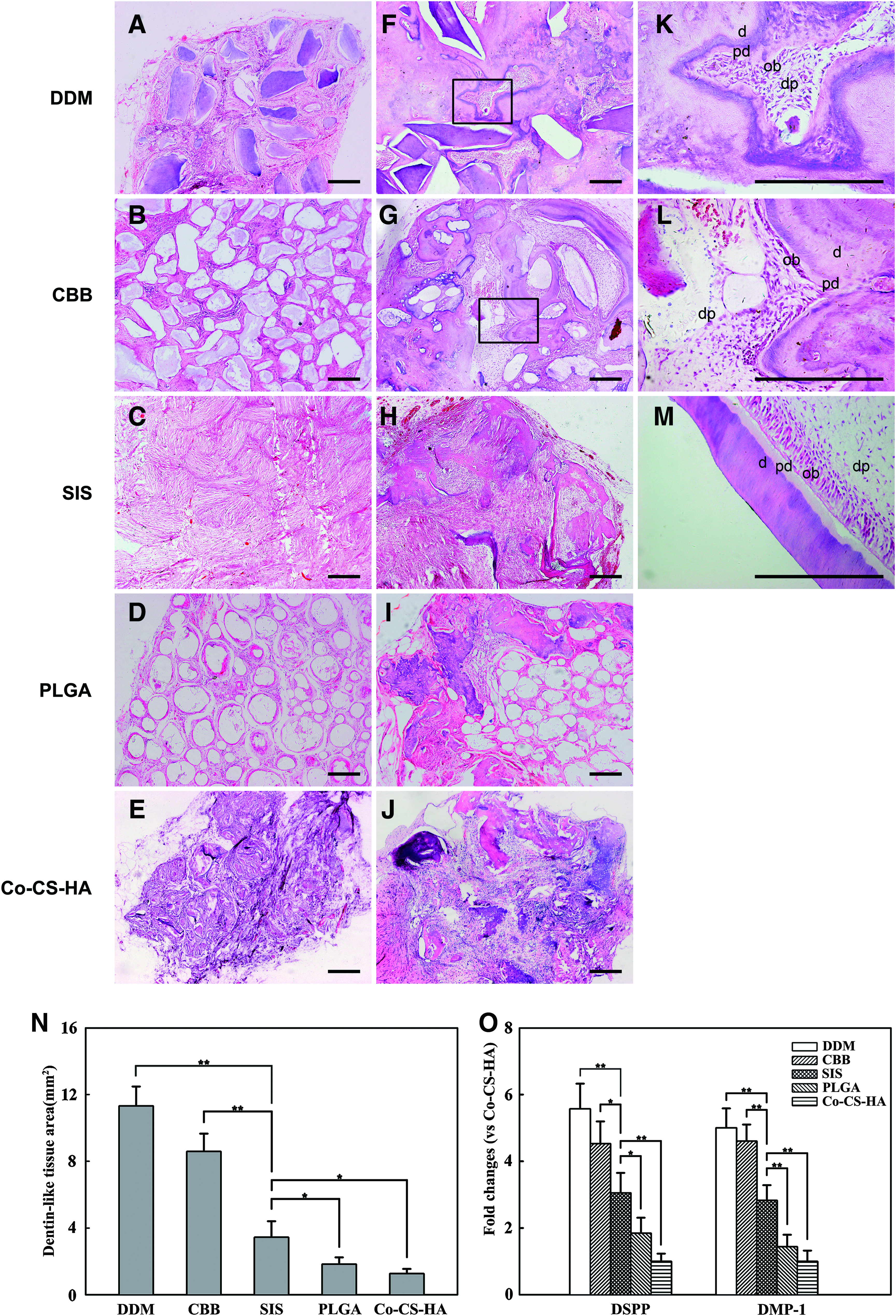
Histology of the implants in vivo.
Real-time PCR. In the real-time PCR analysis, the tissues newly formed by DPSCs expressed DSPP and DMP-1 mRNA (Fig. 7O). The mRNA expression levels of DSPP and DMP-1 were significantly enhanced in DPSCs seeded on DDM and CBB scaffolds, compared with those seeded on the other three scaffolds during the in vivo implantation periods. As for the control groups, no expression of these two genes was found after the 6-week implantation. These results indicated that the tissue formed in vivo carried a tooth-specific phenotype.
Discussion
An essential component in the regeneration of tooth tissue is the use of a proper scaffold material that is able to instruct the seed cells (such as DPSCs) to differentiate into tooth tissue for tooth regeneration and repair. Moreover, the identification of molecular mechanisms by which the scaffolds direct odonto-progenitors into odontogenic differentiation is of great interest to scientists in this field.14,15 In this study, we compared the efficacy of five kinds of scaffolds for DPSC proliferation and odontogenic differentiation in vitro and the regeneration of dentin-like hard tissue in vivo. We further investigated how these scaffolds regulate odontogenic differentiation of DPSC through signaling pathways. Our findings demonstrated that natural scaffolds (DDM, CBB, and SIS) had greater odontogensis capacity than the synthetic and composite scaffolds (PLGA and Co–CS–HA). Among the natural scaffolds, DDM and CBB derived from mineralized tissue had greater odontogensis capacity. We further found for the first time that the scaffolds exert odontogenic conductive effects on DPSCs through the MAPK signaling pathway.
Cell adhesion, spreading, proliferation, and apoptosis are crucial parameters as biocompatibility markers for the cell/biomaterial interaction that determines the cell differentiation and tissue formation. 25 In this study, we found that, under the same 20 mm3 volume condition, none of the scaffolds induced significant cell apoptosis and after 24 h of cultivation, the DPSCs adhered well and spread gradually on all of the scaffold surfaces, indicating that these scaffolds were suitable for the DPSCs in vitro. DPSCs cultured on DDM, SIS, and CBB displayed higher proliferation rates than those on PLGA and Co–CS–HA scaffolds as indicated by the MTS assay and cell cycle analysis. Such a response indicates better cytocompatibility and close interaction of the natural scaffolds with DPSCs. This may be due to the unique advantages of natural scaffolds. DDM and SIS are acellular matrixes, containing many components required for normal cell growth, differentiation, and functioning, including collagen, glycoproteins, proteoglycans, and functional growth factors,26,27 and previous work showed that these factors promote cell growth and differentiation both in vivo and in vitro.4,19 In addition, CBB form sintered bovine cancellous bone that maintains the spongy structure of natural bone and is similar to human natural bone minerals, including the major compositions of HAP [Ca10(PO4)6(OH)2], β-TCP [Ca3(PO4)2], a small quantity of sodium calcium phosphate (NaCaPO4), calcium oxide (CaO), magnesium oxide (MgO), a trace amount of aluminum, iron, magnesium, potassium, silica, sodium, vanadium, and zinc and perhaps carbonated apatite.28,29 A previous study had indicated that inorganic ions (Si, Mg, Ca, P, and Na) could accelerate osteoblasts adhesion and proliferation by delivering the mitogenic stimuli and enhancing channel sensitivity. 30
It is known that if cells are undergoing osteo/odontogenic differentiation, then ALP is widely considered one of the early differentiation markers. 15 In addition, BSP and OCN, which are present in both tooth and bone matrix, have also been used as markers of the potential of hard tissue formation.31,32 DSPP and DMP-1, which play crucial roles during early odontoblast differentiation and late dentin mineralization, have been acknowledged as identifying markers of odontoblasts.33,34 In this study, based on our analysis of ALP activity and the expression of those marker genes, we found that DPSCs cultured on DDM and CBB expressed a higher level of ALP activity and mRNA for odontogenic differentiation-associated genes such as BSP, OCN, DSPP, and DMP-1 as compared with those of SIS, PLGA, and Co–CS–HA, indicating that the DDM and CBB can highly promote cell differentiation compared with the three others. These results are consistent with previous investigations showing that DDM possesses significant odonto-inductive activity, as, aside from conventional acellular matrixes components, it contains some amount of odontogenic bioactive and absorbable ECM molecules (DMP-1, dentin sialoprotein, dentin phosphoprotein, BSP, osteopontin, transforming growth factor-β (TGF-β), and COL-I), which are directly implicated in pulp mineralization or act indirectly on the commitment and differentiation of pulp progenitors during dentin tissue regeneration and dentin repair. 4 In addition, CBB can regulate the cell differentiation by releasing ions into the culture medium or body fluid. Previous studies have shown that Si, Ca, P, and Na ions in bioactive bioceramics may not only regulate the growth and metabolism of various cells but also activate the expression of related genes to enhance the formation and mineralization of ECMs.22,31 The odonto-conductive functionality of these scaffolds prompted us to investigate the underlying mechanisms for this activity.
It has been well documented that cytokine and growth factors such as bone morphogenetic protein-2 and TGF-β often induce changes in cell differentiation and mineralization through the activation of the MAPK pathways.35,36 The bioactive scaffolds have been well documented to coordinate with some cytokines or growth factors in determining differentiation fates.37,38 In the present study, our hypothesis was that the MAPK cascade can serve as a mediator by transducing the signals from the scaffolds into the cells, thereby regulating DPSC differentiation. Our findings confirmed this hypothesis, as we showed that ERK1/2 and p38 phosphorylations were higher in DPSCs cultured on DDM and CBB than in those cultured on SIS, whereas ERK1/2 and p38 phosphorylations were higher in DPSCs cultured on SIS than in those cultured on PLGA and Co–CS–HA. In association with odontogenic gene expression, the increased amount of odontogenic gene expression induced by DDM and CBB was accompanied by the up-regulation of the increased phosphorylations of ERK1/2 and p38 in DPSCs. Further, the induction of ALP activities and mRNA expression of DSPP and DMP-1 of cells by the scaffolds were attenuated by the inhibition of the ERK1/2 or p38 signaling pathway. Therefore, these data indicated that ERK1/2 and p38 may play critical roles in the biomaterials promoted odontogenic differentiation of DPSCs. These results are consistent with those of previous studies, which showed that a β-TCP scaffold promotes osteo-conduction in human osteoblasts through the MAPK/ERK signaling pathway. 38
In the in vivo study, obvious dentin-like tissue formation had occurred in all of the implants loaded with cells after the 6-week implantation, which were further confirmed to be dentin-like tissue by detection of mRNA expression of DSPP and DMP-1 as odontoblast-specific marker using RT-PCR. 39 However, compared with the three others, DPSC/DDM and DPSC/CBB complexes presented significantly larger quantities of dentin-like tissue formation, which presented a dentin-pulp complex with a regular outline containing dentin, predentin, and pulp cavity with a palisade-like parallel arrangement of odontoblast-like cells at higher magnification. These observations suggest that the biocompatibility and odonto-induction of DDM and CBB were more appropriate for the dentin formation in vivo than that of SIS, PLGA, and Co–CS–HA. It was implied that DDM and CBB provide more superior microenvironments where the DPSCs can differentiate into odontoblasts and form typical dentin structure reminiscent of tooth development. Correspondingly, our previous studies have demonstrated that, after induction with DDM, dental follicle cells can differentiate into functional odontoblasts in vitro and complete dentin regeneration. 4 Batouli et al. also showed that DPSCs combined with HA/TCP differentiated into odontoblasts and generated dentin 4 weeks after transplantation. 40 Among SIS, PLGA and Co–CS–HA, the SIS group showed significantly more dentin-like tissue formation. This was consistent with a recent report that bone formation at the defect site in SIS sponge/bone marrow stromal cells (BMSCs) was significantly greater than that in PGA mesh/BMSCs. 19 This result suggests that natural biomaterials are superior to the synthetic or composite scaffold in terms of their effects on cellular function and behavior.
Aside from the chemical nature of the material, its physical properties may directly influence adsorption of adhesion proteins and subsequent cell growth and differentiation.41–43 In this study, all scaffolds were designed with granules of 200–280 μm, which was to provide a large surface area to volume ratio to facilitate cellular-scaffold interactions, and the pore structures among granules could promote the transport of gases, metabolites, and nutrients. Moreover, there appeared to be a limit where increasing biomaterial volume did not lead to continued increase in DPSCs proliferation and differentiation. This result suggested that cellular behavior may be related to the material volume, which is most likely a result of the concentration of active factors released by biomaterials into the local region. The cell cytotoxicity assay further supported this result. Huang et al. also reported similar effects where proliferation was significantly enhanced when BMSC was cultured in lower versus higher concentrations of ceramic extracts. 22
Conclusion
Based on our results, we concluded that all five kinds of scaffolds supported the attachment, growth, and differentiation of DPSCs in vitro, and those combined DPSCs can form dentin-like tissue 6 weeks after implantation. Compared with those on the other scaffolds, DPSCs on DDM and CBB derived from natural mineralized tissue displayed higher odontogenic differentiation, and the cell/scaffold complexes presented significant dentin-pulp structures in vivo. Further, we determined that these scaffolds promoted odontogenic differentiation of DPSCs via the MAPK signaling pathway, and cell behavior including proliferation and differentiation may be affected by the biomaterial volume.
Footnotes
Acknowledgments
This study was supported by grants from the Nature Science Foundation of China (30725042, 30973285, and 31030033) and the National Basic Research Program (973 Program) (2010CB944800).
Disclosure Statement
No competing financial interests exist.