
Abstract
Three-dimensional (3D) collagen hydrogels have been extensively used for cell culture experiments and are more closely representative of in vivo conditions than monolayer (2D) culture. Here we cultured rat bone marrow-derived mesenchymal stem cells (MSCs) in collagen hydrogels containing varying concentrations of basic fibroblast growth factor (bFGF) to examine the effect of bFGF on MSC proliferation and osteogenic differentiation in 3D culture. The optimal bFGF concentration that promoted the greatest degree of cell proliferation and expression of the early osteogenic induction marker alkaline phosphatase was also determined. Subsequent quantitative real-time polymerase chain reaction analysis of gene expression demonstrated that bFGF promoted significant upregulation of the bone-related genes: collagen type I, osteopontin (OPN), bone sialoprotein (BSP), and osteocalcin (OCN) for periods of up to 21 days. Immunofluorescence staining and fluorescence-activated cell sorting analysis further supported the enhanced osteogenic differentiation of cells as a greater proportion of cells were found to express OPN. Matrix mineralization within the collagen hydrogels was enhanced in the presence of bFGF, as assessed by calcium detection using von Kossa staining. These results clearly demonstrate a positive effect of bFGF on proliferation and osteogenic induction of MSCs in 3D collagen hydrogels when applied at the appropriate concentration. Moreover, collagen hydrogel constructs containing MSCs and appropriate growth factor stimulus might be a potentially useful biological tool for 3D bone tissue engineering.
Introduction
Hydrogels are a currently favored choice of scaffold for tissue engineering, as their water content can be controlled to support cells and maintain their migration, growth, and even differentiation into specific tissue types.9,10 Among the hydrogels available now, those composed of collagen have gained greatest interest. 11 As the most abundant constituent of natural extracellular matrices of tissues, collagen has been used for various biomaterials applications, including wound dressings and temporary scaffolds for tissue engineering.12–14 As the major organic component of bone, collagen (mainly type I) provides a structural framework resisting tensile forces, and plays a dominant role in mineralization process, leading to the formation of new bone. 15
Adult stem cells, particularly mesenchymal stem cells (MSCs) that have the capacity to undergo osteogenic differentiation, are a favored source of cells for bone tissue engineering, as they can be sourced with relative ease and can be expanded to achieve the desired quantity in vitro. 16 Stem cell behavior has frequently been characterized in 2D culture but less-so using 3D collagen hydrogel culture models. However, studies have explored methods that incorporate MSCs into 3D collagen hydrogels for prospective tissue engineering of cartilage, 17 tendon, 18 cruciate ligament, 19 and bone. 20 When MSCs are used for tendon repair strategies, ectopic bone formation is seen, 21 suggesting that MSCs placed in a 3D environment will preferentially undergo osteogenic induction. Furthermore, improved osteogenic differentiation of MSCs is achieved when they are cultured in 3D collagen hydrogels compared with monolayer culture on plastic21–23 and this is not always the case when planar collagen scaffold material is used as a monolayer growth substrate. 24 Subsequent studies have demonstrated that MSCs retain multilineage potential in 3D collagen hydrogels and can equally undergo chrondrogenic or adipogenic differentiation in addition to osteogenic differentiation if given the appropriate chemical cues.25,26
Defined osteogenic medium containing dexamethasone, β-glycerophosphate, and ascorbic acid has routinely been used to promote MSC differentiation along the osteoblast lineage for many years.27–30 In addition, soluble factors such as growth factors that can regulate bone regeneration are potential candidates for enhancement of osteogenic differentiation. For instance, basic fibroblast growth factor (bFGF) promotes increased osteoblast proliferation in monolayer31–33 and in organ culture models and leads to enhanced bone formation.34,35 The transforming growth factor β superfamily member, bone morphogenetic protein 2 (BMP2), directly promotes induction of an osteogenic differentiation36,37 by upregulating markers of mature osteoblasts such as type I collagen (Col I), osteocalcin (OCN), and alkaline phosphatase (ALP) activity.38,39 Recently, collagen hydrogels containing MSCs overexpressing recombinant BMP2 have proven very effective at repairing full-thickness cranial defects in a miniature pig model, 40 and a direct comparison of bFGF versus BMP2 suggested also that bFGF is a potent growth factor to promote osteogenesis.
Therefore, in this study we report the effects of bFGF on osteogenic differentiation of MSCs in collagen hydrogels to exploit 3D culture conditions, which better represents the natural in vivo tissue environment. The proliferation of MSCs and their osteogenesis within the collagen hydrogels, including ALP activity, gene expression, protein synthesis, and mineralization in vitro, were investigated with the stimulus of bFGF loaded within the hydrogels.
Materials and Methods
Recombinant bFGF
As described elsewhere, 41 recombinant bFGF was constructed. In brief, the complementary deoxyribonucleic acids (cDNAs) of the bFGF were amplified from an adult human cDNA library. A pair of primers, 5′-CGAGATCTCAGCCGGGAGCATCAC-3′ and 5′-TGCAGATCTCGCTCTTAGCAGACATTG-3′, was designed according to the GenBank™ sequence (GenBank accession number NM002006). The polymerase chain reaction (PCR) products were cloned into pBAD/His A (Invitrogen) in-frame using the NH2-terminal 6× His tag. The recombinant bFGF protein containing the poly-His tag was expressed and purified using a Ni2+ affinity column under denaturing conditions according to the manufacturer's protocol (Invitrogen).
Preparation of collagen hydrogels
Collagen 3D matrix for the culture of cells was prepared by mixing the collagen solution (3.45 mg/mL, rat tail Col I; BD Science) with a mixture solution of 1N NaOH, α-minimum essential medium (α-MEM; 10×; Gibco), and distilled water. The incorporation of NaOH neutralized the mixture to pH 7.4 and the addition of 10× concentrated α-MEM provided an appropriate nutritional environment for the culture of cells. The final concentration of collagen was determined to be 2.277 mg/mL.
When placed in an incubator under a humidified atmosphere of 5% CO2 in air at 37°C for 30 min, the solution was allowed to solidify into a collagen hydrogel matrix. When needed the solutions of cells (50 μL/mL) and bFGF (100 μL/mL) prepared were also added to the mixture of collagen solution before gelation.
MSC isolation and culturing
MSCs were isolated from bone marrow of male adult Sprague-Dawley rats (4–8 weeks old). 42 All protocols involving animals were approved by the Animal Care and Use Committee of Dankook University. Briefly, from the marrow cavities of femora and tibiae bones of rats, bone marrow tissue was removed and then flushed with dispase II and type I collagenase solution in α-MEM. The tissue was centrifuged and resuspended in the standard culture medium (α-MEM supplemented with 10% fetal bovine serum containing 1% penicillin/streptomycin) and then placed in a culture dish, which was then cultured in an incubator under humidified atmosphere of 5% CO2 in air at 37°C. Nonadherent hematopoietic cells were removed from the medium in the course of medium change, and the MSCs underwent passages of two to three were used for further experiments.
Incorporation of bFGF into collagen hydrogel and the release profile
To observe the effects of bFGF on cell behavior, different concentrations of bFGF (0.01, 0.1, 0.5, 1 and 10 μg/mL) were prepared in the culture medium, and then a 100 μL aliquot of the bFGF solutions was supplemented into the mixture of collagen solution before seeding cells. The cell–collagen solutions either containing bFGF or not were incubated at 37°C for 30 min to promote hydrogel formation. Accordingly, the final quantities of bFGF incorporated into each 500 μL hydrogel matrix were 0.5, 5, 25, 50, and 500 ng.
The release profile of bFGF incorporated within the collagen hydrogel matrix was monitored for up to 30 days. For this test, the hydrogels incorporating bFGF at two different concentrations (1 and 2.5 μg) were used. Each hydrogel was placed in culture medium within an incubator at 37°C, and at the predetermined time points, the supernatant was gathered and the bFGF quantity was assessed using an enzyme-linked immunosorbent assay (ELISA) kit (human FGF-basic ELISA development kit, Peprotech) according to the manufacturer's instructions. After the enzymatic reaction, the absorbance recorded at 405 nm was used to determine the quantity of bFGF released referring to the standard curve obtained in the range of 0–4 ng/mL. Based on this, the remaining bFGF within the hydrogel matrices was also assumed.
Cell culture within collagen 3D hydrogel containing bFGF
For the culture of cells within the 3D collagen hydrogel matrix, a 50 μL aliquot of MSCs prepared at varying concentrations (5×104, 1×105, 2×105, and 5×105 cells/mL) were combined and mixed with the collagen solution (either containing bFGF or not). About 500 μL of the cell–collagen solution was plated into each well of 24-well plates. Therefore, the final cell numbers contained within each hydrogel sample varied (2.5×103, 5×103, 1×104, and 2.5×104 cells). The behavior of cell-seeded collagen hydrogels, for example, the contraction with culture time, was monitored for up to 6 weeks. Based on this, the collagen hydrogel matrices loaded with a large number of cells (2.5×104) were shown to undergo substantial contractions (50%–60%) after 3–4 weeks, but those with less cells (2.5×103, 5×103, and 1×104 cells) showed gel contractions either trivial or much less (<10%) after 3–4 weeks, although after 6 weeks those with 5×103 and 1×104 cells also showed high contractions (∼30%–40%). Therefore, we determined the cell culture conditions at the points where the gel contraction was not so significant; cell numbers of 2.5×103, 5×103, and 1×104 and culture periods of up to 3–4 weeks.
The cell-containing hydrogel matrices were cultured with osteogenic medium (standard culture medium plus 50 μg/mL ascorbic acid, 10 mM β-glycerol phosphate, and 0.1 μM dexamethasone) in humidified atmosphere of 5% CO2 in air at 37°C.
Cell proliferation assay
During the culturing, the cell growth kinetics within the collagen hydrogel matrices containing different concentrations of bFGF (0.5, 5, 25, 50, and 500 ng) was determined. At predetermined time points (3, 7, and 14 days), cells were gathered from the collagen hydrogel by means of digesting the collagen hydrogel with type I collagenase (10 mg/mL) and then centrifugation (1500 rpm). Cell proliferation was assessed using an MTS (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium) assay kit (CellTiter 96 Aqueous One Solution from Promega) at an absorbance 490 nm using a spectrophotometer. For the proliferation study, the quantity of cells loaded in the collagen hydrogel was initially varied (2.5×103, 5×103, and 1×104).
Cell morphology observation
The cell morphology within the collagen hydrogels was observed by means of confocal laser scanning microscopy (CLSM; Zeiss M700, Carl ZEISS). The constructs of cell–collagen hydrogel were fixed with 3.7% formaldehyde in phosphate buffered saline (PBS) for 10 min and permeated with 0.1% Triton X-100 in PBS for 1 min. Cells were then incubated in the solution of Alexa Fluor 546 phalloidin (Invitrogen A22283) and 4′,6-diamidino-2-phenylindole (DAPI; Invitrogen P36935) for 1 h, and then washed with distilled water. The actin rings formed and nuclei were revealed under the microscope.
Histological observation of the cells was also made by cryopreservation and sectioning. The constructs of cell–collagen hydrogel were embedded in an optimal cutting temperature compound and snap-frozen in liquid nitrogen at −196°C. Frozen sections with a thickness of ∼10 μm were obtained using a Cryostat (Leitz 1720, Emst Leitz, Wetzlar GmbH), which were then mounted on slide glass, and stained with hematoxylin and eosin (H&E). The stained cells were observed by Olympus IX71 optical microscope (Japan).
ALP determination
To determine the initial osteogenic development of the MSCs within the bFGF-containing collagen hydrogel matrices the ALP activity of the cells was measured. After culturing for 7 and 14 days, cells were gathered from the hydrogel matrices by a digestion with collagenase (for Col I) and then centrifugation. The cell pellets were added with lysis buffer and disrupted by the freezing/thawing processes. Each sample was added to the ALP reaction media to allow an enzymatic reaction according to the manufacturer's instruction (Sigma). The quantity of sample addition was determined based on the total protein content, which was determined by a commercial DC protein assay kit (from BioRad). The product p-nitrophenol appeared as the color change, which was measured at an absorbance of 405 nm using a spectrophotometer. Five replicate samples were used at each condition (n=6).
Gene expression by quantitative real-time PCR (QPCR)
Expression of genes associated with osteogenic differentiation, including Col I, osteopontin (OPN), bone sialoprotein (BSP), and osteocalcin (OCN), was assessed by means of QPCR. Cells cultured for 7, 14, and 21 days were gathered by a digestion of collagen hydrogel with collagenase and then centrifugation. The cell pellets were homogenized with guanidine isothiocyanate-based cell lysis buffer (Qiagen Ltd.). Total ribonucleic acid (RNA) was extracted from the lysate using RNeasy Mini Kit (Qiagen Ltd.). RNA was reverse transcribed to cDNA using the Superscript kit (Invitrogen) with random hexamers. QPCR was performed using Sensimix Plus SYBR master mix (Quantace) in a spectrofluorometric thermal cycler (Rotor-Gene 3000; Corbett Research). The comparative threshold cycle (CT) method was used in which the accumulated PCR products for each gene (Col I, OPN, BSP, and OCN) examined were normalized to the reference gene β-actin. Triplicate samples were tested. The primer sequences of the genes for QPCR are presented in Table 1.
Immunofluorescence staining and fluorescence-activated cell sorting analysis
The MSCs expressed osteogenic marker OPN were immunofluorescence-stained. For the immunostaining, the MSCs cultured within the collagen hydrogels either containing bFGF or not for 14 and 21 days were fixed with 4% paraformaldehyde for 5 min, washed with PBS, and dried. Samples were treated with 0.3% triton-x100 and blocked with 2% bovine serum albumin, and then incubated with primary antibody against OPN (anti-rabbit. 1:100) at 4°C overnight. Double-labeled immunostaining was done with appropriate anti-rabbit fluorescein isothiocyanate (FITC)-conjugated secondary antibodies. The OPN stains were revealed in tandem with the stains of F-actins and cell nuclei by using Alexa Fluor 546 phalloidin and DAPI, respectively. Images were captured using a Zeiss M700 confocal microscope.
To better confirm the cellular fraction that expressed OPN, fluorescence-activated cell sorting (FACS) analysis was carried out. Cells grown for 14 and 21 days were gathered from the collagen hydrogel matrices, and then washed with PBS. The harvest cells were fixed with 4% paraformaldehyde and then treated with 0.2% triton ×100. Five percent bovine serum albumin (BSA) was used to block the sample and the cells were probed with primary antibodies specific for OPN. The cells were then incubated with secondary antibodies conjugated to FITC (Santa Cruz Biotechnology, Inc.). Data were acquired for at least 10000 cells using BD FACS caliber flow cytometry system (BD Biosciences).
Western blot analysis
The extracts of the harvested cells were prepared with radio-immunoprecipitation assay (RIPA) buffer, after which protein samples were resolved on 10% sodium dodecyl sulfate–polyacrylamide gels, transblotted to nitrocellulose membranes blocked with 2.5% BSA in Tris-buffered saline with 0.1% Tween-20, and probed with primary antibody (anti-OPN antibody, 1:1000). The blots were then incubated with horseradish peroxidase-conjugated secondary IgG, and immunoreactive bands were detected using ECL detection reagent (Pierce).
Mineralization
Cellular mineralization was determined by staining the calcium deposition with von Kossa stain and data were quantified by the densitometric analyses. For the von Kossa sliver nitrate staining assay, the cell–collagen hydrogel constructs at day 28 were fixed with 10% formalin for 10 min. After rinsing, the fixed slides were incubated with 5% sliver nitrate solution under UV light for 30 min. After the reaction the calcium deposition was revealed as black spots. Samples were rinsed fully with distilled water and 5% sodium thiosulfate. The relative area of the deposits (mean gray values of deposits per total area) was quantified with densitometric analyses (NIH image 1.61 software).
Statistics
Data were represented as mean±one standard deviation. Statistical difference was analyzed by one-way analysis of variance, and p-values of <0.05 and <0.01 were considered significant.
Results
Characteristics of collagen hydrogels containing cells
We initially seeded various concentrations of cells within the collagen hydrogels to determine the appropriate number of cells that can be used without resulting contraction and remodeling of collagen hydrogels occurring. Collagen hydrogels containing various quantities of MSCs (from 1×103 to 2.5×104 cells per collagen hydrogel matrix) were cultured for periods up to 6 weeks and visually assessed to identify the hydrogel contraction. Representative macroscopic images at certain time points are presented in Figure 1. There were no significant changes in the size of all the hydrogels during the first 2 weeks. However, substantial contractions (∼50%–60%) were noticed at 3–4 weeks when the cell seeding number was high (2.5×104). Moreover, much longer culture period (6 weeks) also resulted in some contractions (∼20%–30%) of the hydrogels even with lower cell numbers. Taking these hydrogel contractions into account, we determined the culture conditions for subsequent experiments at the points where the gel contraction was not so significant: cell numbers <2.5×104 per hydrogel and culture periods of up to 3–4 weeks.
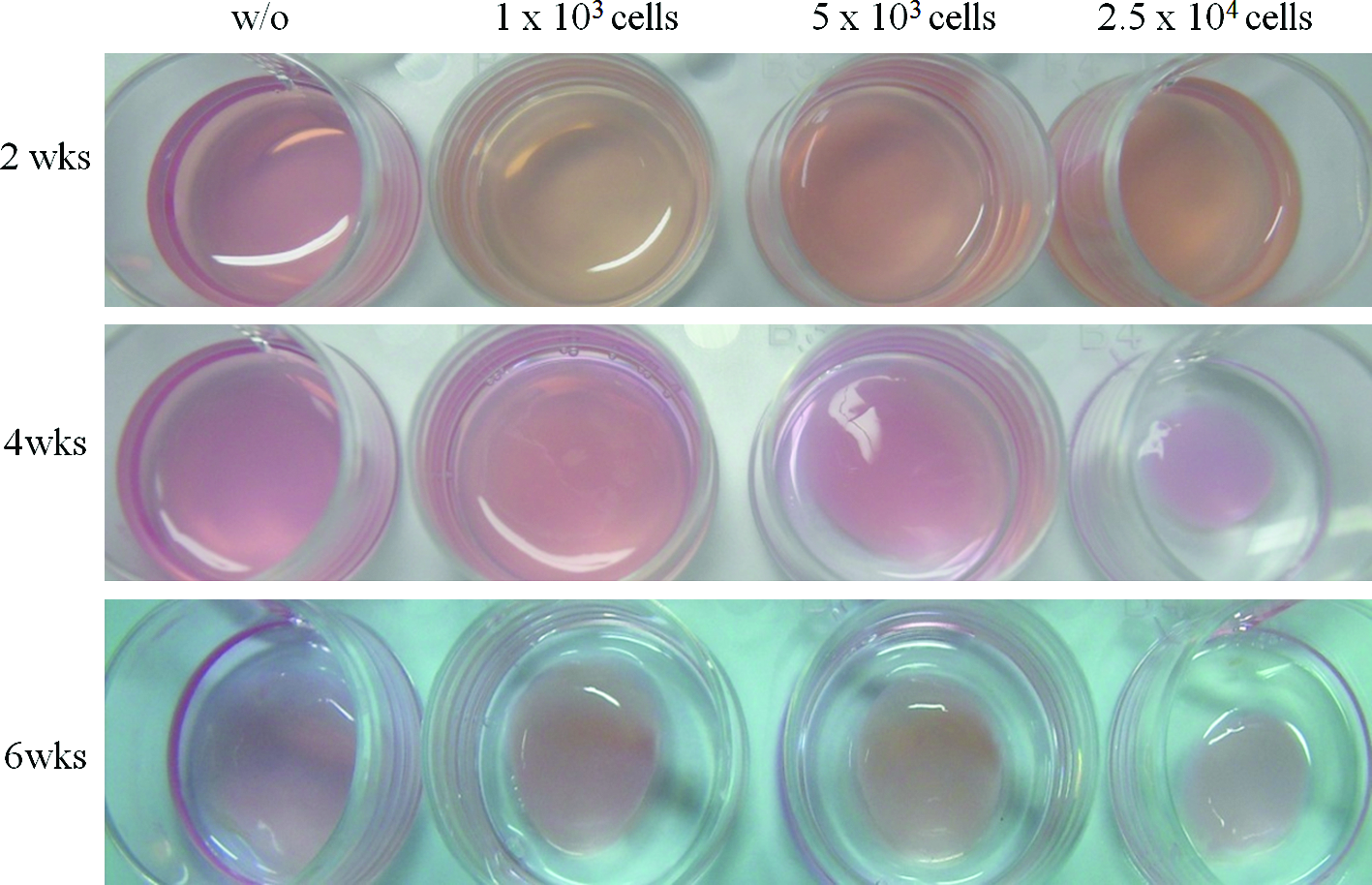
Macroscopic images of collagen hydrogels containing different seedings of MSCs (without and with 1×103, 5×103, and 2.5×104) over culturing for 2, 4, and 6 weeks. Contraction of the collagen hydrogels occurred with high cell loading quantity (2.5×104) and prolonged culture period (6 weeks). MSCs, mesenchymal stem cells. Color images available online at www.liebertonline.com/tea
Release profiles of bFGF from collagen hydrogel
The release profiles of bFGF from the collagen hydrogels were monitored, as shown in Figure 2. bFGF was first incorporated at either 1 or 2.5 μg within the hydrogel matrix, which was then incubated in a culture medium for up to 30 days. At the predetermined time points, the supernatant was gathered and assessed using an ELISA kit. Release profiles showed a stepwise pattern for both bFGF concentrations: an initial rapid and steady release within the first 7 days and then further sustained and steady release up to 30 days. The total amounts of the released bFGF from the hydrogels were found to be about 76.3% and 51.9% of the initial amounts (1 and 2.5 μg), respectively.

Release profiles of bFGF incorporated within the collagen hydrogel matrices. The collagen hydorgel incorporating bFGF of either 1 or 2.5 μg was placed in a culture medium, and the supernatant was assessed by an ELISA kit to detect the released amount of bFGF. Release profiles show a stepwise pattern, namely, relatively rapid but steady release within the first 7 days and a further sustained release up to 30 days. The final released amounts of bFGF measured were ∼76.3% and 51.9% of the initials (1 and 2.5 μg), respectively. bFGF, basic fibroblast growth factor.
Effects of bFGF on the MSC proliferation within collagen hydrogel
In the next set of experiments we determined the effect of bFGF on MSC proliferation within collagen hydrogels. Based on the observations in Figure 1, we seeded 1×104 MSCs into collagen hydrogels, which were loaded with varying concentrations of bFGF (broad range from 5 to 500 ng per hydrogel sample), and the effects of bFGF on the MSCs proliferative potential were assessed using the MTS viable cell assay after culturing cells for 3 days. In hydrogels containing 5 ng bFGF, cell proliferation increased significantly with respect to bFGF-free collagen hydrogels (**p<0.01; Fig. 3) and this increase appeared to maintain up to the bFGF amount of 50 ng. However, further increase in bFGF up to 500 ng resulted in a significant decrease in proliferation.
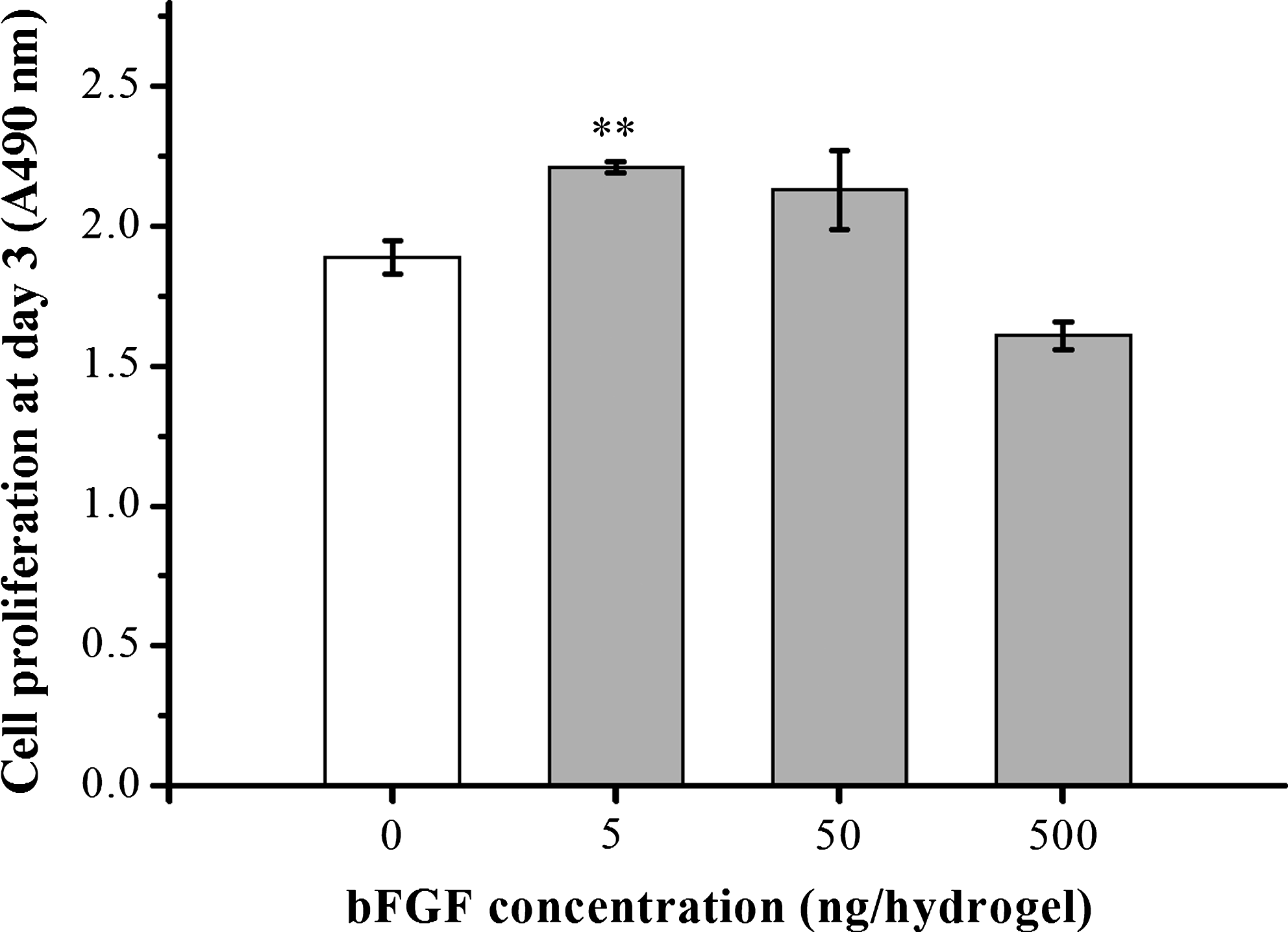
Proliferation of MSCs within collagen hydrogels after 3 days in culture in the presence of a broad range of bFGF concentrations (0, 5, 50, and 500 ng per hydrogel sample), as measured using an MTS assay. Optimal cell number was attained with 5 ng bFGF, but a decline was observed at higher concentrations (**p<0.01 without vs. with 5 ng bFGF, by ANOVA, n=3). MTS, (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium); ANOVA, analysis of variance.
More in-depth investigation into the effects of bFGF on cell proliferation was subsequently made using a narrow range of bFGF concentrations (0 to 50 ng per hydrogel). Moreover, several cell concentrations were assayed (2.5×103, 5×103, and 1×104 cells/sample). After culturing for 3 and 7 days, the cell proliferation was measured. At 3 days (Fig. 4a), increasing bFGF up to 5 ng was accompanied by a dose-dependent increase in proliferation. However, further increasing bFGF up to 50 ng led to a gradual decrease in cell proliferation. This trend was similarly observed for all cell concentrations. At day 7 (Fig. 4b), the effects of bFGF concentration on the cell proliferation were also similarly observed as to the results at day 3. Initial cell seeding density directly correlated with relative cell number obtained upon measurement of proliferation, at both time points.
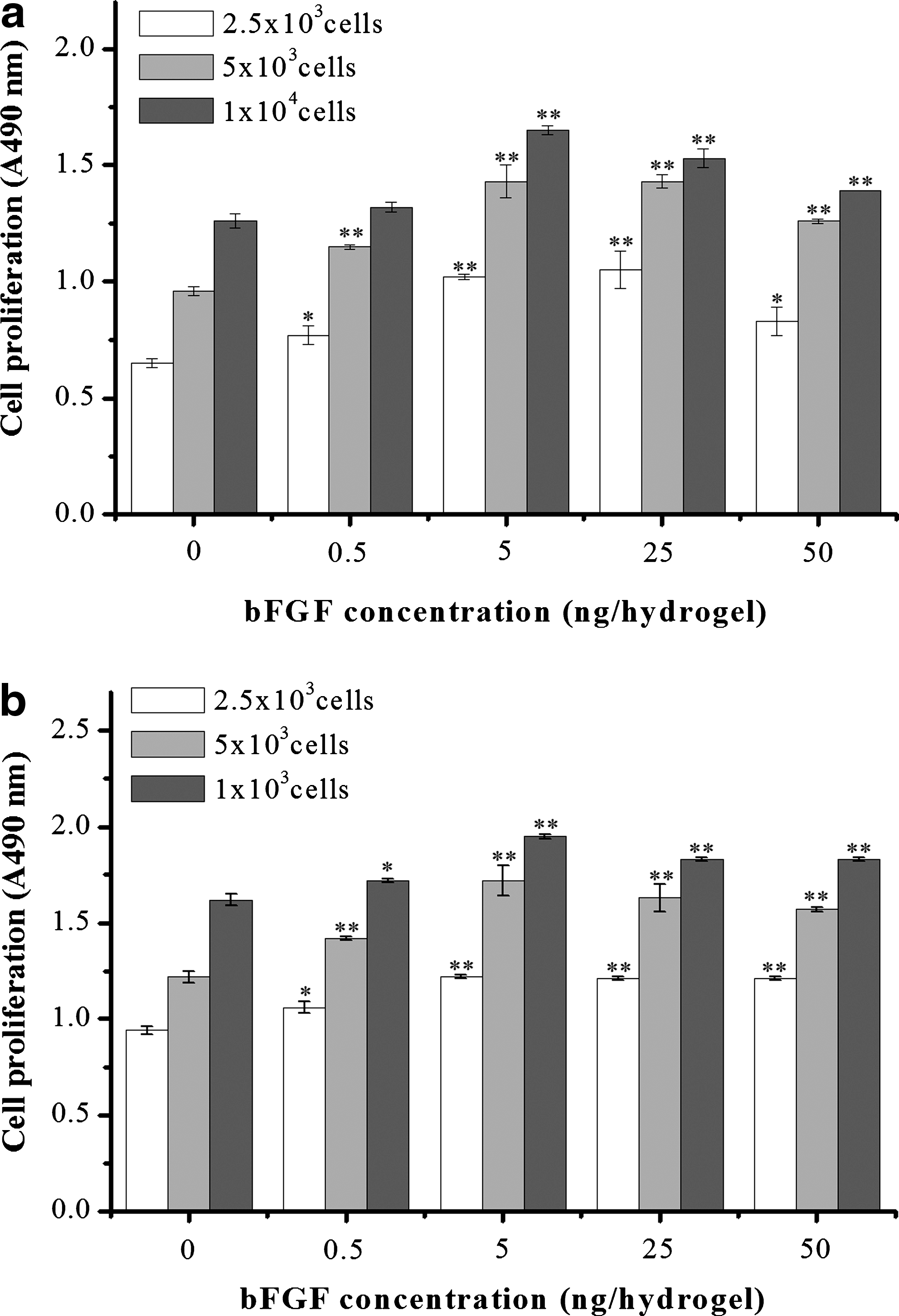
Effect of bFGF on the MSCs proliferation cultured in the collagen hydrogels after
In the last proliferation experiment, collagen hydrogels containing MSCs seeded at a quantity of 5×103 cells per hydrogel were prepared with either 5 ng bFGF or in the absence of bFGF to directly test the effect of bFGF over the time that osteogenic differentiation events usually begin to occur. Proliferation was measured at days 3, 7, and 14. At all time points, we observed significant differences in the cell proliferation (Fig. 5).

Effect of bFGF (5 ng) on the MSC proliferation cultured in collagen hydrogels up to 14 days. An increase in cell proliferation was observed at all time-points in the presence of bFGF (*p<0.05 and **p<0.01 by ANOVA, n=3).
Cell growth morphology within collagen hydrogel
The presence of cellular material was revealed by means of a histological staining. Frozen sections of collagen hydrogels obtained after 14 days of culture with or without 5 ng bFGF are presented in Figure 6. A noticeable increase in H&E staining is evident in sections of hydrogels containing bFGF (Fig. 6a), and small clusters of cells are evident (indicated as arrows) that are lacking in bFGF-free control (Fig. 6b).
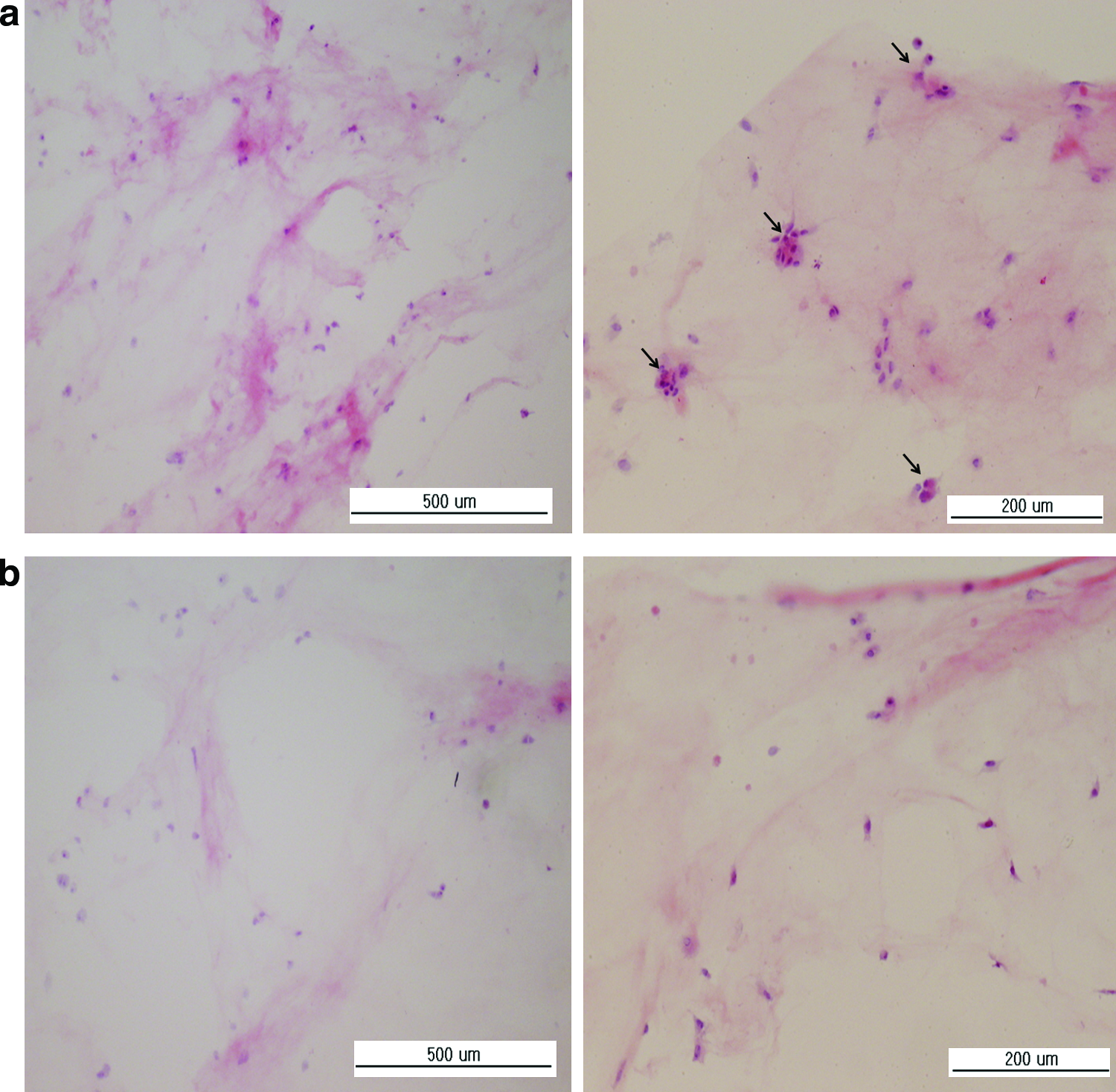
Histological appearance of collagen hydrogels containing MSCs after 14 days of culture.
Effect of bFGF on ALP production
Osteogenic differentiation of MSCs cultured in collagen hydrogels in response to bFGF was determined by detecting ALP activity of cells within the collagen hydrogels after 7 and 14 days. ALP protein level increased from day 7 to 14 both in the presence and absence of bFGF (5 ng; Fig. 7). At day 7, the ALP level in hydrogels containing bFGF was similar to those without bFGF, but by day 14, ALP was much higher in the hydrogels containing bFGF compared to those without (p<0.01) (Fig. 7).
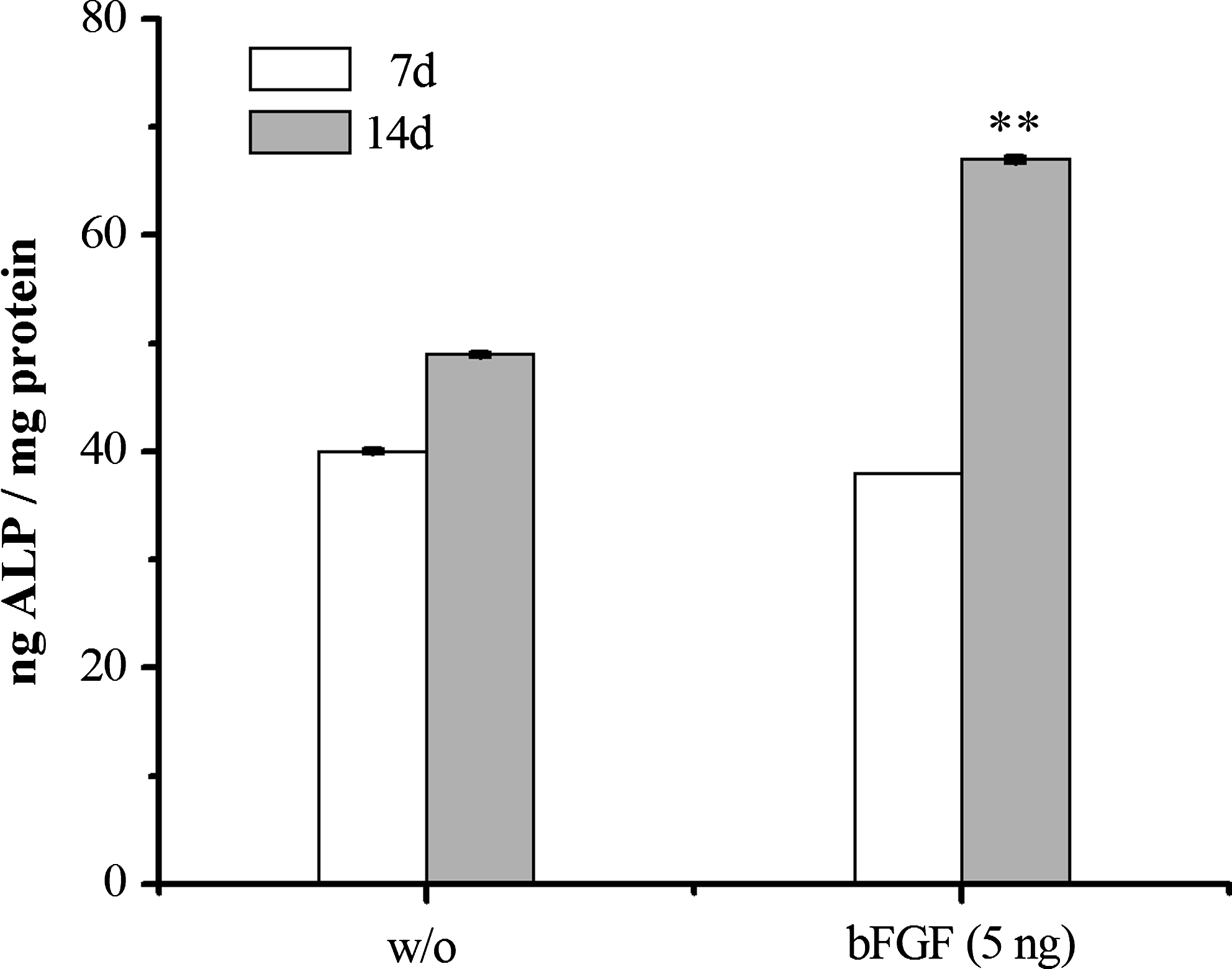
Effect of bFGF (5 ng) on the alkaline phosphatase (ALP) activity of MSCs cultured in collagen hydrogels after 7 and 14 days, demonstrating that osteogenic induction was significantly higher in the presence of bFGF (**p<0.01, by ANOVA, n=6).
QPCR analysis of osteogenic differentiation markers
We then investigated the effect of bFGF on expression of genes that are related to osteoblastic development, including Col I, OPN, BSP, and OCN. After culture periods of 7, 14, and 21 days, levels of mRNA transcripts were quantified by QPCR analysis. Col I expression increased over time and was much greater in the presence of bFGF at all time points (p<0.01 vs. time-matched controls lacking bFGF; Fig. 8a). OPN expression was significantly upregulated at day 14 in the presence of bFGF compared to the time-matched control (Fig. 8b), and this expression level was maintained until day 21, by which time the bFGF-free control also expressed a similar level of OPN. For BSP and OCN, bFGF treatment stimulated a time-dependent increase in transcript expression, with a fourfold increase in expression of both transcripts after 21 days relative to the bFGF-free control (Fig. 8c, d).
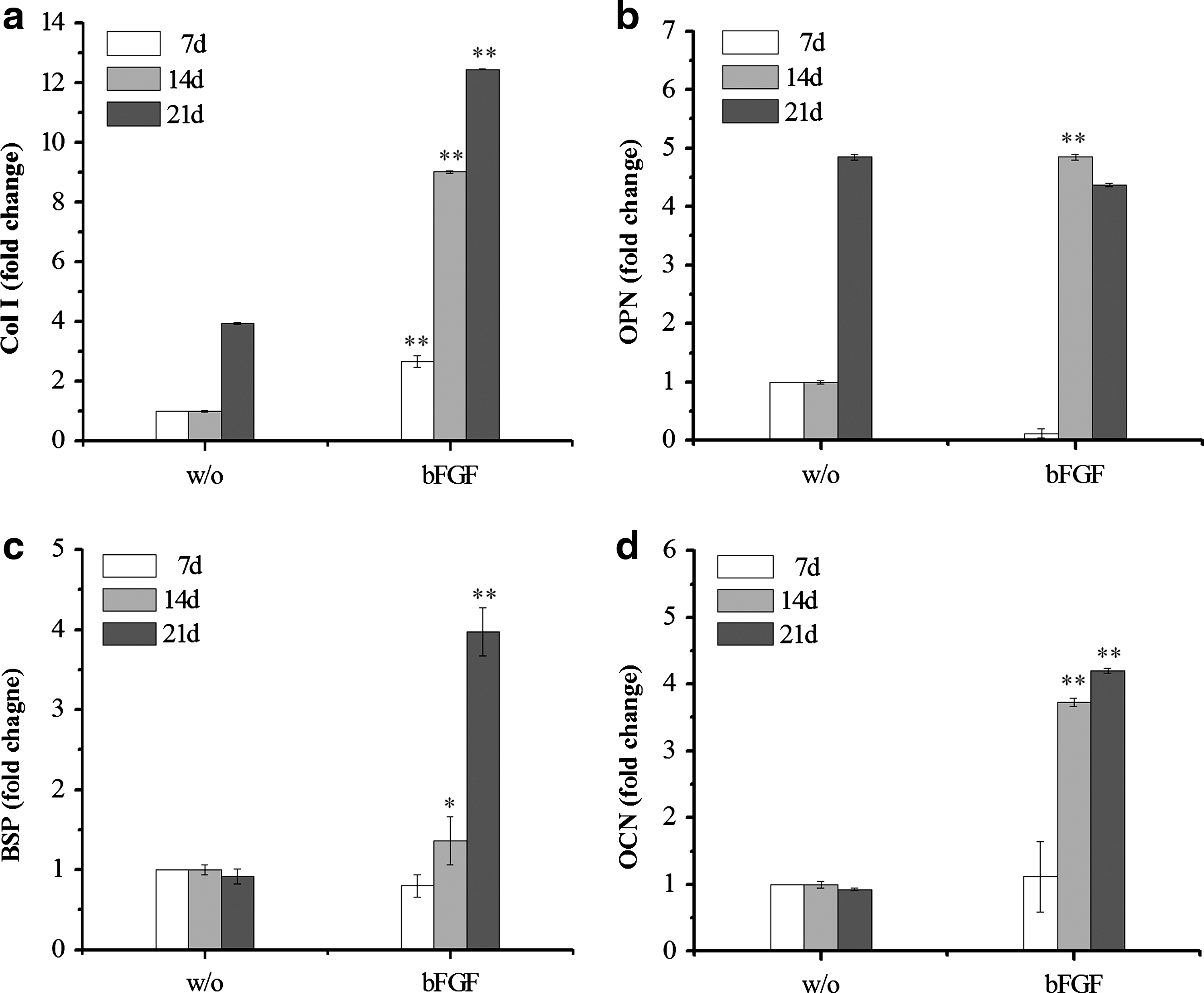
Quantitative real-time PCR analysis of the effect of bFGF (5 ng) on MSC gene expression cultured within the collagen hydrogels. The mRNA transcript levels associated with markers of osteogenic differentiation of MSCs over a culture period of 21 days were quantitatively assessed:
Immunofluorescence staining and FACS analysis
The effect of the presence of bFGF within the collagen hydrogel matrices on the osteogenic differentiation was further examined by immunofluorescence staining. In particular, expression of OPN was assessed as this was highly expressed in the QPCR result and its importance in the later stage of osteogenic differentiation. As revealed in Figure 9, MSCs cultured for 14 and 21 days were stained with OPN antibody (in green). Cytoskeleton processes (F-actins, in red) and cell nuclei (in blue) were also revealed by co-staining. Cells were observed highly viable with numerous cytoskeleton processes, and the more so with prolonged culture period. With regard to OPN stains, many cells cultured in the presence of bFGF for 14 days were shown to be stained positive for OPN, and the more so at 21 days. However, very little stain for OPN was noticed in the cells cultured without bFGF at 14 days and only a part of cells were positive for OPN at 21 days.
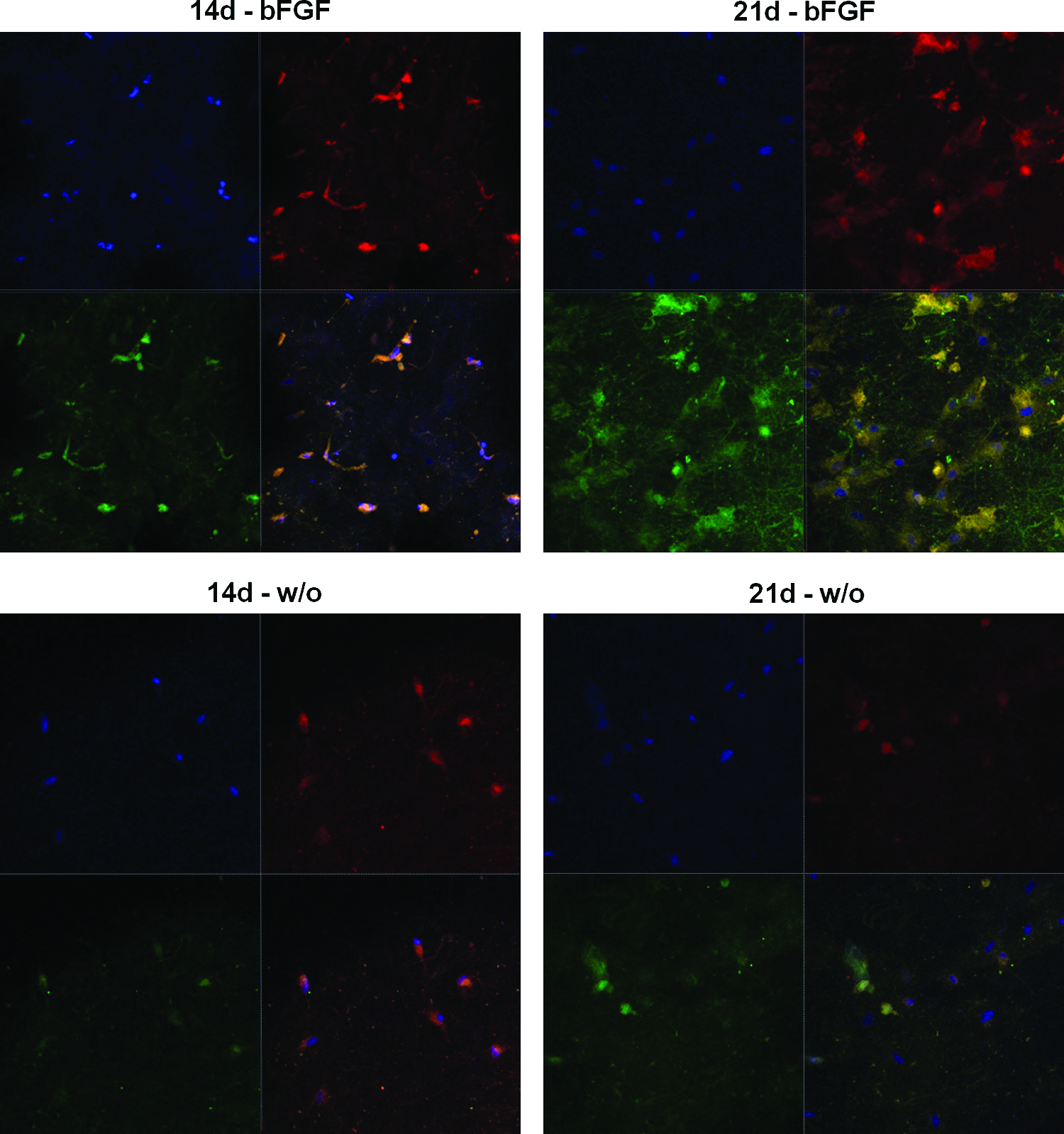
Confocal laser scanning microscopy of the MSCs immunofluorescence-stained for OPN. MSCs were grown within the collagen hydrogel either with bFGF 5 ng or without it, for periods of 14 and 21 days. After culture, cells were fixed and immunostained with antibody against OPN (in green). Co-stains were made to visualize cell nuclei (in blue) and cytoskeleton processes (F-actins, in red) with DAPI and Alexa Fluor 546 phalloidin, respectively. DAPI, 4′-6-diamidino-2-phenylindole. Color images available online at www.liebertonline.com/tea
The OPN immunofluorescence-stained cells were further quantified by the FACS, as presented in Figure 10. After culture for 14 and 21 days, cells immunofluorescent-stained for OPN were analyzed with flow cytometry system. Data were acquired for at least 10000 cells. There were significant up-regulations in the bFGF-containing hydrogel matrices at both culture periods, which was, however, not noticed in the bFGF-free hydrogels.

Fluorescence-activated cell sorting (FACS) analysis performed on the MSCs cultured for 14 and 21 days within the collagen hydrogels either containing 5 ng bFGF or not. Cells immunostained for OPN were characterized by a flow cytometry analysis system. Data are expressed as the cell number (y-axis) plotted as a function of fluorescence intensity (x-axis) and are representative of three separate experiments. Negative control cells were incubated with the secondary antibody alone.
Confirmation of protein level by western blot analysis
The protein level of OPN was further analyzed by western blotting. As shown in Figure 11, the band corresponding to OPN protein was better developed in the collagen hydrogel containing bFGF than that without bFGF at both culture periods, and the difference was much clearer at 21 days.

Western blot analysis of OPN protein expressed by the MSCs after culture for 14 and 21 days within the collagen hydrogels either containing 5 ng bFGF or not.
Cellular mineralization
Based on our observations of elevated osteogenic differentiation marker expression of MSCs in collagen hydrogels containing bFGF, we further assessed the mineralization activity of the cells. After culturing MSCs in collagen hydrogels with or without bFGF for 28 days, the mineralization was detected using the von Kossa staining method. Representative images showing the mineralized product are presented in Figure 12a. Calcium deposits stained as dark spots were observed in the hydrogel matrix. Calcium deposits were more abundantly and uniformly detected in the presence of 5 ng bFGF when compared to bFGF-free control. The formation of deposits was quantified using optical densitometry and significantly higher mineralization activity was detected in the collagen hydrogels containing bFGF, (approximately fivefold increase; p<0.01; Fig. 12b), indicating that bFGF had a significant positive impact on the osteogenic differentiation and mineralization potential of MSCs cultured in 3D collagen hydrogels.

Effect of bFGF (5 ng) on the mineralization of MSCs cultured within the collagen hydrogels during 28 days was determined using von Kossa stain.
Discussion
The importance of culturing cells in a 3D environment that is more representative of tissue architecture in vivo is widely recognized and many strategies for producing 3D scaffolds have been explored. Collagen hydrogels were first used as a 3D growth substrate for cells over 50 years, 43 and subsequent work by Bell and colleagues described the contraction profiles of collagen hydrogels seeded with fibroblasts of varying proliferative potential. 44 These studies laid the foundation for development of culture models to analyze responses of cells, particularly fibroblasts, in 3D culture to aid our understanding of wound healing and scarring.45–47 Collagen hydrogels have also been successfully employed as a scaffold for tissue-engineered skin that is used for routine clinical practice48,49 and hence their potential for use as a scaffold material for tissue engineering of other tissue types is attractive. In this study we used collagen hydrogels to determine the osteogenic responses of MSCs to bFGF in a 3D culture environment that is more representative of in vivo tissue structure.
Initially, we determined the degree of cell proliferation in the collagen hydrogels. We first dosed the hydrogels with varying concentrations of MSCs to determine the appropriate cell seeding density that could be used without promoting a contractile phenotype as researchers working with fibroblasts have reported that collagen hydrogel contraction increases as cell seeding density increases. 44 Based on our observations that only the highest cell seeding density (2.5×104 cells per hydrogel) promoted visible contractions during the culture periods relevant to this study (3–4 weeks), subsequent experiments characterizing the effects of bFGF were conducted using MSC seeding densities up to 1×104 cells per hydrogel. Using this 3D collagen hydrogel culture method, we investigated the effect of bFGF on MSCs, as bFGF is known to be a potent mitogen for mesenchymal/osteoprogenitor cells.33,50,51
Before the observation of bFGF effects on cell behavior, we sought to investigate the quantity of bFGF remaining within the hydrogel matrices during the culture period. Because it is not easy to directly assess the bFGF within the hydrogel, we measured the released amount from the hydrogel and assumed the possible remaining quantity. Results indicated the bFGF incorporated within the hydrogels was released in a steady and sustainable manner over a month and the total released quantities up to 30 days were about 50%–75% depending on the initial loadings, which allows us to presume the bFGF incorporated within the hydrogel was not released completely, rather remained at a certain level within the hydrogel matrix. Although the remaining quantity may not be the direct subtraction between the released and the initial, the result suggests the possible amount of bFGF remaining within the hydrogel and its subsequent effects on MSCs behavior. When we consider this, our system of incorporating growth factors within the hydrogels containing cells inside should have merits for the purposes of cell carriers and tissue engineering as injectable matrices. In fact, the enhancement of MSC proliferation by bFGF has also been observed in 3D scaffolds of β-tricalcium phosphate, 52 those composed of PLGA nanofibers 53 and composite scaffolds of microfibrous silk coated with PLGA nanofibers. 54 bFGF therefore has potential benefit for expanding cells within 3D scaffolds to obtain therapeutic numbers and coupled with its ability to promote osteogenic differentiation of MSCs in vitro55–57 may help to attain functional engineered tissues for bone regeneration purposes. Compared to the porous type of scaffolds, the hydrogel matrices used herein can easily incorporate tissue cells within, providing gel-like tissue mimicking microenvironments. Furthermore, when therapeutic molecules like bFGF were incorporated together, the constructs can be a potential cell delivery and tissue engineering system. This study is thus considered of value compared to other bFGF studies such as the observation of biological effects on osteoblast or MSC behaviors under 2D culture plastic dish31,32,35 or within 3D gel matrices (but not incorporated within)58,59 or those utilizing bFGF within 3D porous type of scaffolds.52–54
We tested whether the same is true for collagen hydrogel cultures by assaying a wide range of bFGF concentrations up to 500 ng and found that an optimal bFGF concentration of 5 ng stimulates cell proliferation, whereas concentrations above 50 ng are inhibitory. We then looked in greater detail at a narrow range of bFGF concentrations with a range of MSC seeding densities to assess more precisely the optimal concentration of bFGF for enhanced MSC proliferation responses in 3D hydrogels. We found that both in the presence and in the absence of bFGF, MSC number increased over time in accordance with initial cell seeding density. This is not surprising as the hydrogels were not released from the sides of the well during culture and are therefore anchored. Other studies using fibroblasts indicate that cell number increases in anchored collagen hydrogels in contrast to the cessation of cell proliferation that is typical of free-floating hydrogels46,60–62 in the absence of externally applied stresses. 63 Therefore, we can conclude that these proliferative responses of fibroblasts in hydrogels are comparable for MSCs.
The enhanced cell proliferation in the presence of up to 5 ng bFGF, at all cell seeding densities, indicates that the mitogen activity of bFGF is maintained in 3D culture, as the case in monolayer culture previously described.33,50,51 However, increasing the concentration of bFGF beyond 5 ng resulted in a decline in cell number at each time point. Whether this was a result of reduced cell proliferation or apoptosis is not clear, but as continued culture of MSCs in the presence of 5 ng bFGF for 14 days did not result in reduced cell number, it is assumed that the effect is simply blockage of proliferation.
Collagen hydrogels provide a suitable scaffold that induces osteogenic differentiation of MSCs. 64 The substrate can support multilineage differentiation depending on instructional signals incorporated into the gel: with BMP2 promoting chondrogenic differentiation, while dexamethasome, ascorbate-2-phosphate, and β-glycerophosphate promoting osteogenic differentiation. 65 The osteogenic and bone regeneration capacity of collagen as a hydrogel substrate was superior to that of fibrin glue and alginate 66 and modifying collagen hydrogels by loading them with connective tissue growth factor (CTGF) could enhance osteogenic differentiation even further. 67
Here, we determined the effect of bFGF on osteogenic differentiation of MSCs in collagen hydrogels. bFGF is a good candidate because we previously found that osteogenic phenotype acquisition of preosteoblastic MC3T3-E1 cells grown on hydroxyapatite bone granules was much greater in the presence of bFGF than without. 68 In the current study, we found that osteogenic differentiation of MSCs in collagen hydrogels was also improved in the presence of bFGF. Enhanced osteogenic differentiation was evident at the molecular, cellular, and extracellular levels. Even though ALP levels after 7 days of culture with bFGF-containing medium were the same as with medium lacking bFGF, after 14 days, there was a significant increase in detectable ALP in the presence of bFGF. Gene expression analysis also confirmed the delayed effect of bFGF upon osteogenic induction with BSP, OPN, and OCN transcription only being significantly upregulated after 14 days of culture. Elevated Col I expression was detected after just 7 days, correlating with the increased cell number present at this point in the early osteogenic phase. Finding significant increases in osteogenic gene expression at 14 days in the presence of bFGF compared to bFGF-free control prompted further investigation, so OPN was selected as a candidate for further examination to determine whether transcriptional enhancement by bFGF was evident at the protein level. Elevated OPN protein production due to bFGF exposure was confirmed by western blot analysis, and a greater proportion of cells within the population becoming OPN producing were confirmed by FACS. Immunofluorescence imaging provided visual confirmation of these results.
Enhanced proliferation and osteogenic differentiation of MSCs in 3D collagen hydrogels containing bFGF is encouraging evidence that fabricated collagen structures can be used as an appropriate 3D scaffold for bone regeneration, with appropriate growth factor addition. The benefits of collagen scaffolds for bone regeneration strategies are also evident for other scaffold types that have been doped with bFGF,52–54 but the natural role of collagen during tissue morphogenesis makes it particularly attractive as a scaffold. Indeed, enhanced bone regeneration in a critical sized defect was obtained when collagen membranes doped with nano-sized bioactive glasses had bFGF incorporated into them. 69 This correlates with data presented in the current study whereby matrix mineralization after several weeks in culture is greater in hydrogels containing bFGF. bFGF is not the only growth factor that can produce improved osteogenic responses and significant benefits have also been reported for other growth factors, including CTGF, 67 platelet-derived growth factor, 70 insulin-like growth factor, 71 and BMP272–74 when loaded into various types of scaffold. However, bone regeneration process is quite complicate requiring numbers of growth factors and the interactions of them, and the optimal functions are expected with administrations at proper time and dose. 75 It is thus considered that the presentation of several growth factors combined in collagen hydrogels may yield better results than either factor alone as have recently been proposed in systems under 2D culture or using porous scaffold materials.33,75–77 The exact balance of combined delivery such as bFGF with other growth factors in terms of optimal mitogenic and osteogenic effects in vitro and in vivo remains to be understood and the work needs to follow as further study.
Conclusions
Osteogenic induction of MSCs can be promoted in 3D collagen hydrogels in the presence of bFGF. The use of 3D collagen hydrogel scaffolds for regenerative medicine approaches such as tissue engineering has great potential for skeletal regeneration and replacement. The combination of natural biomimetic scaffold material and appropriate osteogenic growth factor signals are favorable for clinical use and this strategy of applying bFGF to collagen hydrogels containing MSCs could be of considerable benefit for patients requiring bone replacement therapies.
Footnotes
Acknowledgments
This work was supported by Priority Research Centers Program (grant no. 2009-0093829) and WCU Program (grant no. R31-10069) through the National Research Foundation (NRF) funded by the Ministry of Education, Science and Technology, Republic of Korea. The authors thank Dankook University IBST for the kind support.
Disclosure Statement
No competing financial interests exist.