
Abstract
Directing chondrogenic differentiation of human mesenchymal stem cells (MSCs) is currently a challenging problem in tissue engineering of cartilage. Short-peptide motifs are promising new tools to aid in controlling chondrogenesis. The aim of this study was to investigate whether a short bone morphogenetic protein-2 (BMP-2)-derived peptide (BMP peptide) stimulates chondrogenesis of human MSCs in the absence of other growth factors. A high-throughput pellet culture system was used to rapidly collect biochemical data such as glycosaminoglycan (GAG), total collagen, and DNA content, as well as alkaline phosphatase (AP) activity. Cells cultured with ≥100 μg/mL of the peptide produced 74% of the GAG content that cells cultured with BMP-2 produced. Comparable levels of GAG production were promoted by the peptide and BMP-2 over 4 weeks of culture. However, histology revealed that the peptide promoted a more homogenous distribution of GAG than BMP-2. The BMP peptide directed human MSCs to increase collagen production after 3 weeks, but at significantly lower levels compared to BMP-2. Treatment with BMP-2 resulted in a large increase in hypertrophic markers such as AP activity and gene expression of type X collagen, whereas treatment with the peptide resulted in little-to-no increase in these markers. These results suggest that the BMP peptide could be an effective new tool for cartilage tissue engineering.
Introduction
MSCs are typically differentiated to cartilage through the use of soluble factors. Transforming growth factor-β3 (TGF-β3) is a potent inducer of chondrogenesis. 2 It was included in early medium formulations for chondrogenesis of human MSCs 3 and was shown to promote matrix production more robustly than TGF-β1. 4 A recent study demonstrated that implanting human MSCs and TGF-β3-releasing microparticles into SCID mice resulted in a cartilage-like phenotype in vivo. 5 Although TGF-β3 is an important tool in cartilage engineering, it has been shown to direct the differentiation of human MSCs to hypertrophic chondrocytes in vitro and in vivo. 6 Thus, there exists a need for additional growth factors and cues to control the differentiation state. Bone morphogenetic protein-2 (BMP-2) has also been implicated as a powerful chondrogenic agent for human MSCs in vitro7–9 and encourages articular cartilage healing in vivo.10,11 When BMP-2 is combined with TGF-β3, an enhanced or synergistic response in chondrogenesis is observed.7,12
New bioactive molecules have emerged as promising tools in cartilage engineering. Especially interesting is the diverse range of short-peptide motifs with bioactive properties. Because of their smaller size, peptides are often simpler and cheaper to manufacture than full-length growth factors and are easily incorporated into scaffolds. Motifs derived from fibronectin (RGD),13,14 decorin, 15 and collagen 16 have been shown to affect chondrogenesis of human MSCs. In addition, matrix metalloproteinase sequences have been used to enhance chondrogenesis by allowing human MSCs to release bioactive molecules and enzymatically degrade scaffolds as they secrete an additional matrix.14,17 In addition to short peptides, small chemical functional groups that affect human MSC differentiation have been identified via microarray analysis. 18 These short peptides and chemical moieties can be useful for controlling the differentiation of MSCs.
We investigated an additional tool, a peptide derived from BMP-2 (BMP peptide), for its ability to direct chondrogenesis of human MSCs. The peptide (KIPKASSVPTELSAISTLYL) is derived from residues 73–92 of the knuckle epitope of BMP-2. It increased alkaline phosphatase (AP) activity, an early marker for bone formation, in murine osteoprogenitor cells. 19 It also promoted dose-dependent bone healing when loaded in α-tricalcium phosphate scaffolds and implanted in a rabbit radial bone-defect model. 20 Further studies have demonstrated the osteogenic activity of the peptide using various cell types21–25 ; however, the effect of the peptide on chondrogenesis has not been investigated. Since the BMP peptide is bioactive and BMP-2 promotes cartilage matrix formation, we hypothesized that in the absence of other growth factors, the peptide would direct human MSCs to produce a cartilage matrix in a pellet culture. Thus, this study examines the role of the BMP peptide in inducing chondrogenesis of human MSCs by measuring its effect on gene expression and extracellular matrix production and distribution.
Materials and Methods
Cell culture
Human MSCs (Lonza) were cultured at 37°C and 5% CO2 in a low-glucose Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (Lonza 14-501F), 100 U/mL penicillin, 100 μg/mL streptomycin, and 10 ng/mL basic fibroblast growth factor 26 (bFGF; R&D Systems, courtesy of the National Cancer Institute Preclinical Repository). Cells were subcultured after reaching 60%–80% confluence.
Pellet cultures were initiated with cells in passage 3. Briefly, cells were centrifuged, washed twice with a chondrogenic medium, and pelleted in a high-throughput culture system 27 using 96-well, conical-bottom, polypropylene plates (Nunc 249946). For the BMP peptide concentration studies, cells were cultured using 30,000 cells per pellet (n=5, with each sample containing two pellets). For experiments measuring glycosaminoglycan (GAG) and total collagen content (n=5–6, with each sample containing two pellets) and AP activity (n=12), cells were cultured using 60,000 cells per pellet. For histology and quantitative reverse transcription–polymerase chain reaction (qRT-PCR), pellets contained 250,000 cells and were cultured in 15-mL polypropylene tubes (Denville c1018-p). Pellet cultures were maintained in a chondrogenic medium containing a high-glucose DMEM supplemented with 100 nM dexamethasone, 50 μg/mL ascorbic acid 2-phosphate, 1 mM sodium pyruvate, 50 μM L-proline, 1% ITS+ (BD Biosciences), 100 U/mL penicillin, 100 μg/mL streptomycin, and 4 mM L-glutamine. The medium contained 10 ng/mL TGF-β3 (PeproTech), 200 ng/mL BMP-2 (PeproTech), 100 μg/mL of the BMP peptide (synthesized by Chi Scientific at 95% purity), or no growth factors (negative control). The medium was exchanged three times per week. Pellets were harvested at 3, 7, 14, 21, and 28 days.
Papain digestion
Pellets were rinsed in phosphate-buffered saline (PBS) and digested with 125 μg/mL papain in a digest buffer (5 mM
DNA quantification
DNA was measured as previously described 28 using Hoechst 33258 dye. Briefly, 50 μL of the digested pellet was combined with 50 μL Hoechst dye solution (0.7 μg/mL). Fluorescence measurements were taken with an excitation wavelength of 340 nm and emission wavelength of 465 nm. A standard curve was obtained from calf thymus DNA. GAG and total collagen contents were normalized to the amount of DNA measured per sample.
GAG quantification
GAG content was measured by dimethylmethylene blue assay as previously described. 29 Chondroitin sulfate C from shark cartilage was used as a standard. Briefly, 20 μL of the digested pellet was combined with 250 μL dimethylmethylene blue dye solution, and the absorbance was immediately measured at 525 nm.
Total collagen content
After acid hydrolysis, the total collagen content was determined by measuring the hydroxyproline concentration via a reaction with chloramine-T and p-dimethylaminobenzaldehyde. 30 To make the assay high throughput, the digested pellets were hydrolyzed in capped PCR plates (VWR 37001–272). To remove the yellow color, the reconstituted samples were added to a 96-well filter plate (NUNC 278011) containing activated charcoal, and the plates were centrifuged. After oxidation with chloramine-T and reaction with p-dimethylaminobenzaldehyde, the absorbance was read at 570 nm and compared to a standard of trans-4-Hydroxy-L-proline.
AP activity
Medium samples were removed from pellets and stored at −80°C until assayed. To determine AP activity, 50 μL of the medium was combined with 50 μL of p-nitrophenylphosphate substrate solution (10 mg/mL p-nitrophenylphosphate in 0.1 M glycine and 1 mM MgCl2, pH 9.6) and incubated at 37°C for 2 h. The absorbance was measured at 460 nm, and the activity was compared to the linear range of the standard curve made from p-nitrophenol in the chondrogenic medium.
Histology
All histological analyses were performed by the Purdue Histology and Phenotyping Laboratory. Pellets (n=3) were cultured for 3 weeks, fixed with 10% neutral buffered formalin, dehydrated in graded alcohol solutions, cleared in toluene, infiltrated with paraffin, and sectioned (7 μm). All sections were deparaffinized and routinely rehydrated before staining. To detect proteoglycan distribution, sections were stained in 0.1% Fast Green for 10 min, briefly placed in 1% acetic acid, and then stained in 1% Safranin-O for 1 h. To detect type I collagen, rehydrated sections were placed in hydrogen peroxide for 5 min, subjected to protein blocking for 30 min, incubated with human type I collagen monoclonal antibody (MP Biomedicals, clone I-8H5, diluted 1:100 in Dako S3022) for 60 min, incubated with a horseradish peroxidase-labeled polymer conjugated with anti-mouse secondary antibody (Dako K4001) for 45 min, and finally incubated in a substrate–chromogen solution (Dako K3468) for 10 min for visualization. Hematoxylin was used for counterstaining. A similar protocol was followed to detect type II collagen, except that sections were first treated with 0.25% trypsin for 30 min at 37°C for epitope retrieval. Sections were then treated with 1.5 IU hyaluronidase for 30 min at 37°C before being placed in hydrogen peroxide, and a different antibody was used (MP Biomedicals; clone II-4C11, diluted 1:2000). All sections were dehydrated, cleared, and permanently mounted before being examined by light microscopy.
Quantitative reverse transcription–polymerase chain reaction
Pellets were rinsed in warm PBS, placed in 1 mL TRIzol, and homogenized with syringe needles (16–22 gauge). Total RNA was isolated from the pellets according to the manufacturer's instructions and treated with DNase I (DNA-Free RNA Kit; Zymo Research). Isolated RNA was used to synthesize first-strand complementary DNA (SuperScript III First-Strand Synthesis Kit; Invitrogen). Relative expression levels of the genes were measured by qRT-PCR on Mastercycler ep realplex (Eppendorf) and normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) levels. The primer sequences for type I, II, 31 and X 32 collagens; aggrecan; SOX9; 33 cartilage oligomeric matrix protein (COMP); 7 and GAPDH 34 were obtained from references or designed using Primer-BLAST on the National Institutes of Health Website (Table 1). qRT-PCR was performed as follows: 95°C for 10 min, followed by 40 cycles of 95°C for 15 s, 60°C for 15 s, and 68°C for 20 s. The ΔΔCt method was used to calculate differences in gene expression relative to the negative controls. 35
Gene expression was measured after 3 days of culture (early gene expression) and after 1 and 2 weeks of culture (late gene expression). To obtain sufficient material, 1–5 pellets were combined to make one sample. There were 3–4 samples per experimental group. All samples were run in duplicate or triplicate.
Statistical analysis
Data are represented as a mean, and the error bars represent one standard deviation. Statistical difference between groups and the negative control was determined using a one-tailed, unpaired t-test (Excel) or a one-sided Mann–Whitney U nonparametric test (Statistical Analysis Software [SAS] version 9.2) where appropriate. A one-tailed or one-sided test was used because the BMP-2 growth factor and peptide are expected to stimulate bioactivity at or above the basal level of the negative control.7,8,12,19,22 The assumption of normal distribution was confirmed by the Shapiro–Wilk and Anderson–Darling tests for normality. All data passed normality testing, except the gene expression data, which underwent nonparametric testing. Homogeneity of variance was tested using Levene's test (SAS). All GAG and gene expression data displayed constant variance. Total collagen and AP activity displayed nonconstant variance, and the t-test was adjusted to account for unequal variance. Single-factor analysis of variance (ANOVA) and Tukey's honestly significant difference post hoc tests were performed using SAS on GAG data where indicated. For all statistical tests, a threshold value of α=0.05 was chosen, and a p-value at or below 0.05 indicated significance.
Results
Determining an effective BMP peptide concentration
To determine the concentration of the BMP peptide that affects chondrogenesis, the BMP peptide was added to a medium at concentrations ranging from 0.02 to 200 μg/mL. After 2 weeks, the GAG and DNA contents were measured. GAG production was normalized to DNA content, a relative measure of cell number, by dividing by the amount of DNA. ANOVA revealed that the concentration of the peptide has a significant effect on normalized GAG production (Fig. 1). Human MSCs cultured with ≥100 μg/mL of the peptide produced significantly more GAG than the negative control (cells cultured with no growth factors). The GAG production of human MSCs cultured with 100 μg/mL of the peptide was 74% of the positive control (cells cultured with 200 ng/mL BMP-2). Further studies were performed with 100 μg/mL of the peptide.
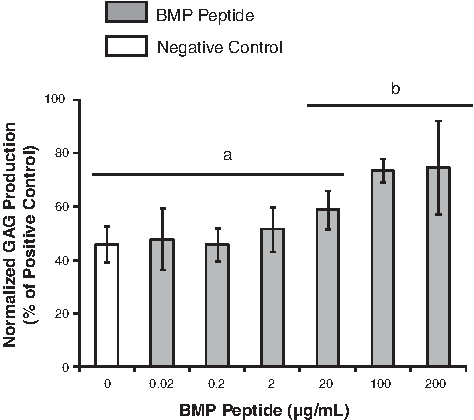
Determination of an effective bone morphogenetic protein (BMP) peptide concentration. The BMP peptide concentration had a significant effect on the normalized glycosaminoglycan (GAG) production (GAG/DNA) of human mesenchymal stem cells (MSCs) after 2 weeks in a pellet culture (single-factor analysis of variance, p<0.05). Tukey's honestly significant difference post hoc tests were performed, and data within the same group (either a or b) are not significantly different. Data (n=6) are represented as the mean±the standard deviation.
BMP peptide increases GAG and total collagen production
The ability of the peptide to promote cartilage matrix production of human MSCs was studied by measuring GAG and total collagen production. Normalized GAG production promoted by the BMP peptide was significantly higher compared to the negative control and was statistically equivalent to the BMP-2-positive control at all time points (Fig. 2A). At 4 weeks, the peptide promoted a 1.8-fold increase in GAG production compared to the negative control, and the BMP-2-positive control promoted a 2.0-fold increase. There was no significant difference between the peptide and BMP-2 measurements at any time point. With medium containing TGF-β3, no significant increase in normalized GAG was observed by adding the peptide (Fig. 2B).

The effect of the BMP peptide on
Total collagen content was measured at 1, 2, and 3 weeks (Fig. 2C). Although the BMP peptide significantly increased total collagen production per cell, the amount was significantly lower compared to the BMP-2-positive control. At 3 weeks, the peptide promoted a 1.7-fold increase in collagen production over the negative control, whereas the BMP-2-positive control produced a 7.8-fold increase.
BMP peptide does not promote large increases in AP activity
The medium was analyzed for AP activity at 1, 2, 3, and 4 weeks (Fig. 2D). As expected, the BMP-2-positive control significantly increased AP activity at each time point with a peak in activity occurring at 2 weeks. The BMP peptide did not promote a similar peak in AP activity and did not significantly increase AP activity from 1 to 3 weeks. After 4 weeks, the peptide increased AP activity 3.8-fold over the negative control; however, BMP-2 increased activity 22-fold.
BMP peptide promotes homogenous matrix distribution
Pellets were cultured for 3 weeks with the BMP peptide and compared to pellets cultured with BMP-2. Safranin-O staining confirmed the biochemical analysis, which indicated that the BMP peptide and BMP-2 promote similar amounts of proteoglycan production (Fig. 3B, C). Interestingly, the pellets cultured with BMP-2 had a heterogeneous distribution of matrix molecules, whereas those treated with peptide resulted in a more homogenous distribution. Immunohistochemical staining showed that the BMP peptide did not promote the production of type I collagen (Fig. 3F), whereas BMP-2 induced type I collagen production with a heterogeneous distribution (Fig. 3G). As expected, pellets cultured with no growth factors or peptides (negative control) showed little staining for proteoglycan (Fig. 3A) or type I collagen (Fig. 3E), whereas pellets cultured with TGF-β3 stained positively for both (Fig. 3D, H). Immunohistochemical staining for type II collagen was conducted, but only pellets cultured with TGF-β3 showed positive staining (data not shown).
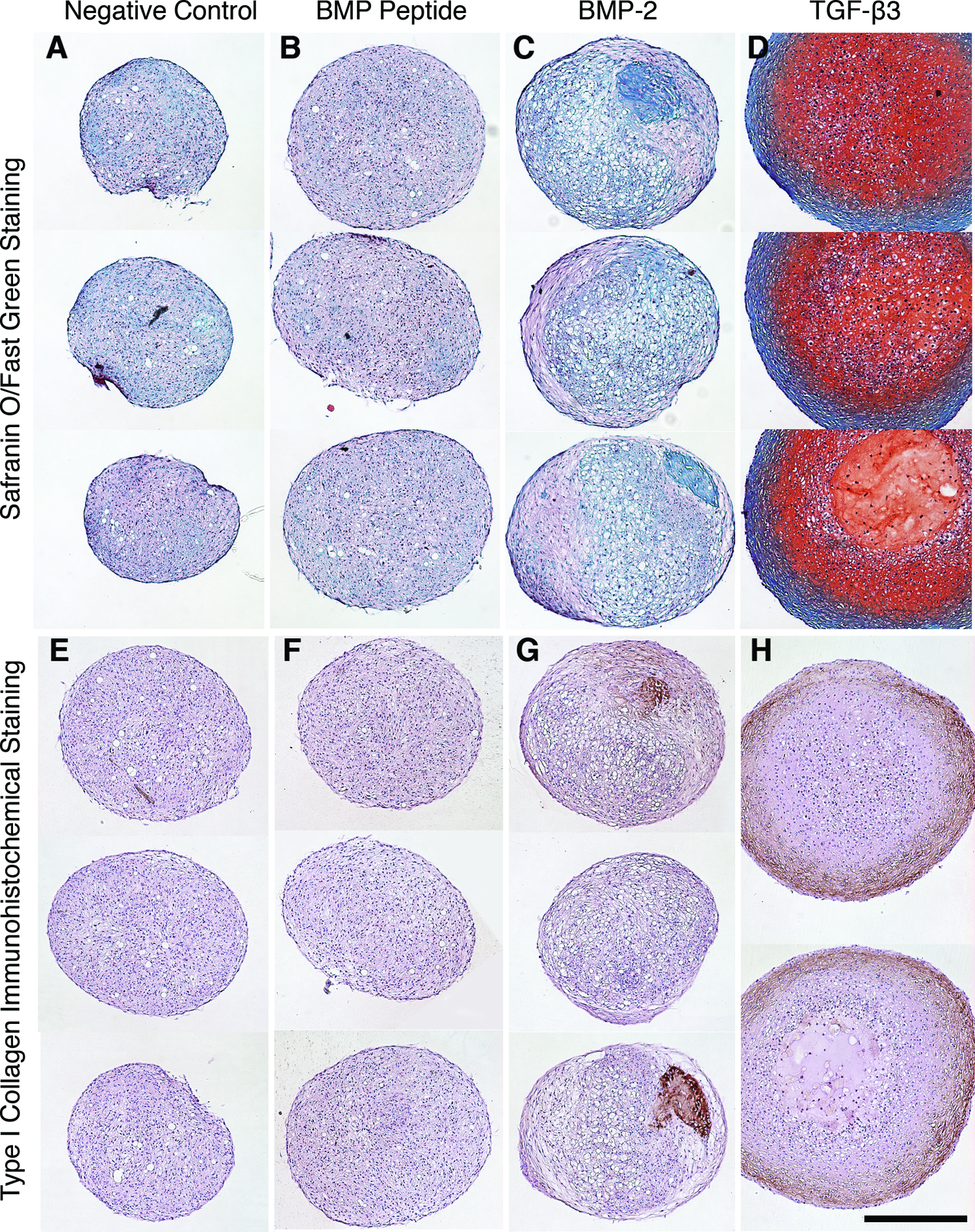
The BMP peptide promoted homogenous matrix distribution. Human MSCs were cultured for 3 weeks (n=2–3) and
BMP peptide induces gene expression at attenuated levels compared to BMP-2
Gene expression was measured after 3 days, 1 week, and 2 weeks of culture (Fig. 4). Cells were cultured without peptide or growth factors (negative control), with the BMP peptide, BMP-2, or TGF-β3. At day 3, the peptide significantly increased aggrecan gene expression 1.8-fold over the negative control, but this increase was not observed at later time points. BMP-2 directed a 350-fold increase in aggrecan gene expression at day 3 and maintained a high level of expression at later time points. The discrepancy in aggrecan gene expression levels stimulated by the peptide and BMP-2 was unexpected, given that the biochemical assay and histology indicated that similar amounts of GAG were produced. One explanation for the discrepancy could be that the qPCR analysis only tested for aggrecan gene expression, whereas the histology results nonspecifically detected all proteoglycan content.
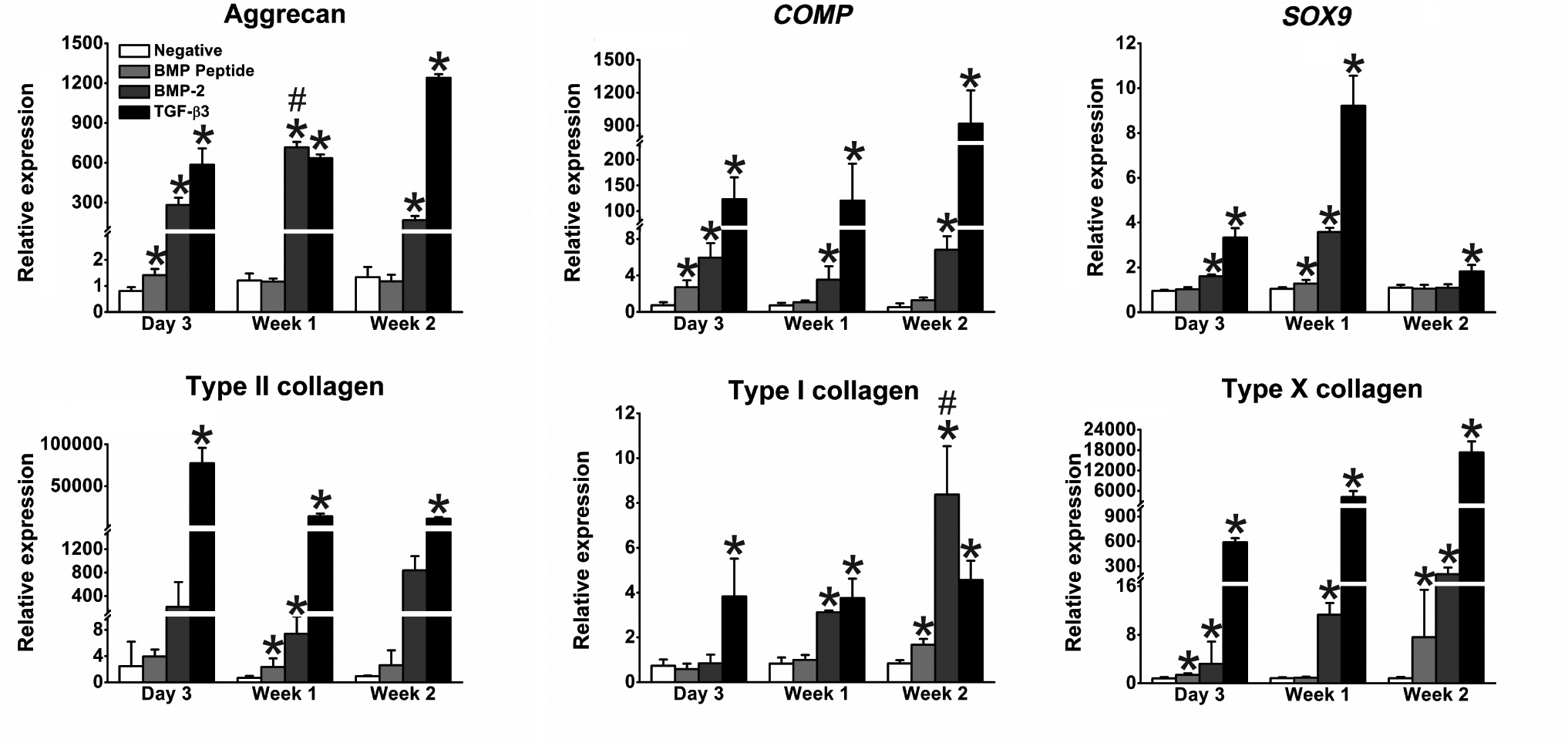
The BMP peptide significantly increased expression levels of several genes compared to negative control. After 3 days, 1 week, and 2 weeks of culture, quantitative reverse transcription–polymerase chain reaction analysis revealed that the BMP peptide had a similar, yet reduced, effect on SOX9 and type I collagen gene expression when compared to BMP-2. It should be noted that data at 3 days were gathered in a separate experiment from data at 1 and 2 weeks; thus, comparisons between these experimental time points should be made with caution. One-sided, Mann–Whitney U nonparametric tests were performed. *represents p≤0.05 compared to the negative control. # represents gene expression significantly greater than that induced by the TGF-β3 control with p≤0.05. Data (n=3–4) are represented as a mean±the standard deviation.
The peptide affected COMP gene expression similarly to the way it affected aggrecan gene expression. At 3 days, the pellets treated with peptide exhibited a 3.7-fold increase in COMP gene expression, whereas BMP-2 directed an 8.2-fold increase compared to the negative control. At 1 and 2 weeks, BMP-2 upregulated COMP significantly, whereas the peptide did not.
The peptide had a similar, though attenuated, effect on Sox9 gene expression compared to BMP-2. SOX9 gene expression was upregulated on days 3 and 7 in BMP-2-treated cells, but not at 2 weeks. A similar pattern was observed with the BMP peptide; a spike occurred at week 1 with a 1.2-fold increase over the negative control.
BMP-2 significantly increased type II collagen gene expression at all time points. The peptide significantly increased type II collagen gene expression levels at 1 week.
The BMP peptide significantly upregulated type I collagen gene expression at 2 weeks, but at a lower level than BMP-2. At 2 weeks, BMP-2 promoted a 10-fold increase in type I collagen gene expression compared to the negative control, which was significantly higher than the type I collagen gene expression induced by TGF-β3. This result is not unexpected given the role of BMP-2 in bone formation.
BMP-2 significantly promoted type X collagen gene expression at all time points with the largest increase occurring at 2 weeks (250-fold over the negative control). The BMP peptide resulted in significant increases at 3 days and 2 weeks, with the largest increase occurring at 2 weeks (9.2-fold increase over the negative control).
Discussion
BMP-2 has been shown to promote chondrogenesis of human MSCs in vitro7–9 and encourage in vivo articular cartilage formation.10,11 Yang et al. recently showed that the combination of microfracture and long-term delivery of BMP-2 promoted the regeneration of hyaline-like cartilage in a rabbit model. 11 A short-peptide mimic of BMP-2 would be an attractive alternative to BMP-2 in cartilage tissue engineering because recombinant BMP-2, which consists of two 115-amino-acid-long polypeptide chains linked by a disulfide bond, is more expensive and difficult to manufacture and incorporate into materials. The BMP-derived peptide investigated in this study is composed of 20 amino acids and does not require additional processing, such as disulfide bond formation or complex folding regimens.
Although some studies found the peptide to be active at 200 ng/mL (0.1 μM),22,36 our results agree with other studies showing that the peptide is only active at concentrations ranging from 100–200 μg/mL (47–94 μM).23,25 For comparison, BMP-2 is typically used at 50–500 ng/mL (0.002–0.02 μM). In our studies, the peptide was effective at 47 μM, which was 6,100 times higher than the BMP-2 concentration (0.008 μM). Despite this concentration difference, the cost of using the peptide in culture was comparable to the cost of using BMP-2. We envision that the peptide can be grafted to a tissue-engineered scaffold, which would further reduce the cost of using the peptide, since it would not be included in the medium, which is periodically replenished. Previous studies demonstrated that the peptide is still active when grafted to a surface.22,25,36
Both histological and biochemical analyses demonstrated that in the absence of growth factors, the peptide promoted similar amounts of GAG, but lower levels of collagen compared to BMP-2. The peptide promoted a homogenous distribution of GAG, whereas BMP-2 promoted a heterogeneous distribution, as evidenced by the varying intensity of staining (Fig. 3C). Two pellets cultured with BMP-2 had dense regions of type I collagen, which varied in size, whereas the third pellet did not have this feature (Fig. 3G). This heterogeneity may explain the high level of variance in the total collagen measurements (Fig. 2C) and the collagen gene expression levels (Fig. 4) for BMP-2. From a tissue-engineering perspective, an advantage of the peptide over BMP-2 is the expected homogenous and predictable distribution of matrix molecules in a construct.
TGF-β3 and BMP-2 synergize and result in enhanced GAG production.7,9,12 Given the similar GAG levels produced by BMP-2 and the peptide, we hypothesized that the peptide would exhibit a similar behavior as BMP-2 when added with TGF-β3. To test this hypothesis, TGF-β3 was added to all groups, and the GAG content was measured at 1, 2, 3, and 4 weeks (Fig. 2B). Surprisingly, the peptide did not enhance GAG production, although BMP-2 did. These results suggest a different mechanism for bioactivity of the peptide. Further exploration of the mechanism of this peptide is needed; however, since our data suggest that the peptide acts through a different biochemical mechanism, it is possible that using the peptide may avoid some of the adverse effects of BMP-2 that are currently observed clinically.37,38
One concern of using BMP-2 is that, in our studies as well as others,7,32,39 BMP-2 increased chondrogenic markers as well as hypertrophic markers such as AP activity (Fig. 2D) and type X collagen gene expression (Fig. 4). Our study demonstrated that the peptide did not increase these hypertrophic markers to as large an extent as BMP-2 and maintained a cellular phenotype more reminiscent of cartilage. In our studies, BMP-2 promoted AP activity at early time points, and the activity peaked at 2 weeks. Previously published literature showed that the peptide promoted AP activity in 2D at early time points (3 days and 1 and 2 weeks).19,22,25 Because we expected the peptide to behave similarly to BMP-2 and because of the previous results with the peptide in 2D systems, it was surprising that the peptide did not promote AP activity in the pellet culture system at early time points, resulted in a minimal response at a late time point (4 weeks), and did not display a peak in activity over the culture time. The peptide also did not upregulate type X collagen gene expression to the extent that BMP-2 did, but it was able to maintain similar levels of GAG production. In addition, the peptide upregulated gene expression of aggrecan, COMP, and collagen II, which are key components of the cartilage matrix. Given the importance of maintaining the cartilage phenotype and suppressing chondrocyte hypertrophy, the BMP peptide is an attractive tool in cartilage tissue engineering.
The concentrations of GAG and type II collagen vary between zones of cartilage. The BMP peptide studied here induced GAG production at levels similar to BMP-2, but did not induce similar amounts collagen. The BMP peptide could be useful in engineering cartilage tissue that has reduced levels of collagen, such as in the deep zone cartilage. 40 Therefore, the peptide could potentially be incorporated in patterned, 3D grafts that mimic the zonal architecture of native cartilage.41–43
In conclusion, this study has shown that the BMP peptide promoted a small amount of collagen production and promoted GAG production in human MSCs at levels similar to BMP-2. Unlike BMP-2, the BMP peptide encouraged matrix accumulation without extensive upregulation of hypertrophic markers, which makes the peptide a valuable tool for differentiation of human MSCs in cartilage engineering. Future directions include tethering the BMP peptide to a 3D matrix and studying its effect on chondrogenesis.
Footnotes
Acknowledgments
This work was supported by the Purdue School of Chemical Engineering and the College of Engineering and the National Science Foundation (Award No. 0927100-EEC and a Graduate Fellowship to J.N.R.). The National Cancer Institute Biometric Research Branch Preclinical Repository kindly provided bFGF. We thank Carol Bain at the Purdue Histology and Phenotyping Laboratory for performing all histology and Kevin M. Cherry for his editorial assistance.
Disclosure Statement
No competing financial interests exist.