
Abstract
Activation of the protein kinase A (PKA) pathway with dibutyryl cyclic adenosine monophosphate (db-cAMP) was recently shown to enhance osteogenic differentiation of human mesenchymal stromal cells (hMSCs) in vitro and bone formation in vivo. The major drawback of this compound is its inhibitory effect on proliferation of hMSCs. Therefore, we investigated whether fine-tuning of the dose and timing of PKA activation could enhance bone formation even further, with minimum effects on proliferation. To test this, we selected two different PKA activators (8-bromo-cAMP (8-br-cAMP) and forskolin) and compared their effects on proliferation and osteogenic differentiation with those of db-cAMP. We found that all three compounds induced alkaline phosphatase levels, bone-specific target genes, and secretion of insulin-like growth factor-1, although 8-br-cAMP induced adipogenic differentiation in long-term cultures and was thus considered unsuitable for further in vivo testing. All three compounds inhibited proliferation of hMSCs in a dose-dependent manner, with forskolin inhibiting proliferation most. The effect of forskolin on in vivo bone formation was tested by pretreating hMSCs before implantation, and we observed greater amounts of bone using forskolin than db-cAMP. Our data show forskolin to be a novel agent that can be used to increase bone formation and also suggests a role for PKA in the delicate balance between adipogenic and osteogenic differentiation.
Introduction

Schematic overview of the cyclic adenosine monophosphate (cAMP)/protein kinase A (PKA) pathway. G-protein coupled receptor (GPCR) ligands bind to their receptors on the cell surface, which results in dissociation of the alpha subunit from its G-protein. The Gα-protein activates adenylate cyclase, which converts adenosine triphosphate into cAMP. cAMP binds to the regulatory subunits of PKA, inducing a conformational change that leads to activation of PKA. PKA in turn phosphorylates cAMP-responsive element binding protein (CREB), which translocates into the nucleus where it binds to the cAMP-responsive element (CRE) to induce transcription of target genes. Dibutyryl-cAMP and its analogue 8-bromo-cAMP directly activate PKA, whereas forskolin activates the more-upstream adenylate cyclase. DMSO, dimethyl sulfoxide. Color images available online at www.liebertonline.com/tea
Materials and Methods
Isolation and culture of hMSCs
Bone marrow aspirates were obtained from donors after they provided written informed consent. hMSCs were isolated and proliferated as described previously. 26 Briefly, aspirates were resuspended using 20G needles, plated at a density of 5×105 cells/cm2, and cultured in hMSCs proliferation medium consisting of alpha-minimal essential medium (α-MEM, Life Technologies), 10% fetal bovine serum (Cambrex Bio Sciences Verviers), 0.2mM ascorbic acid 2-phosphate (Sigma Aldrich), 2 mM L-glutamine (Invitrogen, Life Technologies), 100 U/mL penicillin (Life Technologies), 10 μg/mL streptomycin (Life Technologies), and 1 ng/mL basic fibroblast growth factor (Instruchemie, The Netherlands). Cells were grown at 37°C in a humid atmosphere with 5% carbon dioxide (CO2). Medium was refreshed twice a week, and cells were used for further subculturing or cryopreservation upon reaching near confluence. hMSCs basic medium was composed of proliferation medium without basic fibroblast growth factor, and osteogenic medium was composed of basic medium supplemented with 10−8M dexamethasone (Sigma).
PKA activators
Three different PKA activators were selected: db-cAMP (N 6 ,2′-O-dibutyryladenosine 3′,5′-cyclic monophosphate sodium salt, stock solution 20mM in water), 8-br-cAMP, (8-bromoadenosine 3′,5′-cyclic monophosphate sodium salt, stock solution 20mM in water), and forskolin (from Coleus Forskohlii, stock solution 2mM; 5 mg/mL dimethyl sulfoxide (DMSO) supplemented with basic medium) (all from Sigma). Concentrations of 0.1, 0.5, 0.75, 1.0, and 2.0mM were used for db- and 8-br-cAMP and of 0.01, 0.05, 0.075, 0.10, 0.15, and 0.2 mM. for forskolin
Proliferation
hMSCs were seeded at 5000 cells/cm2 in 24-well plates in triplicate and cultured in basic medium supplemented with denoted concentrations of PKA activators. As a control, cells were cultured in basic medium. A 10% v/v Alamar Blue (Biosource) solution was prepared in basic medium. After 4 and 7 days of culture, the medium was removed, and 1 mL of Alamar Blue solution was added to each well. For background measurements, Alamar Blue solution alone was used. After an incubation period of 4 hours, 200 μL of the solution was transferred to a 96-well plate, and fluorescence intensity was measured using an LS50B luminescence spectrometer (Perkin-Elmer) with an excitation wavelength of 545 nm and an emission wavelength of 590 nm. After measurements, cells were washed with phosphate-buffered saline (PBS), fresh medium was added, and cells were returned to culture.
ALP expression
To determine ALP expression levels, hMSCs were seeded in triplicate at 5000 cells/cm2 in 6-well plates. Cells were kept overnight in proliferation medium to attach, after which the medium was replaced with basic or osteogenic medium supplemented with denoted concentrations of PKA activators. lCells were cultured in basic medium as a negative control and in osteogenic medium as a positive control. After 3 days, cells were trypsinized (0.25% trypsin, Invitrogen Life Technologies), and 1 mL of blocking buffer (PBS with 5% bovine serum albumin (BSA, Sigma) and 0.1% sodium azide (Sigma)) was added and incubated for 30 minutes to prevent a-specific binding. After centrifugation and removal of the supernatant, cells were incubated with 50 μL of primary antibody (anti-ALP, B4-78, Developmental Studies Hybridoma Bank, University of Iowa,) diluted 1:50 in wash buffer (PBS with 1% BSA and 0.05% sodium azide) for 1 hour. For each condition, part of the cells was used for the isotype control to determine a-specific binding. These cells were incubated with immunoglobulin (Ig)G-1 (BD Bioscience, diluted 1:50 in wash buffer), a randomly binding antibody. Cells were then washed twice with 500 μL of wash buffer. Next, 100 μL of secondary antibody (rat anti-mouse IgG1-PE (BD Bioscience), diluted 1:100 in wash buffer) was added. After incubation for 30 minutes, cells were washed again three times, 10 μL of nucleic acid dye solution (Viaprobe, Pharmingen) was added for live/dead staining, and ALP expression was determined using a FACS Calibur measuring 10,000 events (Becton Dickinson, Immunocytometer Systems). FACS data were analyzed using CellQuest software. Cells with the right size (forward scatter) and granularity (side scatter) were gated, and debris and dead cells were excluded from analysis. The percentage of ALP-positive cells was determined, with a minimum of 7500 gated events. Experiments were repeated with cells from at least three different donors.
Reverse transcriptase polymerase chain reaction
hMSCs were seeded in triplicate at 5000 cells/cm2 in 6-well plates and kept overnight in proliferation medium to attach. Then the medium was replaced with basic medium containing various concentrations of PKA activators. As a control, cells were cultured in basic medium or, as a control for BMP-2 target genes, basic medium supplemented with 100 ng/mL BMP-2. After 5 days, RNA was isolated using a Bioke RNA II nucleospin RNA isolation kit (Machery Nagel), and RNA concentrations were measured using an ND100 spectrophotometer (Nanodrop Technologies,). Complementary DNA was synthesized from 250 ng of RNA using iScript (BioRad) according to the manufacturer's protocol. For semiquantitative polymerase chain reaction (PCR), a master mix containing distilled water, forward primer, reverse primer (Sigma Genosys), BSA, and SYBR green I mix (all from Invitrogen) was prepared, and real-time qualitative PCR was performed using a Light-Cycler (Roche). Light-Cycler data were analyzed using the fit points method of Light-Cycler software. The baseline was set at the lower log-linear part above baseline noise, and the crossing temperature (Ct value) was determined. Ct values were normalized to the 18S housekeeping gene, and ΔCt (Ct,control – Ct, sample) was used to calculate the upregulation in gene expression. 27 Primer sequences are listed in Table 1.
IGF-1 secretion
hMSCs were seeded in triplicate at 5000 cells/cm2 in 24-well plates and allowed to attach overnight in proliferation medium. Then the medium was removed and replaced with basic medium supplemented with various concentrations of PKA activators. As a negative control, cells were cultured in basic medium. After 5 days of culture, the medium was removed, and the concentration of IGF-1 in the medium was determined using enzyme-linked immunosorbent assay (human IGF-1, Quantikine, R&D Systems), according to the manufacturer's protocol.
In vivo bone formation
Two- to 3-mm biphasic calcium phosphate particles were prepared and sintered at 1,150°C, as described previously. 28 hMSCs were seeded at 200,000 cells/three particles in proliferation medium. After 4 days, the medium was changed to basic medium or basic medium supplemented with 1mM db-cAMP, 0.1mM forskolin, or 0.15mM forskolin, and cells were cultured for 4 more days. Twenty minutes before surgery, six nude male mice (Hsd-cpb:NMRI-nu, Harlan) were injected subcutaneously with 0.05 to 0.1 mg/kg of buprenorphine hydrochloride and subsequently anesthetized with isoflurane. Particles were implanted in subcutaneous pockets, with three particles per pocket, four pockets per mouse, and six samples per condition. Incisions were closed using Vicryl 5-0 sutures, and after 6 weeks, the mice were killed using CO2. Particles were explanted, fixed in 4% paraformaldehyde (Merck), dehydrated, and embedded in methyl methacrylate (L.T.I. Bilthoven) for sectioning. Undecalcified sections were processed on a histological diamond saw (Leica). Sections were stained with methylene blue and basic fuchsin to visualize bone formation. Quantitative histomorphometry was performed by selecting three sections, with an intermediate section in between, after which a high-resolution (5400 dpi) micrograph was made (Minolta Dimage Scan). Micrographs were pseudocolored (bone in red and scaffold in green), and image analysis was performed using the KS400 version 3.0 software (Carl Zeiss Vision, Oberkochen, Germany). Before measurements, the system was geometrically calibrated using an image of known dimensions. Bone was quantified as the percentage of bone area per scaffold area.
Statistics
All experiments were performed in triplicate. Data were analyzed in SPSS (PASW statistics, SPSS, Inc., Chicago, IL) using one-way analysis of variance, followed by Tukey's multiple comparison test.
Results
PKA activators inhibit proliferation
Three different PKA activators (db-cAMP, 8-br-cAMP, and forskolin) were investigated for their dose-dependent effects on hMSCs proliferation. Concentrations of 0.1, 0.5, 0.75, 1.0, and 2.0mM for db- and 8-br-cAMP and 0.01, 0.05, 0.075, 0.10, 0.15, and 0.2mM for forskolin were used. As a measure for cell number, metabolic activity was determined after 4 and 7 days; Figure 2 depicts values relative to control (basic medium). Six donors were tested, and for forskolin, which was dissolved in DMSO, the solvent alone was included as a control. After 4 days, proliferation was slightly lower compared to control in two of six donors using db-cAMP (Fig. 2A), whereas it did not affect proliferation of the other four. After 7 days, proliferation was inhibited in a concentration-dependent manner in every donor tested; 8-br-cAMP displayed similar effects (Fig. 2B): no change in proliferation after 4 days of treatment and a concentration-dependent decrease in cell number after 7 days. For two donors, 8-br-cAMP did not affect proliferation at all. Treatment with forskolin (Fig. 2C) resulted in a strong concentration-dependent decrease in proliferation already after 4 days, with an even larger decrease after 7 days, which was partly due to the DMSO (Fig. 2D). For all compounds, the level of decrease was strongly donor dependent, varying from 25% to 60% and from 10% to 55% using the highest concentrations of db-cAMP and forskolin, respectively. (For additional data on various donors see Supplementary data S1.) DMSO had similar donor-dependent effects, varying from 0% to 5% in the lowest concentration and 10% to 25% in the highest concentration.
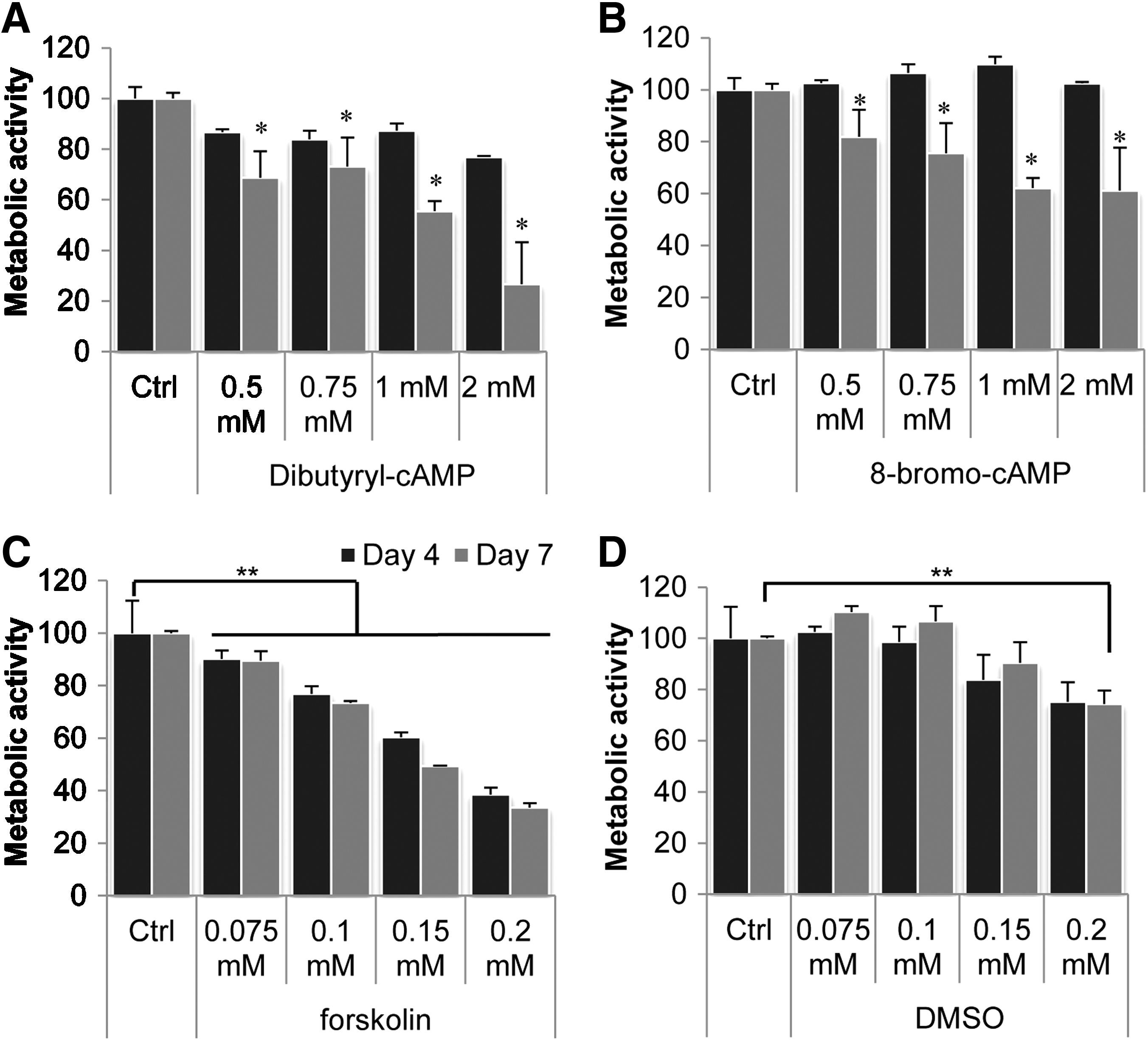
Protein kinase A (PKA) activators inhibit proliferation of human mesenchymal stem cells in a dose-dependent manner. Cells were treated with dibutyryl cyclic adenosine monophosphate (db-cAMP) or 8-bromo-cAMP (8-br-cAMP) in concentrations ranging from 0.5 to 2.0mM or with forskolin in concentrations ranging from 0.075 to 0.2mM. A concentration of 0.5mM db-cAMP did not affect proliferation. Other concentrations significantly decreased proliferation after 7 days, but after 4 days, only a slight decrease in cell number was observed
8-br-cAMP does not affect ALP expression but enhances expression of osteogenic marker genes
Next, we investigated whether osteogenic differentiation was dependent on the type of activator or the concentration by measuring expression of the early osteogenic marker ALP. Concentrations used were the same as for proliferation experiments, and each compound was tested on hMSCs from at least three donors. Representative results from one donor are depicted in Figure 3. Treatment with 0.1mM db-cAMP did not enhance ALP expression in any of the donors tested, consistent with cell morphology observations (data not shown). Concentrations of 0.5 to 2.0mM db-cAMP resulted in two to three times greater ALP expression, comparable with dexamethasone-induced ALP expression in that specific donor, without differences between these concentrations (Fig. 3A). In contrast, 8-br-cAMP did not significantly affect ALP expression in any of the donors tested (Fig. 3B). Treatment with 0.05 to 0.20mM forskolin resulted in a dose-dependent increase in ALP expression, with 0.10mM resulting in highest expression levels (Fig. 3C). Treatment with 0.01mM forskolin or DMSO alone did not affect ALP expression. We then selected concentrations of 1.0mM db- and 8-br-cAMP and 0.10mM forskolin to compare the three compounds (Fig. 3D). In hMSCs from every donor tested, db-cAMP induced the highest ALP expression. (For additional data on other donors, see Supplementary data S2.) Because we previously observed synergistic effects of db-cAMP and dexamethasone on ALP expression levels, we also examined the combinatorial effects of 8-br-cAMP and forskolin with dexamethasone. As shown in Figure 3E, although 8-br-cAMP alone did not affect ALP levels, upon addition to dexamethasone, there was a small but nonsignificant increase in ALP levels over dexamethasone alone. For forskolin, there was no significant increase in ALP levels upon addition of dexamethasone. In this donor, forskolin induced ALP to a significantly higher level than dexamethasone, indicating that forskolin might have potential for hMSCs that do not respond to dexamethasone.

Dibutyryl cyclic adenosine monophosphate (db-cAMP) and forskolin enhance alkaline phosphatase (ALP) expression of human mesenchymal stem cells. Cells were cultured in the presence of protein kinase A (PKA) activators for 3 days, after which ALP expression was measured. A concentration of 0.1mM db-cAMP did not enhance ALP expression, but concentrations ranging from 0.5 to 2.0mM increased ALP expression to the same extent as dexamethasone
Similar to the proliferation results described above, ALP expression levels were donor dependent, but no correlation was found between the inhibitory effect on proliferation and the greater ALP expression. Because 1mM db-cAMP was previously shown to increase gene expression of the BMP-2 target genes ID1 and ID2, 20 we also examined gene expression levels of these genes after treatment with the three compounds in addition to the osteogenic genes BMP-2 and IGF-1. As shown in Figure 4, all three compounds increased expression of ID1, ID2, BMP-2, and IGF-1, generally in a dose-dependent manner. Treatment with the optimum concentration of db-cAMP (1mM) resulted in significantly higher expression of BMP-2 and IGF-1 than with 8-br-cAMP and forskolin. In contrast, expression of ID1 was higher in forskolin-treated cells, and no differences were found in ID2 expression between the optimal concentrations of these two compounds. 8-br-cAMP did not induce expression of ID1 or ID2. Because it has been found that forskolin and 8-br-cAMP reduce expression of the osteogenic transcription factor osterix in UMR-106-01 cells, 29 the expression of this gene was also examined. In contrast to this study, Figure 4E shows that all three compounds induce high expression of this gene and that 8-br-cAMP and forskolin induce significantly higher levels than db-cAMP.

All three protein kinase A (PKA) activators increased expression of osteogenic genes in a dose-dependent manner. Human mesenchymal stem cells were treated with dibutyryl cyclic adenosine monophosphate (db-cAMP), 8-bromo-cAMP (8-br-cAMP), or forskolin for 5 days, after which gene expression was determined. All three activators increased expression of insulin-like growth factor (IGF)-1
db-cAMP and forskolin induce IGF-1 secretion to a higher extent than 8-br-cAMP
We previously demonstrated that PKA activation enhances secretion of bone-specific growth factors such as IGF-1, interleukin-8 (IL-8), and IL-11. 20 Because quantitative PCR experiments showed higher IGF-1 expression after treatment with db-cAMP, we investigated whether the secretion of IGF-1, one of the most abundant growth factors in bone, was dependent on the type of PKA activator and the concentration. Figure 5A shows that all three PKA activators increased IGF-1 secretion, but db-cAMP and forskolin did so to significantly higher concentrations than 8-br-cAMP. No differences were found between different concentrations of these compounds, although when IGF-1 concentrations were normalized to cell numbers, a clear concentration-dependent increase was observed, as shown in Figure 5B. Of the concentrations tested here, 2.0mM db-cAMP and 0.2mM forskolin resulted in highest IGF-1 secretion per cell, but because of the severe inhibition of proliferation at these concentrations, the total amount of secreted IGF-1 was highest after treatment with 0.75 to 1.0mM db-cAMP and 0.10mM forskolin. These optimum concentrations were then used to examine IGF-1 secretion in two more donors (Supplementary data S3), and in these donors, an increase in IGF-1 secretion, also by 8-br-cAMP, was found, although again, db-cAMP and forskolin had more profound effects.
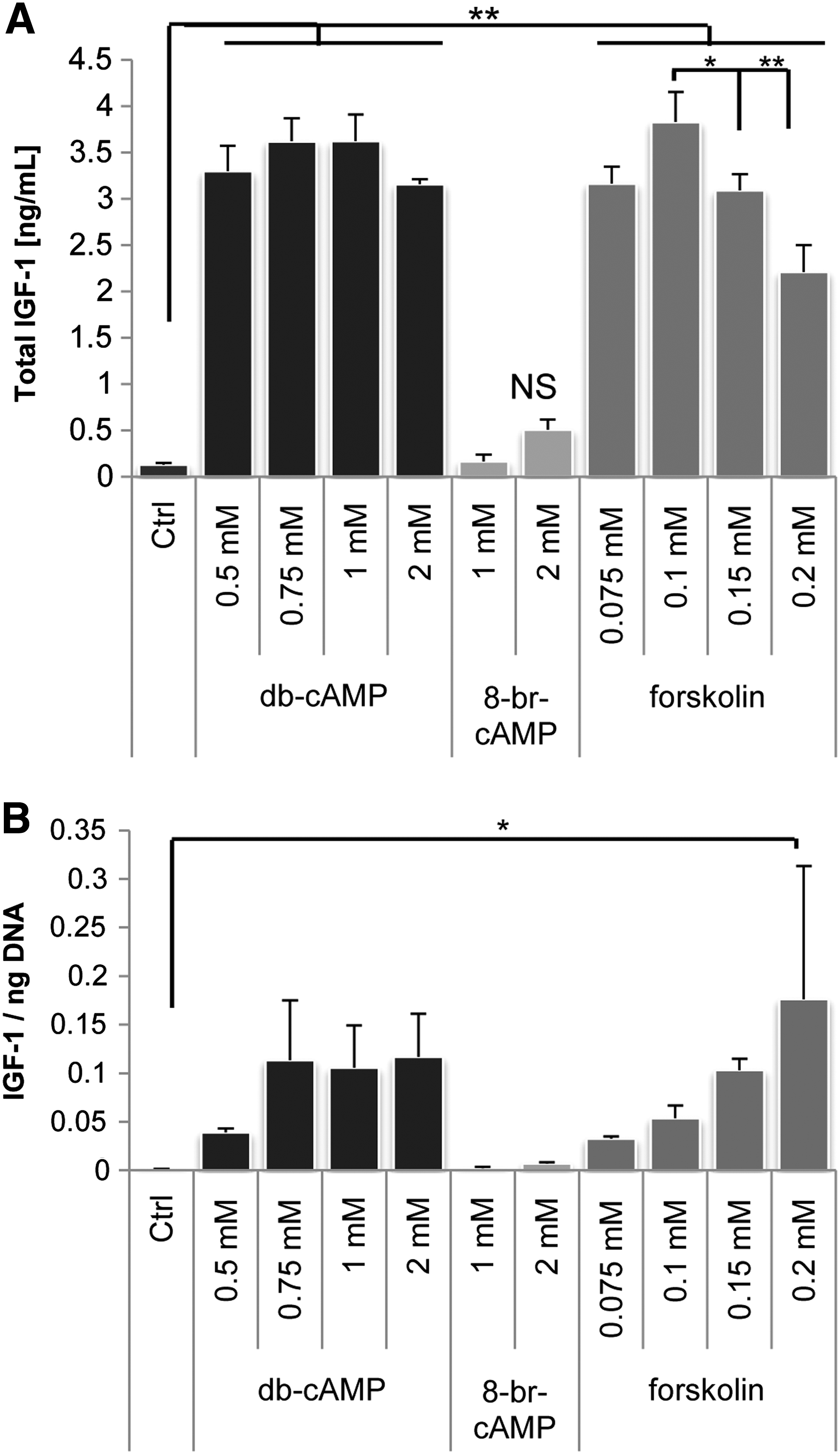
Dibutyryl cyclic adenosine monophosphate (db-cAMP) and forskolin enhanced insulin-like growth factor (IGF)-1 secretion. Human mesenchymal stem cells were cultured in the presence of PKA activators for 5 days, after which the concentration of IGF-1 in the medium was measured using enzyme-linked immunosorbent assay. No significant differences in IGF-1 secretion between various concentrations of db-cAMP and forskolin were detected, whereas 8-br-cAMP treatment resulted in significantly lower amounts of IGF-1
Preconditioning with forskolin enhances in vivo bone formation of hMSCs
As described, differentiation of hMSCs in vitro does not necessarily result in enhanced bone formation in vivo. Because 8-br-cAMP treatment resulted in adipogenic differentiation in long-term mineralization cultures (data not shown), only db-cAMP and forskolin were used to pretreat hMSCs before implantation in an ectopic location in nude mice. Figure 6 shows the amount of bone per scaffold area (A) and histological sections (B) after 6 weeks of implantation. Pretreatment with 0.10 or 0.15mM forskolin or 1mM db-cAMP resulted in greater bone formation than in untreated hMSCs, confirming previous data. Although not significant, treatment with 0.10mM forskolin resulted in approximately twice the amount of bone as with the higher concentration or with db-cAMP. The newly formed bone had a mature appearance.
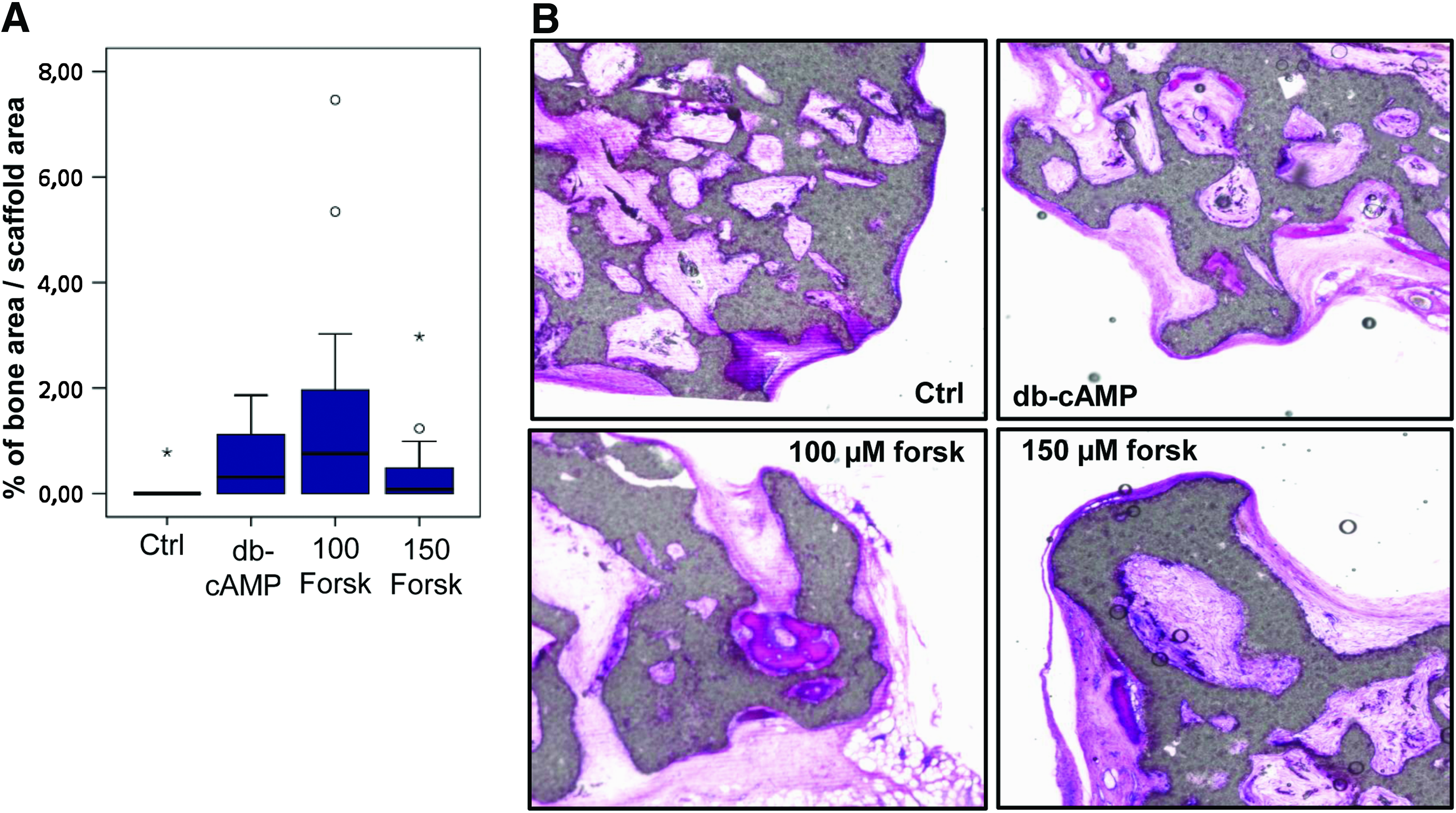
Forskolin enhances bone formation in vivo. Human mesenchymal stem cells were cultured on biphasic calcium phosphate scaffolds for 4 days in proliferation medium, followed by 3 days in basic medium in the presence of dibutyryl cyclic adenosine monophosphate (db-cAMP) and forskolin. Then, samples were implanted for 6 weeks, after which the amounts of bone were analyzed. Although not significant, treatment with 0.10mM forskolin resulted in greater amounts of bone than 1mM db-cAMP or 0.15mM forskolin
Discussion
We previously demonstrated that activation of PKA with the small molecule db-cAMP significantly enhanced bone formation of hMSCs in vivo, but the severe inhibition of cell proliferation by this compound led us to investigate whether osteogenic differentiation and bone formation as a result of PKA activation can be improved further by reducing concentrations or using other PKA activators. In line with our previous findings, we here confirm greater bone formation upon pretreatment of hMSCs with the PKA activator db-cAMP, and in addition, we show enhanced bone formation after treatment of hMSCs with forskolin, a more-upstream activator of the PKA pathway. As demonstrated before with dexamethasone,19,30 we found a donor-dependent effect with both compounds. Although ALP levels and expression of IGF-1, BMP-2, and BMP-2 target genes were consistently enhanced in the donors tested, basal levels and relative increases varied between donors. The inhibitory effect on proliferation also varied between donors, but we did not find a link between the induced ALP expression or gene expression and the decrease in proliferation. A decrease in proliferation will ultimately result in smaller numbers of implanted cells, and it has been demonstrated that implantation of larger cell numbers results in more bone formation, although maximums have been observed. For human periosteum–derived cells, it was shown that implants seeded with fewer than 1.106 cells did not form any bone, whereas implants with 1 to 5.106 cells showed similar amounts of bone. 31 For hMSCs, maximum bone was observed at a cell number of 5.105 cells per implant, 32 suggesting that, above this threshold, the effect on proliferation becomes irrelevant and that differentiation and release of bone-specific growth factors may be more important parameters. In our in vivo assay, we initially seeded 200,000 cells/implant, well below this threshold. Although cell numbers were not quantified before implantation, a proliferation phase of 4 more days with 3 additional days of differentiation before implantation most certainly resulted in implantation of higher cell numbers. We found larger amounts of bone after pretreatment of the cells with 0.10mM forskolin than with 0.15mM forskolin, probably because of the severe inhibition of proliferation after treatment with 0.15mM forskolin. In addition, 0.10mM forskolin resulted in larger amounts of bone than 1.0mM db-cAMP, although in vitro data, which showed higher expression of IGF-1 and BMP-2 after treatment with db-cAMP, did not reflect this. Forskolin highly induced expression of osterix, which demonstrates the need for a proper in vitro marker for in vivo bone formation. It is possible that the target of forskolin, adenylyl cyclase, being more upstream, eventually also results in activation of pathways other than via cAMP,33,34 but this was not investigated here. Because we observed a concentration-dependent effect in vitro and in vivo, optimal concentrations may even be specific for each donor, depending on the compounds' effect on proliferation. The effect of forskolin could potentially be further enhanced by using another solvent that would have less-detrimental effects on proliferation. Ethanol would probably interfere less with proliferation and is also compatible but was demonstrated to interfere with the activity of forskolin, whereas DMSO was shown to be least interfering and was therefore also used in this study. 35 Forskolin has been tested before in clinical settings for the treatment of dilated cardiomyopathy36,37 and is known as a medicinal herb, and its use thus has been shown to be safe. Translation to the clinic would thus be easier than with other PKA activators. In addition to differentiation of hMSCs, cAMP/PKA activation is involved in a wide range of cellular processes, and in vivo application of these compounds could potentially induce side effects. An overview of various biological applications for which these specific PKA activators have been used is given in Table 2.
hMSCs, human mesenchymal stem cells; PLGA, poly(lactic-co-glycolic acid; cAMP, cyclic adenosine monophosphate.
Differences in downstream activation mechanisms could possibly explain the distinct effects of db-cAMP and 8-br-cAMP. Recently we demonstrated that intermittent stimulation of PKA with db-cAMP results in a decrease in ALP expression, which suggests a dual role for PKA in the differentiation of hMSCs, depending on the duration of the signal. 38 Differences in the stability of db-cAMP and 8-br-cAMP could result in variations of downstream activation regimes and ultimately in alternative differentiation routes. For example, whereas for some time PKA was thought to be the sole mediator of cAMP signaling, it is now known that cAMP can also activate exchange protein directly activated by cAMP, a factor demonstrated to be indispensable for adipogenic differentiation.39,40 In conclusion, we introduce forskolin as a novel agent to induce osteogenic differentiation of hMSCs in vitro and to enhance bone formation in vivo, and we suggest a role for the cAMP/PKA pathway, and possibly other downstream mediators, in controlling the balance between adipogenic and osteogenic differentiation.
Footnotes
Acknowledgments
We would like to thank Hugo Andre Alves and Anandkumar Nandakumar for their technical support and Dr. Auke Renard (Medisch Spectrum Twente, Enschede, The Netherlands) for kindly providing us with bone marrow aspirates. We also gratefully acknowledge the support of the Smart Mix Program of the Netherlands Ministry of Economic Affairs and the Netherlands Ministry of Education, Culture and Science and STW for a VIDI grant to J.d.B.
All animal experiments were approved by the local Animal Experimental Committee.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.