
Abstract
Osteoporosis is a skeletal disorder characterized by reduced bone mineral density (BMD) and increased risk of fracture. We studied the effects of cell therapy of human adipose tissue-derived stromal cell (ADSC) on ovariectomy-induced bone loss in T cell deficient nude mice. Twelve-week-old female nude mice underwent ovariectomy and were treated with ADSC, estrogen, or phosphate buffered saline (PBS). Whole body BMD revealed that treatment of ADSC was more protective against ovariectomy-induced attenuation in bone mass gain compared with PBS control after cell therapy (8.4±1.1 vs. 2.4%±1.4%, p<0.05 at 4 weeks, 13.7±1.3 vs. 7.7%±1.8%, p<0.05 at 8 weeks) and this effect was comparable to that of estrogen. μCT analysis revealed that the effect of ADSCs was specific to trabecular bone. Serum osteocalcin levels were increased 4 weeks after ovariectomy and treatment with ADSCs (76.4±11.6 ng/mL) increased osteocalcin to a greater extent when compared with estrogen (63.1±6.7 ng/mL, p<0.05) or PBS treatment (58.0±9.2 ng/mL, p<0.05). Flow cytometry analysis for PKH26-labeled ADSCs and quantitative real-time PCR analysis for human β-globin from bone revealed that transplanted ADSCs were trafficking in bone 48 h after injection and subsequently disappeared. There was no evidence of long-term engraftment of infused ADSCs in bone. In vitro, treatment with ADSC-conditioned medium enhanced osteogenic differentiation in stromal cells and preosteoblasts. These results suggest that cell therapy of ADSCs protects against ovariectomy-induced bone loss in nude mice in a paracrine manner.
Introduction
The imbalance of bone remodeling in which bone formation by osteoblasts is surpassed by osteoclast-mediated bone resorption is a main pathophysiology of bone loss in osteoporosis.4–6 Therefore, although most of current therapies for osteoporosis are aiming at inhibition of bone resorption, promoting bone formation using anabolic agents that can enhance osteoblastic activity is a reasonable approach for treatment of osteoporosis. However, teriparatide, the only currently available anabolic agent for osteoporosis, has to be injected everyday and very expensive. A number of bone growth factors including BMPs 7 are being explored as new anabolic therapy but they can be applicable only for local therapy in selected cases of non-union or spinal fusions.8–10 In this regard, it would be plausible to consider cell therapy with mesenchymal stem cells (MSCs), a prospective source of osteoblastic precursors, for physiologic treatment of osteoporosis. After being considered as a proper source for cell therapy in bone regeneration, 11 most studies were focused on bone marrow derived MSC (BMMSC). However, recently, white adipose tissue has emerged as a suitable and alternative reservoir of progenitor cells. 12 Compared to BMMSC, adipose tissue-derived stromal cells (ADSC) are similarly able to differentiate into lineages of mesodermal cells and tissues such as adipocytes, bone, cartilage, and skeletal muscle (reviewed in ref 13 ). Further, the abundance in the body, easy and repeatable accessibility, and larger yield by simple isolation procedure compared with BMMSC represent clear advantages of the ADSC over BMMSC. 13 For this reason, there are growing numbers of in vivo applications utilizing ADSC therapy in various diseases such as osteoarthritis, 14 disc degeneration, 15 liver regeneration, 16 and vascular ischemia. 17
We previously performed syngeneic murine BMMSC mediated gene therapy for preventing bone loss in ovariectomized mice18,19 and confirmed that MSCs could clearly protect against overiectomy (OVX)-induced bone loss in these animals, 18 although the mechanism for this positive effect still remained unclear. Having confirmed the feasibility of syngeneic MSC therapy in osteoporosis treatment and/or prevention, we next decided to study the effects of xenotransplantation of human MSC in mice model of osteoporosis. Toward this end, we used human ADSCs as they are more readily available as previously described and would be a more feasible approach in human study in the future as well.
We found that human ADSCs effectively prevented OVX-induced bone loss not by long-term engraftment of transplanted cells but by short-term cell trafficking in bone maybe in a paracrine manner.
Materials and Methods
Source of adipose tissue and isolation of ADSC
Human visceral adipose tissues obtained from patients who received abdominal surgery were used for cell cultures. This study was approved by the Institutional Review Board of Seoul National University Hospital (H-0809-075-258) and all participants gave their written, informed consent to participate in the study. Adipose tissues were digested in 0.075% collagenase type I solution (type A; Roche) with gentle agitation for five digestions of 15 min each in a 37°C incubator. The first digestion was discharged and second to fifth suspensions were pooled together and filtered. Cells were centrifuged at 1200g for 5 min, supernatants were discarded and the cell fraction was treated with red blood cell lysis buffer for 5 min. Cells were cultured in α-MEM (Gibco) supplemented with 10% FBS. After 3∼4 days, non attached cells were discarded and the remaining cells were designated as “passage 0 (P0)”. P1 to P2 cells were used for all experiments. To characterize ADSCs, flow cytometry was performed using appropriate antibodies. The antibodies anti-human CD 29, CD 31, CD 106, STRO-1, and CD 44 were obtained from BD Bioscience and anti-human CD 34 and CD 105 were purchased from AbD Serotec.
Experimental design to study the effects of ADSC transplantation on OVX nude mice
Female BALB/c-nu Slc mice aged 12 weeks were purchased from Japan SLC Inc. All animal experiments were performed under approval from the Institutional Animal Care and Use Committee of Seoul National University. On study day −1, bilateral OVX or Sham operation was performed by standard method 20 for “OVX” or “Sham” groups, respectively. On study day 0 and 7, 4∼5×105/19∼20 g mice of human ADSCs were injected into the lateral tail veins of mice. Phosphate buffered saline (PBS) was injected intravenously as a negative control and estrogen (50 μg/kg, 3 days/week for 8 weeks) was injected intraperitoneally as a positive control. Whole body BMD (g/cm2) and body composition were measured using a PIXImus densitometer (software version 2.0; GE Lunar) as previously described 19 at week 0, 4, and 8 after surgery. Sera were obtained from retro-orbital plexus at week 0, 4, and 8 and serum osteocalcin was measured by the mouse osteocalcin ELISA kit (Biomedical Technologies, Inc.). After 8 weeks, the mice were sacrificed by CO2 gas asphyxiation and right femurs from each mouse were fixed in 70% ethanol at least 1 week. Fixed femurs were scanned by micro-computed tomography (μCT, Skyscan 1076, Kontich, Belgium).
Experimental design to evaluate short-term cell trafficking
Experimental design
Isolated ADSCs (5∼10×106) were stained with the PKH26 labeling kit (Sigma Chemical Co.) according to the manufacturer's protocol. PKH26-labeled ADSCs were counted and viable cells were suspended in 70 μL of PBS for in vivo injection. To evaluate stability of the dye, in vitro culture of PKH26-labeled ADSCs were performed in parallel. Flow cytometric analysis showed that PKH26-positive cells were over 99% at each time point till 1 week although the intensity of fluorescence was decreased suggesting that the dye was divided and diluted as the cells divided. On study day −1, bilateral OVX was performed and PKH26-labeled ADSC (4∼5×106) or PBS was infused via tail vein on study day 0. On 8, 24, 48, 72 h, and 1 week after one-time cell infusion, mice were sacrificed (n=4 per each group).
Quantitative real-time PCR for human β-globin
To quantify the amount of human DNA in each mouse tissue (femur, heart, kidney, liver, lung, and spleen), all tissues including whole femurs were immediately frozen with liquid nitrogen and crashed. Genomic DNA was extracted using DNA purification and extraction kit (Promega) per manufacturer's protocol and 20 ng of purified DNA was amplified to detect human specific β-globin using TaqMan universal PCR master mix (Applied Biosytems). Mouse β-actin gene was used as an internal control. The primers and probe for human β-globin were forward primer 5′–CTCCTTAAAC CTGTCTTGTAACCTTGA–3′; reverse primer 5′–GGGCA AGGTGAACGTGGAT–3′; and the probe labeled with fluorescent reporter and quencher 5′–FAM–CCTCACCACCA ACTTC–3′.
FACS analysis for PKH26-labeled cells
Whole bone marrow cells were flushed from femurs and analyzed for the presence of PKH26-positive cells by flow cytomtery (FACS Vantage SE; Becton Dickinson) using 106 cells per sample. Cells from PBS injected mice were used as negative controls. Bone marrow cells collected immediately after direct intratibial injection of PKH26-labeled cells were used as positive controls.
Histologic analysis
Right femurs from each mouse were directly embedded in frozen Optimal Cutting Temperature (Tissue-Tek® O.C.T.) compound (Sakura Fineteck USA Inc.). Serial sections (5 μm) were cut on a cryostat microtome at −20°C and frozen sections were mounted on glass slides, air-dried, and acetone-fixed. The sections were evaluated using fluorescence microscopy. Diaphysis and an area ∼200 μm proximal from the distal metaphysis of the femur were used for counting PKH26-positive cells. Negative control tissues were obtained from PBS treated mice.
Preparation of ADSC-conditioned medium
To generate conditioned medium (CM), human ADSCs, dermal fibroblasts (DFB; a kind gift from Dr. Jin Ho Chung, Seoul National University, Seoul, Korea) or human embryonic kidney 293T cells (purchased from ATCC) were cultured in 9-cm2 tissue culture dishes with α-MEM including 10% fetal bovine serum. At 80% confluency, medium was replaced with 5 mL of serum free α-MEM and harvested after 48 h to yield ADSC-CM or DFB-CM.
In vitro differentiation study
For osteogenic differentiation, UAMS-32 stromal cells (a kind gift from Dr. Mijung Yim, Sookmyung Women's University, Seoul, Korea) or MC3T3-E1 cells (purchased from ATCC) were seeded in 12-well plates at a density of 20,000 cells per well. After reaching confluence, ADSC-CM (50% or 100%), DFB-CM (50% or 100%) or Dexamethasone (10 nM) in α-MEM was added and maintained for 7 days in the presence of L-ascorbic acid (50 μM) and β-glycerophosphate (2 mM) to induce osteogenic differentiation. Alkaline phosphatase (ALP) activity was measured as previously described. 21 For osteogenic marker gene assay, MC3T3-E1 cells were culture with ADSC-CM (100%), DFB-CM (100%), 293T-CM (100%), or Dexamethasone (10 nM) in α-MEM combined with L-ascorbic acid (50 μM) and β-glycerophosphate (2 mM) for 5 days.
For bone marrow macrophage (BMM) culture, bone marrow was flushed with α-MEM from long bones of 6-week-old mice. To induce osteoclast differentiation, bone marrow cells were placed in a 48-well culture plate at a density of 2×105 cells/well and cultured with ADSC-CM, DFB-CM (50%, 100%), or 293T-CM (100%) containing 30 ng/mL M-CSF (Peprotech) and 50 ng/mL RANKL (Peprotech). After 5∼6 days of culture, cells were fixed and stained for TRAP, a marker of OC differentiation, according to manufacturer's instructions (Sigma-Aldrich).
For gene marker analysis, cells were washed with PBS twice and mRNA was isolated by TRIzol (Invitrogen) per manufacturer's protocol. cDNA was synthesized from RNA (1 μg) using Multiscribe reverse transcriptase. TaqMan universal PCR master mix was used with the mouse ALP (Mm00475831_m1; FAM dye), osteocalcin (Mm03413826_mH; FAM dye), TRAP 5b (Mm00475698_m1; FAM dye), and cathepsin k (Mm00484039_m1; FAM dye) probe/primer sets from Applied Biosystems. Rodent glyceraldehyde-3-phosphate dehydrogenase (VIC reporter dye) was used as an endogenous control. Amplification of cDNA was performed using the ABI Prism 7700 Sequence Detection System (Applied Biosystems). The amplification program was set for 1 cycle at 50°C for 2 min, 1 cycle at 10°C for 10 min followed by 40 cycles of 95°C, 15 s; 60°C, 1 min using the Applied Biosystems 7500 Real-Time PCR System. Relative quantification of data generated using this system was performed using the standard curve method.
ELISA
Immunoassays of human RANKL (BioVendor) or human osteoprotegrin (R&D Systems) were performed per the manufacturer's protocols.
Statistical methods
All data are presented as mean±S.E. The data were analyzed with the nonparametric Mann–Whitney test as appropriate. A significance level of 5% was assigned to all tests.
Results
Phenotypic characteristics of human ADSC
Cells isolated from human intra-abdominal fat were initially plated in 9-cm2 culture plates. After 4∼5 days culture, ADSC reached 80% confluence in a monolayer and exhibited uniform fibroblast-like cell morphology. To further characterize the ADSC population, cell surface marker profile was examined by flow cytometry. Consistent with previous data, 22 ADSCs showed positive staining for mesechymal linage related cell adhesion molecules such as CD29 (99%, high), CD44 (98%∼99%, high), CD 105 (94%∼95%, high), CD 106 (94%∼95%, high), and STRO-1 (14%∼30%, low) and negative staining for hematopoietic (CD34) and endothelial (CD31) lineage markers (1%–2%, respectively). Passage 1, 2, and 3 ADSCs were able to display not only osteogenic differentiation in culture medium containing L-ascorbic acid and β-glycerophosphate but also adipogenic differentiation in medium containing rosiglitazone (1 μM), isobutylmethylxanthine (0.2 mM), insulin (10 μg/mL), and dexamethasone (data not shown).
Transplantation of ADSCs prevents OVX induced bone loss in nude mice
At study day 1, 12-week-old female nude mice underwent OVX or sham operation and human ADSCs (OVX/ADSC), estrogen (OVX/E2), or PBS (OVX/PBS) were systemically administered twice at day 0 and 7. Sham operated mice with PBS injection (Sham/PBS) were used as control. Eight weeks after operation, uterine weights significantly decreased by 84%∼88% in all OVX mice compared with sham-operated mice indicating successful induction of estrogen deficiency (data not shown). There was no significant difference in total body weight and body fat percent between groups. Although the mice in all groups gained bone mass, the OVX/PBS mice gained significantly less bone mass compared with Sham/PBS group as expected (Fig. 1). Transplantation of ADSCs in this setting has significantly prevented OVX-induced bone mass attenuation measured over 8 weeks postsurgery. Indeed, OVX/ADSC showed a significant increase in percent change of BMD compared with OVX/PBS (8.4±1.1 vs. 2.4%±1.4%, p<0.05 at 4 weeks, 13.7±1.3 vs. 7.7%±1.8%, p<0.05 at 8 weeks; Fig. 1). Of note, the effect of ADSC therapy was comparable to that of estrogen treatment (Fig. 1).
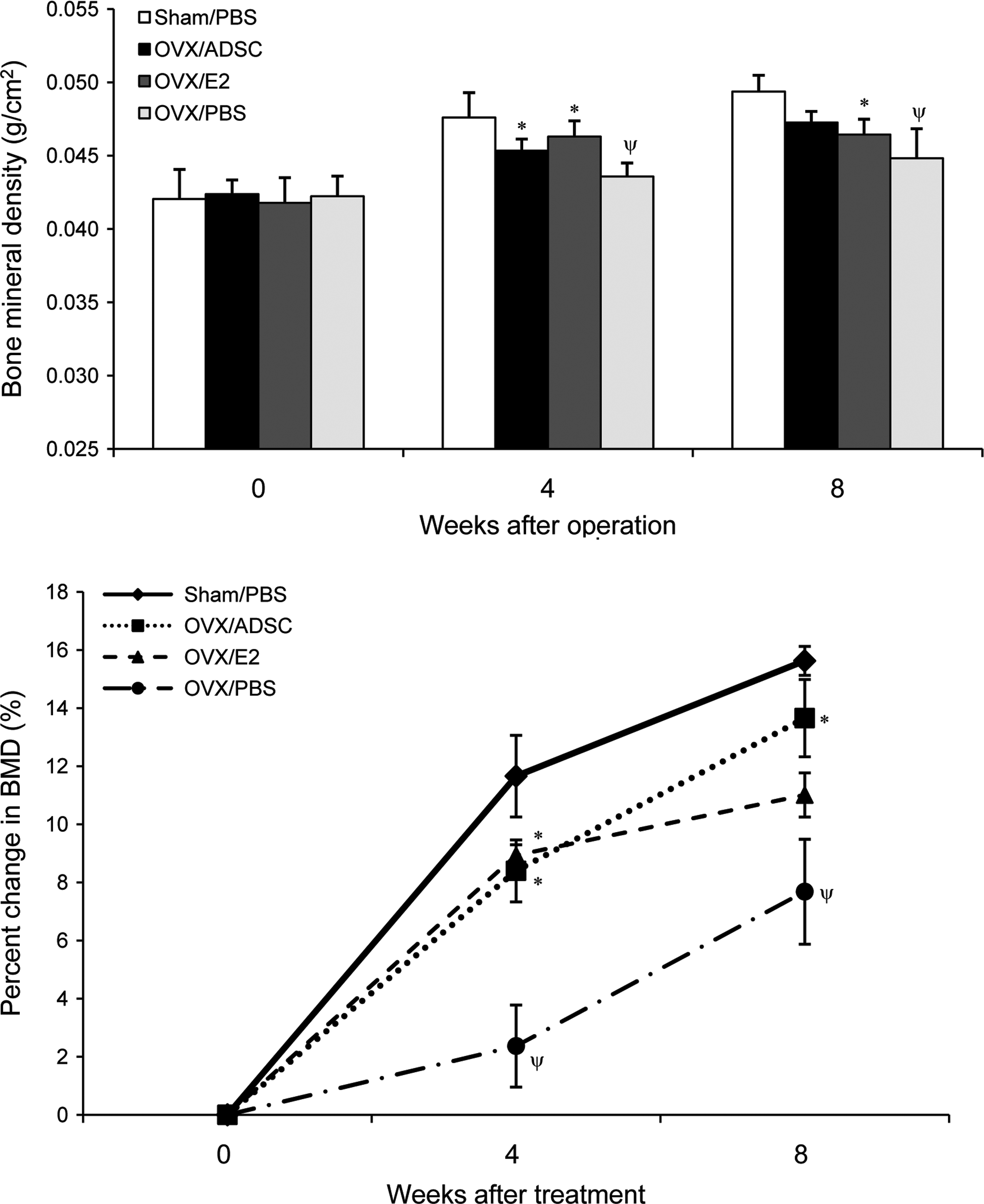
Therapeutic effect of ADSCs on bone mineral density (BMD) in ovariectomized nude mice. Twelve-week-old female nude mice were ovariectomized and treated with ADSCs (OVX/ADSC, n=9), estrogen (OVX/E2, n=6), or PBS (OVX/PBS, n=6) at day 0 and 7. Sham-operated mice were treated with PBS on the same days (Sham/PBS, n=6). In vivo total body BMD (g/cm2) was measured by dual energy X-ray absortiometry (PIXImus) on weeks 0, 4, and 8 and absolute values (upper graph) or percent changes (lower graph) of BMD were analyzed. Data are presented as mean±S.E., *p<0.05 versus OVX/PBS, Ψp<0.05 versus Sham/PBS. OVX, ovariectomy; ADSC, adipose tissue-derived stromal cell; PBS, phosphate buffered saline.
Measurement of μCT at 8 weeks suggested that the protective effect of ADSCs was specific to the trabecular component of bone (Fig. 2). OVX/ADSC showed a significant increase in fractional trabecular bone volume (BV/TV,%) compared with OVX/PBS (Fig. 2B). In addition, trabecular number (Fig. 2C) and thickness (Fig. 2D) were significantly increased while trabecular separation (Fig. 2E) showed a reciprocal decrease in OVX/ADSC compared with OVX/PBS. This increase in trabecular bone mass of OVX/ADSC was comparable to that of OVX/E2 or Sham/PBS. On the contrary, cortical bone volume (Fig. 2F) and thickness (Fig. 2G) measured in mid-diaphysis showed no significant difference between OVX/ADSC and OVX/PBS.
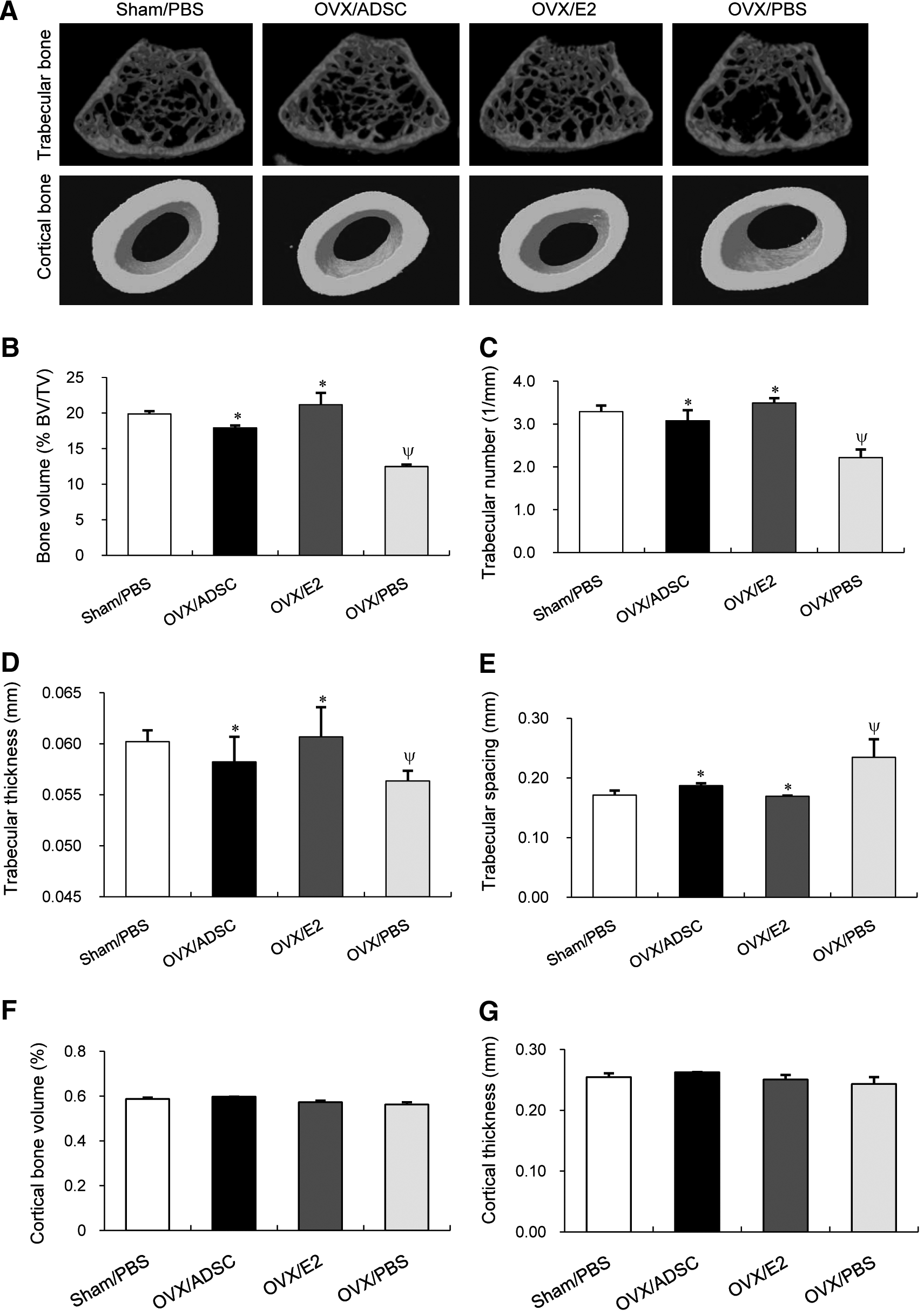
Therapeutic effect of ADSCs on trabecular bone measured by μCT. Twelve-week-old female nude mice were ovariectomized and treated with ADSCs (OVX/ADSC, n=9), estrogen (OVX/E2, n=6), or PBS (OVX/PBS, n=6) at day 0 and 7. Sham-operated mice were treated with PBS on the same days (Sham/PBS, n=6). Eight weeks after surgery, right femurs were harvested and analyzed by μCT.
Serum level of osteocalcin, an index of bone formation rate and osteoblastic activity, 23 was significantly increased by OVX by 45% (p<0.01) at 4 weeks and recovered at 8 weeks after operation (Fig. 3). Interestingly, OVX/ADSC group (76.4±11.6 ng/mL) showed significant increase in serum osteocalcin level compared with OVX/E2 (63.1±6.7 ng/mL, p<0.05) or OVX/PBS (58.0±9.2 ng/mL, p<0.05) at 4 weeks, suggesting that ADSC therapy may positively regulate bone formation (Fig. 3). Difference in osteocalcin levels between all groups disappeared at 8 weeks. Taken together, our results suggest that injection of human ADSCs prevented OVX-induced bone mass attenuation in nude mice, which seems to result from increase in bone formation especially at the trabecular bone compartment.
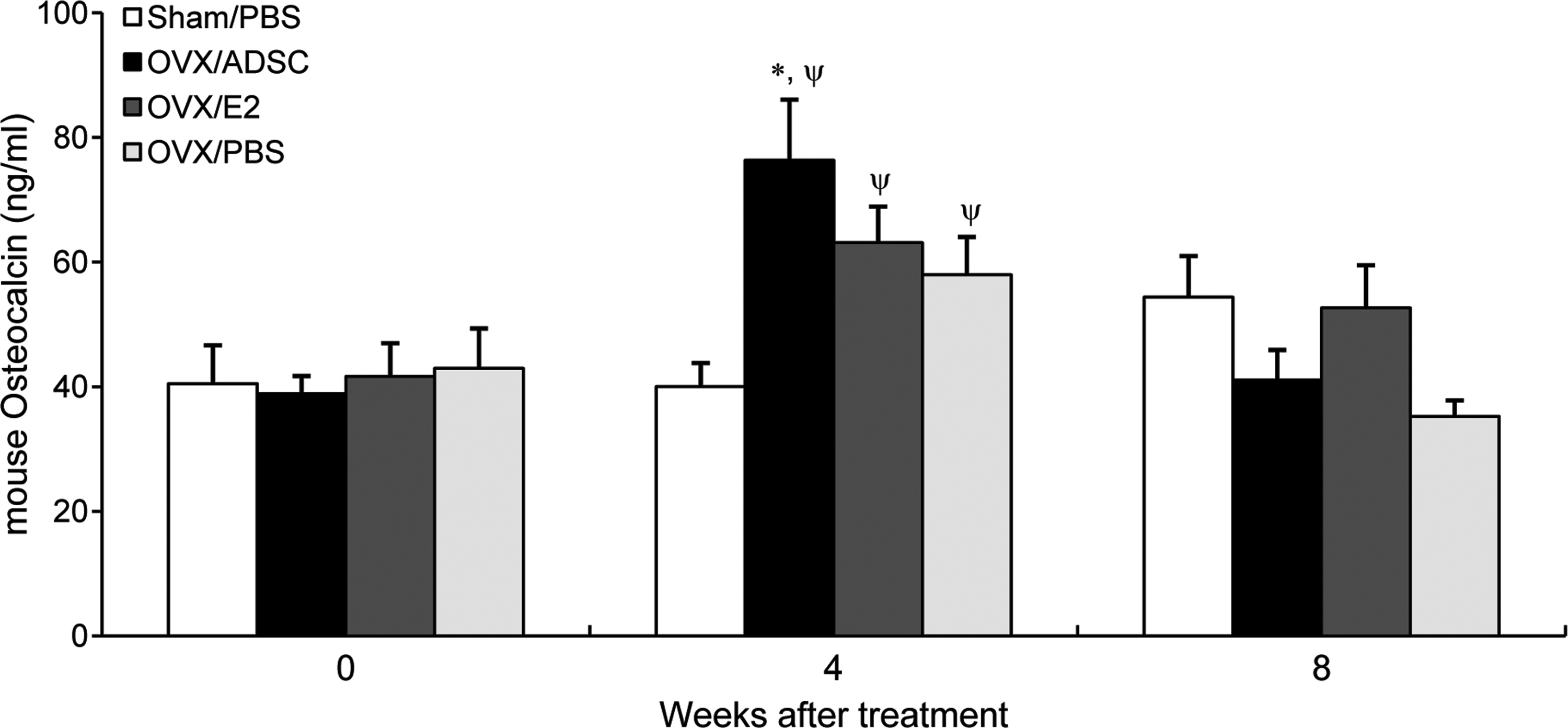
Serum osteocalcin, a marker of bone formation. Twelve-week-old female nude mice were ovariectomized and treated with ADSCs (OVX/ADSC, n=9), estrogen (OVX/E2, n=6), or PBS (OVX/PBS, n=6) at day 0 and 7. Sham-operated mice were treated with PBS on the same days (Sham/PBS, n=6). Sera were collected at week 0, 4, and 8 and serum osteocalcin levels were measured by ELISA. Data are presented as mean±S.E., *p<0.05 versus OVX/E2, Ψp<0.05 versus Sham/PBS. OVX, ovariectomy.
Short-term trafficking of ADSC after transplantation
To investigate the transplanted ADSC trafficking and engraftment efficacy, PKH26-labeled ADSC or PBS was injected to OVX mice and femurs and other tissues were harvested at different time points (8, 24, 48, 72 h, and 1 week after cell injection) as a parallel experiment.
First, we evaluated the in vivo tissue distribution of injected ADSCs at 48 or 72 h after cell injection using PCR amplification of human specific β-globin gene from various tissues (lung, heart, femur, kidney, liver, and spleen). Forty eight hours after injection, human β-globin gene copy numbers were highly detectable in the lung, while much less amount was distributed to various organs including femur. At 72 h, the signal from lung decreased while heart and kidney exhibited slightly increased copy numbers. In femurs, human specific β-globin gene was undetectable at 72 h, suggesting that transplanted human ADSCs had all passed through the bone after 48 h (Fig. 4A).
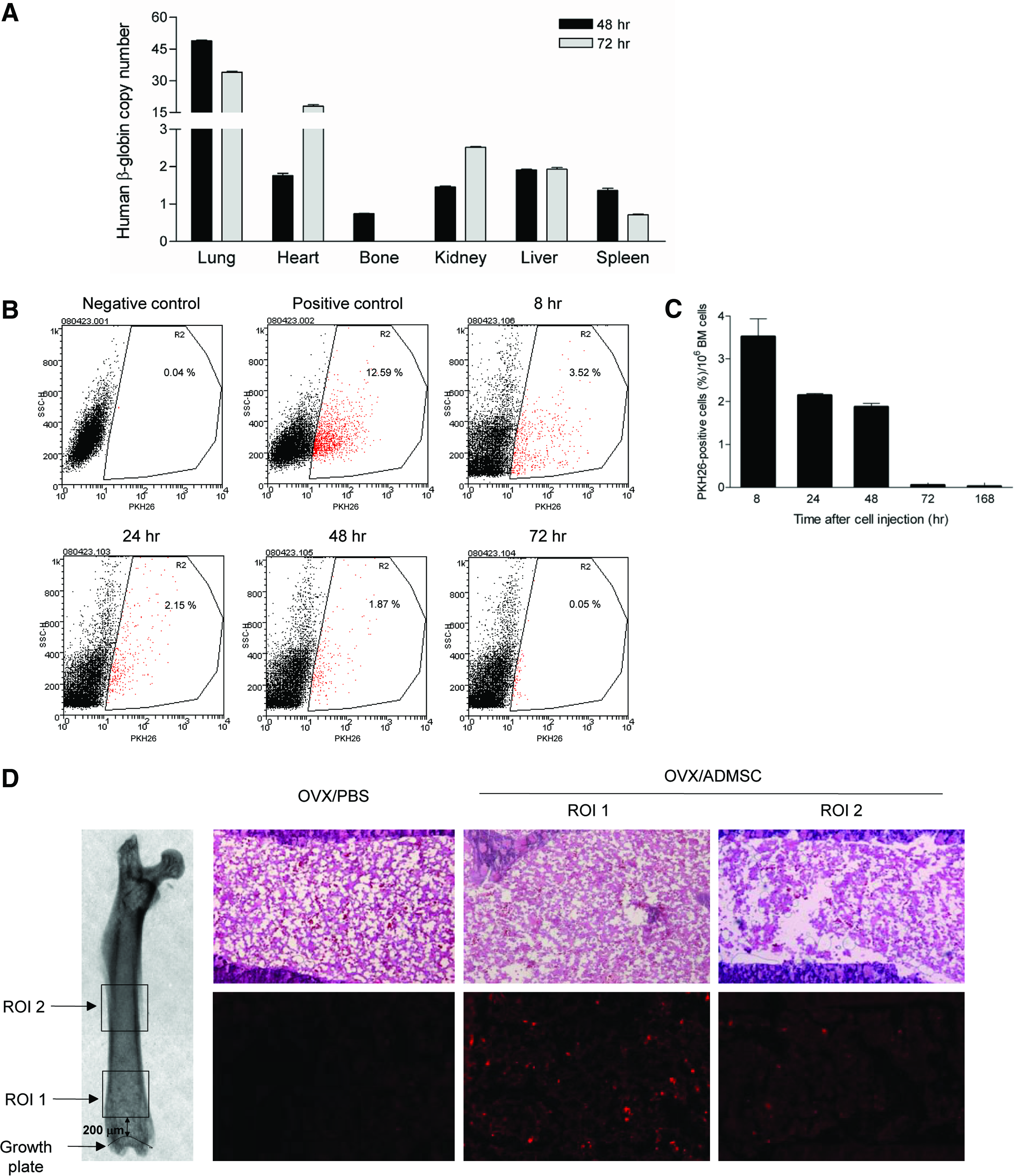
Cell trafficking assay. PKH26-labeled ADSCs or PBS was injected to OVX mice. Bone and other tissues were harvested at 8, 24, 48, 72 h, and 1 week after cell injection.
The direct evidence of ADSC trafficking to bone was further studied by flow cytometry. Bone marrow cells were flushed at 8, 24, 48, 72 h, and 1 week after injection of PKH26-labeled cells and the presence of these labeled cells were analyzed by flow cytometry. The fraction of PKH26-positive cells in bone marrow of OVX/ADSC was 3.52%±0.42% at 8 h after cell transplantation and it gradually decreased from 8 to 48 h. The negative control group (OVX/PBS) showed less than 0.05% (Fig. 4B–C). At 72 h after cell transplantation, PKH26-positive cells were not observed by flow cytometry in the OVX/ADSC group. In parallel, in vitro analysis under fluorescence microscope showed that PKH-26 labeling was maintained for 1 week.
Finally, we used histological assessment to demonstrate the presence of transplanted cells in vivo. To avoid false positive effects of autofluorescence, we used frozen sections and the metaphysical area was excluded from analysis. Consistent with the flow cytometry data, femurs harvested at 48 h clearly showed PKH26-positive cells but the fluorescent cells were not detectable at 72 h and 1 week. Average numbers of PKH26-positive cells were 28±4/mm2 in the diaphysis and 48±4/area in distal metaphysis at 48 h (Fig. 4D). Collectively, transplanted ADSCs systemically distributed in various organs with large amount of trapping at the lung. In bone, ADSCs were detectable during the initial 48 h but they subsequently disappeared with no evidence of long-term engraftment.
Effect of ADSC-CM on cell differentiation
Since the engraftment of ADSCs in bone seemed to be transient, we postulated that the therapeutic effect of transplanted ADSCs on OVX-induced bone loss resulted from paracrine action. To verify this hypothesis, the effect of ADSC-CM was tested with in vitro osteoblastic and osteoclastic differentiation assays. To induce osteogenic differentiation, UAMS-32 stromal cells or preosteoblast MC3T3-E1 cells were cultured in osteogenic medium including L-ascorbic acid and β-glycerophosphate. As shown in Fig. 5A, treatment of ADSC-CM (50% or 100%) significantly increased ALP activity compared with the control DFB-CM (50% or 100%), suggesting that ADSCs may secrete paracrine factors that can enhance osteoblastic differentiation. The effect of ADSC-CM was ∼35% of dexamethasone treatment (Fig. 5A). MC3T3-E1 cells yielded similar results (data not shown). Consistent with ALP activity data, ADSC-CM treatment significantly increased mRNA expression levels of ALP or osteocalcin compared with control-CM (293T- or DFB-CM; Fig. 5B). Next, we tested the effects of ADSC-CM on osteoclastogenesis. BMM cells were cultured with ADSC-CM or DFB-CM (50% or 100%) in the presence of M-CSF and RANKL to induce osteoclast differentiation. However, the number of TRAP-positive cells (Fig. 5C) or the mRNA expression of TRAP 5b and cathepsinK (Fig. 5D) were not changed by ADSC-CM. Interestingly, DFB-CM at both 50% and 100% has significantly inhibited osteoclast differentiation by 39% and 55%, respectively (Fig. 5C). The level of OPG was significantly higher in DFB-CM (48 pg/mL) compared with that of ADSC-CM (undetectable) although the level of RANKL was not significantly different between ADSC-CM (13.4 pmol/L) and DFB (16.2 pmol/L). Taken together, these results suggest that ADSCs secrete some factors that can promote osteoblastic differentiation in a paracrine manner, which could contribute to protection of OVX-mediated bone mass attenuation in nude mice model.

In vitro cell differentiation study with conditioned medium (CM). For osteogenic differentiation, cells were cultured in control or osteogenic medium (OM), containing L-ascorbic acid (50 μM) and β-glycerophosphate (2 mM), with indicated CM (293T-, DFB-, or ADSC-CM) or dexamethasone (10 nM) for 5 days.
Discussion
This study has demonstrated that systemic transplantation of human ADSCs, which are easily obtained and expanded with high yield, could efficiently prevent OVX-induced bone mass attenuation in nude mice. The therapeutic effect of transplanted ADSCs was as effective as estrogen and persisted for 8 weeks. Since transplanted ADSC showed short-term cell trafficking in bone within the initial 48 h after injection and there was no evidence of long-term engraftment of MSC in bone in vivo, we assumed that the protective effect of transplanted ADSC in bone was due to paracrine effects. This hypothesis was supported by the cell differentiation assay using ADSC-CM in which there was a significant increase of osteogenic differentiation of stromal cells in vitro.
Although osteoporosis has been ranked one of the top 10 potential target diseases for stem cell therapy, 24 systemic transplantation of MSCs has been rarely tried in osteoporotic models. One of the main issues in stem cell therapy is the cell homing and engraftment capacity to the affected tissue. Unlike acute tissue injuries such as skin wound, 25 ischemic hind limb, 26 or infracted myocardium (reviewed in ref 27 ) which could promptly recruit transplanted stem cells into the damaged sites, osteoporosis is a chronic progressing disease which involves the whole skeleton with relatively gradual environmental changes. For this reason, it is hard to expect that systemic transplantation of MSCs would efficiently recruit into osteoporotic bone and achieve therapeutic benefits. Most of the studies showing skeletal advantages with MSC therapy were performed by using local injection method in models of fracture healing, 28 calvarial defect, 29 or spinal fusion. 30 In our previous study, we found that systemically administrated murine BMMSC could effectively prevent bone loss after OVX, 18 and here we validated that xenotransplantation of human ADSC could similarly protect against OVX-induced bone mass attenuation, suggesting that systemic therapy of MSCs might have therapeutic potential. In this study we used T cell deficient mice to avoid immune response against human cell transplantation. T cell deficient nude animals have been showed conflicting results of bone phenotype after OVX.31,32 Cenci et al. showed that nude mice did not lose trabecular bone mass after estrogen withdrawal as measured by pQCT. 32 However, using histomorphometry, Sass et al. reported that nude rats underwent similar trabecular bone loss after OVX compared with wild-type control animals. 31 Recently, Lee et al. demonstrated that 4∼5-week-old nude mice showed comparable trabecular bone loss compared with wild-type mice at 4 weeks after OVX using μCT analysis. 33 Although these conflicting results seem to stem from the difference in sensitivity to detect the trabecular specific bone loss in response to estrogen withdrawal, we could successfully demonstrate bone mass attenuation using whole body DXA in 12-week-old nude mice. It is interesting to note that whole body BMD has actually increased in nude mice even after OVX because the mice we used in this study are still in growing phase. However, the percentage gain of BMD was significantly lower in OVX group compared with sham operated group indicating that OVX resulted in attenuation in bone mass gain rather than actual bone loss.
Notably, two times of systemic injection of ADSCs resulted in ∼6% increase in BMD compared with the control group from 4 to 8 weeks after OVX and this effect was comparable to that of persistent estrogen therapy. As the response to ADSC does not seem to result from engraftment of the cells per se, it would be less likely that the response is directly proportional to the number of doses. However, multiple injections could in theory increase the effect size compared with single injection because the estrogen deficiency-mediated bone loss should be continued. In clinical situations, reducing therapeutic frequency is a major strategy for treatment of chronic disease such as osteoporosis, prolonged effect of two injections in the present study might be promising. The beneficial effects of ADSCs seem to result from its anabolic effects on trabecular bone compartment. Indeed, the mice injected with ADSCs showed greater trabecular bone volume, trabecular number, and trabecular thickness, while no demonstrable effects were noticed in cortical bone. Since trabecular bone is a highly active tissue in bone metabolism and is the most sensitive to estrogen withdrawal, 34 trabecular bone loss is a major pathophysiologic change in postmenopausal osteoporosis in humans and rodents. 35
In this study, we observed that serum level of osteocalcin was increased after OVX, which was recovered at 8 weeks. Given that osteocalcin, a bone gla protein, is synthesized by osteoblasts and a proportion of these protein escape into the circulation, its serum level has been regarded as a marker of bone forming activity of the individual. However, as bone formation is tightly coupled with bone resorption under most conditions, the increased level of osteocalcin after OVX appears to represent increased turnover as a result of acute estrogen deprivation induced by ovariectomy.36–40 In contrast, significant increase in osteocalcin in mice injected with ADSC may truly reflect increased bone formation rate consistent with the increased BV/TV we have identified by microCT analysis.
Of note, the therapeutic effect of ADSCs appears to be mediated by paracrine manner. This notion comes from the finding that systemically injected ADSCs, which initially distributed in various tissues eventually disappeared 48∼72 h after administration, particularly in bone. At 48 h after injection, the ADSC trafficking in bone was 50%∼60% less than other tissues such as liver, kidney, and spleen analyzed by quantitative PCR of human specific β-globin gene, let alone the lung which has the pulmonary first-pass effect.41,42 Further, both human β-globin PCR signal and flow cytometry data of PKH26-labeled ADSCs demonstrated that ADSCs were detectable only during the initial 48 h and had disappeared in bone at 72 h after administration. These results are analogous with the previous study by us 18 and others18,43 that showed low homing efficacy and lack of long-term engraftment in murine BMMSC transplantation. Even with direct intrafemoral infusion of MSCs, Zhang et al. showed that only 2% of injected MSCs remain in bone at 24 h and the donor cells in bone compartment decreased over the first 2 weeks after injection. 43 Although there was limited cell trafficking and no engraftment of cells in bone, the therapeutic effect of ADSCs was as effective as that of estrogen and was maintained for 8 weeks. This apparent discrepancy suggested that instead of direct engraftment into bone and differentiation into osteoblasts, ADSCs which were distributed to the whole body and perhaps those specifically in bone marrow compartment produced secretory factors to exert their therapeutic effects on bone.
One of the major concerns in systemic therapy of MSCs is a possible systemic adverse event. Consistent with previous studies,41,42 our study showed that more than 50% of cells were trapped in the lung and passed to liver and spleen which belong to mononuclear phagocyte system. Although we did not observe signs of sickness or gross abnormalities in lung tissue, this initial lung trapping may not only limit the available number of ADSCs in systemic circulation but also result in embolization and local production of cytokines. 44 Another potential risk would be the possibility of developing tumors upon transplantation of ADSCs. Indeed, it is well recognized that injection of both human and mice MSCs after long-term in vitro expansion led to development of tumors possibly due to DNA mutation or chromosomal instability.45–48 Further, a recent report demonstrated similar chromosomal abnormalities and malignant tumor formation even after short-term culture of MSCs, underscoring the need for careful monitoring of chromosomal aberration in the process of in vitro expansion of MSCs. 49
Although we did not investigate specific factors which could be secreted by ADSCs, it is clear from the cell differentiation assay that ADSC-CM had an osteogenic effect on stromal cells in vitro as assessed by greater (∼60%) increase in ALP activity compared with control medium. This hypothesis is supported by previous studies which revealed that several cytokines relevant to osteogenesis such as hepatocyte growth factor, vascular endothelial growth factor (VEGF), insulin-like growth factor-1, and transforming growth factor β were expressed by human ADSCs.50–52 Moreover, it has been shown that ADSCs are also able to promote angiogenesis by secreting a number of pro-angiogenic factors including angiopoietin-1, stromal derived factor-1, and VEGF (reviewed in ref 53 ). These angiogenic factors may also play roles in the anabolic effect of ADSCs by recruiting host endothelial progenitor cells and stimulating neovascularization. Among the factors that regulate osteoclast differentiation, we were unable to identify significant difference in RANKL levels between ADSC CM and DFB CM although the level of OPG was significantly higher in DFB CM consistent with a recent study. 54
The results of our study are in good agreement with a recent study that also showed potential therapeutic effects of ADSCs on OVX-induced murine osteoporosis models. 55 In that study, Lee and colleagues have also demonstrated increased bone mass by systemic transplantation of ADSC in mice OVX model. One of the major differences from our study is that they have isolated ADSC from subcutaneous fat tissue, while we used visceral fat. We chose visceral adipose tissue because previous studies showed that visceral adipose tissues are less affected by age-dependent decrease of cell proliferation, 56 and exhibited higher osteogenic potential compared with subcutaneous fat tissues. 57 In addition, to minimize potential host immune reactions to human cells, T-cell deficient nude mice were used in the present study while immunocompetent ddY mice were used previously. It may in part contribute to more potent therapeutic effects of ADSC in this study compared with previous work, which used a half-dose of cells. In addition, in our study we have included an estrogen positive control group, which received estrogen in our study, for a quantitative comparison of the effect size mediated by ADSC therapy.
In summary, we have demonstrated that systemic administration of human ADSCs protected against OVX-induced bone mass attenuation in T-cell deficient nude mice. Given that there was no evidence of long-term engraftment, transplanted ADSCs appears to attenuate bone loss via a paracrine mode of action. The results of this study will shed more light on the potential use of ADSC therapy in the prevention or treatment of osteoporosis, a common but chronic debilitating disease that represents a major worldwide health problem.
Footnotes
Acknowledgments
This work was supported by a grant from Seoul National University Hospital (grant # 03-2007-053-0) and a grant from the Ministry of Education, Science and Technology of Korea (grant # 2009-0077579).
Disclosure Statement
No competing financial interests exist.