
Abstract
Current therapies for tissue regeneration rely on the presence or direct delivery of growth factors to sites of repair. Bone morphogenetic protein-2 (BMP-2), combined with a carrier (usually collagen), is clinically proven to induce new bone formation during spinal fusion and nonunion repair. However, due to BMP-2's short half-life and its diffusive properties, orders of magnitude above physiological levels are required to ensure effectiveness. In addition, a high dose of this multifunctional growth factor is known to induce adverse effects in patients. To circumvent these challenges, we proposed and tested a new approach for BMP-2 delivery, by controlling BMP activity via carrier binding and localized proteolysis. BMP-2 was covalently bound to gelatin through site-specific enzymatic crosslinking using a microbial transglutaminase. Binding of BMP-2 to gelatin can completely switch off BMP-2 activity, as evidenced by loss of its transdifferentiating ability toward C2C12 promyoblasts. When gelatin sequestered BMP-2 is incubated with either microbial collagenase or tissue-derived matrix metalloproteinases, BMP-2 activity is fully restored. The activity of released BMP-2 correlates with the protease activity in a dose- and time-dependent manner. This observation suggests a novel way of delivering BMP-2 and controlling its activity. This improved delivery method, which relies on a physiological feedback, should enhance the known potential of this and other growth factors for tissue repair and regeneration.
Introduction
Currently, the most common delivery system for BMP-2 involves a degradation/diffusion-based delivery that utilizes the physical properties of the scaffold materials to regulate its availability.14–16 Scaffold materials used include ceramics, 17 nano/microparticles, 18 biodegradable synthetic polymers,19,20 and collagen. 21 In most cases, the scaffolds are constructed in such a way that BMPs are noncovalently immobilized for the purpose of increasing retention time. 22 This allows osteoprogenitor cells to migrate and differentiate into osteoblasts at the repair sites. 17
The major problem associated with this approach is the high initial burst release. To compensate for this initial rapid depletion and decreased amount of BMP-2 remaining in the scaffolds for sustained activity, BMP-2 is always applied in excessive concentrations, which increases the occurrence of adverse effects such as heterotopic bone and bone cyst formation. 22 High doses can also trigger a negative feedback by releasing BMP-2 inhibitors 23 and increasing osteoclastic activity.24,25
To circumvent this problem associated with burst release, the growth factor itself can be modified by enhancing its affinity for a particular substrate by affinity binding. Earlier studies from our laboratory used this approach to enhance the ability of collagen matrices to sequester transforming growth factor beta (TGF-β) and its family members.26–28 We synthesized recombinant fusion proteins of TGF-β and BMP-3 that contained a collagen-binding domain derived from von Willebrand factor to allow noncovalent immobilization on the surface of collagen-based materials. 28 The modified recombinant fusion proteins bound to collagen with considerably higher affinity and promoted higher levels of migration, growth, and differentiation of bone marrow mesenchymal cells in the implantation areas.28–30
Besides noncovalent bounding of growth factor to matrices, covalently tethering of BMP-2 to a substrate was explored to prevent growth factor from diffusion.31,32 An important consideration in using these approaches is to ensure that the chemical process of immobilizing the active growth factor onto the scaffold does not affect its efficacy or biological activity by uncontrolled crosslinking 33 or protein denaturation.34,35 Even though site-specific crosslinking may be achieved through chemical crosslinking, multisteps and complicated chemistry are usually involved.31,36
In contrast to conventional chemical conjugation, substrate-specific enzymatic crosslinking methods have been attempted to generate control delivery of growth factors. Crosslinking enzymes such as factor XIII or tissue transglutaminase (TGase),32,37 and tyrosinase,38,39 which selectively recognize certain amino acid sequences, have been used to immobilize growth factors such as vascular endothelial growth factor (VEGF) 40 and nerve growth factor (NGF) 41 to matrices without compromising their activities. Results from these studies demonstrated the feasibility of using this single step reaction; however, growth factors had to be modified or re-engineered to contain additional sequences to make them compatible with the enzymatic reaction.
In this study, we covalently bound native BMP-2 to a gelatin substrate through an enzymatic approach, and then explored its release through proteolytic digestion. The enzyme selected was microbial TGase, due to its availability and site-specificity. 31 TGase has been proven to be biocompatible and noncytotoxic in crosslinking collagen-based scaffolds in our previous study 42 and that of others.43–46 In addition, gelatin as a derivative of collagen not only provides naturally derived cellular microenvironments, but also is proteolytically digestible. The proteolytic digestion of gelatin thus enables the release of the bound growth factor. BMP-2 activity, before and after crosslinking and during its subsequent release, was studied biochemically and in cell-based assays. The feasibility of a cell-instructive release of BMP-2 for bone induction was also demonstrated in vivo.
Methods
Preparation of materials
Gelatin (Type B 225 bloom; Sigma Aldrich) was dissolved and autoclaved in distilled water to make a 10% gelatin stock. The autoclaved gelatin was aliquoted and stored at 4°C until use. A 2% percent gelatin solution was made by diluting from a 10% gelatin stock at 37°C with BMP-2 buffer (25 mM tricine, pH 7.2, 15 mM sucrose, 1.7 mM NaCl, and 0.01% Tween 80).
Microbial TGase (ACTIVA TI Ajinomoto) from Streptomyces mobaraense was purified using a Sepharose Fast Flow column. 47 Briefly, 3 g of crude TGase was dissolved in a phosphate buffer (20 mM phosphate and 2 mM EDTA, pH 6.0) and gently mixed with 3 mL of pre-equilibrated S Sepharose Fast Flow beads (Sigma). After incubation at 4°C overnight with occasional vortexing, the protein solution and beads mixture were batch loaded into a column. After washing with four volumes of phosphate buffer, TGase was eluted with eluting buffer (phosphate buffer with 800 mM NaCl). Protein concentration was monitored by the Bradford method (Bio-Rad) utilizing BSA as a standard. BMP-2 (R&D Systems) was kept in stock concentrations of 20 ng/μL at −20°C in buffer solution (5 mM glutamic acid, 2.5% glycine, 0.5% sucrose, and 0.01% Tween 80).
Preparation of gelatin/BMP-2 and gelatin-BMP-2 complex
Gelatin/BMP-2
Two percent gelatin was mixed with 20 ng/μL BMP-2 stock at a ratio of 5:2 at room temperature. The final BMP-2 concentration was 4 ng/μL.
Gelatin-BMP-2 complex
The covalent binding of BMP-2 to gelatin was prepared using TGase. TGase was added to the gelatin/BMP-2 mixture at a final concentration of 25 μg/mL. The reaction was carried out at room temperature for 18 h. Mixtures were either prepared fresh or stored at −80°C before determining BMP-2 activity by using the C2C12 cell assay.
Dosage and time response using collagenase
Gelatin-BMP-2 complex was treated with collagenase to evaluate BMP-2 release and its activation profile. The complex was treated with 1 U/mL (final) bacterial collagenase (190 U/mg, Type 2, Worthington) at 37°C for 1 h. Samples were collected and taken for the C2C12 cell-based BMP-2 activity assay.
For the dose–response study, the gelatin-BMP-2 complex was treated with collagenase (0.2–2 U/mL for 1 h at 37°C) before samples were taken to the C2C12 cell-based BMP-2 activity assay. For the time course study, a final concentration of 1 U/mL collagenase was added to separate aliquots of the gelatin-BMP-2 complex solution. The solution was incubated at 37°C and retrieved at various time points from 0 to 180 min, and stored at −80°C before assaying for BMP-2 activity.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis
To examine the BMP-2 interaction with gelatin, sample concentrations were increased because concentrations used in the C2C12 cell-based BMP-2 activity assay were too low to be visible on sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). The final concentrations of BMP-2 and TGase in the mixtures were raised to 100 ng/μL and 40 μg/mL, respectively. Gelatin (1 mg/mL) was generated from heat denaturing rat tail type I collagen (lab prepared) by single run of autoclave. Collagenase final concentration was 1.5 U/mL.
Reactants were mixed at 1:1 (v/v) with SDS sample buffer (125 mM Tris pH 6.8, 2% SDS, 0.1% bromophenol blue, and 25% glycerol) and heated at 100°C for 5 min before loading onto the 3%–18% gradient SDS-PAGE gel. The samples underwent electrophoresis at 90 V until the frontier reached the end of the gel. The gels were stained with Coomassie Blue solution (62.5% methanol, 25% acetic acid, and 0.125% Coomassie Blue R250) (Bio-Rad) overnight and destained with 30% methanol and 1% formic acid for 5 h.
Extraction of rat skin matrix metalloproteinase
Within 2 h after the euthanization of a Fisher 344 rat, a 2×2 cm square piece of skin was removed from the abdominal area after being shaved and the surface-treated with 70% ethanol. The dermis was separated from the keratin layer and the attached muscle with a scalpel. The dermis was incubated in phosphate-buffered saline (PBS) containing a 2% antibiotic–antimycotic solution (Mediatech) at 4°C for 15 h. The skin was subsequently rinsed twice with PBS, excised into 0.5×0.5 cm pieces, and placed into a 60 mm tissue culture plate with 5 mL of Dulbecco's modified Eagle's medium (DMEM) containing 10 ng/mL (final) of tumor necrosis factor α (R&D Systems). Aliquots of 200 μL were collected after 1, 2, 3, 4, and 5 days of incubation at 37°C 5% CO2. Matrix metalloproteinases (MMPs) were measured by a gelatinolytic zymograph.
Gelatin-BMP-2 complex treatment with tissue-derived MMPs
Samples of 20 μL from the collected organ culture media were added to each of 100 μL aliquots of gelatin-BMP-2 complex and incubated at 37°C for 1 h. A subsequent C2C12 cell-based BMP-2 activity assay was conducted on the gelatin-BMP-2 complex solution treated with organ culture media. Collected organ culture media without gelatin-BMP-2 complex was also assayed for BMP-2 activity as a control.
Gelatinolytic zymograph assay
Media collected from the organ culture were analyzed for MMP activity through a gelatin zymograph as described. 48 In short, the culture medium was mixed with sample buffer at a 1:1 ratio (250 mM Tris pH 6.8, 10% SDS, 50% glycerol, and 2.5 mg/mL of bromophenol blue) without reducing agent or heating. The sample was loaded into a gelatin (5.5 mg/mL) containing 10% acrylamide/biascrylamide (Bio-Rad) and underwent electrophoresis with constant voltage (90 V) for 5.5 h. Afterward, the gel was washed with 2.5% Triton X-100 to remove the SDS, rinsed with 50 mM Tris–HCl pH 7.5, and then incubated overnight at 37°C with the developing buffer (50 mM Tris pH 7.5, 5 mM CaCl2, and 200 mM NaCl) for 16 h. The zymographic activities were revealed by 1 h staining with Coomassie Blue staining solution and subsequent overnight destaining with 30% methanol and 1% formic acid.
C2C12 cell-based BMP-2 activity assay
BMP-2 dose dependently induces alkaline phosphatase (ALP) activity in a C2C12 mouse myoblast cell line 5 ; therefore, activities of sequestered and released BMP-2 can be determined by ALP assay with C2C12 cells. C2C12 cells (ATCC) were seeded onto a 96-well-plate at a concentration of 1.25×104 cells per well with 100 μL of 10% fetal bovine serum (FBS)/DMEM. The plate was incubated at 37°C, 5% CO2 overnight for attachment. The medium was exchanged with the test medium with 200 μL of 1% FBS/DMEM. Aliquots of 10 μL samples were added to each well and incubated for 48 h. At the end of the incubation period, the medium was removed, and the cells were washed twice with cold PBS. The cells were then lysed by adding 30 μL of 0.5% Triton X-100/PBS and undergoing three freeze/thaw cycles. For p-nitrophenyl-phosphate (pNPP) substrate solution, each pNPP tablet (5 mg pNPP disodium salt/tablet; Thermo Scientific) was dissolved in 5 mL diethanolamine buffer (1.02 M diethanolamine 0.5 mM MgCl2 pH 9.8). One hundred microliters of pNPP substrate solution was added to the cell lysates and incubated at 37°C for 30 min. ALP activity was determined by recording absorbance at 405 nm and normalized by protein content using a BCA Protein Assay kit (Bio-Rad).
Bone induction of gelatin-BMP-2 complex in vivo
For implantation, a higher percentage of 5% gelatin instead of the 2% gelatin was used creating a longer lasting scaffold, which will aid in carrying the growth factor. Each gelatin-BMP-2 complex sample destined to be implanted was prepared by mixing 3 μg of BMP-2 (30 μL) into a 200 μL of 5% gelatin solution and incubated with TGase (25 μg/mL, final concentration) overnight. Gelatin/BMP-2 was prepared in a similar manner by mixing 3 μg of BMP-2 in a 200 μL of 5% gelatin solution but without TGase. All samples were lyophilized. Before lyophilization, aliquots of the mixtures, with or without collagenase digestion, were assayed in vitro with C2C12 cells for BMP-2 activity to confirm BMP-2 binding and release. Lyophilized 5% gelatin gels that were crosslinked with 25 μg/mL of TGase were used as controls.
Animal use protocols were approved by Institutional Animal Care and Use Committee of the University of Southern California. A total of 3 Fisher 344 rats (male, 8 weeks, weight 190–210 g) were used in the in vivo study. Muscle pouches were created in the abdominal muscles bilaterally at six sites by sharp and blunt dissection and subsequently packed with lyophilized gels. Samples were harvested after 35 days. Each sample was divided into two, one half fixed in 10% neutral buffered formalin for histology (hematoxylin and eosin, H&E) and the other half homogenized in 0.5% Triton X-100 for ALP activity using pNPP as a substrate. 49 Absorbance was measured at 405 nm after 15 min of incubation at 37°C. The activity was normalized by total protein content using a BCA Protein Assay kit (Bio-Rad).
Statistical analysis
The Student's t-test was performed to evaluate differences in all C2C12 cell-based BMP-2 activity assays. Data were expressed as mean±standard deviation. The Pearson correlation test was used for evaluating the time and dose-dependent effects of collagenase digestion. Statistical significance was set at p<0.05 in all analyses.
Results
De-activation of BMP-2 when crosslinked to gelatin by TGase
BMP-2 is known to transdifferentiate a premyoblast C2C12 cell line by dose dependently increasing ALP activity. BMP-2 alone (Fig. 1A), gelatin/BMP-2 (Fig. 1B), and BMP-2 treated with TGase (Fig. 1F) all induced ALP activity at comparable levels. The high ALP activity suggested that TGase or gelatin had no inhibitory effect on BMP-2 activity. However, when BMP-2 and gelatin mixture was treated with TGase, BMP-2 activity was completely lost as exhibited by ALP (Fig. 1G). This indicated that as BMP-2 and gelatin bonds and forms a complex by the action of TGase, the formation of the complex (gelatin-BMP-2 complex) shields the BMP-2 activity. Significant differences were observed between gelatin-BMP-2 complex and all other BMP-2 containing samples (p<0.001).
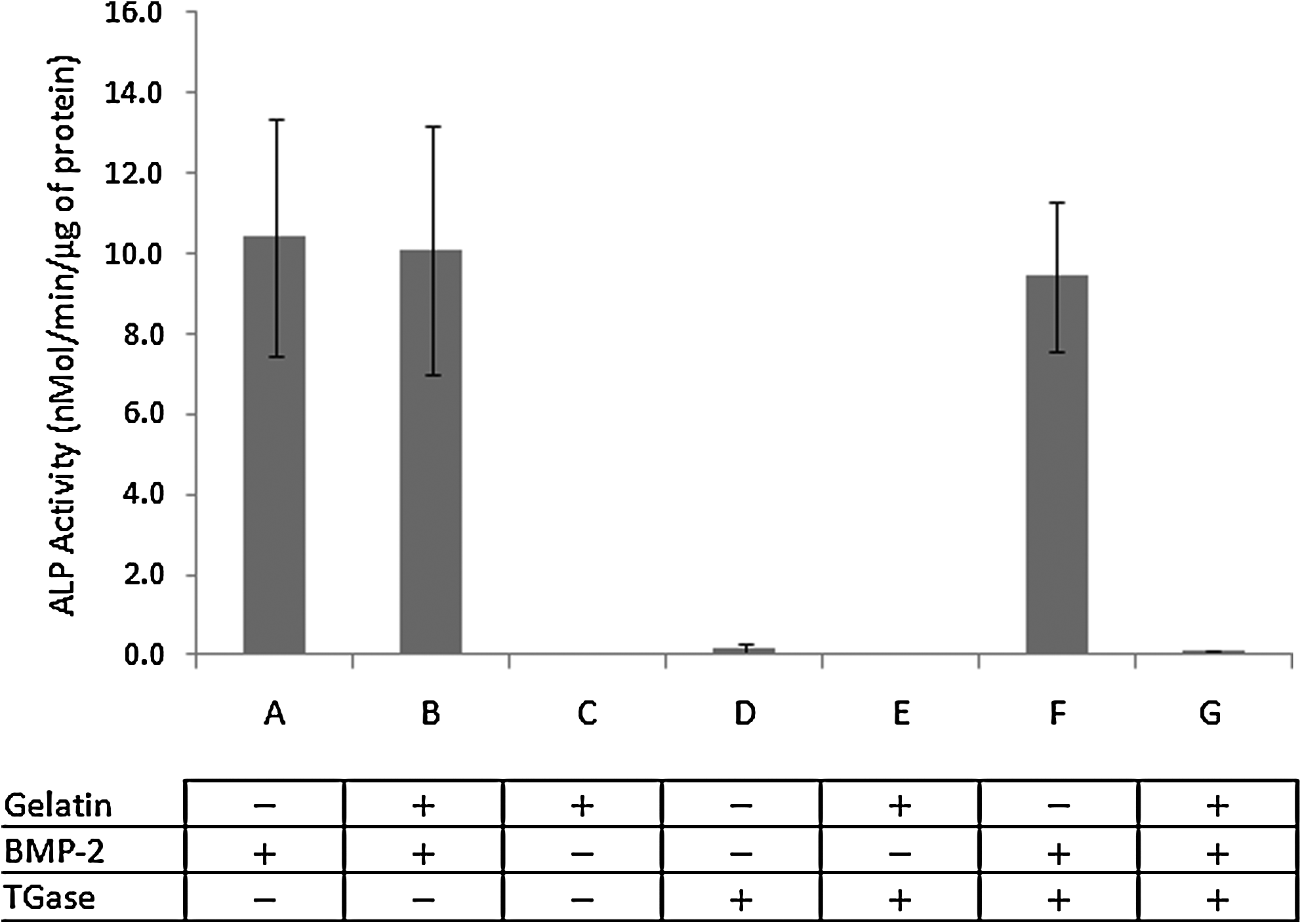
De-activation of bone morphogenetic protein-2 (BMP-2) by gelatin and transglutaminase (TGase). BMP-2 activity was determined by alkaline phosphatase (ALP) activity from the C2C12 cell-based BMP-2 activity assay.
Re-activation of BMP-2 from a gelatin-BMP-2 complex by bacterial collagenase
To determine whether BMP-2 activity can be restored, gelatin-BMP-2 complexes were digested by bacterial collagenase. As shown in Figure 2, collagenase restored BMP-2 activity (Fig. 2D) to a level that did not significantly differ from free BMP-2 (Fig. 2A). Collagenase itself showed no effect on BMP-2 activity (Fig. 2E). This indicated that BMP-2 is not only de-activated by TGase-gelatin crosslinking to gelatin, but that it can also be re-activated by collagenase digestion.
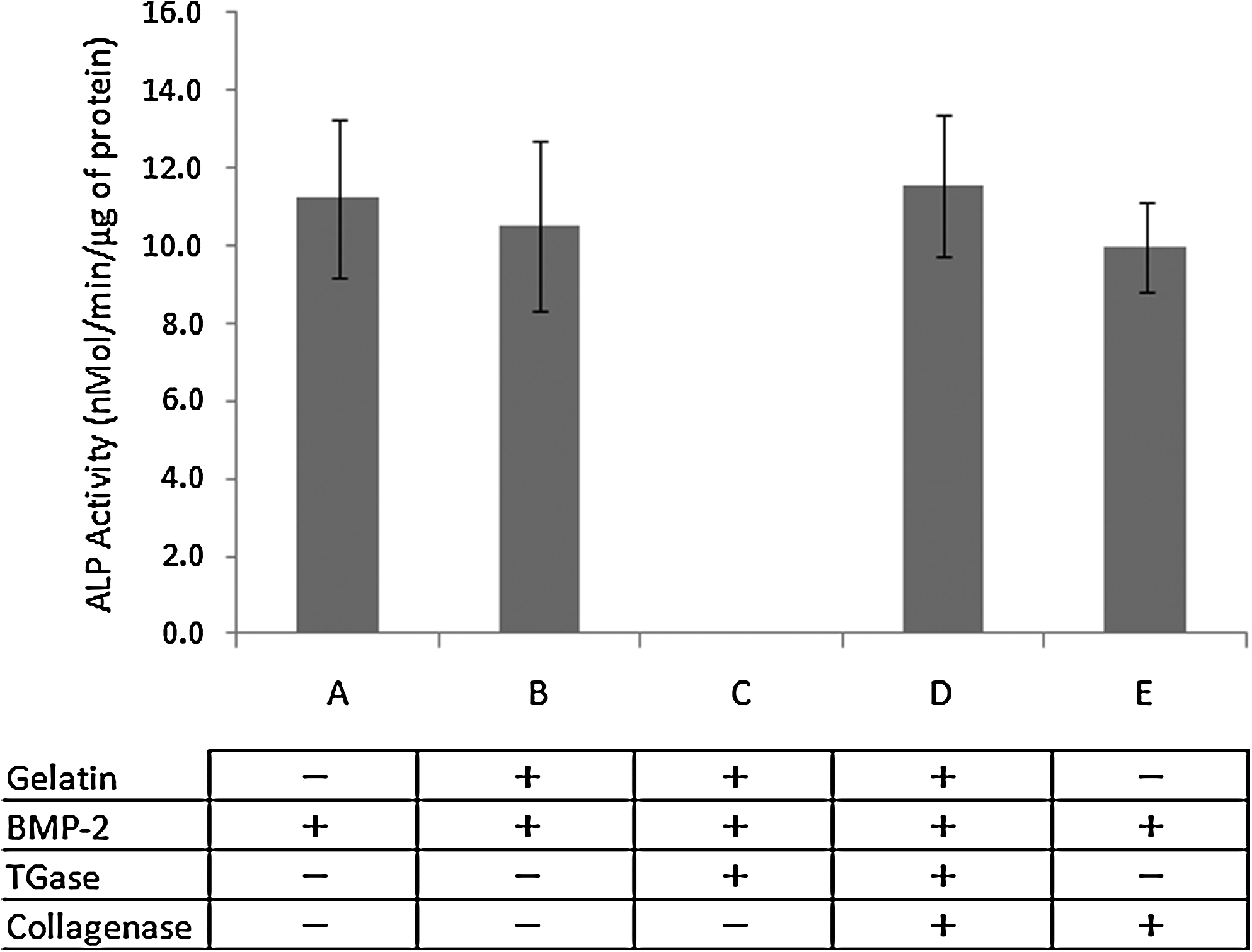
Re-activation of BMP-2 from the gelatin-BMP-2 complex by collagenase. BMP-2 activity of
Collagenase dose- and time-dependent re-activation of BMP-2
To demonstrate controllable activation, various amounts of collagenase were added to each gelatin-BMP-2 complex. Figure 3 exhibits an increased BMP-2 activity as the dose of collagenase was increased (r=0.8870, p<0.001), showing that BMP-2 can be re-activated by collagenase in a dose-dependent manner.
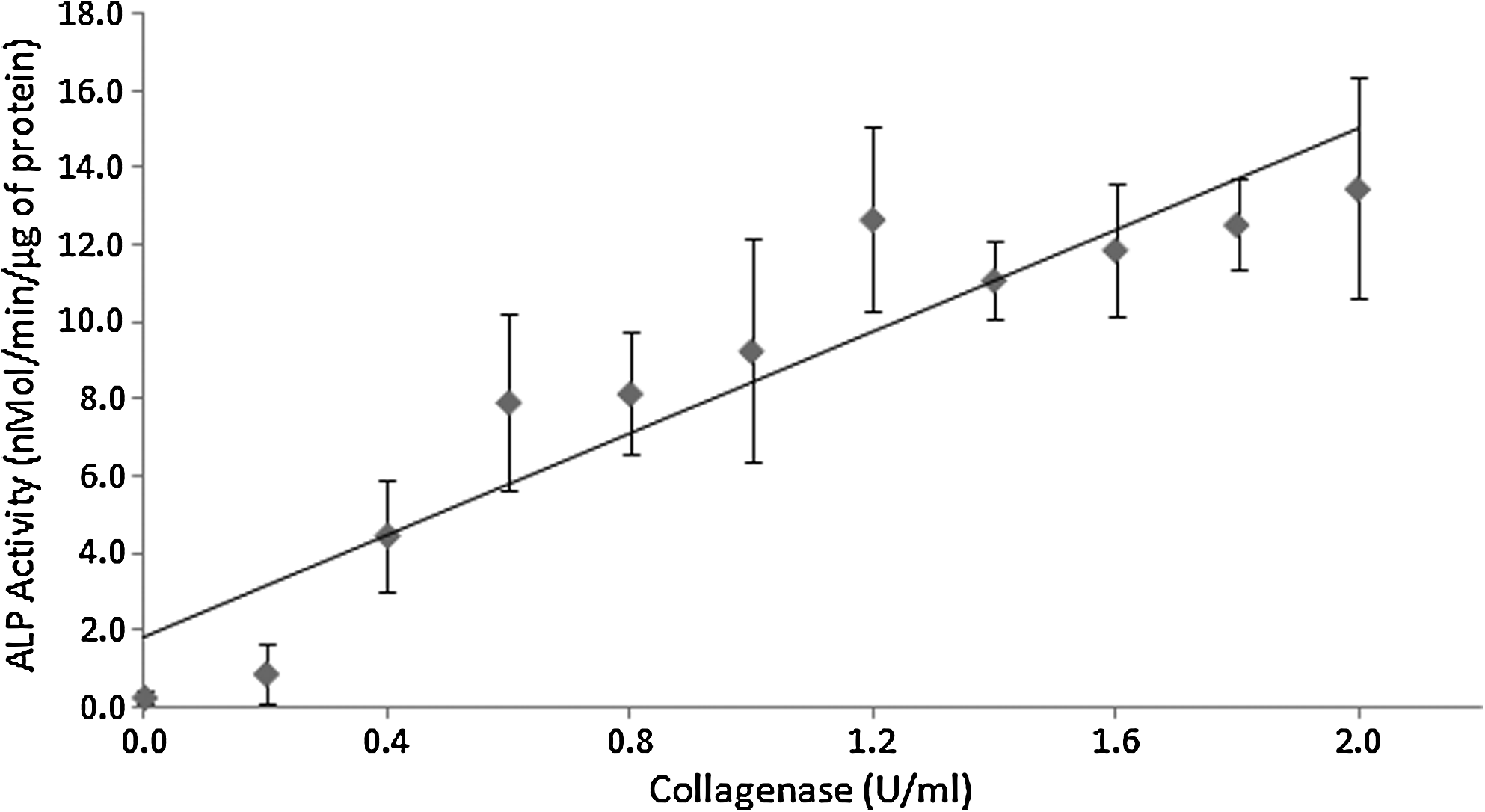
Dose–response relationship of re-activated BMP-2 activity and collagenase concentration. The indicated concentrations were the final concentrations of collagenase that was incubated with gelatin-BMP-2 complex. Aliquots were retrieved after 1 h of incubation at 37°C for each collagenase concentration before subjected to C2C12 cell-based BMP-2 activity assay. The Pearson correlation coefficient and associated p-value were r=0.8870 and p<0.001. Each point represented the mean±SD (n=4).
Temporal effects of re-activation of BMP-2 were evaluated by varying the incubation time using a defined concentration of collagenase. Figure 4 exhibits BMP-2 activity to increase as incubation time was increased (r=0.7388, p<0.001), showing that a prolonged digestion results in increased BMP-2 re-activation from the gelatin-BMP-2 complex.
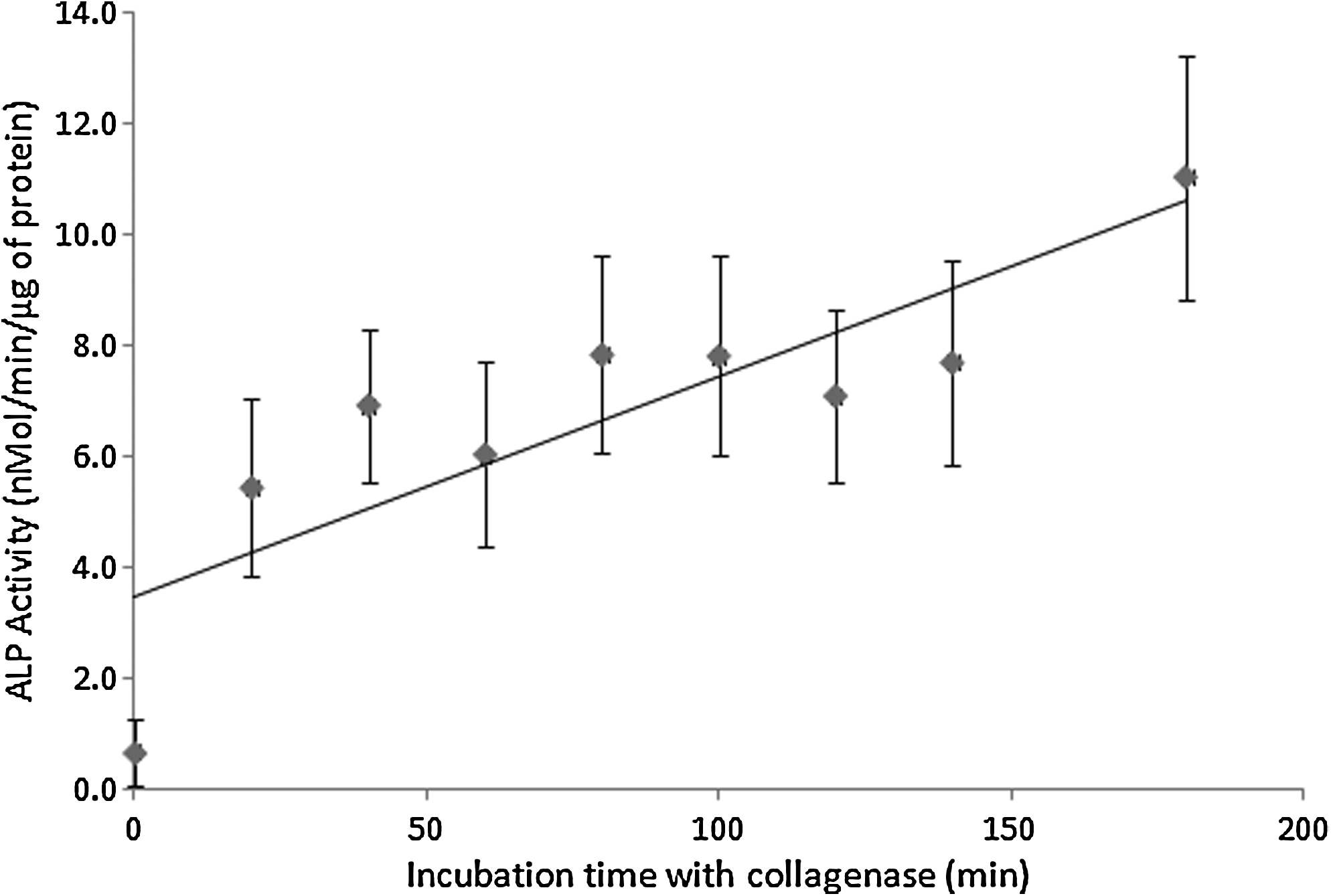
Time course re-activation of BMP-2 from gelatin-BMP-2 complex by collagenase. The gelatin-BMP-2 complex was incubated at 37°C with collagenase with a final incubation concentration of 1 U/mL for all time points. Aliquots were retrieved at various time points and subjected to C2C12 cell-based BMP-2 activity assay The Pearson correlation coefficient and associated p-value were r=0.7388 and p<0.001. Each point represented the mean±SD (n=4).
Examination of the gelatin-BMP-2 complex formation and digestion on SDS-PAGE gel
To elucidate the possible reaction mechanism, SDS-PAGE was used to monitor the protein complex formation and its digested byproducts after collagenase (Fig. 5). SDS-PAGE displayed BMP-2 with a molecular weight of 26 kDa in lane 2, gelatin as band fragments of different molecular weight peptides between 37 and 116 kDa in lane 3 and TGase as a single band with molecular weight around 37 kDa in lane 4. Separate bands of BMP-2 and gelatin were viewed in lane 6, indicating that no binding transpired between BMP-2 and gelatin without TGase. The addition of TGase created a smear out of the fragmented gelatin peptide bands (lane 7), suggesting that TGase generated the crosslinking among the gelatin fragments. The same smear was observed in lane 8 (gelatin-BMP-2 complex), when BMP-2 and gelatin underwent the reaction by TGase.
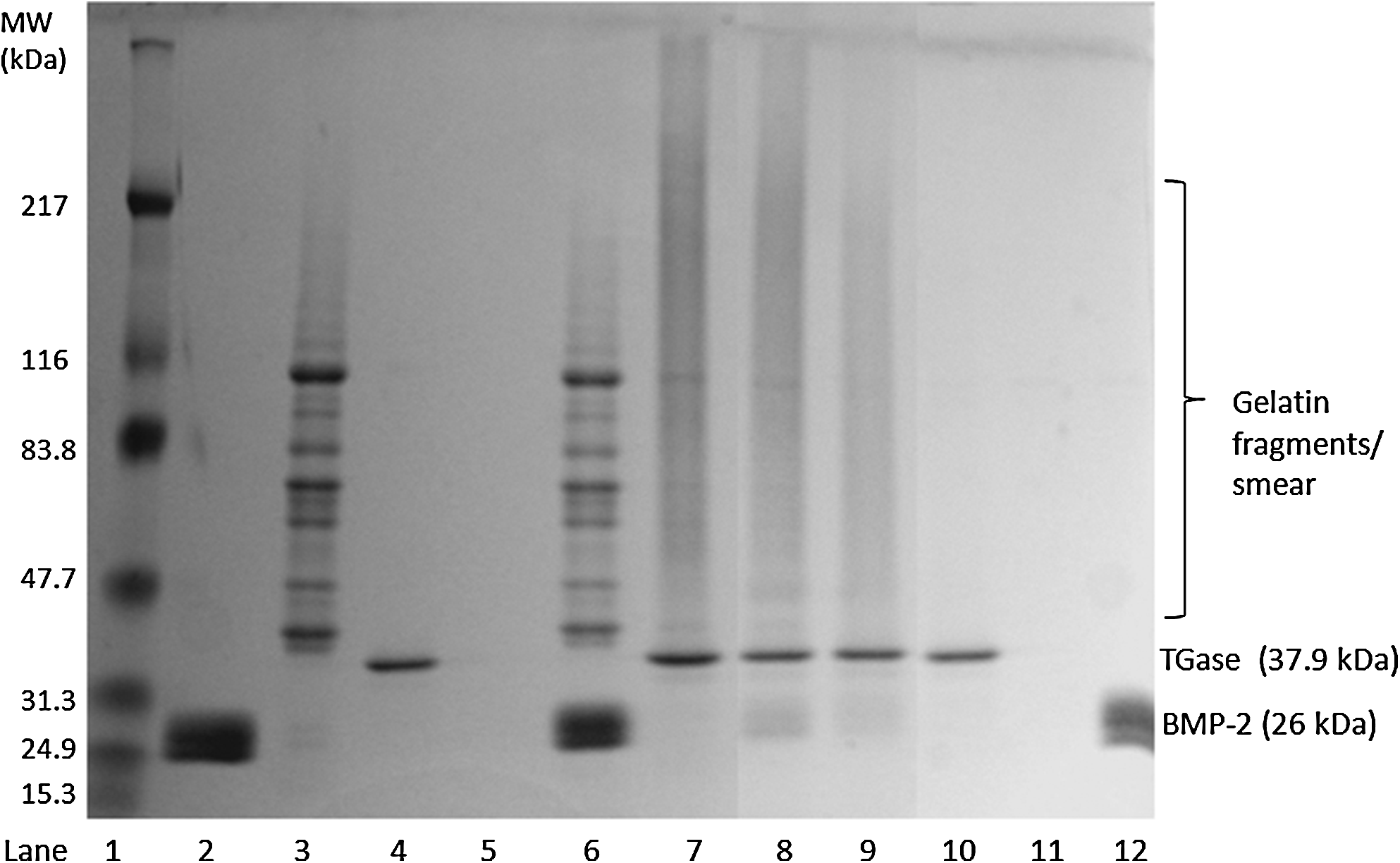
Crosslinking reaction between BMP-2 and gelatin by TGase and their collagenase digested products. Sodium dodecyl sulfate–polyacrylamide gel electrophoresis gradient (3%–18%) was run at 90 V under non-reducing conditions. Lane 1, molecular weight marker; lane 2, BMP-2 alone; lane 3, gelatin alone; lane 4, TGase alone; lane 5, collagenase alone; lane 6, gelatin/BMP-2; lane 7, TGase crosslinked gelatin; lane 8, gelatin-BMP-2 complex; lane 9, gelatin-BMP-2 complex treated with collagenase; lane 10, TGase crosslinked gelatin treated with collagenase; lane 11, gelatin treated with collagenase; lane 12, BMP-2 treated with collagenase.
Collagenase alone was run in lane 5, but concentration was not high enough to be observed on the gel. However, the concentration was enough to digest the gelatin and TGase crosslinked gelatin as evidenced by the absence of bands in lanes 10 and 11. The elimination of all bands of TGase crosslinked gelatin (lane 10) and gelatin (lane 11) show that collagenase completely digested gelatin and TGase crosslinked gelatin. However, in lane 9 (gelatin-BMP-2 complex with collagenase), collagenase digestion left behind a smear indicating components of the complex to be indigestible. In comparing BMP-2 bands in lane 8 to lane 2 (BMP-2 alone) or lane 6 (gelatin/BMP-2), a fading band density of BMP-2 was observed. This suggested an uptake of BMP-2 as it binds to gelatin. Together with the effects associated with digestion, the smear on lane 9 most likely involved the bound BMP-2. Further, BMP-2 in lane 12 showed no fragmentation or signs of digestion when treated with collagenase, suggesting that BMP-2 was the un-degradable component left behind in lane 9. The smear also indicated that small undigested pieces of gelatin remained attached to BMP-2 after collagenase treatment.
Re-activation of BMP-2 by MMPs
To test the effects of mammalian collagenase activity on gelatin-BMP-2 complex, MMPs were obtained from rat skin. Gelatin zymographs revealed that different types of MMPs were expressed from the dermis rat skin layer after prolonged incubation periods in organ culture. Signs of MMP2 production were observed from day 1. MMP9 and MMP3 began to manifest from day 2. All MMPs were observed to increase with longer organ culture incubation (Fig. 6A); thus, a gradient of MMPs was created as daily collections from media were performed.
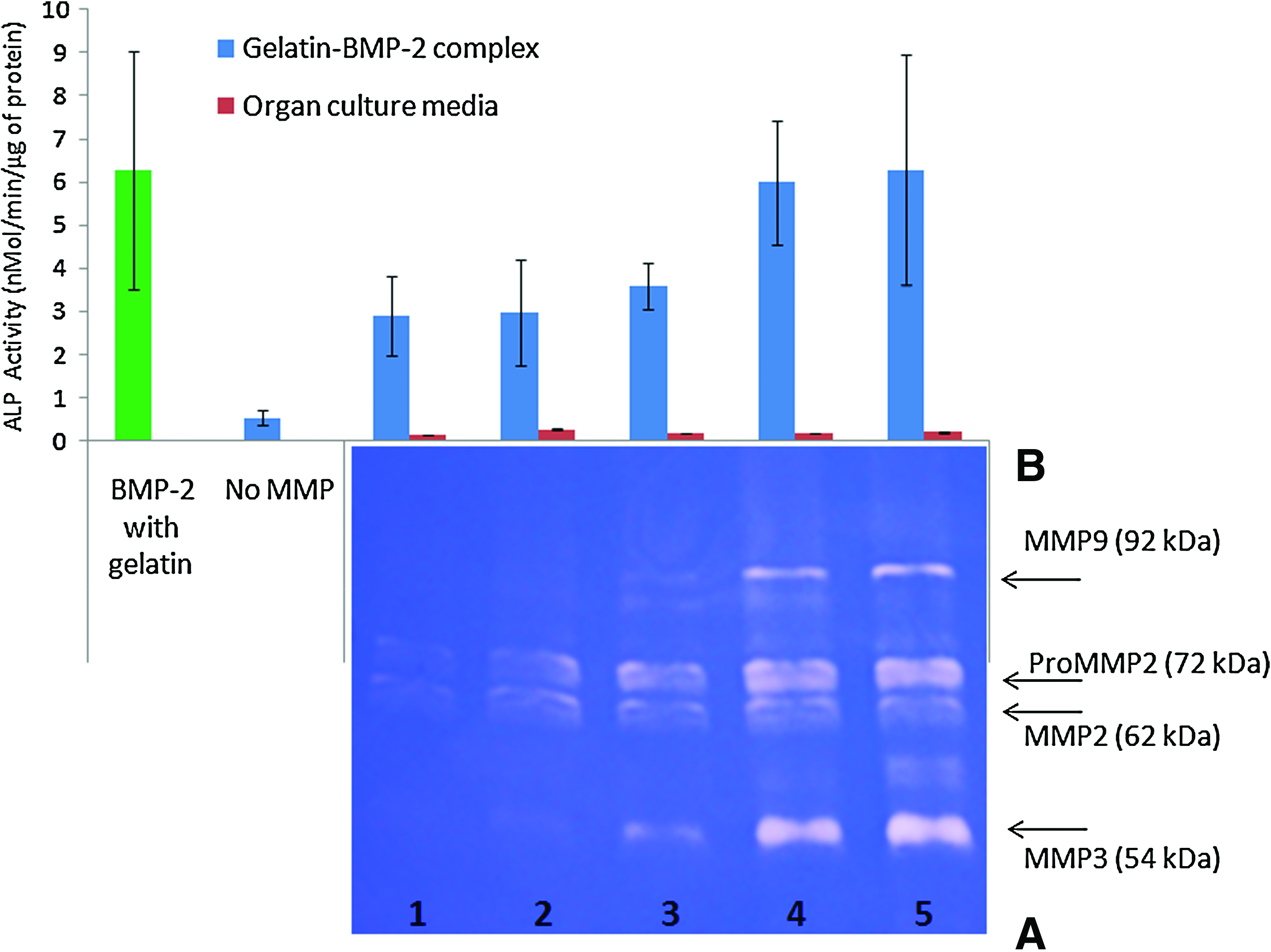
Re-activation of BMP-2 from gelatin-BMP-2 complex by tissue-derived matrix metalloproteinases (MMPs).
For the re-activation of BMP-2, the gelatin-BMP-2 complex was treated with a gradient of MMPs. Results show that media containing native mammalian MMPs re-activated BMP-2 from the gelatin-BMP-2 complex in a dose-dependent manner (Fig. 6B).
Formation of ectopic bone with gelatin-BMP-2 complex
Before conducting the in vivo bone induction test for the gelatin-BMP-2 complex, samples of gelatin-BMP-2 complex before lyophilization were taken and tested by the C2C12 cell-based BMP-2 activity assay. The results confirmed that BMP-2 was completely de-activated and that collagenase digestion of the samples led to re-activation (data not shown). Gelatin crosslinked by TGase 42 was co-lyophilized with a pre-formed gelatin-BMP-2 complex before implantation.
After 35 days of implantation in the abdominal muscle pouch, gelatin/BMP-2, which had no TGase treatment, completely degraded and was unretrievable. The surrounding tissues of the gelatin/BMP-2 implantation site (marked by a suture) were instead taken for histological analysis. All the samples were found to have decreased in size; however, explants with gelatin-BMP-2 complex were larger than TGase crosslinked gelatin control. H&E staining showed active cellular infiltration around and inside these explants. For TGase crosslinked gelatin, we observed it to be partially degraded with fibrous tissue ingrowths and no bone formation (Fig. 7A). However, when implants were supplemented with gelatin-BMP-2 complex, new bone formation was clearly evident. Interestingly, areas of new bone formation were focused at the outer edge of the implant (Fig. 7C). The lack of bone formation in the center part of implant indicated that cell-instructive activation only occurred on the surface of the construct under the current design and time course of the study. The surrounding tissue of gelatin/BMP-2 displayed no bone formation and showed some fibrous tissue growth (Fig. 7B). No gelatin remnants were observed, confirming the complete degradation of the sample. The results of ALP activity from explants correlated with the histological findings. Gelatin-BMP-2 complex group exhibited higher levels of ALP activity compared to gelatin/BMP2 and TGase crosslinked gelatin (Fig. 7D), indicating active bone formation in the gelatin-BMP-2 complex containing explants.
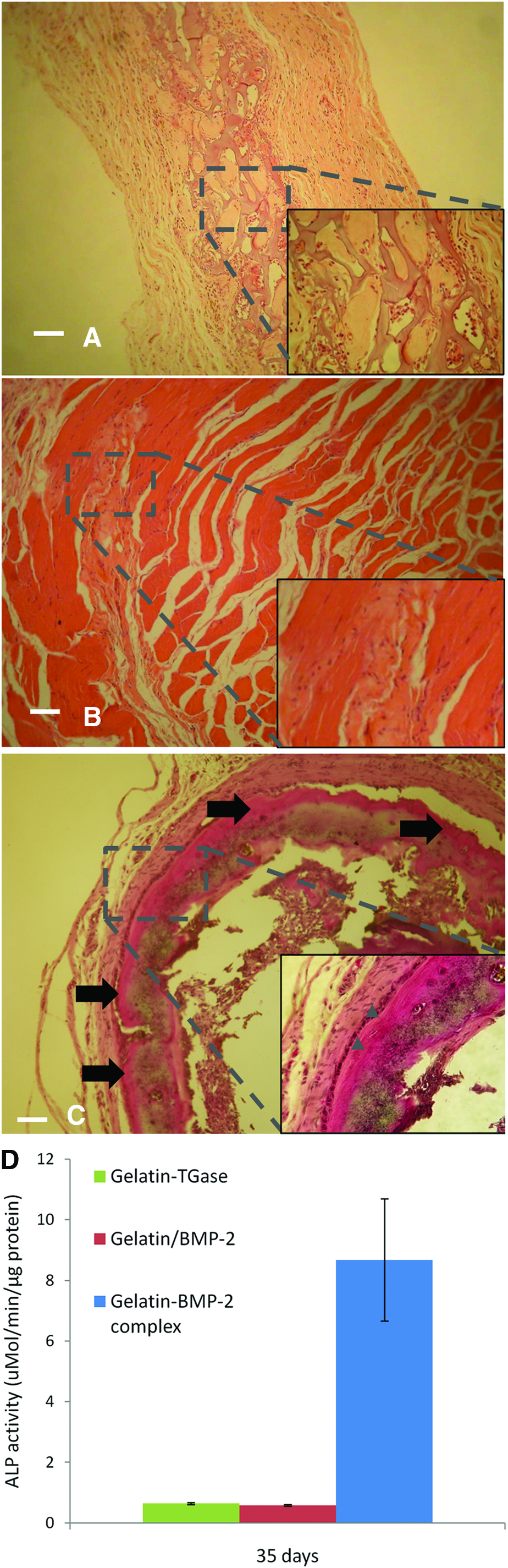
Ectopic bone formation of gelatin-BMP-2 complex. After 35 days of intramuscular implantation, explants underwent histology and ALP assay. Hematoxylin and eosin staining was performed for TGase crosslinked gelatin
Discussion
Immobilization of soluble regulatory peptides on carrier molecules or scaffolds that are later released by cellular activation may provide a powerful means to control cell behavior and enable complex processes of tissue formation and regeneration. To the best of our knowledge, this study is the first to report the novel discovery that the activity of growth factor when conjugated to a carrier protein by enzymatic action can be switched off converting from an active ligand into its latent form. Strikingly, through the action of proteolytic action such as MMPs, the growth factor can be re-activated into its full potential. Schematic mechanism is illustrated in Figure 8. This new drug delivery method by immobilization of bioactive ligand onto a carrier molecule not only changes its biomolecular characteristics (active-latent-active) but also creates signal controlling microenvironment by cell matrix interaction.
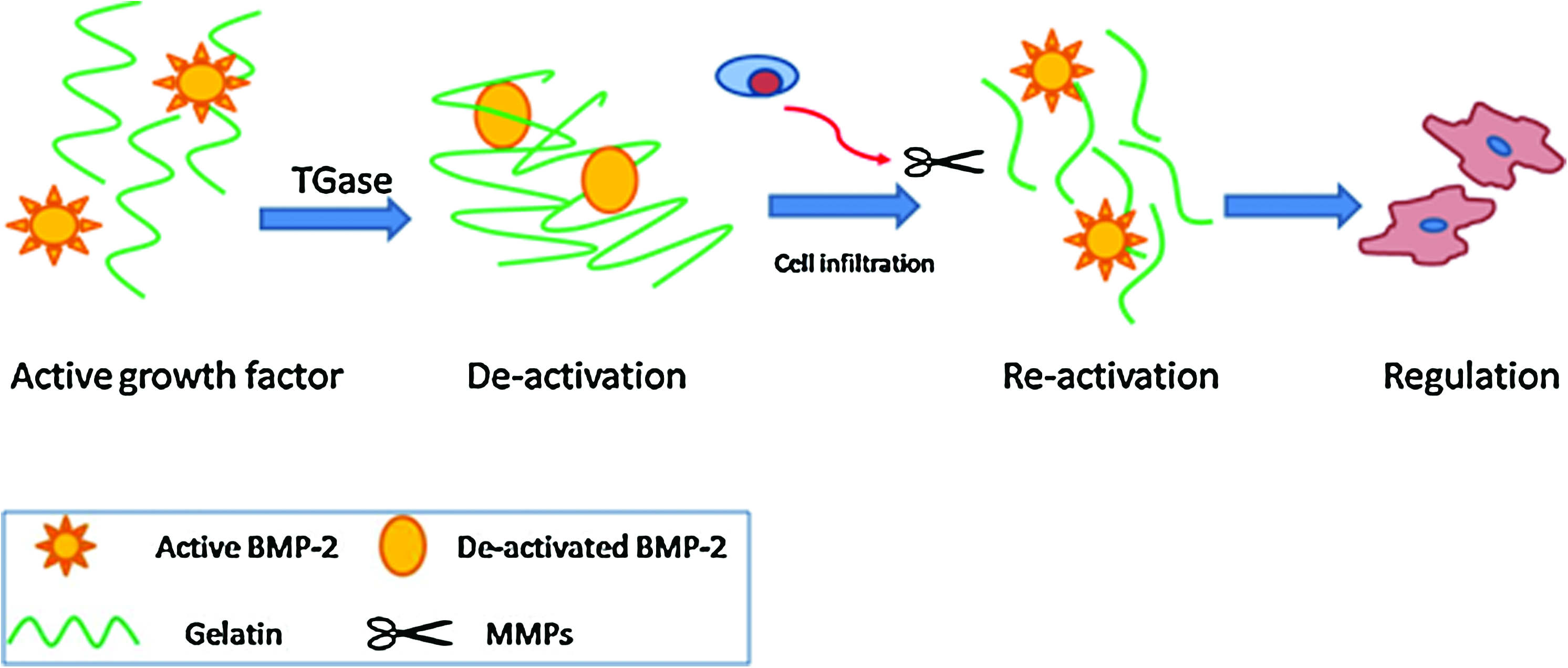
Schematic illustration of the de-activation and re-activation of BMP-2. Active signal molecules, such as BMP-2, are proportionally mixed with protective peptides, such as gelatin. After enzymatic crosslinking by TGase, the gelatin tethers to BMP-2 rendering it to become de-activated. The BMP-2 is re-activated by MMPs digestion secreted from the infiltrated cells. The consequential release of active BMP-2 regulates the cellular functions in the microenvironment. Color images available online at www.liebertonline.com/tea
Several groups have previously demonstrated that enzymatic reaction can be exploited for a variety of applications in drug delivery and tissue engineering. Sakiyama-Elbert et al. 41 modified the NGF sequence to incorporate TGase binding sites with a plasmin degradable domain. The modified NGF was later conjugated to fibrin that would be released by cell-associated enzymatic activity. Similarly, Schmoekel et al. 32 immobilized a modified BMP-2 that contained a binding site for TGase to crosslink and plasmin digestion sites for cell associated release from a fibrin scaffold. Ehrbar et al. 37 used modified VEGF with TGase binding sites covalently linked with a synthetic scaffold containing polyethylene glycol-peptide (PEG-peptide) conjugates. The PEG-peptide was composed of TGase binding sites and MMP1-sensitive degradation sites for cell-activated degradation and VEGF release. In a different study, cell-activated release of fibrin-bound VEGF created by TGase was found to be more effective in promoting vessel growth as compared to passive diffusion of VEGF. 40 Gobin and West 50 found that when tethered PEG-EGF-containing proteolytic degradable sequences was embedded in a PEG diacrylate hydrogel, the diffusion of EGF was prevented, and EGF was released as cells invaded into the scaffold.
In contrast to our study, which uses a native form of growth factor, all these known enzymatically regulated release of growth factors requires the engineering of new fusion proteins that contain exogenous substrates for enzymatic crosslinking and/or for release by enzymatic degradation. In addition to the extra protein expression work, utilizing structurally modified recombinant proteins may be of concern for clinical applications. Our work clearly demonstrates that the shielding of growth factor activity from cells can be achieved through binding a soluble biomaterial to the growth factor at the molecular level. In our case, the active sites are masked with protective peptide, gelatin. This contrasts with the traditional methods of using solid or semisolid biomaterials for creating a separation barrier between the growth factor and cells.
The TGase exhibits its effect by covalently binding the ɛ-amino group of a lysine residue to the γ-carboxamide group of a glutamine. 51 However, for most TGase protein reactions, TGase was found to be highly selective toward only one single or rather few glutamines (Q) among the many other glutamine residues present in proteins. 52 The substrate reactivity toward TGase is determined by the amino acids surrounding the glutamine in the peptide chain. Repeated glutamines were shown to increase reactivity with TGase. 53 Ohtsuka et al. 54 reported that the substrate reactivity is enhanced when the leucine (L), glutamic acid (E), or valine (V) is placed in front of the Q. Ito et al. 44 enhanced microbial TGase crosslinking of RGD to gelatin by synthesizing a peptide with the Ls in front the Q as seen in the resulting sequence RGDLLQ. TGase also shows reactivity preference toward different lysines depending on their location or sequence. 55 If growth factors have endogenous reactive sites for TGase to bind to, the growth factor can be tethered onto a suitable reactive scaffold like gelatin 44 or collagen 46 that contain both reactive lysine and glutamine residues. In our study, unmodified native BMP-2 was crosslinked directly onto gelatin, revealing that endogenous TGase reaction sites are available on BMP-2.
The de-activation of the growth factor as a result of this crosslinking reaction was initially discouraging, but the later re-activation of BMP-2 after the addition of collagenase brought to light an interesting phenomenon. Because collagenase was found to be selective toward digesting both TGase crosslinked and noncrosslinked gelatin and had no effect on BMP-2, gelatin appears to be responsible for de-activation. Although the binding sites where gelatin attaches onto BMP-2 still need to be elucidated, they do not seem to be binding to the active sites of BMP-2. Covalent bonds between ɛ-amino group and γ-carboxamide group are not digestible by collagenase, and if the bonds occurred at the BMP-2 active site, the residual gelatin fragment after digestion would have blocked the BMP-2 active site permanently. Biological active regions of BMP-2 are reported to be located on the two epitopes, the wrist epitope and knuckle epitope. 56 The wrist epitope associates and binds with the BMP receptor IA and the knuckle epitope associates with the BMP receptor type II. It has been shown that mutating regions of one or the other epitope of BMP-2 reduces but not entirely demolishes the biological effect of BMP-2. 56 In this study, BMP-2 activity in the gelatin-BMP-2 complex was completely lost. Since C2C12 cells have both BMP receptor type IA 57 and II, 58 it is possible that the bound gelatin, which serves as a latent sequence of BMP-2, shielded both BMP-2 epitopes that binds to the C2C12 cell receptors. Although BMP-2 activity assays and SDS-PAGE seem to support this pathway, the detailed mechanism requires further investigation.
Construction of MMP-sensitive biomaterial for guided tissue repair has been drawn a lot of attention in recent years.37,59–64 As a denatured form of collagen, gelatin is susceptible to degradation by various proteolytic enzymes, including MMPs. MMPs are usually released by cells at the defect site for purposes of tissue repair and remodeling.65,66 Besides their role in remodeling, MMPs affect other cell functions such as proliferation and apoptosis. Secreted as inactive proenzymes and activated near the cell surface or expressed at the surface in activated form as membrane-type MMPs, these enzymes can cleave virtually all constituents of the extracellular matrix.67,68
In our study, we demonstrated that, besides bacterial collagenase, gelatin-BMP-2 complex can be readily activated by media containing tissue-derived MMPs. It is possible that the presence of other proteases other than MMPs released from the skin could have played a role in the BMP-2 re-activation. Further characterizations of individual MMPs, combination of MMPs, or other cellular proteases are currently under investigation.
A most important feature in this study involves the use of gelatin as a carrier. Gelatin, under the action of TGase, serves dual roles in implant construction, both as a scaffold and as a growth factor switch. On one hand, intracrosslinking of gelatin modulates its resorption rate and prevents it from rapidly degrading in vivo.42,46,69 On the other hand, intercrosslinking between gelatin and BMP-2 entraps the growth factors in the matrix where the BMP-2 remains in its latent state until it switches on by the MMPs secreted from repair cells.
Overall, the multistep osteoinductive events can be described as follows: BMP-2 is covalently immobilized and the bioactive osteoinductive signal switched off in the gelatin matrix when implanted. The MMP-sensitive TGase crosslinked gelatin scaffold guides the inflammatory and osteoprogenitor cells to migrate into the matrix. The cells produced the MMPs that, in addition to degrading the matrix, released BMP-2 into its active form. BMP-2's osteoinductive signaling causes the differentiation of the osteoprogenitor cells in this microenvironment into osteoblasts. Although in vivo ectopic bone formation studies demonstrate feasibility, further selection, modification, and construction of a tissue-specific scaffold and optimization of growth factor release will be needed, before applications for surgical implants that will promote repair of tissue defects in situ can be implemented. Currently, new studies aimed at extending the implantation beyond 35 days are under investigation. The longer implantation duration will answer whether bone will continue to form as the cells infiltrate deeper into the scaffold.
Footnotes
Acknowledgment
The authors wish to thank Dr. Marcel Nimni for his encouragement and his critically reading of this article.
Disclosure Statement
No competing financial interests exist.