
Abstract
Epithelial–mesenchymal interactions during embryogenesis are critical in defining the phenotype of tissues and organs. The initial elongation of the mammary bud represents a central morphological event requiring extensive epithelial–mesenchymal crosstalk. The precise mechanism orchestrating this outgrowth is still unknown and mostly animal models have been relied upon to explore this process. Highly tunable three-dimensional (3D) culture models are a complementary approach to address the question of phenotypic determination. Here, we used a 3D in vitro culture to study the roles of stromal cells and extracellular matrix components during mammary tubulogenesis. Fibroblasts, adipocytes, and type I collagen actively participated in this process, whereas reconstituted basement membrane inhibited tubulogenesis by affecting collagen organization. We conclude that biochemical and biomechanical signals mediate the interaction between cells and matrix components and are necessary to induce tubulogenesis in vitro.
Introduction
Tubulogenesis, the initial elongation of the mammary bud, represents a central event in mammogenesis, requiring reciprocal epithelial–mesenchymal interactions. Other branching organs such as teeth, lungs, kidneys, and salivary glands undergo similar processes. Most genes known to be required for the development of the embryonic mammary gland have been identified using knockout mouse models and genetic tools. The disruption or complete absence of certain proteins led to the arrest of mammogenesis at specific stages.9–13 Still, the underlying mechanisms that coordinate the initial steps of mammary gland morphogenesis remain largely unknown.
When searching for answers to those unknowns, animal models provide only limited assistance in identifying the roles played by the different cells and tissue components affecting ductal morphogenesis. A complementary approach to address the issue of tissue patterning, particularly when assessing the biophysical properties of tissue components, benefits from the use of 3-dimensional (3D) tissue culture models.14,15
We have previously developed 3D in vitro tissue culture models in which stromal–epithelial interactions can be manipulated to form mammary structures closely resembling those found in vivo.16,17 The composition of the extracellular matrix (ECM) and the presence of fibroblasts influenced the epithelial phenotype. For example, mammary ducts formed in the absence of human breast fibroblasts when MCF10A breast epithelial cells grew in type I collagen gels. In contrast, fibroblasts were necessary for the formation of ducts in a matrix containing a mixture of reconstituted basement membrane (rBM) and type I collagen. 17 Furthermore the ECM is an important player in tubulogenesis as demonstrated in our earlier work by the inhibition of ECM in the presence of rBM. 17 Also, epithelial cells and fibroblasts modified the organization of collagen fibers, and tubulogenesis was associated with the development of thick collagen bundles located along the direction of ductal elongation. 16
Herein, we explore further the role of ECM composition and the contribution of two stromal cell types, namely fibroblasts and adipocytes, to tubulogenesis using nontumorigenic human breast MCF10A cells. We also assessed the role of ECM composition in defining the epithelial phenotype in the absence of stromal cells, the role of fibroblasts and adipocytes in defining the epithelial phenotype in the presence of various ECM, and whether paracrine signals or physical contact between epithelial and stromal cells are needed for ductal formation.
Materials and Methods
Chemicals and cell culture reagents
Mayer's hematoxylin, 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, hydrocortisone, cholera toxin, insulin, and carmine were purchased from Sigma-Aldrich (St. Louis, MO). Dulbecco's modified Eagle medium (DMEM) was purchased from MP Biomedicals (Solon, OH). DMEM/F12 and penicillin–streptomycin solution were obtained from Gibco/Invitrogen (Carlsbad, CA). Equine serum and fetal bovine serum were purchased from HyClone (Logan, UT). Bovine type I collagen was purchased from Organogenesis (Canton, MA). Epidermal growth factor, rBM (Matrigel), and rat tail collagen were purchased from BD Biosciences (San Jose, CA). Recombinant human hepatocyte growth factor (rhHGF) was obtained from R&D Systems (Minneapolis, MN). Formalin was obtained from Fisher Scientific (Atlanta, GA). Human preadipocytes (PA) and proliferation and differentiation media were purchased from ZenBio (Research Triangle Park, NC).
Cell maintenance
Primary human fibroblasts obtained from reduction mammoplasties (RMF) were purchased from ScienCell Research Laboratories (San Diego, CA). RMF and the nontumorigenic human breast epithelial MCF10A cells were maintained and expanded in Corning cell culture plastic flasks (Corning, NY). MCF10A cells were grown in DMEM/F12 containing 5% equine serum, 20 ng/mL epidermal growth factor, 0.5 μg/mL hydrocortisone, 0.1 μg/mL cholera toxin, 10 μg/mL insulin and 1% penicillin–streptomycin solution. RMF were grown in DMEM containing 10% fetal bovine serum. Human primary PA were used immediately after thawing. All cultures were maintained at 37°C and 6% carbon dioxide. For co-culture experiments, a combined medium containing equal volumes of each of the individual cell media was used. The combined media were tested containing each cell type alone to assure proper growth and behavior of cells (data not shown).
ECM preparation
Type I collagen gels (collagen gels) and mixed rBM–collagen gels (mixed gels) were prepared as previously described. 17 For the tubulogenesis assay, the mixed gels were modified to include a broad spectrum of collagen to rBM ratios. These gels were prepared using 50%, 40%, 30%, 20%, 10%, and 5% volume ratios of rBM while maintaining the final collagen concentration at 1 mg/mL.
3D cell culture
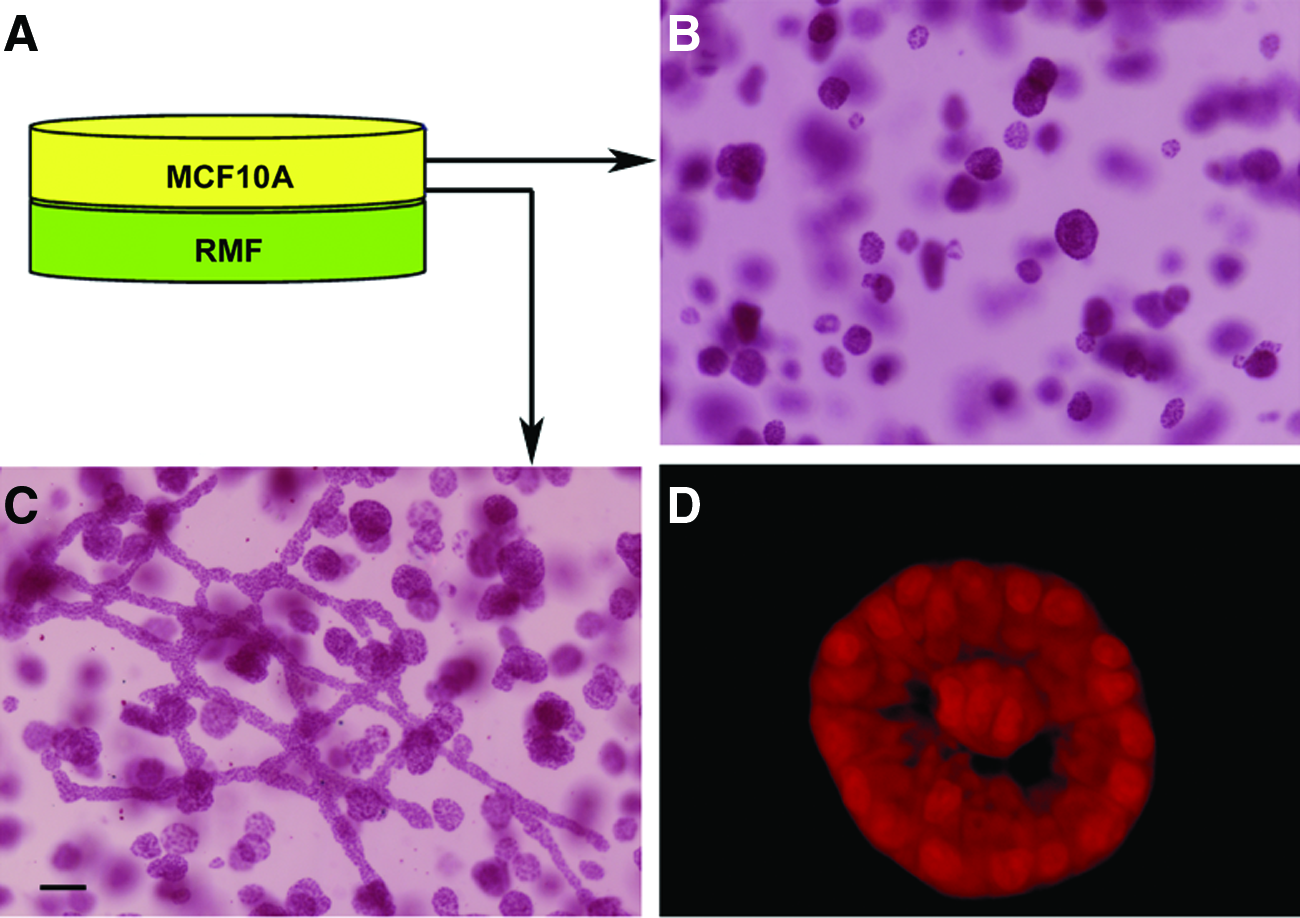
The 3D cultures were prepared as previously described 17 with modifications as indicated in Fig. 1. To obtain differentiated adipocytes from PA, 300,000 freshly thawed PA were embedded in collagen or mixed gels and allowed to proliferate for 2 weeks in proliferation medium. After 2 weeks in culture, the medium was changed to differentiation medium containing the necessary supplements, including insulin, dexamethasone, and Peroxisome proliferator-activated receptor gamma agonist, to induce adipocyte differentiation. Adipocytes were allowed to differentiate for 4 weeks, after which the gels were harvested, or a new gel was cast on top of the adipocyte-containing gel. These will be referred to as “sandwich” gels (Fig. 1); the second gel contained MCF10A cells or MCF10A cells and RMF in co-culture.

Experimental gel formats. Schematic depicts the following gel formats used in these studies: the basic three-dimensional gel (left), sandwich gel comprising two gels cast on top of each other with or without a porous membrane separating them (middle), and drop gels in which a drop of collagen–reconstituted basement membrane matrix is suspended within a collagen gel before solidification (right). Color images available online at www.liebertonline.com/tea
In co-cultures, cells were seeded as 300,000 MCF10A cells and 100,000 RMF or 300,000 MCF10A cells and 300,000 PA. The same cell number was seeded for each cell type cultured independently. Unless otherwise specified, the same cell numbers were used in all experimental conditions. The co-cultures were maintained for 1 to 3 weeks depending on the experimental design. The medium was changed every 2 to 3 days.
Conditions necessary for tubulogenesis
RMF-conditioned medium
MCF10A cells and RMF embedded in mixed gels containing 50% rBM and collagen (1 mg/mL) were seeded concomitantly into separate inserts. Every 2 to 3 days, the MCF10A cells were fed with 75% RMF-conditioned medium and 25% fresh medium. The RMF-conditioned medium was used immediately after collection from the RMF-containing gels and was never frozen. The cells were cultured for up to 3 weeks.
Interface of collagen and rBM
Before the acellular collagen gel solidified, a drop of mixed matrix containing MCF10A cells was pipetted into the center of the gel, forming a drop gel (Fig. 1).
Physical separation between MCF10A and RMF
Mixed gels containing RMF or MCF10A cells were seeded simultaneously. Upon solidification, the gel containing the MCF10A cells was carefully removed from its insert and placed on top of the solidified RMF-containing gel. These sandwich gels were incubated for up to 3 weeks, and the medium was changed every 2 to 3 days.
Assessing the role of HGF in tubulogenesis
MCF10A cells were cultured in mixed gels containing 10 or 20 ng/mL recombinant human HGF. The culture medium was also supplemented with rhHGF; medium was changed every 2 or 3 days. The gels were harvested after 1 or 2 weeks in culture.
Gel processing and staining
At the end of each experiment, the gels were cut into pieces. One piece was fixed overnight in 10% phosphate-buffered formalin and paraffin embedded. Another piece was whole-mounted onto a slide and fixed overnight in 10% phosphate-buffered formalin for 3D morphological analysis. The whole-mounted gels were stained with carmine alum overnight following a protocol described previously 18 and mounted with Permount (Fisher Scientific, Atlanta, GA). Some paraffin sections were stained with hematoxylin and eosin for histological analysis, and picrosirius red was used to assess collagen fiber organization as previously described. 16
Immunohistochemical analysis
All primary antibody concentrations and supplier information are listed in Table 1. An antigen-retrieval method using microwave pretreatment and 0.01M sodium citrate buffer (pH 6) was used for all antibodies except laminin 5, for which 0.05% proteinase XXIV treatment was used instead, 19 and the analyses were performed as previously described. 17 Paraffin-embedded sections were also stained using picrosirius red solution (0.1% Sirius red diluted in saturated picric acid) as described previously. 16 Images were captured using an AxioCam HR color digital camera attached to a Zeiss Axioscope 2 plus microscope (Carl Zeiss MicroImaging Inc, Thornwood, NY). The picrosirius red stained sections were analyzed using the same microscope equipped with a polarizer and an analyzer.
Leptin quantification
Leptin, a marker of adipocyte differentiation, was assessed using an enzyme-linked immunosorbent assay for human leptin (R&D Systems) following the manufacturer's instructions. An aliquot of medium was collected from the PA cultures after 2 weeks and just before changing the proliferation medium to the differentiation medium. After that, the medium was collected once a week for 4 weeks. All the aliquots were kept frozen until the assay was performed. Media were collected from experiments that included cells grown in collagen gels and mixed gels.
Confocal microscopy
Whole-mounted gels were analyzed using a Zeiss LSM 510 microscope (Carl Zeiss MicroImaging Inc.), as previously described. 17
Morphometric analysis
Three experiments for each condition were analyzed, and for each experiment, three arbitrarily chosen fields at ×10 magnification per section were examined. Images were captured using a Zeiss Axioscope 2 plus at ×10 and 3900 dpi and analyzed using Zeiss Axiovision version 4.5 software. The following parameters were measured per section: total number of alveolar structures versus extending and tubular structures, total number of cells per structure, and number of Ki67-positive cells within those structures. Proliferating epithelial cells were expressed as %Ki67-positive cells per total epithelial cell number. Extending (defined as not yet duct-like but rather elongating structures) and tubular structures were counted as one regardless of the numbers of branches per tubular structure.
Statistics
SPSS software package 15.0 (SPSS, Inc., Chicago, IL) was used for all statistical analyses. An ANOVA with a Bonferroni post hoc test was performed to assess the differences in the percentage of the cells positive for Ki67 as a proliferation index. Independent-sample t-tests were used to compare morphological parameters. For all statistical tests, results were considered significant at p<0.05. All results are presented as means and standard errors of the mean.
Results
rBM prevents tubulogenesis in a concentration-dependent manner
We have previously shown that MCF10A cells grown in a collagen matrix (1 mg/mL) were able to form ducts while the same cells exclusively formed alveolar structures in mixed gels containing the same concentration of collagen and 50% rBM. 17 To investigate the role of the ECM composition in tubulogenesis, MCF10A cells were grown in matrices containing decreasing volumes of rBM while maintaining the final collagen concentration at 1 mg/mL (tubulogenesis assay).
Ducts were first observed in 20% rBM gels after 1 week in culture, and the number of elongated alveoli increased at week 2 (Fig. 2A). The number of ducts increased steadily with decreasing amounts of rBM, although the number of alveoli was significantly higher than that of tubular and extending structures in gels containing 20% or more rBM (Fig. 2B) (p=0.003, p=0.04, p=0.006, p=0.02, for 50%, 40%, 30%, and 20% rBM, respectively). Ducts were most abundant in gels containing as little as 10% or 5% rBM after 2 weeks in culture (Fig. 2A, B). The 10% rBM gels had a similar number of ducts and alveoli, but the number of ducts was significantly higher than the number of alveoli in 5% rBM gels (p=0.01) (Fig. 2B). The pattern of epithelial organization observed in 5% rBM matrix was similar to that observed in 1 mg/mL collagen matrix alone (0% rBM). 17 The almost complete switch of the epithelial phenotype from only alveolar in the presence of 50% rBM to mostly ductal in 5% rBM strongly suggests that epithelial cells grown alone in a matrix containing large quantities of rBM were inhibited from organizing as ducts and that the inhibitory effect disappeared when the amount of rBM was reduced.
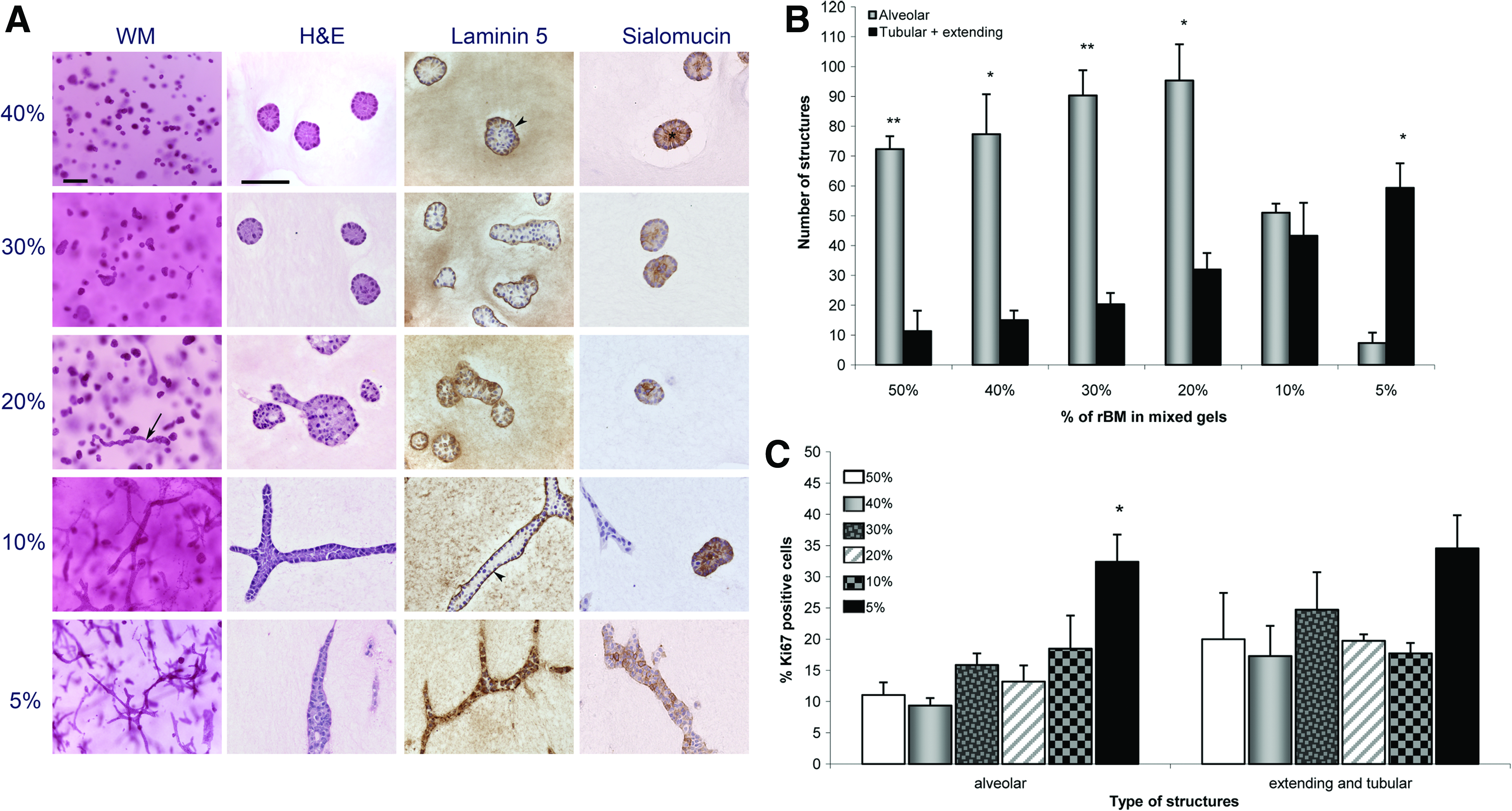
Epithelial structures obtained in mixed reconstituted basement membrane (rBM) –collagen gels containing MCF10A cells alone. Decreasing amounts of rBM (Matrigel) were used while the collagen concentration was maintained at 1 mg/mL in all cases.
We assessed the proliferation index by measuring the percentage of epithelial cells expressing Ki67 protein. After 2 weeks in culture, the percentage of Ki67 positive cells in the tubular structures was 34.6% in the 5% rBM gels compared to 20.0% in the 50% rBM gels (Fig. 2C). Similarly, after 2 weeks in culture, the proliferation index in alveolar structures also increased with decreasing content of rBM, and the percentage of Ki67 expressing cells was significantly higher in the 5% rBM (32.4%) than in the 50% rBM gels (11.0%) (p=0.008) (Fig. 2C).These results indicate that rBM may also have an inhibitory effect on epithelial cell proliferation regardless of the phenotype.
After 1 and 2 weeks in culture, basement membrane formation, characterized by the presence of laminin 5 staining, was observed in alveoli and ducts in the majority of the matrices (Fig. 2A), although in 5% rBM gels, the staining became diffuse and discontinuous. This result correlates with our previous observation that MCF10A cells cultured alone in collagen gels were unable to form a basement membrane. 17 In addition, after 2 weeks, epithelial structures formed in most matrices were polarized, as demonstrated by the apically secreted protein sialomucin, although the staining pattern varied according to the matrix composition (Fig. 2A). A typical pattern of polarity with apical expression of sialomucin was observed in gels containing high concentrations of rBM, whereas the sialomucin staining became pericellular in gels containing 10% and 5% rBM, indicating a loss of cell polarity (Fig. 2A). These findings are consistent with our previous results. 17
Collagen fiber organization plays an important role in tubulogenesis. 16 We used picrosirius red staining to examine whether the addition of rBM to rat tail collagen affects the fibers' organization. This method is based on the birefringence properties of collagen when packed into parallel fibers and dense bundles; they appear as bright structures varying in color from red to orange and yellow. When the collagen fibers are not packed in an orderly fashion or are not dense, the intensity of the birefringence diminishes. 20 Figure 3 shows that, after 2 weeks in culture, the presence of rBM significantly influenced collagen fiber organization. The collagen fibers formed an organized pattern of long, thick bundles when low content of rBM was present, as demonstrated by the greater birefringence, manifested as a bright yellow color (Fig. 3). On the contrary, in the presence of high content of rBM, fibers organized in shorter, thinner bundles that formed a random pattern. The collagen fibers appeared to be organized in two different patterns in the presence of elongating alveoli: the birefringence diminished immediately around the epithelial cells, which appeared with a darker halo, and the birefringence increased in areas near the points of elongation, where the staining clearly showed bundles of fibers organized along the direction of elongation (Fig. 3, lower panels).

Picrosirius red staining and polarized light microscopy showing collagen fiber organization in gels containing MCF10A cells alone with decreasing volumes of reconstituted basement membrane (indicated in %). Scale bar: 50μm. Time point: 2 weeks. *Epithelial structures. Color images available online at www.liebertonline.com/tea
Type I collagen is necessary for tubulogenesis
To further understand the roles of collagen and rBM in tubulogenesis, MCF10A cells were suspended in 100% rBM or 50% rBM–collagen matrix. A drop of the matrix containing the cells was placed within a 1 mg/mL collagen matrix just before the solidification of the gels (drop gel). The two matrices did not mix and were easily identifiable throughout the 2 weeks in culture (Fig. 4A). Only MCF10A cells located at the interface between the two matrices formed ducts that invaded the collagen gel (Fig. 4B,C). These ducts were well organized, contained a lumen and polarized cells (Fig. 4C), and were surrounded by a basement membrane (Fig. 4D). In addition, the epithelial cells within the drop of rBM-based matrix formed alveolar structures exclusively (Fig. 4C, inset), consistent with our previous findings. 17 These results indicate that collagen is necessary to induce tubulogenesis and that rBM inhibits this effect.
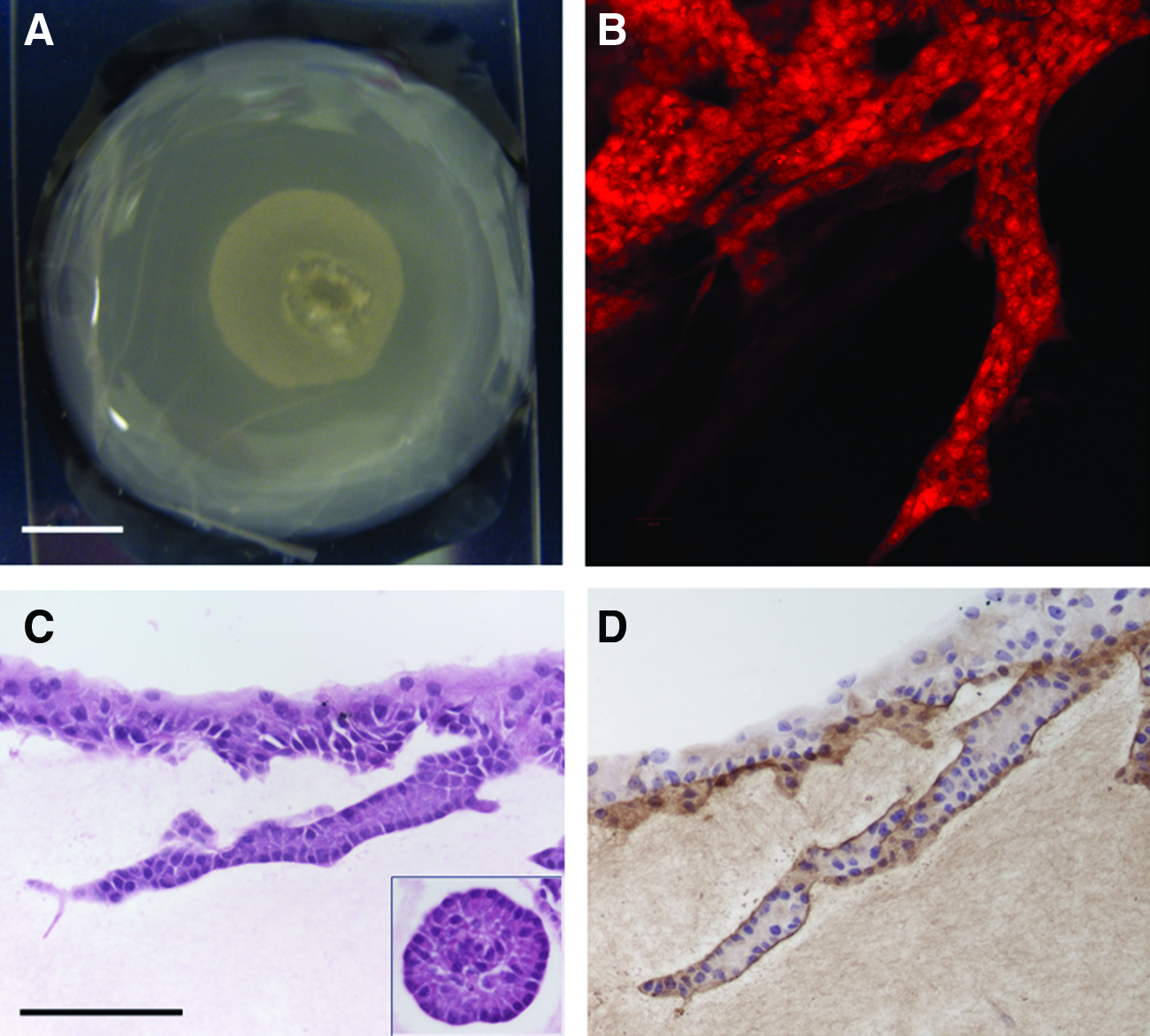
Presence of collagen was necessary for ductal formation.
Stromal cells initiate ductal formation in matrices containing high levels of rBM-breast fibroblasts
We have previously observed that MCF10A cells exclusively developed alveolar structures when grown in mixed gels containing 50% rBM and collagen (1 mg/mL), whereas they also formed ductal structures when grown in co-culture with RMF in the same matrix. 17 To further investigate the role of fibroblasts in promoting ductal morphogenesis, we tested whether soluble factors secreted by RMF would be sufficient mediators and whether physical contact between RMF and MCF10A cells was necessary to induce tubulogenesis in a 50% rBM–type I collagen matrix (mixed gels).
RMF-conditioned medium was not sufficient to induce tubulogenesis
After 2 weeks in culture, MCF10A cells grown in mixed gels and fed with conditioned medium derived from RMF grown separately in the same matrix did not form ductal structures (data not shown). Instead, only alveolar structures were observed. MCF10A cells co-cultured with RMF were used as positive controls for tubulogenesis. The use of conditioned medium derived from RMF cultured in two dimensions produced the same results (data not shown).
RMF in close proximity to MCF10A cells promoted tubulogenesis
A mixed gel containing MCF10A cells was cast on top of a mixed gel containing RMF, forming a sandwich gel (Fig. 5A). These gels were separated by a porous membrane of 3 or 0.22μm pore size, and the cells were cultured for 2 weeks. Alveoli were observed in the top gel away from the RMF-containing gel (Fig. 5B, D). When a 3-μm porous membrane separated the gels, ducts formed in areas where the top gel abutted the RMF-containing gel (Fig. 5C). Some RMF were observed within the MCF10A cell–containing gels, indicating that the fibroblasts had migrated through the membrane to the top gel. Although ductal structures in the top gel in areas abutting the bottom gel were obtained as well using the smaller 0.22-μm pore size membrane, which effectively prevented fibroblast migration. These findings indicate that fibroblasts need to be near epithelial cells to initiate tubulogenesis but that direct physical contact is not necessary.
MCF10A cells and primary human fibroblasts obtained from reduction mammoplasties (RMF) need to be in close proximity for tubulogenesis to occur in a 50% reconstituted basement membrane (rBM)-based matrix.
HGF insufficient to induce tubulogenesis in a rBM-containing matrix
HGF is considered a short-range factor secreted by RMF, and it was previously identified as being involved in tubulogenesis.2,21–23 Thus, we tested whether HGF might induce tubulogenesis in our system. The addition of rhHGF at 10 and 20 ng/mL to MCF10A cells grown in mixed gels was insufficient to induce tubulogenesis after 2 weeks in culture (data not shown). After 1 and 2 weeks, only acini were observed in all conditions; this was similar to when MCF10A cells were grown alone. MCF10A cells co-cultured with RMF were used as positive controls for ductal formation (data not shown).
Breast adipose cells
Fetal mammary ductal morphogenesis is initiated after signs of adipocyte differentiation appear in the stroma, 24 although the role of adipocytes and their precursors have yet to be thoroughly explored during mammary development. Given the abundance of fat cells in the human breast connective tissue, we examined their effect on epithelial phenotypes in our 3D tissue morphogenesis model.
Functional adipocytes were generated in the 3D morphogenesis model
Primary culture human PA were spindle shaped and morphologically similar to fibroblasts. We assessed whether these cells were able to differentiate into functional adipocytes when grown in a 1 mg/mL collagen matrix or a mixed gel containing 50% rBM. First, the PA were allowed to proliferate for 2 weeks; after differentiation was induced, PA started to accumulate lipid droplets and consequently became larger and rounder. After 4 weeks in differentiation medium (a total of 6 weeks in culture considering the first 2 weeks in proliferation medium), unilocular adipocytes and cells containing few, large lipid droplets were observed (Fig. 6A). These adipocytes showed the typically displaced nucleus, whereas some formed clusters containing a few cells and secreted the basement membrane protein type IV collagen (Fig. 6A). The adipocytes expressed this phenotype in collagen (top panel) and mixed gels with 50% rBM (lower panel), although the adipocytes in mixed gels seemed to contain fewer and bigger lipid droplets than those grown in a collagen- only matrix. In addition, we assessed the presence of leptin in the culture medium throughout the differentiation period. Leptin is a protein produced and secreted only by differentiated adipose cells and not by PA.25,26 The amount of leptin was negligible during the PA proliferation period; the range of leptin concentration reached 12.7±7.0 pg/mL in collagen and was undetectable in mixed gels, but it increased over time during the 4-week differentiation period from 34.8±14.9 pg/mL to 390.0±203.4 pg/mL in collagen, and from 17.5±7.1 pg/mL to 191.6±84.8 pg/mL in mixed gels (Fig. 6B). Although adipocytes grown in collagen gels seemed to secrete greater amounts of leptin than the mixed gels, the differences were not statistically significant, probably because of the large variation in measurements.

Adipocytes close to MCF10A cells promoted tubulogenesis in mixed gels containing 50% rBM
The hormones and chemicals necessary to induce the accumulation of lipids in PA also induced lipid accumulation in MCF10A cells when co-cultured with differentiating PA (data not shown). To maintain the phenotype of MCF10A cells, adipocytes were differentiated in a mixed gel containing 50% rBM, and then a second mixed gel containing MCF10A cells was cast on top of the gel containing the already differentiated adipocytes, forming a sandwich gel.
Confocal microscopy demonstrated the presence of ducts in areas where the top gel containing MCF10A cells abutted the adipocyte-containing gel (Fig. 7A); these ducts were first observed after 1 week in culture. In addition, alveoli exclusively formed in areas of the gel located further from the adipocytes (Fig. 7B, upper panels). These results are similar to those described for RMF (Fig. 5). After 2 weeks in culture, the ductal and alveolar structures showed positive staining for laminin 5, indicating the presence of a basement membrane (Fig. 7B). Sialomucin staining also showed apical secretion toward the lumen in most epithelial structures, confirming polarization (data not shown).
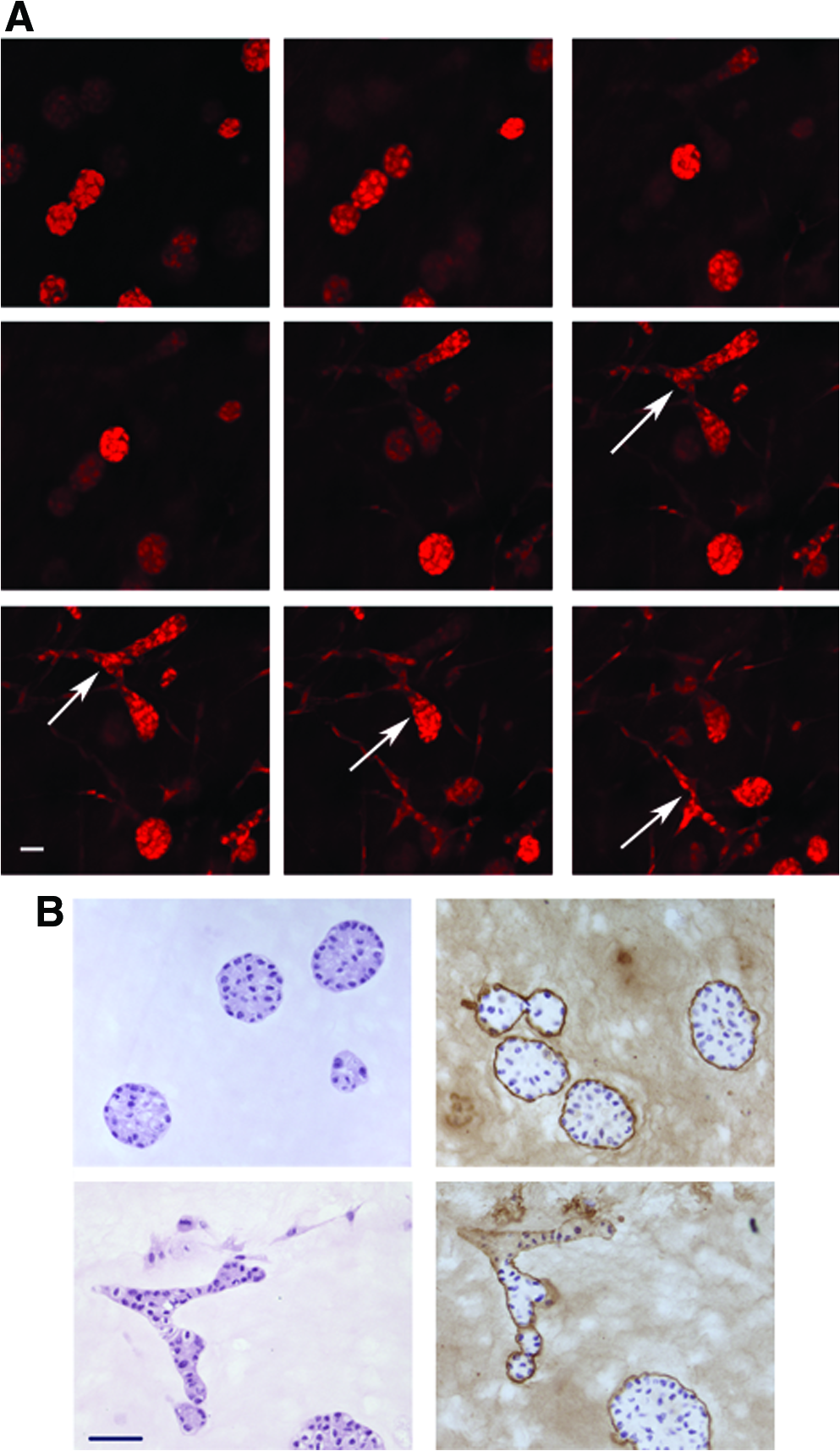
Proximity of adipocytes to MCF10A cells is necessary for tubulogenesis.
Adipocytes did not affect the epithelial phenotype in collagen gels
Contrary to what was observed in the mixed gels, the presence of adipocytes in the bottom gel did not affect the phenotype of MCF10A cells grown in collagen gels; these cells formed ductal and alveolar structures (Fig. 8A).
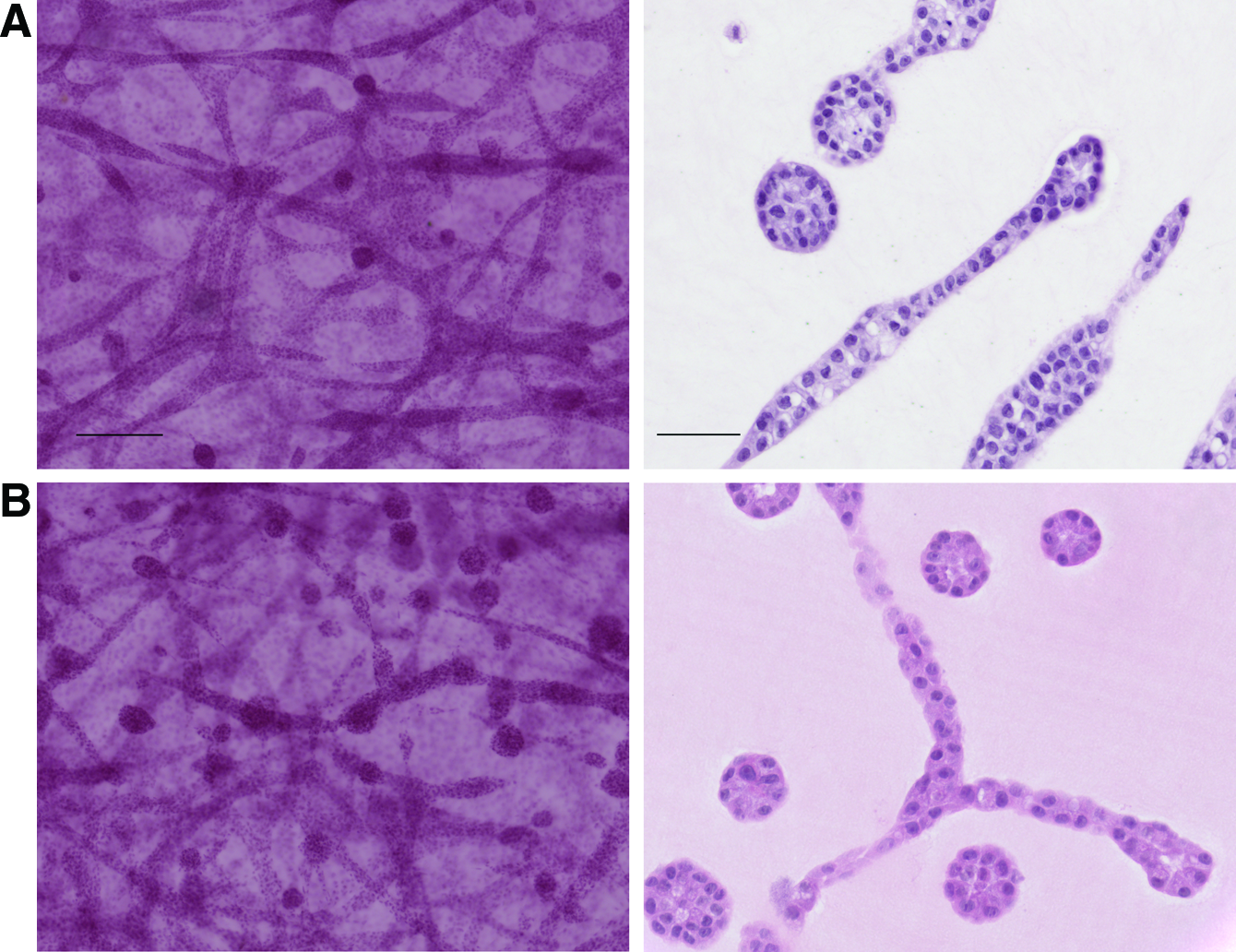
Human adipocytes did not affect the epithelial phenotype of
Increasing complexity: culturing MCF10A cells with RMF and adipocytes
Next, MCF10A cells were grown in the presence of the two most abundant breast stromal cells (fibroblasts and adipocytes) by casting a gel containing the epithelial cells and the RMF on top of a gel containing differentiated adipocytes. The cells were cultured for up to 3 weeks, and the experiments were performed using collagen and mixed gels. In all conditions, MCF10A formed branching ducts and alveoli (Fig. 8B), indicating that the two combined stromal cells did not significantly alter the epithelial phenotype. At the end of 3 weeks, the gels started to become crowded with cells, but no significant apoptosis or deterioration was observed in the structures.
In mixed gels, alveolar and ductal structures generated a basement membrane, as evidenced by positive laminin 5 staining (data not shown). After 2 weeks, a large number of these structures contained a lumen. Sialomucin was also expressed at the apical surface of cells in ductal and alveolar structures (data not shown).
In collagen gels, most epithelial cells became polarized by week 1. Although the epithelial structures did not generate a basement membrane; only a diffuse and discontinuous laminin 5 staining pattern was observed, even though they polarized, as shown by apical sialomucin secretion (data not shown). Adipocytes did not induce the formation of a basement membrane by MCF10A cells in collagen gels.
Preadipocyes induce tubulogenesis even in the presence of rBM-containing matrix
MCF10A cells were grown in co-cultures with PA in collagen and mixed gels with 50% rBM. Similar to results obtained with MCF10A cells co-cultured with RMF, alveoli and branching ducts were observed in both types of gels after weeks 1 and 2 in culture. Confocal microscopy confirmed that most epithelial structures also contained a lumen (Fig. 9A). In the mixed gels, the epithelial structures were polarized (Fig. 9B), and some showed positive staining for sialomucin and laminin 5 (Fig. 9C). Thus, PA were also able to induce tubulogenesis in 50% rBM–containing matrix. In collagen gels, the epithelial phenotype was similar to the one observed in the presence of RMF, namely, acini and ductal structures were observed (data not shown).
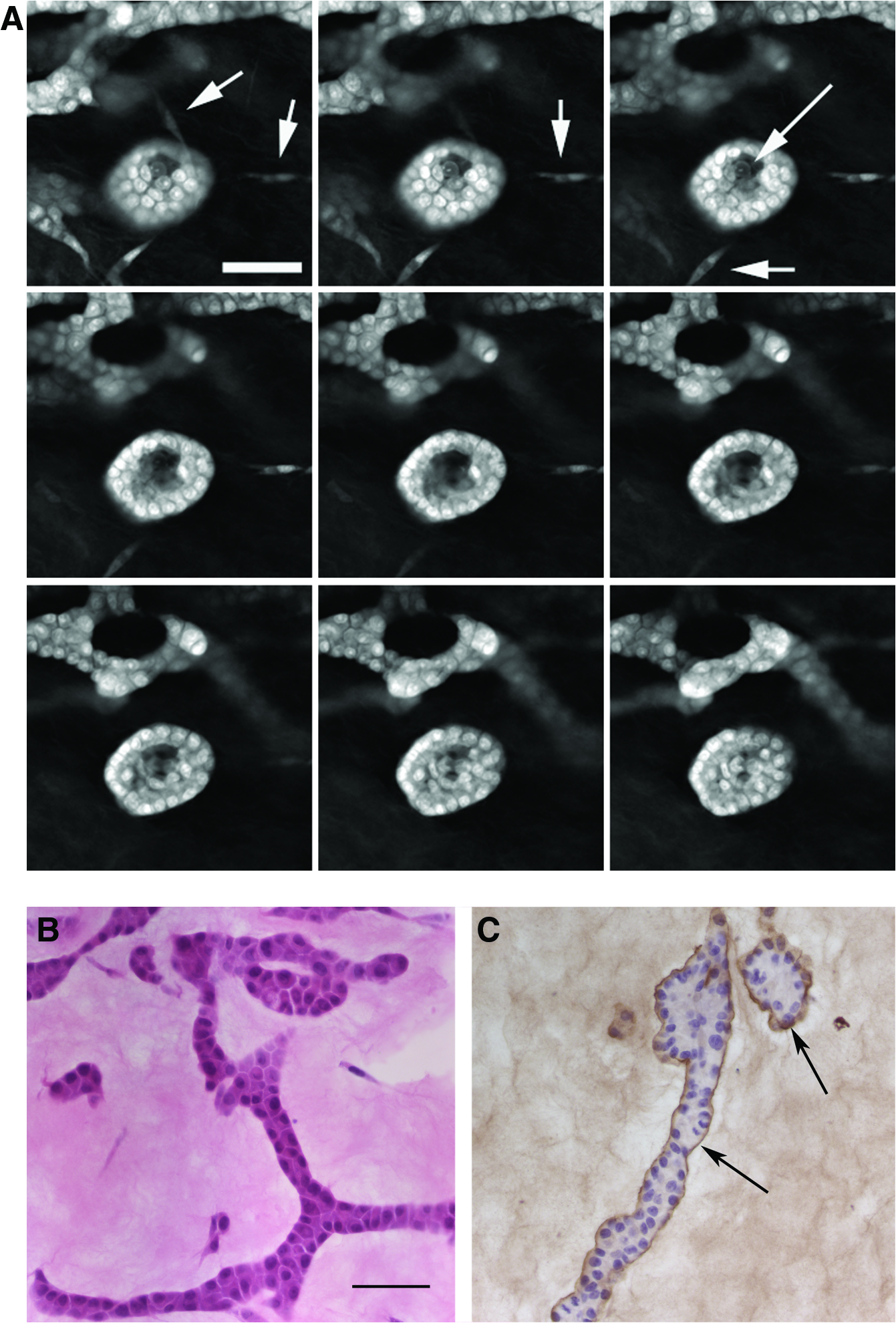
Preadipocytes allow polarization and basement membrane and lumen formation.
Discussion
One of the critical steps in embryonic breast development is the invagination of the mammary bud into the underlying mesenchyme and the formation of a sprout, a phenomenon called tubulogenesis. Little is known about the mechanism governing tubulogenesis, and studying this process in vivo has proven to be difficult. Understanding the contribution of cell–cell and cell–ECM interactions in breast morphogenesis requires careful analysis of the available surrogate models. We have explored the roles of the main breast cell types involved in the formation of ducts and alveoli and of the ECM that surrounds them using various 3D morphogenesis models.
We and others have previously shown that the composition of the ECM plays an important role in determining the phenotype of the epithelial structures that MCF10A cells form in 3D matrices.17,27,28 Based on our observation that MCF10A cells exclusively formed alveolar structures in mixed rBM–collagen gels, we hypothesized that rBM would inhibit ductal formation in our 3D model. By decreasing the content of rBM in mixed gels, we confirmed that tubulogenesis was initiated only when the amount of rBM was sufficiently low.
The inhibitory effect of rBM could be because of biochemical signaling by one or more of its components or rBM's mechanical properties. In support of the former hypothesis, Santos and colleagues reported that rBM was able to inhibit kidney branching morphogenesis and that type IV collagen and heparin sulfate were associated with the inhibitory effect observed in Mardin-Darby canine kidney epithelial cells. 29 MCF10A cells grown in a 3D matrix made of egg white developed a similar phenotype to the one observed in our rBM-containing matrix (alveolar structures containing a lumen and polarized cells, 28 although the major proteins in egg white are not present in rBM, which suggests that the biochemical composition of the matrix may not be the only factor in determining epithelial phenotype. Thus, the mechanical properties of matrices are likely to play an equally important role in preventing epithelial cells from initiating tubulogenesis. Alternatively, some proteins in egg white might also inhibit tubulogenesis.
We consistently observed that mixed gels appeared softer and contracted less than collagen gels. Decreasing rBM content progressively stiffened the matrix. The consistency of the gels containing only 5% rBM appeared to be similar to that of collagen gels; this experimental observation suggests that a potentially “stiffer” matrix may promote tubulogenesis. We are currently optimizing the compliance quantification of the various matrices used in the studies, which is a challenging task because of the gel's soft nature. Another interesting observation was that cell proliferation also progressively increased as the amount of rBM present in the matrix decreased. These results support the notion that changes in the biochemical composition and mechanical properties of gels can profoundly affect cell behavior.14,30–33
Our data demonstrated that type I collagen is necessary to initiate tubulogenesis. MCF10A cells embedded in a mixed collagen–rBM matrix placed within a type I collagen gel formed ducts and invaded the collagen gel and were seen only in the interface between the two matrices. Type I collagen is an abundant component of the breast ECM, whose expression is spatially and temporally regulated during mammary ductal formation. 34
As mentioned above, the role of collagen in tubulogenesis may also be attributed to the biomechanical properties of the matrix, mostly because of fiber organization. We addressed this option by analyzing fiber organization using picrosirius red staining and polarized-light microscopy. Our results showed that a high concentration of rBM disrupted collagen fiber organization. Also, the organization of collagen fibers was different in the vicinity of elongating epithelial structures, where the collagen fibers seemed to be pulling the epithelial structures at the elongating site; this was not observed near the round alveoli. Similarly, we previously observed that MCF10A cells facilitated the organization of collagen into thick fibers at the tip of the ducts and parallel to the direction of ductal elongation. 16 MCF10A cells formed ducts along the collagen fiber axis when cultured in collagen gels under uniaxial tension, which induced anisotropic arrangement of collagen bundles. 16 The collagen remodeling occurring in mixed gels containing low rBM concentrations appeared to be sufficient to create conditions of local biomechanical anisotropy in the vicinity of epithelial structures. The arrangement of collagen fibers along the axis of ductal elongation suggests that the locally generated anisotropy could be responsible for the initiation and direction of tubulogenesis. Collagen fiber organization was also shown to play a role in vivo by facilitating migration and invasion of tumor cells.35,36
Establishing the correct polarity of the epithelial cells is essential for tissue morphogenesis and function. Our results show that the presence of rBM in the matrices is critical for cell polarization; more importantly, with concentrations below a threshold of 10% rBM, sialomucin expression shifted its location from apical to pericellular. Coincidentally, with lower rBM concentrations, the expression of the basement membrane protein laminin 5 also became diffuse and discontinuous. These data are consistent with reports showing that ECM preparations containing laminin 1 impart the correct apical–basal cell polarity to spheroid arrangements of primary luminal epithelial cells, in contrast to type I collagen.37,38 Gudjonsson and colleagues also showed that the addition of normal myoepithelial cells, which secrete laminins, can reverse the resulting “inside-out” epithelial polarity typical of cells cultured within collagen gels. 37 Therefore, both biochemical and biomechanical signals seem to be important to maintaining epithelial polarity and differentiation.
In addition to the role played by the ECM, we investigated the contribution of stromal cells to mammary tubulogenesis. We observed that RMF could reverse the inhibitory effect that high concentrations of rBM had on tubulogenesis, although this phenomenon may not be fully attributed to secreted molecules because the RMF-conditioned medium was not sufficient to induce ducts in mixed gels. Ducts developed only when RMF were near MCF10A cells. Therefore, short-range factors secreted by RMF or a local remodeling of the ECM is required for tubulogenesis. Because fibroblast filopodia are capable of traversing a 0.2-μm porous membrane, we cannot exclude that physical contact might be necessary; live imaging analysis of growing ducts will be necessary to exclude this possibility.
HGF was previously shown to stimulate ductal formation in other 3D culture models.22,39 In our model, the addition of two different concentrations of HGF to mixed gels containing MCF10A cells was insufficient to induce ducts. Together, these findings highlight the complexity of 3D culture models containing various ECM components and more than one cell type. Data gathered from different 3D culture models ought to be interpreted in the context of each experimental design. To address whether the local remodeling of ECM was responsible for tubulogenesis, preliminary data in our laboratory showed that the inhibition of matrix metalloproteinases was not sufficient to abrogate tubulogenesis when RMF and MCF10A cells were co-cultured in mixed gels (data not shown). Therefore, the role of RMF in inducing tubulogenesis is most likely due to multiple biochemical and biomechanical factors.
Only a few in vitro studies have focused on the interactions between adipocytes and epithelial cells. Most of the studies used the nonmammary mouse preadipocyte cell line 3T3-L1,23,40–43 conditioned media from other adipocyte cell lines, or mammary adipose tissue.44,45 To overcome these limitations, we added human PA and adipocytes to our 3D morphogenesis model. We demonstrated that human PA were able to differentiate into mature adipocytes in 3D cultures in type I collagen and mixed gels. Differentiated cells were capable of producing leptin and secreted basement membrane. To our knowledge, this is the first time that this level of adipocyte differentiation and leptin production has been shown in a 3D culture model.
Adipocyte-secreted factors have been shown to influence normal and tumor cells by promoting the development of branching epithelial networks and tumor invasion through increased motility and proliferation. 46 When we explored the interactions between human adipocytes and breast epithelial cells cultured in various ECM, we observed that the epithelial phenotype developed in the presence of adipocytes was similar to that of MCF10A cells cultured with RMF in either matrix. Briefly, acini and ducts were observed in collagen matrices, and only acini formed in mixed gels containing 50% rBM, but the proximity of adipocytes to MCF10A cells reversed this phenomenon.
Also, undifferentiated pre-adipocytes induced tubulogenesis and branching when co-cultured with MCF10A cells in a mixed collagen and rBM matrix. We previously showed that MCF10A cells can undergo ductal morphogenesis when co-cultured with pre-adipocytes on 3D porous silk scaffolds. 47 Thus, we conclude that stromal cells at various differentiation states secrete factors and reorganize the matrix to enable breast epithelial cells to form ducts.
Conclusions
We have used a physiologically relevant 3D culture model to study epithelial tubulogenesis. This in vitro system shed light on the complex interactions between multiple cell types and the chemical and physical cues originating in the microenvironment surrounding mammary epithelial cells. Substrate pliability and collagen fiber organization are important physical factors influencing the response of many cell types. The stroma, and in particular the ECM, profoundly enhance morphological and functional differentiation of mammary epithelial cells. Reciprocally, stromal cells are able to modify the matrix by modulating the stiffness of the gels, generating mechanical forces, and changing collagen fiber organization. All of these factors contribute to tubulogenesis. In the experiments reported here, all stromal cell types contributed to the fate of the epithelial cells and their phenotype in a similar manner.
Our modified in vitro model combines three different cell types and allows each of them to grow and maintain a differentiated state alone or in combination with others, leading toward an increasingly complex engineered mammary gland surrogate model. This tissue morphogenesis model could be exploited in future studies to further define the mechanisms whereby ECM components and biomechanical properties of the matrix and tissue-specific stromal cells regulate tissue development, to identify which mechanisms have been disrupted when tissue disorganization and abnormal growth occurs, and to explore new pharmacological approaches to restore normality.
Footnotes
Acknowledgments
We appreciate the excellent technical contributions of Maria Banica, Elizabeth Colby, Cheryl Schaeberle, Andrew Tharp, and Jennifer Iordachescu. Department of Defense grant DAMD17-03-1-0467 supported the graduate training of Silva Krause. The research was supported in part by Phillip Morris International and the Parsemus Foundation.
Disclosure Statement
The authors confirm that there are no known conflicts of interest associated with this publication and there has been no significant financial support for this work that could have influenced its outcome.