
Abstract
Clinical utilization of tissue-engineered cartilage constructs has been limited by their inferior mechanical properties compared to native articular cartilage. A number of strategies have been investigated to increase the accumulation of major extracellular matrix components within in vitro-formed cartilage, including the administration of growth factors and mechanical stimulation. In this study, the anabolic effect of inorganic polyphosphates, a linear polymer of orthophosphate residues linked by phosphoanhydride bonds, was demonstrated in both chondrocyte cultures and native articular cartilage cultured ex vivo. Compared to untreated controls, polyphosphate treatment of three-dimensional primary chondrocyte cultures induced increased glycosaminoglycan and collagen accumulation in a concentration- and chain length-dependent manner. This effect was transient, because chondrocytes express exopolyphosphatases that hydrolyze polyphosphate. The anabolic effect of polyphosphates was accompanied by a lower rate of DNA increase within the chondrocyte cultures treated with inorganic polyphosphate. Inorganic polyphosphate enhances cartilage matrix accumulation and is a promising approach to improve the quality of tissue-engineered cartilage constructs.
Introduction
Current surgical interventions to enhance the repair of cartilage such as marrow stimulation techniques and autologous chondrocyte implantation (ACI) have yielded limited long-term clinical success, partly because of the fibrocartilagenous nature of the resulting repair tissue. 1 Efforts to improve the functional outcome of cell-based techniques such as ACI have resulted in the development of numerous tissue-engineering approaches to deliver chondrogenic cells or in vitro-formed hyaline-like cartilage to the injury site. However, tissue-engineered cartilage is often characterized by low content of extracellular matrix components, specifically collagen type II, compared to native articular cartilage, which results in inferior tissue mechanical properties.4,5
Various approaches have been investigated to improve the quality of tissue-engineered cartilage, including the stimulation of tissue formation with growth factors during the in vitro maturation period and/or after implantation in vivo. Growth factors with demonstrated anabolic effects on cartilage include insulin-like growth factor 1, fibroblast growth factor 2 (FGF-2), FGF-18, transforming growth factor β1, bone morphogenetic protein 2 (BMP-2), BMP-4, BMP-5, and BMP-7 and have been reviewed in detail. 6 Our group and others have shown that mechanical stimulation of in vitro-formed cartilage increases extracellular matrix accumulation and yields improved tissue mechanical properties.4,7–10 Other techniques such as the application of ultrasound,11,12 laser stimulation, 13 as well as changes in the culture conditions such as oxygen tension14,15 and temperature 16 have all resulted in modest improvements in matrix accumulation. Despite these efforts, in vitro-formed cartilage remains qualitatively and functionally inferior to native articular cartilage and our understanding of the signals that stimulate extracellular matrix deposition by chondrocytes is still lacking.
Inorganic polyphosphate is a linear polymer of orthophosphate residues linked by energy-rich phosphoanhydride bonds that is ubiquitous in biological systems, as it has been identified in bacteria, fungi, algae, insects, plants, and animals.17,18 While polyphosphate has been extensively investigated in lower organisms, its functions in mammalian cells and tissues have only recently started to be uncovered. Treatment with the phosphate polymer has been shown to modulate mineralization in different tissue types through direct physicochemical interactions with nucleating mineral crystals, as well as biological responses.19–24 Polyphosphate also exhibits other biological effects, including the ability to direct mesenchymal stem cell differentiation toward the osteoblastic phenotype,25,26 to act as a pro-coagulant and pro-inflammatory mediator,27–29 to modulate the activity of ion channels, 30 to block tumour metastasis, 31 to regulate cell energy metabolism, 32 and to enhance the mitogenic activity of different cell types through stabilization of growth factors such as basic FGF and activation of the mTor pathway.33,34
Our group has demonstrated that exogenously administered polyphosphate reversibly inhibits the formation of a zone of calcified cartilage within in vitro-formed hyaline cartilage. 35 It was also observed that cartilage formed in vitro on a porous titanium substrate that was treated with condensed phosphate appeared thicker than untreated in vitro-formed cartilage (unpublished data). Furthermore, polyphosphate has been shown by others to functionally stabilize FGF-2, a growth factor that has been shown to exhibit an anabolic effect on chondrocytes. 33 Thus, it is hypothesized that the exogenous administration of polyphosphate to three-dimensional in vitro cultures of chondrocytes enhances cartilage tissue formation. In this study, the effect of polyphosphates of average chain lengths ranging from 5 to 75 phosphate units on matrix accumulation within in vitro-formed cartilage was examined along with the ability of chondrocytes to metabolize condensed phosphate through enzymes with an exopolyphosphatase activity. The responsiveness of different articular chondrocyte zonal subpopulations cultured in vitro and of native cartilage samples cultured ex vivo to polyphosphate was also investigated. Identifying molecules that positively impact matrix accumulation in cartilage will facilitate successful engineering of cartilage for joint repair.
Materials and Methods
Substrates
Membrane inserts (Millicell-CMR; Millipore Corp., Bedford, MA) were coated with type II collagen (0.5 mg/mL in 0.1 N acetic acid; Sigma Chemical Co., St. Louis, MO) and dried overnight. The membrane inserts were ultraviolet-sterilized for 30 min and soaked in Ham's F12 for 30 min before cell culturing.
Tissue culture
Cartilage was aseptically excised from the full thickness of metacarpal-phalangeal articular cartilage from 9- to 12-month-old calves within 24 h of death as previously described. 36 Chondrocytes were isolated from the tissue by sequential enzymatic digestion (0.5% protease [Sigma Chemical Co.] for 2 h followed by 0.1% collagenase [Roche Diagnostics GmbH, Mannheim, Germany] overnight). The cells were then seeded on top of membrane inserts (1×106 cells per membrane; 12-mm diameter) in Ham's F-12 supplemented with 5% fetal bovine serum (HyClone, Logan, UT) and incubated at 37°C in an atmosphere characterized by 95% relative humidity and 5% CO2. On day 5, the serum concentration was increased to 20%, and the medium was supplemented with ascorbic acid (100 μg/mL; Sigma Chemical Co.).5,13,35,37 At this time, cultures were incubated in the presence or absence of various concentrations of sodium phosphate glasses (inorganic polyphosphate; average chain lengths of 5, 45, and 75 phosphate residues; Sigma Chemical Co.) or sodium phosphate buffer. The concentrations of polyphosphate utilized for each experiment are calculated based on the phosphate content. Cultures were grown for various times up to 4 weeks after initiation of the inorganic polyphosphate treatment. The culture medium was changed every 2–3 days.
For the experiments investigating the response of chondrocyte zonal subpopulations to polyphosphate, the upper 70% (superficial-mid zone) and the bottom 30% (deep zone) of the cartilage layer were harvested separately and processed similarly to full-thickness chondrocytes. The efficacy of this dissection has been reported previously. 38
To investigate the effect of polyphosphate on native cartilage, 4-mm-diameter cartilage samples were obtained from excised full-thickness cartilage using a biopsy punch and cultured directly in 24-well plates under the same conditions as the in vitro-formed cartilage.
Histological evaluation
In vitro- and ex vivo-cultured cartilage tissues were harvested and washed twice in phosphate-buffered saline (PBS). The tissues were fixed in 10% formalin and embedded in paraffin. Five-micron-thick sections were cut, stained with toluidine blue, and examined by light microscopy. To visualize the presence of polyphosphate in the tissues, sections were cut, dewaxed in xylene, and stained with 4′-6-diamidino-2-phenylindole (DAPI) (5 μg mL−1 in dH2O; Pierce Biotechnology, Inc., Rockford, IL). The fluorescence was visualized with a Zeiss Axioplan epifluorescence microscope using a wide-pass DAPI filter. Inorganic polyphosphate specifically shifts the emission peak of DAPI from 456 to 526 nm, permitting its visualization in the yellow–green spectrum rather than the blue spectrum associated with nucleic acids or glycosaminoglycans (GAG).39,40
Dry weight of cartilaginous tissue
Cartilage tissues formed in vitro on membrane inserts were harvested, washed twice in PBS, and removed from membrane inserts. The tissues were then lyophilized overnight and weighed using an electrical balance (Explorer; Ohaus Corp., Florham Park, NJ).
Determination of the DNA content
Cartilage tissues were digested with papain (40 μg mL−1; Sigma Chemical Co.) in a digestion buffer (20 mM ammonium acetate, 1 mM ethylenediaminetetraacetic acid [EDTA], and 2 mM dithiothreitol) for 48 h at 65°C. Papain digests are then stored at −20°C until further analysis. The DNA content was assessed using the Hoechst 33258 dye (Polysciences, Inc., Washington, PA) binding assay and fluorometry (excitation wavelength, 365 nm; emission wavelength, 458 nm). 41 The standard curve was generated with calf thymus DNA (Sigma Chemical Co.).
Determination of the proteoglycan and collagen content
The proteoglycan and collagen contents of cartilage tissues were measured from aliquots of the papain digest. The proteoglycan content was estimated by quantifying the amount of sulfated GAGs using the dimethylmethylene blue dye (Polysciences, Inc.) binding assay and by quantifying the color spectrophotometrically at 525 nm.
42
The standard curve was generated with bovine trachea chondroitin sulfate A (Sigma Chemical Co.). The collagen content was estimated by quantifying hydroxyproline. Papain digest aliquots were hydrolyzed in 6 N HCl at 110°C for 18 h. The hydroxyproline of the hydrolysate was determined using the choramine-T/Ehrlich's reagent assay and the color change quantified spectrophotometrically at 560 nm.
43
The standard curve was generated with
Analysis of gene expression
Total RNA was extracted from in vitro-formed cartilage after culture in the presence or absence of inorganic polyphosphate for various time points up to 1 week using Trizol (Invitrogen, Carlsbad, CA). Total RNA was reverse transcribed into cDNA using Superscript II and Random Hexamers (Invitrogen) according to the protocol provided by the manufacturer. The resulting cDNA was diluted five times and subjected to relative quantitative polymerase chain reaction (qPCR) with a Realplex 2 Master Cycler (Eppendorf, Hamburg, Germany). Briefly, 4 μL of cDNA was amplified in 10-μL total volume with 0.6 μM of each primer suspended in Express SYBR Green ER (Invitrogen). Reaction mixtures were subjected to 10-min incubation at 95°C to activate the enzyme followed by 15-s denaturation at 95°C and 30-s annealing at 60°C. Amplification data were collected for 40 cycles. Mean relative quantification values were calculated with 18S rRNA as endogenous control and tissues harvested before the initiation of the polyphosphate treatment and media change (time point of 0 h) as calibrator. The genes investigated include the major cartilage matrix components collagen type II and aggrecan, as well as the chondrogenic transcription factor Sox9. The expression of collagen type I, which is generally associated with chondrocyte dedifferentiation, was also investigated. The efficiency of each primer pair was verified. Only primer pairs with an efficiency superior to 90% were used. Primer sequences have previously been published. 44
Exopolyphosphatase activity of cartilaginous tissues
In vitro-formed cartilage constructs were harvested and washed twice in PBS. Proteins were extracted and assessed for the exopolyphosphatase activity according to a protocol modified from Lorenz et al. 45 Briefly, cartilage tissues were freeze–thawed three times in 50 mM Tris-HCl buffer (pH 7.5) containing 10 mM MgCl2, 0.5 mM EDTA, 150 mM NaCl, and 0.2% Triton X-100 (homogenization buffer; 150 μL per construct) and mechanically homogenized. The resulting solutions were centrifuged (14,000 g for 15 min), and the protein content of the supernatant was measured using a Pierce BCA protein assay. Reaction mixtures were prepared for each sample containing 25 μg of protein and 0.4 μmol of sodium phosphate glass (average chain length of 45 phosphate units) in a reaction buffer (50 mM Tris-HCl buffer [pH 7.5], 10 mM MgCl2, 0.5 mM EDTA, and 150 mM NaCl) to a final volume of 200 μL. To correct for the phosphate content of protein extracts and hydrolysis of the condensed phosphate, mixtures containing only the protein extract (for each sample) or inorganic polyphosphate were also prepared. The reaction mixtures were incubated at 37°C for 24 h (the enzymatic phosphate release from polyphosphate is linear between 1 and 48 h). The phosphate released was measured spectrophotometrically at 620 nm after incubation with a solution containing 1:6 vol/vol ratio of 10% ascorbic acid and 0.42% ammonium molybdate (Sigma Chemical Co.) in 1 N H2SO4 (1:3 vol/vol ratio) at 37°C for 1 h. The phosphate standard curve was prepared using a sodium phosphate dibasic solution (pH 7.5), the homogenization buffer, and bovine serum albumin at protein levels equivalent to that of samples.
Statistical analysis
Experiments were done in triplicate or duplicate (for qPCR data) and repeated three times with cells from different extractions. Results are expressed as the mean±standard error of the mean unless specified otherwise and analyzed using a one-way analysis of variance (for more than two conditions) or Student's t-test (between groups). The Tukey's test post hoc analysis was performed. p-Values≤0.05 were considered statistically significant.
Results
Inorganic polyphosphate stimulates chondrocyte matrix accumulation
Chondrocytes obtained from full-thickness cartilage were grown in vitro on membrane inserts for 2 weeks in media supplemented with different concentrations of inorganic polyphosphate (chain length of 45 phosphate units). Treatment with polyphosphate at a concentration of 1 mM resulted in a significant increase in the total GAG (but not collagen) content per culture compared to cartilage formed in the medium that was not supplemented with polyphosphate (Fig. 1A). When the data were normalized to DNA, both the GAG and collagen contents were significantly increased by treatment with 1 mM polyphosphate (Fig. 1B). Lower concentrations of polyphosphate resulted in more modest increases in matrix accumulation, which were not significant. The DNA content of in vitro-formed cartilage treated with 1 mM polyphosphate was significantly lower than that in nontreated tissues (Fig. 1C).
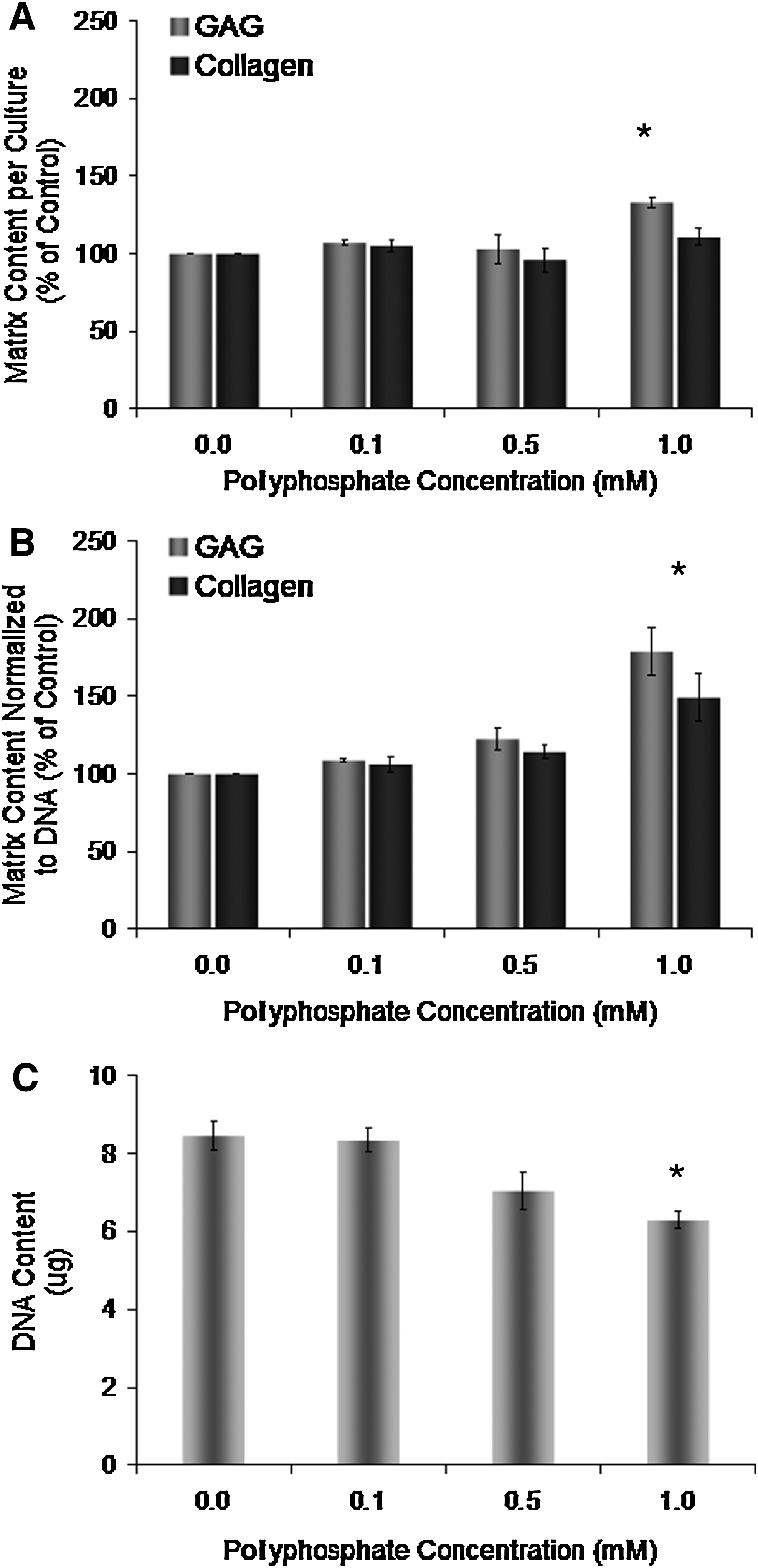
Effect of inorganic polyphosphate on in vitro-formed cartilage matrix and DNA accumulation.
Supplementation of the medium with equivalent concentrations of sodium phosphate to those used for polyphosphate treatment did not significantly alter the DNA, GAG, and collagen contents in these tissues compared to cartilage formed in the nonsupplemented medium (Table 1). This suggests that the stimulatory effect of polyphosphate on cartilage matrix deposition is specific to condensed phosphate rather than its degradation product, orthophosphate.
Chondrocytes were grown in the presence of phosphate (1 mM) or polyphosphate (1 mM based on the phosphate content; a chain length of 45 phosphate units). DNA, GAG, and collagen contents were determined after 2 weeks in culture. The data are from three experiments performed in triplicate and expressed as mean±standard error of the mean. The matrix content is normalized to DNA and expressed relative to untreated controls.
Significantly different content from untreated tissues (p<0.05).
GAG, glycosaminoglycan.
Inorganic polyphosphate does not upregulate the expression of cartilage matrix genes
Chondrocytes were grown in vitro on membrane inserts for up to 1 week in media supplemented with polyphosphate or left untreated. No significant difference was observed between the gene expression of polyphosphate-treated and nontreated tissues at any time point for cartilage-specific genes (collagen type II, aggrecan, and Sox9; Fig. 2) or the gene indicative of dedifferentiation in chondrocytes (collagen type I; data not shown).

Effect of polyphosphate treatment on cartilage-related gene expression. Total RNA was extracted from in vitro-formed full-thickness cartilage cultured in the absence or presence of polyphosphate (1 mM, calculated based on the phosphate content; an average chain length of 45 phosphate units) for different time points from 6 h to 1 week. The differential gene expression of
Effect of inorganic polyphosphate chain length on matrix accumulation
Chondrocytes were grown in vitro on membrane inserts for 2 weeks in a medium supplemented with polyphosphates characterized by various average chain lengths (5, 45, and 75 phosphate units; 1 mM). The GAG content per culture was significantly increased by treatment with polyphosphates characterized by chain lengths of 45 and 75 phosphate units compared to untreated controls, while collagen content per culture was not significantly influenced by the treatments (data not shown). When normalized to DNA, only polyphosphate with an average chain length of 45 phosphate units exhibited significant increases in both GAG and collagen contents in the tissues (Fig. 3A). Based on these results, polyphosphate with an average chain length of 45 phosphate units was employed for all further studies.
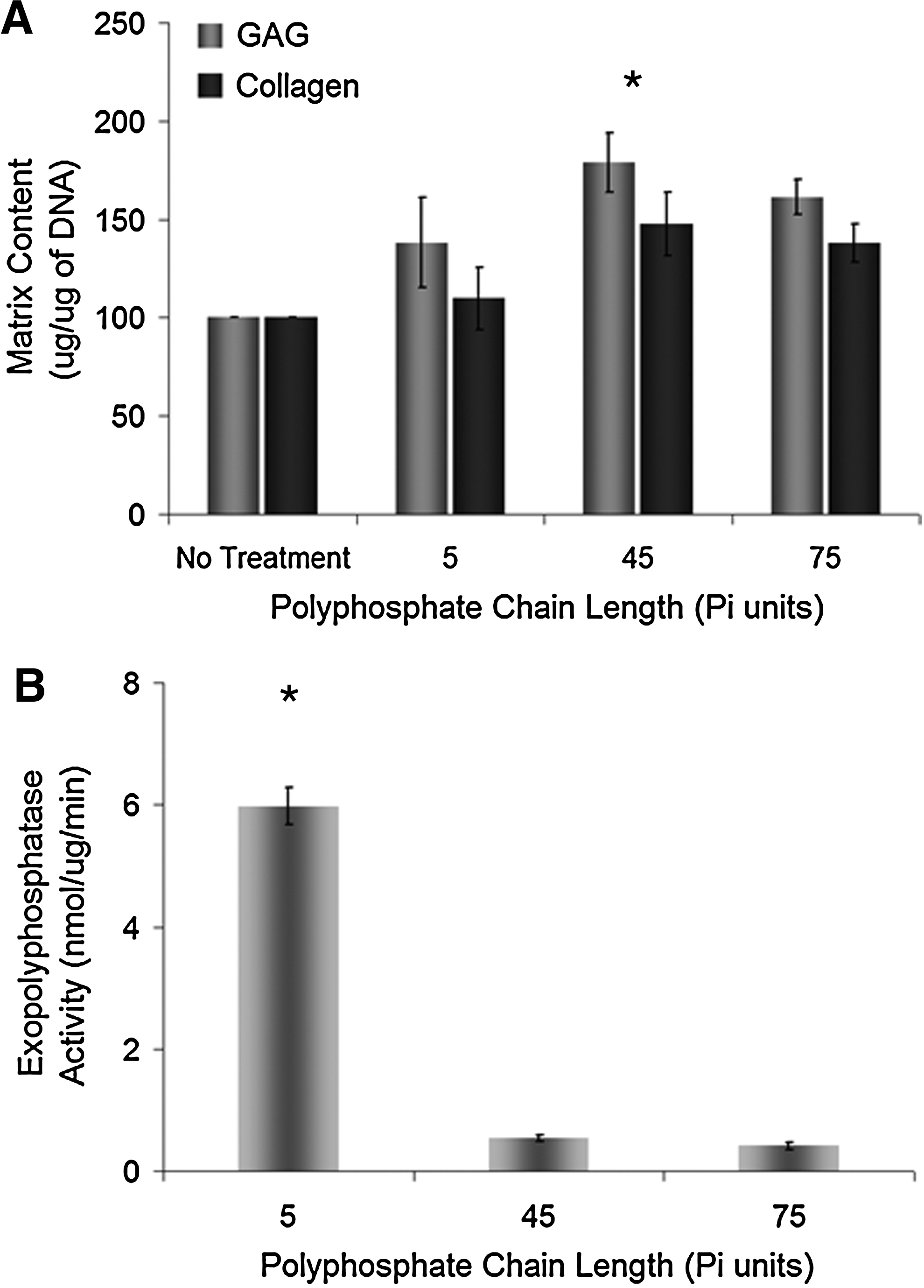
Effects of inorganic polyphosphate chain length on in vitro-formed cartilage matrix accumulation and the exopolyphosphatase activity.
To further understand the effect of polyphosphate chain length on cartilage matrix accumulation, the activity of exopolyphosphatases in protein extracts from 3-day-old in vitro-formed full-thickness cartilage was measured after reactions with polyphosphates of the three chain lengths investigated in this study. As shown in Figure 3B, exopolyphosphatases present in the in vitro-formed cartilage cleaved short chain polyphosphate (5 phosphate units) at a significantly higher rate than longer chain polyphosphate (45 and 75 phosphate units). No significant difference was observed between the degradation rates of the two longer chain length polyphosphates. This experiment was repeated with polyphosphate concentrations measured based on the polyphosphate rather than the phosphate content, and similar trends were observed (data not shown).
Time course of inorganic polyphosphate effect on matrix accumulation
Chondrocytes were grown in vitro on membrane inserts for 0, 1, 2, or 4 weeks after initiation of the treatment with 1 mM polyphosphate. The DNA content of nontreated tissues increased by >125% in 4 weeks, while polyphosphate-treated tissues exhibited a more modest rate of DNA accumulation of 75% (Fig. 4A). The DNA content of treated tissues was significantly lower than that of nontreated controls at 2 and 4 weeks. The GAG (Fig. 4B) and collagen (Fig. 4C) accumulation within tissues increased over the 4 weeks. The GAG content was significantly increased by treatment with polyphosphates compared to untreated controls at 4 weeks. After normalization to DNA, GAG (Fig. 4D), and collagen (Fig. 4E) accumulation within tissues formed in the absence of polyphosphate increased significantly in the first week of culture and then remained stable up to 4 weeks. Matrix accumulation within tissues treated with polyphosphate significantly increased up to 2 weeks of culture and remained stable until 4 weeks. The GAG content was significantly higher in tissues treated with polyphosphate compared to the untreated condition at 2 and 4 weeks. However, a significant difference between polyphosphate-treated and untreated tissues was only observed at 4 weeks for the collagen content.
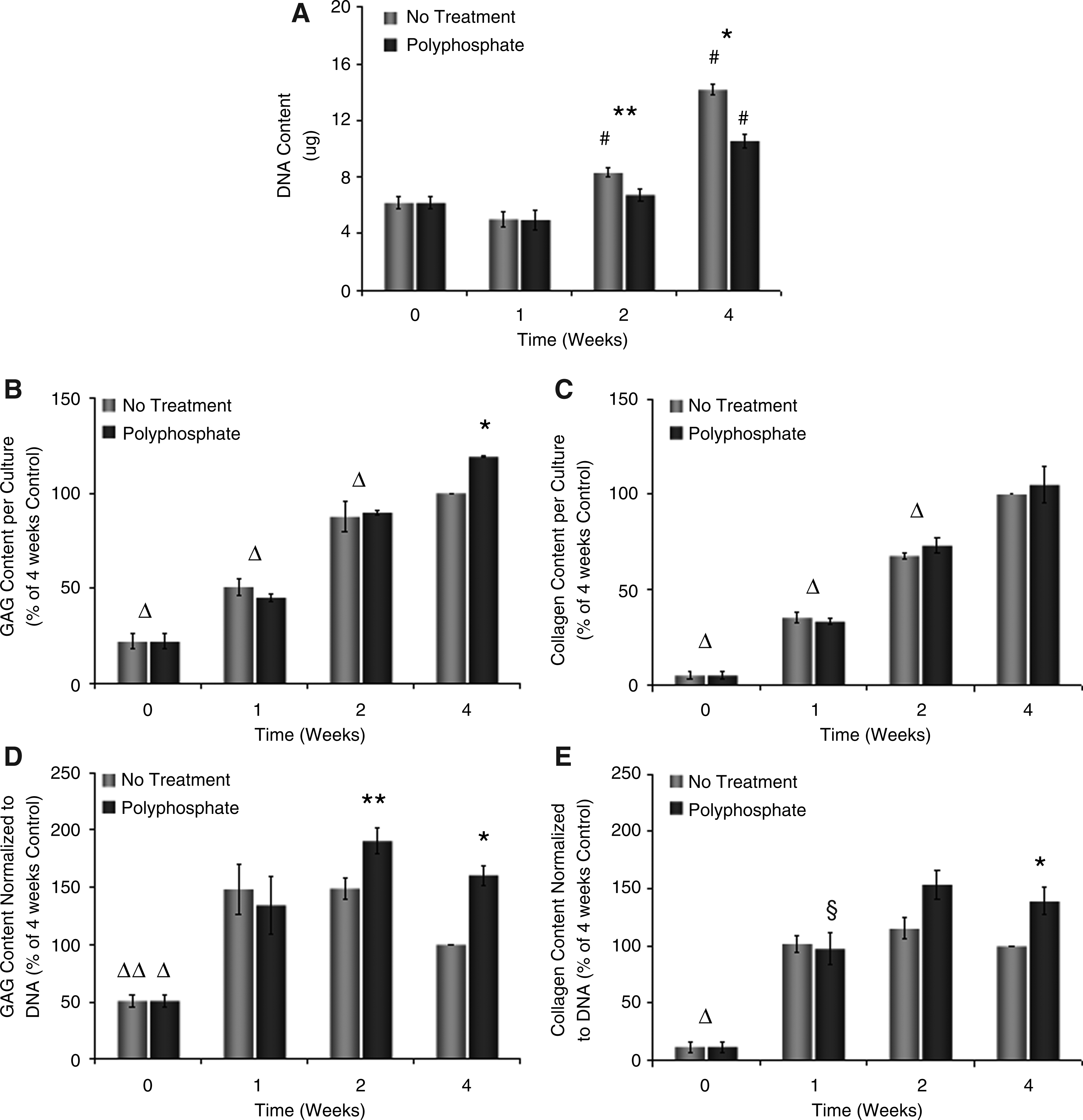
Time course of matrix and DNA accumulation within untreated and polyphosphate-treated in vitro-formed cartilage.
The stimulatory effect of inorganic polyphosphate requires continuous administration
Chondrocytes were grown in vitro on membrane inserts for 4 weeks. The medium was supplemented with 1 mM polyphosphate for the first 1 or 2 weeks of culture or for the entire 4-week period. Significant increases in GAG and collagen contents were detected only after 4 weeks of treatment, suggesting that polyphosphate must be present continuously (Fig. 5A).
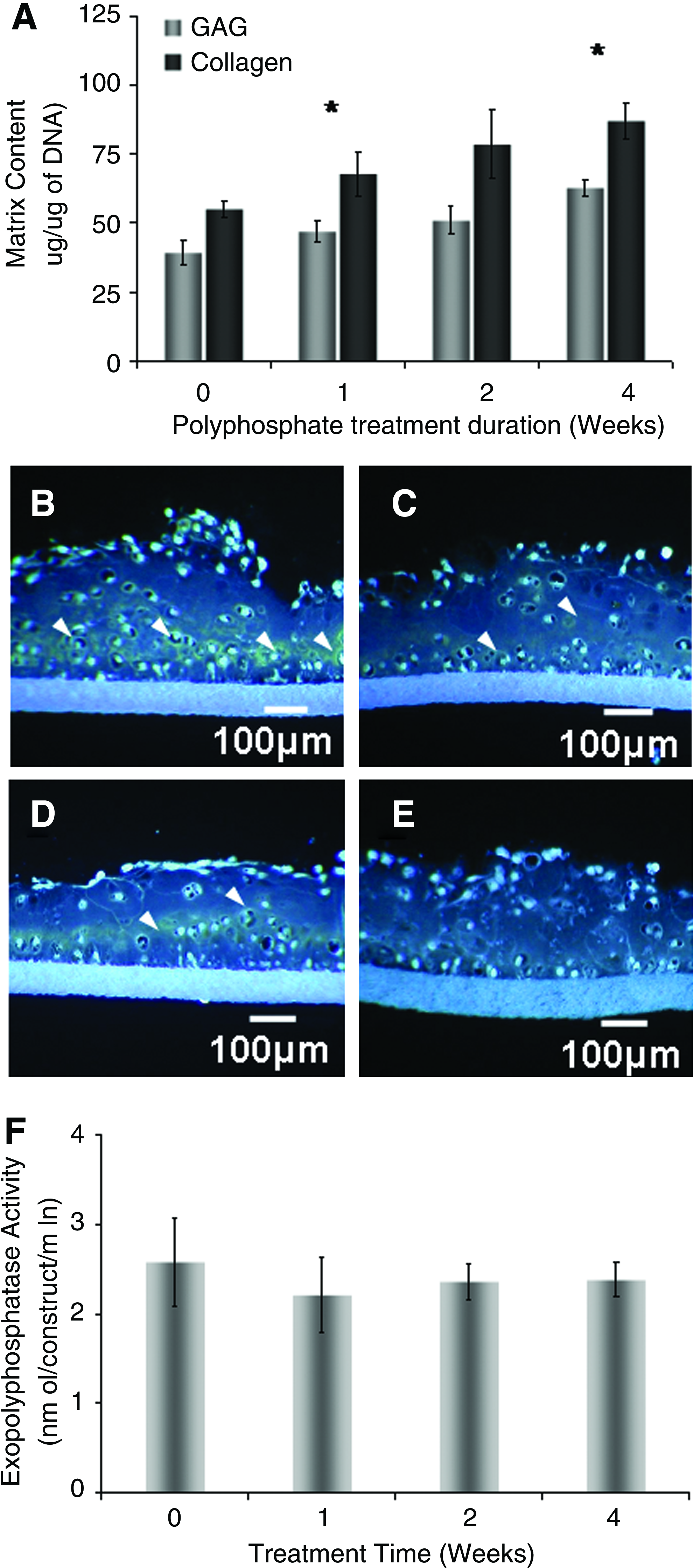
Effect of transient inorganic polyphosphate treatment on in vitro-formed cartilage matrix accumulation and the exopolyphosphatase activity.
Epifluorescence imaging of DAPI-stained tissue sections demonstrated that the polyphosphate, as indicated by the yellow-green staining, accumulates within in vitro-formed cartilage treated with condensed phosphate for 4 weeks (Fig. 5B). This accumulation was less obvious with a short-term treatment (2 or 1 weeks) (Fig. 5C, D). In contrast, nontreated cartilage exhibits the lowest levels of polyphosphate (Fig. 5E). Treatment of cartilage tissues with polyphosphate did not affect exopolyphosphatase activity levels (Fig. 5F).
Effect of polyphosphate on tissues formed by deep-zone and superficial-mid zone chondrocytes
Two chondrocyte subpopulations (superficial-mid zone and deep-zone) were grown in vitro on membrane inserts for 2 weeks in a medium supplemented with 1 mM inorganic polyphosphate and compared to zonal subpopulation tissues formed without supplementation and full-thickness chondrocytes. Figure 6 shows the DNA, GAG, and collagen contents of tissues formed by treatment of the different chondrocyte subpopulations with 1 mM polyphosphate as a percentage of the content in untreated tissues. No significant difference was observed in the responsiveness of the different subpopulations to polyphosphate, but a trend toward an increased effect in deep-zone cultures was observed for proteoglycan and collagen contents (p=0.093, 0.063, and 0.066 for DNA, GAG, and collagen contents, respectively).
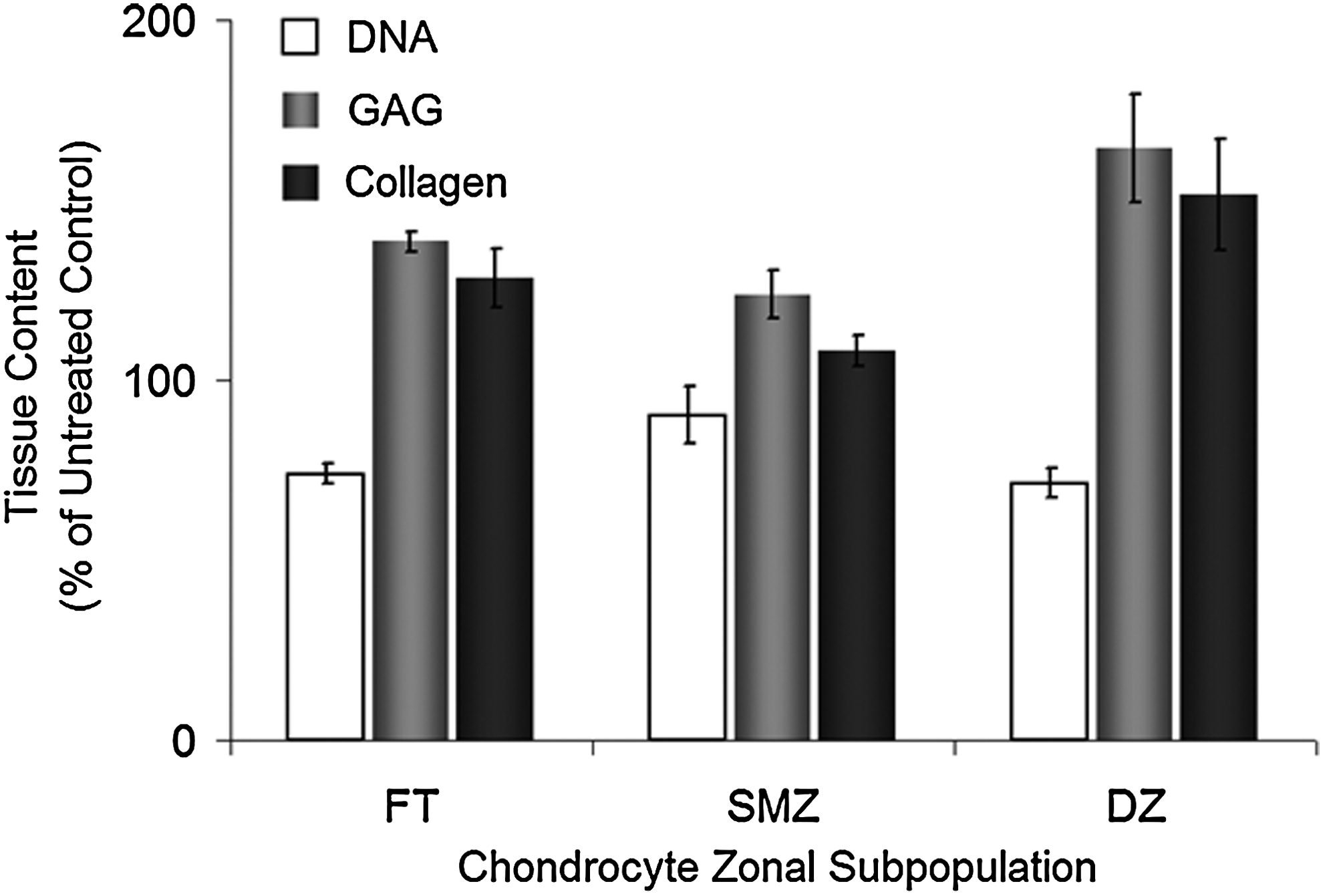
Effect of inorganic polyphosphate on the matrix content of different chondrocyte subpopulation cultures. DNA, GAG, and collagen contents of cartilage formed by full-thickness (FT), superficial-mid zone (SMZ), or deep-zone (DZ) chondrocytes cultured in vitro on membrane inserts in a medium with or without supplementation with 1 mM polyphosphate (an average chain length of 45 phosphate units) at 2 weeks. The matrix content data were normalized to DNA. All data from cultures treated with polyphosphate are expressed as a percentage of untreated cultures for each chondrocyte population. Each condition was done in triplicate, and the experiment was repeated three times. The results are from one representative experiment and expressed as mean±standard deviation.
Inorganic polyphosphate stimulates matrix accumulation in native cartilage
Native cartilage samples were obtained from full-thickness cartilage and cultured ex vivo directly in tissue culture plates for 1 week. Selected samples were treated with polyphosphate at concentrations of 1 or 2 mM. This study was performed to verify that the anabolic effects of polyphosphate were not limited to immature in vitro-formed cartilage. As is shown in Figure 7A, the GAG content of native cartilage samples was significantly increased by ∼45% after supplementation of the culture medium with 2 mM polyphosphate. The collagen content was also increased by ∼30%, but this difference was not significant. The DNA content of cartilage samples was not significantly decreased by treatment with polyphosphate (Fig. 7B).
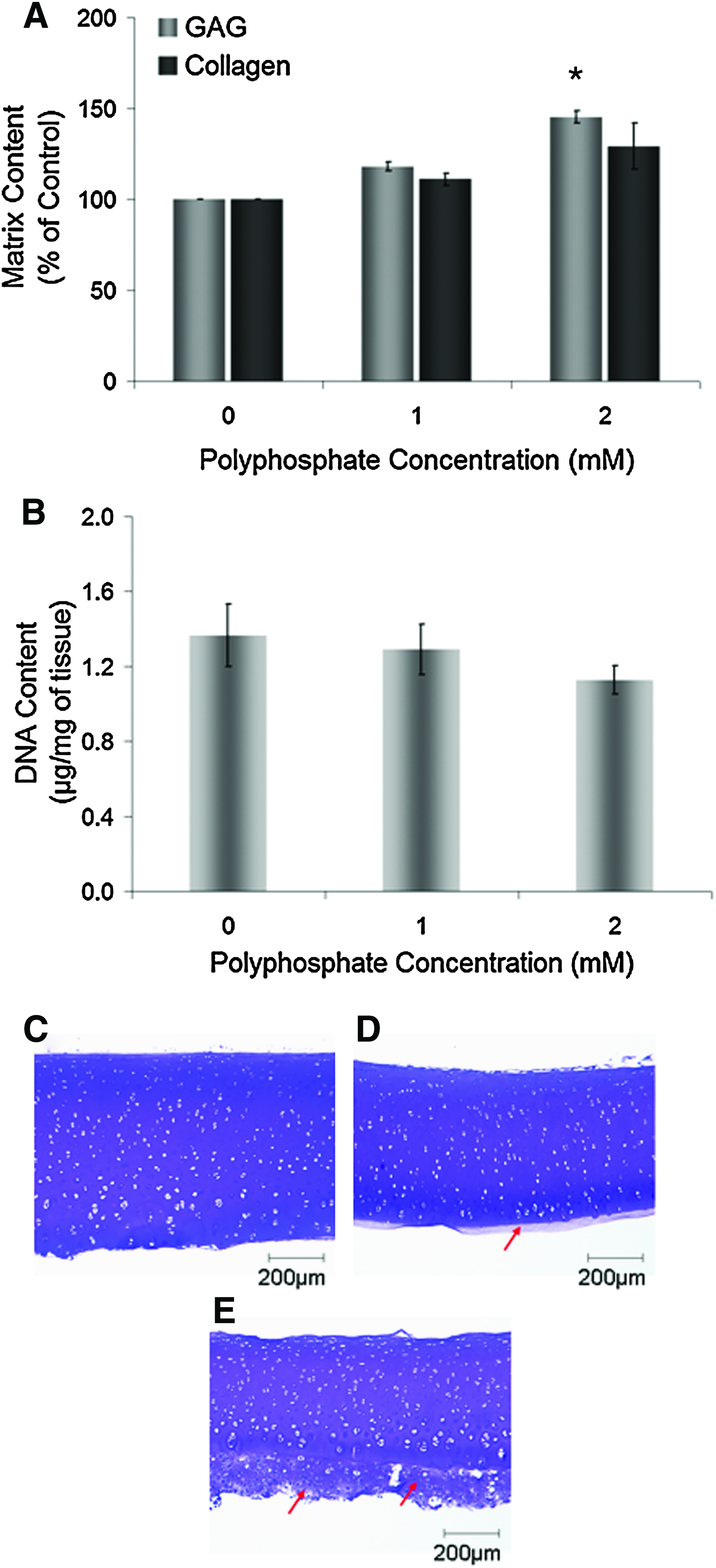
Effect of inorganic polyphosphate on articular cartilage cultured ex vivo.
Histological evaluation of ex vivo-cultured native cartilage samples stained with toluidine blue shows a new matrix that was deposited around tissues stimulated with 2 mM polyphosphate compared to nontreated cartilage (Fig. 7C–E). This matrix was deposited predominantly at the deep-zone aspect of cartilage samples.
Discussion
This study demonstrates that polyphosphate administered to tissue-engineered cartilage during in vitro growth or to native articular cartilage cultured ex vivo promotes GAG and collagen accumulation in the extracellular matrix. The anabolic effect of polyphosphate was dependent on concentration and chain length. Conversely, polyphosphate treatment resulted in a smaller increase in DNA content of in vitro-formed cartilage compared to untreated tissues. The continuous presence of condensed phosphate in the culture is essential to obtain the full stimulatory effects on matrix accumulation, as cartilage expresses exopolyphosphatases that degrade polyphosphate. These results suggest that polyphosphate can be used to stimulate the accumulation of extracellular matrix components within in vitro-formed cartilage.
To the best of our knowledge, this is the first report of the stimulatory effect of polyphosphate on articular cartilage matrix accumulation. However, polyphosphate has been shown to exhibit anabolic effects on bone. Hacchou et al. (2007) demonstrated that polyphosphate treatment enhanced bone regeneration in alveolar bone defects. 46 Similarly, porous calcium hydroxyapatite scaffolds containing adsorbed inorganic polyphosphate induced increased new bone regeneration compared to uncoated scaffolds in a rabbit femoral defect model. 47 While the mechanism by which polyphosphate stimulates cartilage matrix deposition has not yet been determined, it may be acting by stabilizing endogenous growth factors and perhaps presenting them to cell surface receptors in an appropriate conformation as has been shown to occur for FGF-2. 33 Interestingly, treatment of in vitro-formed cartilage with polyphosphate did not lead to the upregulation of cartilage matrix genes aggrecan and collagen type II, nor did it affect the gene expression of Sox9 and collagen type I. While it is possible that gene expression changes occur at other time points than those investigated, these results suggest that the effects of inorganic polyphosphate on cartilage matrix accumulation may be independent of the regulation of the expression of the main cartilage matrix genes. Further investigations are required to elucidate the mechanisms by which polyphosphate stimulates the accumulation of cartilage matrix.
While increases in GAG and collagen contents were observed in cartilage tissues treated with polyphosphates of an average chain length of 5, 45, and 75 phosphate units, only polyphosphate with an average chain length of 45 phosphate units exhibited a significant increase in matrix accumulation over nontreated tissue. Similarly, our group has shown that polyphosphate with an average chain length of 45 phosphate units was optimal for inhibition of cartilage calcification. 35 The chain length specificity was not unexpected and has been shown by other groups as well. For example, short-chain polyphosphates, including pyrophosphate, are less potent inhibitors of mineralization than longer chain polyphosphates.19,24 Short-chain polyphosphates (5 phosphate units) did not stimulate mTor activity in mammary cancer cells, whereas polyphosphate chains with 15 or more phosphate units exhibited an increased activity. 34
Our data suggest that there may be faster removal of short-chain polyphosphates, as a significantly higher exopolyphosphatase activity was detected in protein extracts incubated with short-chain polyphosphate (5 phosphate units) compared to longer chain polyphosphates (45 and 75 phosphate units). This could explain the weaker effects of short-chain polyphosphate on cartilage matrix deposition. As for the discrepancy observed between the anabolic effects of polyphosphates with chain lengths of 45 and 75 phosphate units and their associated exopolyphosphatase activities, this may be attributed to the normalization of polyphosphate concentrations on a phosphate basis resulting in a lower effective condensed phosphate concentration for longer chain polyphosphates. However, this potential explanation conflicts with the observations of Hernandez-Ruiz et al. (2006), who demonstrated that polyphosphate with a chain length of 75 phosphate units lowers the survival rate of U266 myeloma cells significantly compared to those treated with polyphosphate characterized by 45 and 25 phosphate units 48 and supports the possibility of a chain-length specificity on the biological functions of polyphosphates.
Interestingly, treatment with inorganic polyphosphate resulted in an increase in the DNA content of the in vitro-formed cartilage over time, but at a much slower rate than in nontreated tissues, suggesting that polyphosphate inhibits chondrocyte proliferation in our system. However, polyphosphate has been shown to stimulate proliferation in human fibroblasts and a murine preosteoblastic cell line through the FGF signaling pathway 33 as well as in mammary cancer cells through the mTor signaling pathway. 34 This discrepancy may be due to cell-type-specific effects. Alternatively, polyphosphate may have an effect on chondrocyte apoptosis, while the mitogenic activity remains unaffected; however, this seems less likely as the treated tissues did not appear to show greater apoptosis histologically.
The extent of cartilage matrix accumulation is directly related to the length of the culture period in which the in vitro-formed tissues are exposed to polyphosphate. Our group and others have demonstrated that the action of exopolyphosphatases, with the ability to cleave phosphoanhydride bonds of the condensed phosphates, along with hydrolysis can lead to a rapid loss of the effects of polyphosphate.35,49,50 In this study, polyphosphate accumulation was much more prominent in tissues that were treated with the condensed polymers for the entire culture period compared to shorter times (1 or 2 weeks) corroborating these results. Surprisingly, the exopolyphosphatase activity of cartilage formed by full-thickness chondrocytes is not significantly affected by treatment with polyphosphate. This is in contrast with our observations of deep-zone chondrocytes that showed significantly increased exopolyphosphatase activity when treated with condensed phosphate. 35
Polyphosphate treatment of native cartilage samples ex vivo showed increased GAG and collagen contents compared to untreated controls, supporting results obtained with in vitro-formed cartilage. Histological evaluation of the native cartilage tissues cultured ex vivo showed that new matrix accumulation was deposited predominantly on the deep-zone cartilage. This is in keeping with the possible increased responsiveness of deep-zone chondrocytes to polyphosphate treatment in vitro compared to superficial-mid zone chondrocytes. This potential differential response of chondrocytes subpopulations to inorganic polyphosphate may provide a useful system to understand the pathways by which condensed phosphate stimulates cartilage matrix deposition.
In summary, this study demonstrates that inorganic polyphosphate stimulates cartilage matrix accumulation in both in vitro-formed cartilage and ex vivo-cultured native cartilage. Based on these results, polyphosphate treatment may be useful to improve the quality of tissue-engineered cartilage.
Footnotes
Acknowledgments
This work was supported by CIHR and an NSERC fellowship and University of Toronto Open scholarship to J.P.S. The authors thank Dr. Ian Rogers, Dr. Sidney Omelon, Shawn Chua, and Ryszard Bielecki for their support. We thank Harry Bojarski and Ryding-Regency Meat Packers for providing tissues that allowed us to perform these studies.
Disclosure Statement
No competing financial interests exist.