
Abstract
Natural extracellular matrix (ECM) deposited in situ by cultured endothelial cells (ECs) has been proven effective in accelerating endothelialization of titanium (Ti) cardiovascular implants (CVIs) in our previous studies. In this study, the ECM deposited by smooth muscle cells (SMCs) was used in comparison to investigate the effects of tissue specificity of the ECM on the ability to accelerate endothelialization of CVIs. The results demonstrated that the ECM deposited by ECs and SMCs (EC-ECM, SMC-ECM, respectively) differed considerably in components and fibril morphology. Surface modification of Ti CVIs with both types of natural ECM was effective in improving their in vitro hemocompatibility and cytocompatibility simultaneously. However, the endothelialization of ECM-modified Ti CVIs in a canine model demonstrated a high tissue specificity of the ECM. Although the ECM deposited by SMCs (SMC-ECM) induced fewer platelet adhesion and sustained better growth and viability of ECs in vitro, its performance in accelerating in vivo endothelialization of Ti CVIs was extremely poor. In contrast, the ECM deposited by ECs (EC-ECM) led to complete endothelium formation in vivo.
Introduction
The present ECM modification of CVIs aims to mimic the microenvironment of ECs, namely the basement membrane. At first, along with the identification and separation of ECM components, more and more ECM proteins have been used to modify CVIs, such as fibronectin (FN), laminin (LN), and collagens (Col).1,10,11 Furthermore, during the construction of an artificial ECM on CVIs, several techniques, including electrostatic spinning and microcontact printing, have been taken to mimic the physical characteristics of the natural ECM.12,13 Even so, when these artificially constructed ECMs were compared with ECMs deposited in situ by cultured cells, the latter ones could mimic the natural ECM to a much greater extent, and presented far superior biological properties. Matsubara et al. found that the ECM deposited by primary ECs outperformed collagen type IV (Col IV), LN, basic fibroblast grow factors (bFGF), and even Matrigel in sustaining proliferation and maintaining differentiation potential of mesenchymal stem cells. 14 Moreover, the ECM deposited by ECs could sustain proliferation of sparsely seeded ECs in a bFGF-free culture medium. 15
In our previous study, the ECM deposited in situ by human umbilical vein endothelial cells (HUVECs) endowed porous titanium (Ti) CVIs with better hemocompatibility, better cytocompatibility, and accelerated in vivo endothelialization. 16 However, natural ECMs have obvious tissue specificity. Wicha et al. reported that mammary epithelial cells could maintain the secretion of the milk sugar lactose when cultured on an acellular matrix of mammary origin, whereas the secretion behavior was rapidly lost when cultured on a collagen gel or a matrix of liver origin. 17 Zhang et al. further confirmed the tissue specificity of the ECM by studying the influences of ECM origin on growth of the corresponding cells, and found that the cells of specific origin obtained a higher proliferation and differentiation rate when they were cultured on the matched ECM. 18 Accordingly, we hypothesized that the ECM of endothelial origin would perform better in sustaining ECs growth and functions, thus perform better in accelerating endothelialization of CVIs. To warrant the hypothesis, ECMs deposited in situ by ECs and smooth muscle cells (SMCs) (referred to as EC-EMC, SMC-ECM, respectively) were applied to modify Ti CVIs, and their performances in improving hemocompatibility and cytocompatibility and in accelerating the in vivo endothelialization rate of CVIs were systematically compared.
Materials and Methods
Materials
Commercial pure Ti was purchased from Baoji Non-ferrous Metal Co. Ltd. Cell culture reagents, Medium 199 (M199), Dulbecco's Modified Eagle's Medium/Nutrient Mixture F12 (DMEM/F12), and fetal bovine serum (FBS) were from Hyclone. Endothelial cell growth supplement (ECGS) was from Millipore. Antibodies to human FN, LN, and Col IV were from Abcam; antibodies to actin and α-smooth muscle actin (α-SMA), and fluorescein isothiocyanate - and TRITC-labeled immunoglobulin G were from Sigma-Aldrich. Alamar Blue reagent was from BioSource. All other reagents beyond those mentioned were purchased from Kelong chemical reagents Co.
Cells isolation and maintenance
HUVECs were isolated according to the method by Jaffe et al. 19 and then were maintained in an M199 medium containing 15% FBS and 20 μg/mL ECGS in a humidified incubator under 5% CO2 at 37°C. Primary human umbilical artery smooth muscle cells (HUASMCs) were obtained by slow emigration from small pieces of the umbilical artery medium in the DMEM/F12 medium with 10% FBS under 5% CO2 at 37°C. The culture medium was changed every 3 days. Cells were subcultured when the cells were grown to 80% confluency, and cells between second and fifth passage were used for experiments.
Preparation of ECM-coated Ti disks
For better retention of the ECM on Ti disks, polished Ti disks were eroded with 5 M NaOH solution before deposition of the ECM as described previously, 16 since porous surfaces have been shown to enhance the protein-loading capability. 20 Then, HUVECs were seeded onto Ti disks at 1×105 cells/cm2, and HUASMCs were seeded at 5×104 cells/cm2. The samples were maintained in the aforementioned culture medium, respectively. Seven days later, all the samples were treated with 0.5% Triton X-100 and 20 mM NH3 in phosphate-buffered saline (PBS) for 20 min, resulting in the shedding of cells and exposure of the ECM. The ECM-coated disks were washed five times with PBS for subsequent experiments. Bare NaOH-treated Ti disks were treated in the same manner.
Difference in composition of the ECM secreted by HUVECs and HUASMCs
To roughly compare the difference in components of ECMs secreted by HUVECs and HUASMCs, immunofluorescent staining was performed by using antibodies to human FN, LN, and Col IV, respectively. Briefly, ECM-coated Ti disks were fixed with 4% paraformaldehyde, washed with PBS, immersed in 1% bovine serum albumin (BSA) for 30 min to block nonspecific staining, and then incubated with the aforementioned antibodies for 30 min at 37°C. The samples were washed thoroughly and incubated with a fluorescently labeled secondary antibody for another 30 min at 37°C, and finally observed by an inverted fluorescence microscope (Olympus IX51). In addition, enzyme-linked immunosorbent assays (ELISAs) using the same antibodies were adopted to acquire relative quantifications of the above components on EC-ECM- and SMC-ECM-coated Ti disks.
Characteristics of ECM-coated Ti disks
The surface topography of both NaOH-treated and ECM-coated Ti disks was detected by an atomic force microscope (AFM; SPI3800N Seiko). The surface chemical groups were further detected using diffuse reflectance Fourier-transform infrared spectroscopy (DR-FTIR, ST-IR20SX; Nicolet Co. Ltd.). To characterize the differences in surface wettability between the NaOH-treated and ECM-coated sample, contact angles were measured using a contact angle goniometer (DSA100; Krüss).
Hemocompatibility assays
Preparation of platelet-rich plasma (PRP), platelet adhesion assay, and detection of lactate dehydrogenase (LDH) activity of adhered platelets were performed according to Tu et al. 16
For the fibrinogen activation assay, a portion of PRP was centrifuged at 3000 rpm for 20 min to precipitate platelets and obtain platelet-poor plasma (PPP). One hundred microliters of PPP was added onto each sample and incubated at 37°C for 1 h. All the samples were washed three times with PBS and then immersed in 1% BSA solution for 30 min to block nonspecific binding. Then, 20 μL mouse-anti-human γ-fibrinogen monoclonal antibody (Accurate Chemical and Scientific Corp.; 1:100 diluted) was added onto each sample, incubated at 37°C for 1 h, and then washed three times with PBS. Twenty microliters of horseradish peroxidase-labeled sheep-anti-mouse polyclonal antibody was added onto each sample and incubated for 1 h at 37°C. After thorough washing with PBS (5 min for three times), 100 μL chromogenic substrate tetramethyl benzidine (TMB) solution was added onto each sample for 10 min. The reaction was stopped by adding 50 μL H2SO4 to each sample, and each 130 μL reactive solution was collected for detection of the optical density by using a Bio Assay Reader (HTS7000 plus; Perkin Elmer) within 10 min.
HUVECs and HUASMCs reseeding assay
Before cell seeding, both the NaOH-eroded Ti and ECM-coated Ti disks were incubated with an HUVEC or HUASMC complete medium for 1 h, respectively. Both HUVECs and HUASMCs were seeded onto the control, EC-ECM-coated, and SMC-ECM-coated Ti disks at 5×104 cells/cm2. Culture conditions were similar to those described above. The medium was changed every 3 days. To investigate the morphology of adherent cells on the different substrates, samples were taken after 1 day and 3 days of culture for immunofluorescent staining, using an antibody to actin to label HUVECs and antibody to α-SMA to label HUASMCs.
To study the proliferation and viability of cells on different substrata, after 1 and 3 days in culture, 400 μL phenol red-free medium containing 5% FBS and 10% Alamar Blue reagent was added to each sample and incubated at 37°C for 3 h, and then each 200 μL medium was transferred to a new 96-well cell culture plate for detection of the optical density at 570 nm (A570) and 600 nm (A600), respectively. A570 minus A600 was used to indicate the viability of adherent cells according to the instructions provided by the vendor. A higher value implies a larger number and higher viability of adherent cells.
Nitric oxide and prostacyclin release of adherent HUVECs
For nitric oxide (NO) and prostacyclin (PGI2) detection, HUVECs were seeded onto each sample at a high density (5×105 cells/cm2) to get rapid formation of a confluent monolayer. After adhesion for 6 h, all the samples were transferred to new 24-well culture plates, and 1 mL fresh culture medium was added to each well. One hundred fifty microliter medium was collected from each well after 24 h. After 2 days, the culture medium was changed (1 mL) and another 150 μL culture medium was collected from each well 24 h later. All the collected culture media were stored at −20°C until analysis.
NO release was indicated by the content of nitrite, which is a stable metabolite of NO. Nitrite was detected using Griess reagent (Sigma), and PGI2 was detected using a human PGI2 ELISA Kit (R&D) according to the product instructions. The contents of both NO and PGI2 were ultimately divided by the cell number.
HUVEC and HUASMC migration assay
HUVECs and HUASMCs were seeded onto the NaOH-eroded Ti, EC-ECM-, and SMC-ECM-coated Ti disks at a high density of 5×105 cells/cm2 to ensure the formation of confluent monolayer after culture for 1 day. The confluent layers of HUVECs and HUASMCs were scratched by a pipette tip to create artificial scratches. All the disks with artificial scratch were cultured for another 1 and 3 days. All the collected samples were kept in a warm cell culture medium until they were fixed as soon as possible to avoid cell shrinkage. Actin was stained for HUVECs and α-SMA for HUASMCs, and then cells were inspected by a fluorescence microscope. Images were processed and analyzed to obtain percentages of wound closure by the migrating and proliferating cells (total area of the migrating cells to area of the artificial scratch) using ImageJ software. At least three samples and nine random fields of view were evaluated to obtain each value.
In vivo endothelialization experiments
For in vivo endothelialization, commercial Ti was cut into small elliptic slices (7×4 mm), with a hole drilled at each end. The slices were polished down to a thickness below 0.2 mm, then eroded, and deposited with the ECM as described above. Before implantation, all the samples were bent in compliance with the curvature of the artery. All animal studies were performed according to the Guide for the Care and Use of Laboratory Animals of China. Canines weighing 20–25 kg were anesthetized; the skin was disinfected, and then their bilateral femoral arteries were exposed. The control, EC-ECM-coated, and SMC-ECM-coated Ti slices were then implanted into the femoral arteries, as shown in Figure 1. The bilateral femoral arteries of a canine were implanted with different types of samples to provide intraindividual comparison. Three animals were enrolled in each study.
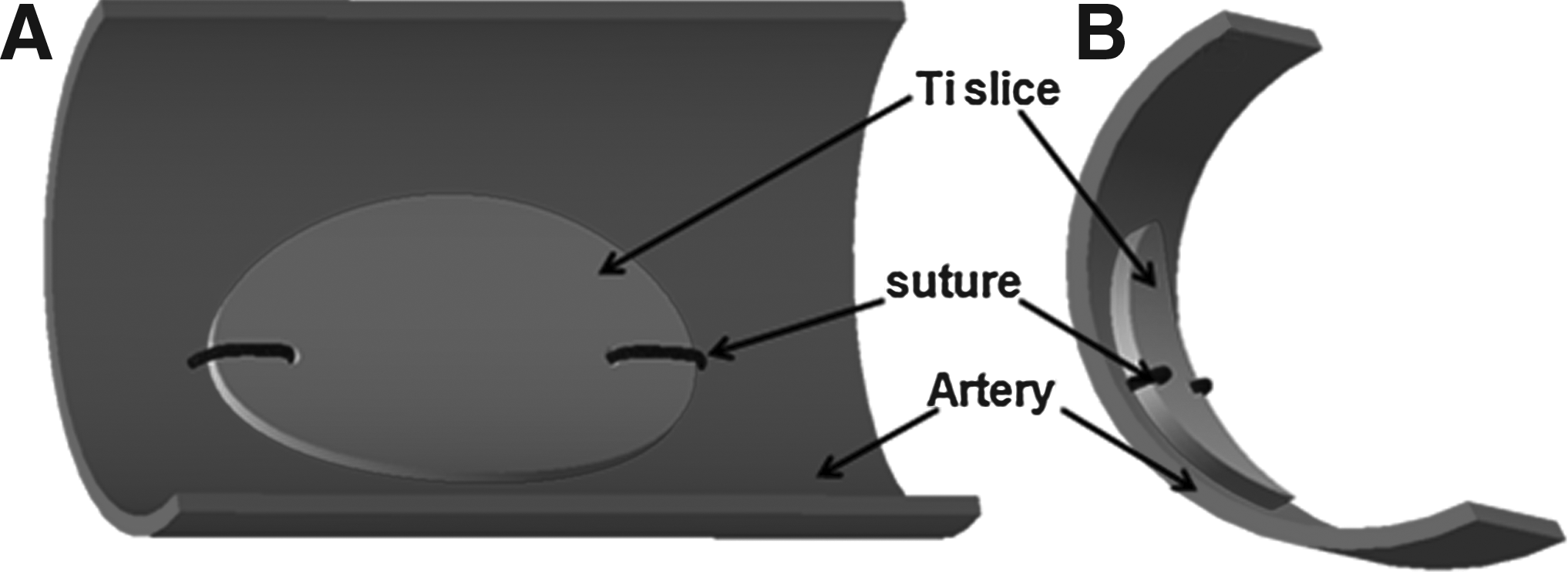
Sketch of the anastomosis of a control or ECM-coated Ti slice in a canine femoral artery.
Six months after implantation, the artery segments with implanted samples were explanted, cut longitudinally, washed, and fixed in 2.5% glutaraldehyde for 12 h for a scanning electron microscope (SEM) observation. The SEM samples were prepared according to the Tu et al.ref 16 .
Statistical analysis
All data were representative of at least three experiments, and data were expressed as means±standard deviations. All statistical comparisons were performed by using analysis of variance, and a probability value <0.05 (p<0.05) was considered significant.
Results
Difference in composition of the ECM secreted by HUVECs and HUASMCs
After exposure to Triton X-100 and NH3 solution for 20 min, the majority of both HUVECs and HUASMCs were washed out, and only few nuclei could be detected on Ti disks via 4′,6-diamidino-2-phenylindole (DAPI) staining (results not shown).
Three major components of the ECM, namely FN, LN, and Col IV, on EC-ECM- and SMC-ECM-coated Ti disks were, respectively, analyzed via immunofluorescent staining. The results are shown in Figure 2. All of the above-mentioned components were secreted by both ECs and SMCs. Both FN and LN adopted obvious fibrillar structures, while Col IV showed dispersed nonfibrillar clusters. There was a minor difference in FN morphology between EC-ECM and SMC-ECM at high magnification. The FN fibers showed a fine reticular network in EC-ECM, whereas in SMC-ECM, the fibers were longer, thicker, and usually distributed in a parallel pattern (Fig. 2B).
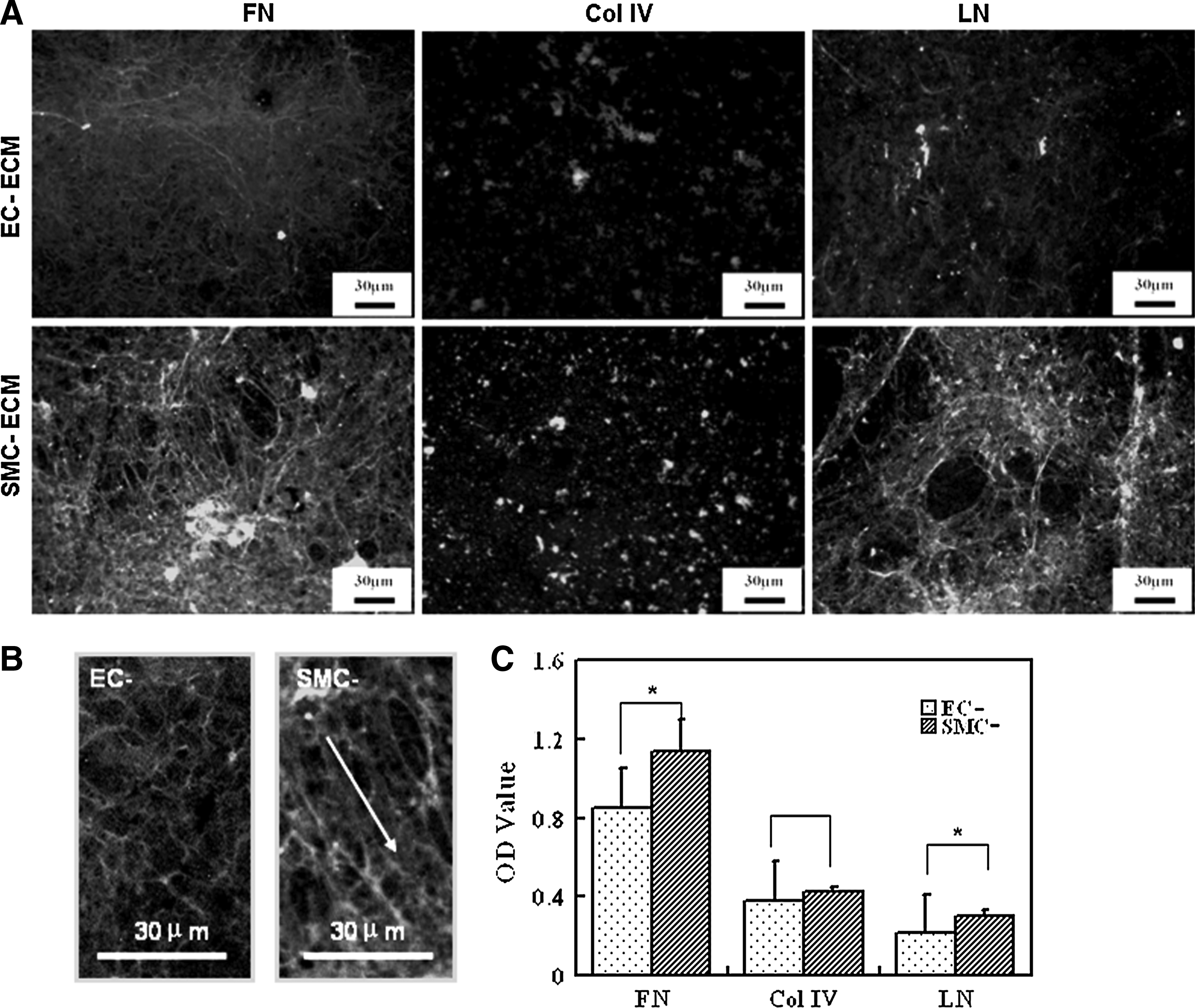
Immunofluorescent images of FN, Col IV, and LN in EC-ECM and SMC-ECM
These above-mentioned components were relatively quantified via ELISA assay. The results showed that there was significantly larger amount of FN and LN in SMC-ECM than that in EC-ECM, but there was no significant difference in the amount of Col IV (Fig. 2C).
Characteristics of ECM-coated Ti disks
The topography of the control and ECM-coated Ti was detected using AFM. Both EC-ECM and SMC-ECM mainly appeared as uniformly distributed granules. However, there were some obvious fibrillar structures on SMC-ECM (Fig. 3).

Atomic force microscope images with an area of 25×25 μm of the control
Figure 4A shows the DR-FTIR spectra of NaOH-treated and ECM-coated Ti disks. In addition to −OH stretch at 3405 cm−1 of NaOH-treated Ti, there were more other distinctive peaks, such as C–H stretch at 2960 cm−1 and 2916 cm−1, C=O stretch at 1650 cm−1, and N–H bend at 1545 cm−1 after ECM deposition. Moreover, the peaks in the spectra of SMC-ECM were sharper than that of EC-ECM.
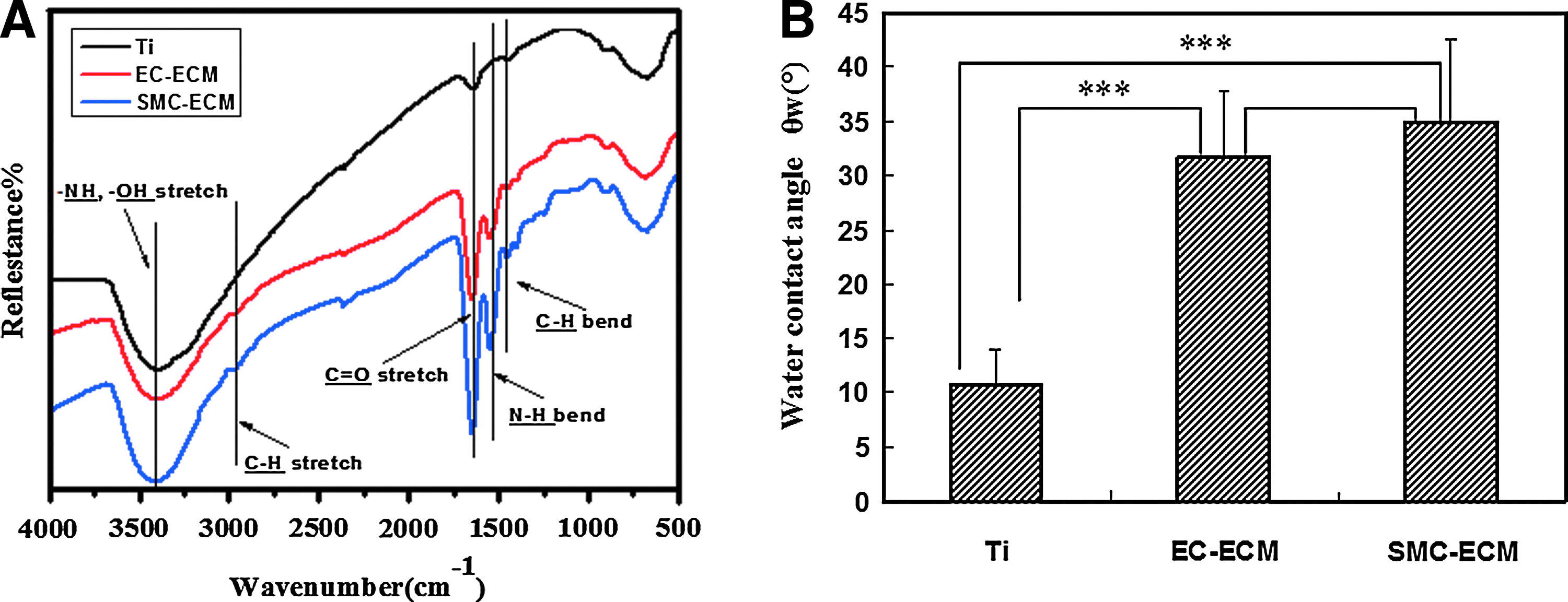
Changes in diffuse reflectance Fourier-transform infrared spectroscopy spectra
In addition to the changes in morphology and DR-FTIR spectra, the contact angles of Ti disks significantly increased after ECM deposition, as shown in Figure 4B, but there was no significant difference between EC-ECM- and SMC-ECM-modified Ti disks.
Effects of ECM coating on hemocompatibility of Ti disks in vitro
Via immunofluorescent staining for P-selectin of adherent platelets, we found that many fewer platelets adhered to either EC-ECM- or SMC-ECM-coated Ti disks as compared with the control Ti disks (Fig. 5A–C). An LDH assay after cell lysis was used to quantify the adherent platelets, and the results further confirmed the reduced platelet adhesion after ECM deposition (Fig. 5D). Moreover, there was a statistically significant difference between the number of adherent platelets on EC-ECM and SMC-ECM, where only about half amount of platelets adhered to SMC-ECM compared to EC-ECM.

Immunofluorescent images of the platelets adherent to the control Ti
Fibrinogen absorbs immediately onto artificial surfaces after exposure to blood. It is generally considered that conformational changes of fibrinogen, such as the exposure of γ-chain C-terminus, remarkably influence platelet adhesion. 21 In this study, exposure of γ-chain C-terminus of adherent fibrinogen was detected via ELISA assay. The results indicated that both ECM modifications effectively diminished fibrinogen conformational changes as compared to the control Ti disks, but there was no statistically significant difference between EC-ECM and SMC-ECM (Fig. 5E).
Effects of ECM coating on HUVECs in vitro
Adhesion and viability of HUVECs
The tendency of EC adhesion onto the control and ECM-coated Ti was different from that of platelets. ECM modification resulted in better EC adhesion and proliferation (Fig. 6). Both qualitative results from immunofluorescent staining and quantitative results from Alamar Blue assay indicated that a larger number of ECs adhered to ECM-coated Ti disks and displayed higher viability and proliferation rates. In addition, SMC-ECM sustained significantly more EC adhesion than EC-ECM (Fig. 6B). Moreover, the source of ECM also evidently influenced the morphology of ECs grown upon it, especially when ECs were grown to confluence. In detail, ECs showed typical cobblestone-like appearance when grown on EC-ECM, and confluent ECs formed capillary-like structures with extremely high-density cells surrounding low-cell-density or cell-free areas. However, ECs cultured on SMC-ECM were strongly elongated into spindle-like shape and in parallel rows, which are typical characteristics of confluent SMCs (Fig. 6A). The aspect ratios, indicated by the ratio of the major axis to the minor axis, of ECs grown on SMC-ECM-modified Ti showed significantly higher values than those on the control or EC-ECM-modified Ti (Fig. 6C).

Growth of ECs on the control Ti, EC-, and SMC-ECM-coated Ti for 1 and 3 days.
NO and PGI2 release of HUVECs
The secretions of NO and PGI2 were detected to partly judge the antithrombogenic potential of ECs. After 1 and 3 days culture, respectively, the conditioned medium of HUVECs was collected for detection of NO and PGI2. The results are shown in Figure 7. There was no significant difference in NO secretion per cell between the HUVECs cultured on the control and ECM-coated Ti for 1 day. However, there was a significantly higher NO secretion per cell cultured on EC-ECM-coated Ti compared to the control Ti and the SMC-ECM-coated Ti after 3 days (Fig. 7A). There were no statistically significant differences in PGI2 production between the control, EC-ECM-, and SMC-ECM-coated Ti (Fig. 7B).
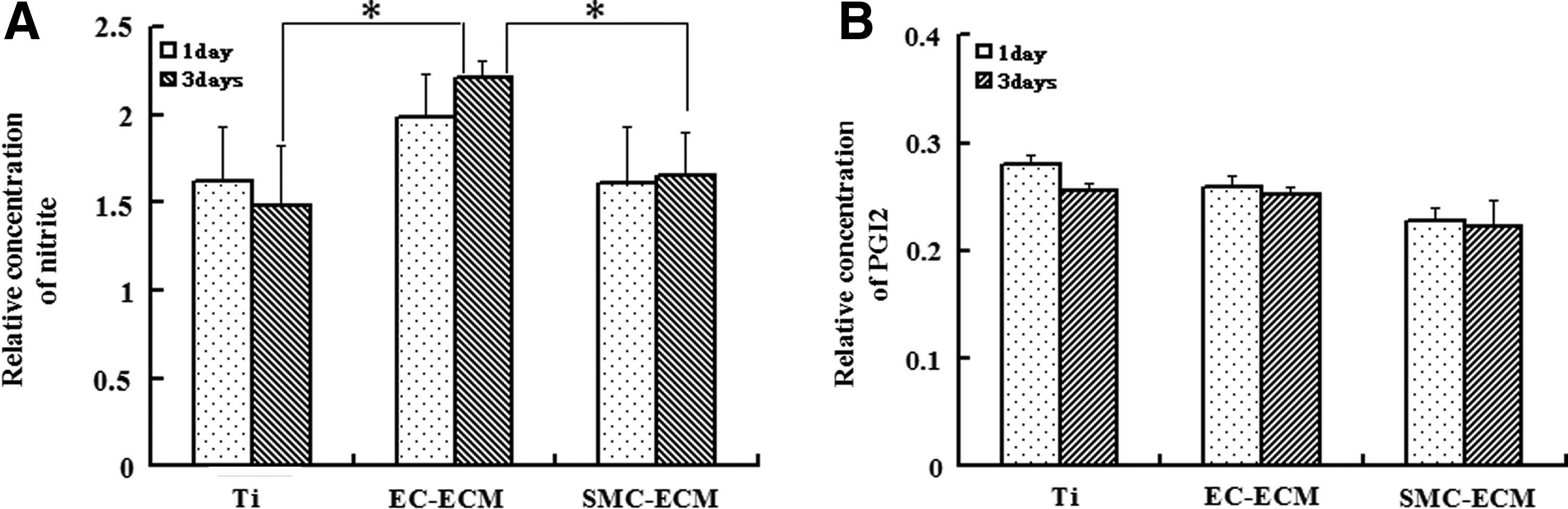
Production of NO
Effect of ECM coating on SMC adhesion in vitro
The influence of both types of ECM on adhesion and proliferation of SMCs was nearly the same as on ECs, as is shown in Figure 8. Although SMCs adhered and proliferated well even on the control Ti disks (Fig. 8A), their growth rate was significantly lower than on SMC-ECM (Fig. 8B). On the EC-ECM-coated Ti disks, the amount of adhered SMCs at 3-day culture, but not at 1-day culture, was a little higher than that on the control Ti disks. SMC-ECM induced significantly more SMC adhesion and a higher proliferation rate than EC-ECM (Fig. 8B).

Growth of SMCs on the control Ti, EC-ECM-, and SMC-ECM-coated Ti for 1 and 3 days.
Effect of ECM coating on migration of ECs and SMCs in vitro
In this study, scratch wound assays were designed to study the migration behavior of ECs and SMCs (Fig. 9A). The results showed that both EC-ECM and SMC-ECM significantly improved the ability of ECs and SMCs to migrate. After 3 days, the migrating ECs or SMCs, together with their offspring, almost covered the artificial scratches on EC- and SMC-ECM-modified Ti disks, but not on the control Ti disks (Fig. 9B, C). The migration of ECs on the control Ti disks was the worst (Fig. 9B). Images were processed and analyzed to obtain approximate area coverage percentages of the artificial scratches by migrating and proliferating cells. The results shown in Table 1 further demonstrate that both types of ECM modification significantly improved both EC and SMC migrations, without statistically significant difference between EC-ECM and SMC-ECM.
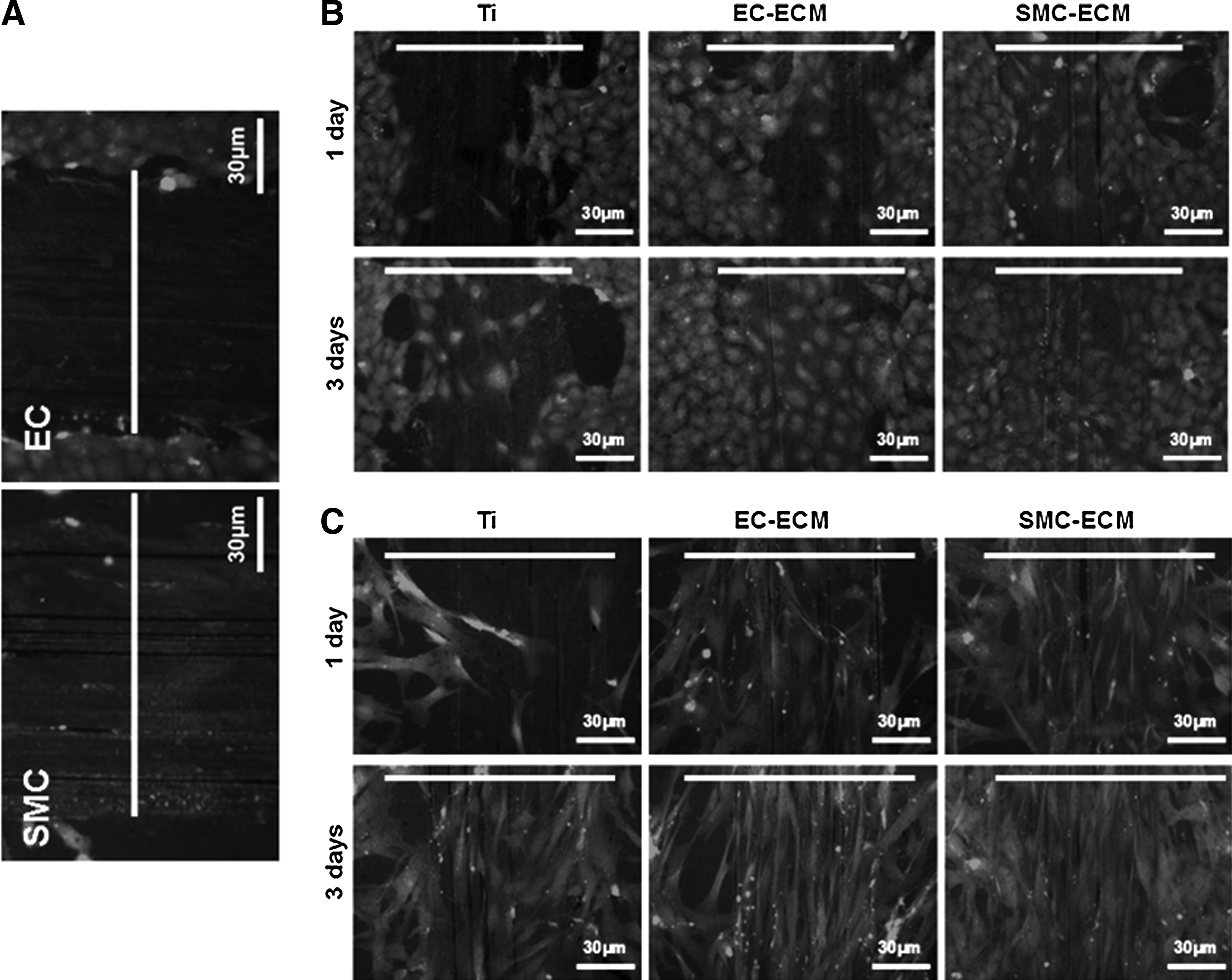
Migration of both ECs and SMCs on the control Ti, EC-, and SMC-ECM-coated Ti.
Significant difference from EC-ECM (ap<0.01, bp<0.005).
Significant difference from Ti (cp<0.01, dp<0.005, ep<0.05).
Data are expressed as means±standard deviation.
Ti, titanium, EC, endothelial cell; SMC, smooth muscle cell; ECM, extracellular matrix.
In vivo endothelialization of the control and ECM-coated Ti slices
All of the femoral arteries implanted with both the control and ECM-coated Ti slices kept patent for up to 6 months. ECs of natural vascular intima migrated through the anastomotic region to the center of the implanted Ti slices. SEM inspection was performed at two separated sites per sample, at the proximal anastomotic region and near the central region of the implant. The results are shown in Figure 10. Only on the surface of EC-ECM-coated Ti slices, a confluent monolayer of cells covered both the anastomotic and central region, and the cells presented their typical cobblestone-like morphology. However, the control and SMC-ECM-coated implants were poorly endothelialized, where the proximal regions were partly covered with ECs, and the central regions were nearly free of cells. The central regions of the control and SMC-ECM-coated Ti slices were mainly covered with fibrils. Moreover, cells on these two types of Ti slices were elongated and spindle-like, with the major axis parallel to the blood fluid in vivo.
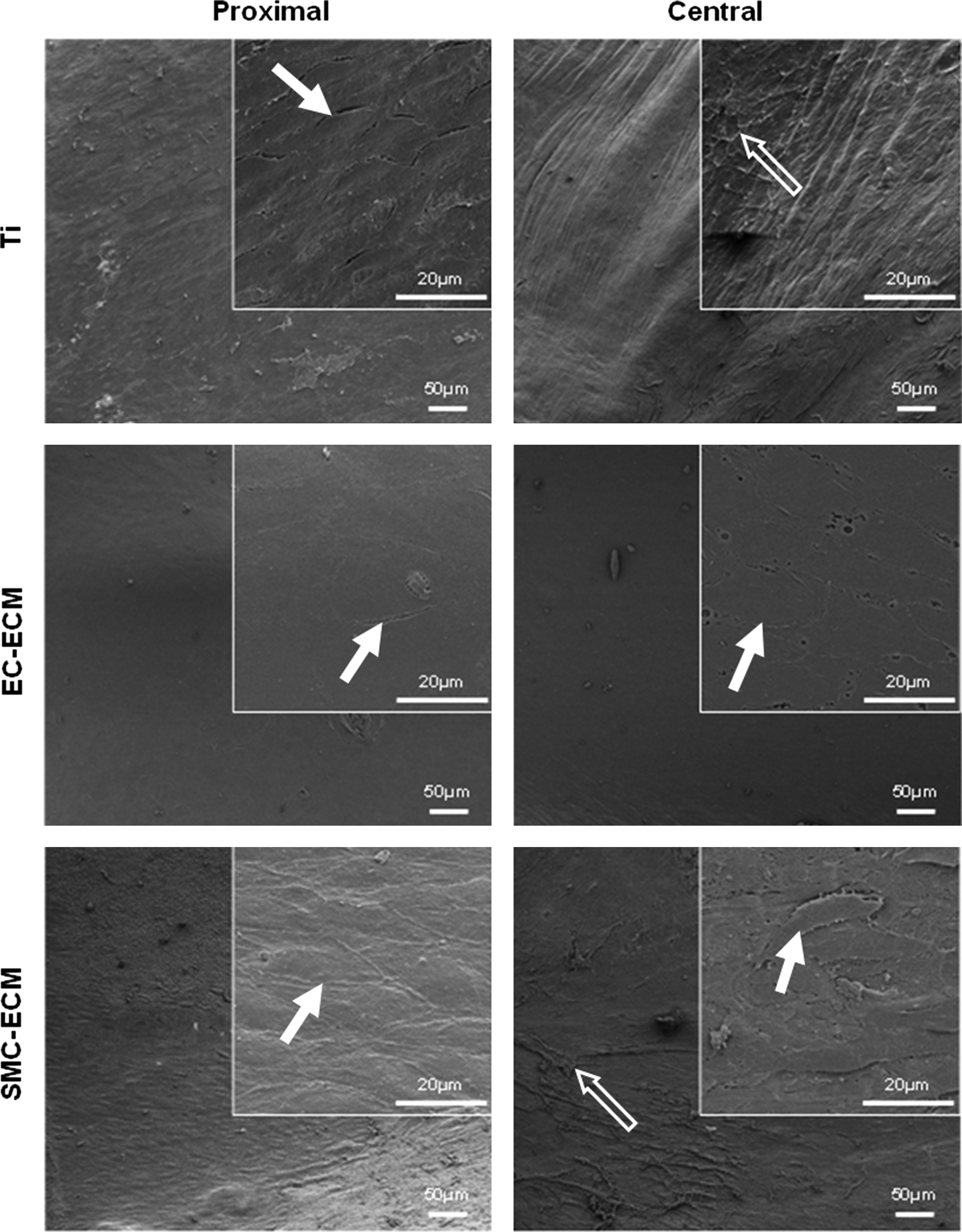
Scanning electron microscope images of explanted Ti slices after 6-month implantation in canine arteries, including the control Ti and Ti slices modified by EC-ECM and SMC-ECM, respectively. Inserts are images at higher magnification (bar=20 μm). The images were obtained from two separate regions, close to the proximal anastomosis and at the central region of each sample. The solid arrows point to the cells, and the hollow arrows point to fibrils.
Discussion
Two reasons that result in the failure of CVIs are intima hyperplasia and thrombus formation, both of which are related to absent or incomplete endothelium formation.22,23 Therefore, constructing an appropriate bioactive surface for ECs to adhere, proliferate, and maintain their normal phenotype is of great importance. Naturally, almost all cells are embedded or in intimate contact with the ECM, a complex meshwork of interlacing macromolecules, such as collagens, proteoglycans. and glycoproteins. 24 Once differentiated cells are removed from their native matrix, they can rapidly lose their important biological characteristics. 17 Physiologically, ECs tightly adhere to the basal lamina, which is mainly secreted by ECs grown upon, and therefore we deduce that the ECM deposited in situ by cultured ECs may mimic their natural habitat to a greater extent. In our previous study, the ECM deposited by ECs had been proven effective in improving in vivo endothelialization of Ti vascular implants. 16 In this study, we test whether EC-ECM is superior to the ECM of other origin for ECs to migrate, proliferate, and maintain their proper phenotype. If so, the reasons should be elucidated. To partly answer the question, EC-ECM and SMC-ECM were deposited in situ onto Ti vascular implants. Their composition, surface characteristics, and their influences on platelet adhesion, on adhesion, proliferation, and migration of ECs and SMCs, and on in vivo endothelialization were systematically studied.
Both ECs and SMCs were capable of depositing a network of ECM on Ti implants, but there were some differences in composition and morphology between EC-ECM and SMC-ECM. Relative quantification of three important ECM components indicated that SMC-ECM contained significantly more FN and LN compared to EC-ECM (Fig. 2). In addition, SMC-ECM showed sharper peaks in the DR-FTIR spectra (Fig. 3). Tan et al. had studied the ECM gene expression by ECs and SMCs from different origins and also found that both cell type and cell origin could influence the expression of ECM genes. 25 Furthermore, the morphology of FN in EC-ECM was obviously different to that in SMC-ECM. The FN fibers presented a uniform network in EC-ECM, whereas a markedly longer and parallel-distributed pattern appeared in SMC-ECM (Fig. 2A). Similar results are reported by Hsieh and Bourn. 26
Moreover, tissue specificity of ECM was also reflected in its biological properties. The different biological properties between EC-ECM and SMC-ECM are summarized in Table 2. SMC-ECM induced a higher number of adherent ECs and higher proliferation rate than EC-ECM did, which was different from Wicha et al. 17 and Zhang et al. 18 The reports showed that cells from special tissue obtained optimal growth and phenotype when they were cultured on the ECM from the same origin.17,18 However, in our study, ECs grown on SMC-ECM showed elongated morphology and secreted much less NO than those on EC-ECM. Most importantly, the Ti CVIs modified by EC-ECM obtained complete in vivo endothelialization, but not by SMC-ECM. In the following section, we try to discuss the relationship between ECM components and their biological properties.
“−” means lower amount or lower speed, and “+” means higher amount or higher speed.
NO, nitric oxide; PGI2, prostacyclin.
What is the reason for the better in vitro hemocompatibility of SMC-ECM compared to EC-ECM? Although both types of ECM deposition on Ti improved the hemocompatibility, platelets adherent on SMC-ECM were significantly fewer compared to EC-ECM (Fig. 5D). In this study, we demonstrated that the quantity of a certain ECM component was different between ECMs of different origin. There was a higher amount of FN in SMC-ECM, which can be regarded as responsible for the platelet adhesion to the ECM, 27 in parallel with higher amount of LN, which has been proven not to cause platelet aggregation. 28 So, the available data about the component differences between EC-ECM and SMC-ECM were not sufficient to elucidate their differences in hemocompatibility. With the development of proteomics methods, it is possible to compare the total set of ECM proteins synthesized by various types of cells, 29 which may provide further insight into the relationship between ECM components and hemocompatibility. However, it is well known that von Willebrand factor (vWF) is specifically expressed and deposited onto the ECM by the ECs, but not by SMCs, and some researchers thought that the adhesion site in EC-ECM for platelets was probably a complex of vWF, FN, and collagen molecules. 27 So, the lack of vWF may be partly responsible for the fewer platelet adhesion on SMC-ECM than on EC-ECM.
What is the reason for the larger number of adherent cells and the higher proliferation rate on SMC-ECM compared to EC-ECM? Surface modification of Ti disks with both types of ECM resulted in better cytocompatibility. However, it seemed there was no tissue specificity in a strict sense between these two types of ECM according to the in vitro cell growth and migration assays, in that EC-ECM failed to inhibit the growth and migration of SMCs as expected (Fig. 8), and vice versa. The relationship between ECs and SMCs has been studied in various models, including direct coculture, conditioned media, and the ECM model. SMCs and their ECM either promote adhesion and growth of ECs 30 or inhibit these activities. 31 ECs and their ECM either promote adhesion and growth of SMCs 32 or inhibit them. 33 Many factors have been reported to be able to influence the relationship between ECs and SMCs, such as species from which the cells are obtained, cell culture conditions, and cell confluence degree.34,35 In this study, both EC-ECM and SMC-ECM stimulated the growth of ECs and SMCs, but SMC-ECM achieved a higher degree of stimulation compared to EC-ECM. This may be partly attributed to the larger amount of ECM secreted by SMCs, as reported by Bareille et al. 36 However, our subsequent results indicate an unequivocal effect of ECM composition and not of quantity on cell adhesion and proliferation: ECs cultured for 7 days and SMCs cultured for 4 days on Ti disks deposited nearly equal amount of ECM; however, SMC-ECM still sustained higher cell adhesion (data not shown).
What is the reason for the limited in vivo endothelialization on SMC-ECM compared to EC-ECM? Although Ti disks modified by SMC-ECM induced the least platelet adhesion, the highest adhesion, and proliferation rate of ECs in vitro, their ability to accelerate endothelialization in vivo was relatively limited (Fig. 10). It is comprehensible that there were contradictory results between in vitro and in vivo experiments, because re-endothelialization in vivo is a complicated process, and there are many conditions impossible to be replicated in vitro. Although the SMC-ECMs had better hemocompatibility and sustained higher adhesion and proliferation rates of ECs in vitro, there are still some factors that strongly impact their in vivo performance. For example, ECs grown on SMC-ECM displayed an elongated morphology with parallel orientation, similar to the physiological appearance of SMCs. In contrast, ECs grown on EC-ECM displayed a morphology that was more similar to that observed in vivo, and cells formed capillary-like structures after reaching confluence. The influence of the ECM on the morphology of ECs grown upon also has been reported by Hsieh and Bourn. 26 They found that ECs grown on the ECM of fibroblasts with a parallel fiber distribution showed an elongated and parallel-oriented cell morphology. Kim et al. summarized that cell morphology can be regulated by designing its contacting surface, and the change in morphology can regulate cell signal transduction, cell growth, migration, differentiation etc. 37 In this study, ECs grown on EC-ECM secreted a significant higher amount of NO than on SMC-ECM, which was proven effective in inhibiting platelet adhesion and intima hyperplasia. 38 Another possible explanation is the more powerful ability of SMC-ECM to sustain the growth and migration of SMCs than EC-ECM. Walpoth et al. used fibrin-releasing vascular endothelial growth factor (VEGF) to modify small-diameter expanded polytetrafluoroethylene (ePTFE) grafts, and found that fibrin-releasing-VEGF resulted in obvious neointima hyperplasia beyond that seen in uncoated grafts. 39 The stimulating effect of VEGF on growth of ECs has been generally accepted, 40 so Walpoth et al. deduced that probable activation of VEGF on SMCs resulted in this deleterious outcome. We therefore postulate that for the evaluation of the long-term potency of a specific modification of CVIs, it is not sufficient to perform in vitro assays of platelet adhesion, and adhesion and migration tests of ECs. Detrimental influences of various origins on the functions of ECs or inappropriate promotion effects on SMCs may neutralize or even subvert the in vitro advantages, eventually resulting in a failure in vivo. That is to say, the artificial ECM that favors ECs instead of SMCs will be more suitable for surface modification of CVIs. As a whole, tissue specificity of the ECM was eventually embodied from the in vivo endothelialization experiments.
Conclusion
In this study, we systematically studied the ability of the ECM, deposited in situ by cultured cells, in accelerating endothelialization of Ti CVIs. Tissue specificity of the ECM was taken into account for the first time in this field. We found that both EC-ECM and SMC-ECM were efficiently improved in vitro hemocompatibility and cytocompatibility of Ti CVIs, and in this regard, SMC-ECM performed better than EC-ECM. However, the in vivo canine assay indicated that complete endothelialization was only achieved on the EC-ECM, but not on the bare or SMC-ECM-modified Ti CVIs. The contradiction between in vitro and in vivo performances of SMC-ECM may be attributed to the abnormal shape and function of ECs induced by SMC-ECM, or the higher ability of SMC-ECM to induce proliferation and migration rate of SMCs.
Footnotes
Acknowledgments
This work was supported by the National Basic Research Program of China (Grant No. 2011CB606204), the Natural Science Foundation for the Youth of China (Grant No. 31000426), the Fundamental Research Funds for the Central University (Grant No. SWJTU11CX057), and the Natural Science Foundation of China (Grant No. 50971107).
Disclosure Statement
No competing financial interests exist.