
Abstract
Introduction:
Current cartilage repair procedures using autologous chondrocytes rely on a variety of carriers for implantation. Collagen types I and II are frequently used and valuable properties of both were shown earlier in vitro, although a preference for either was not demonstrated. Recently, however, fibrillar collagens were shown to promote cartilage degradation. The goal of this study was to evaluate the effects of collagen type I and type II coating on chondrogenic properties of in vitro cultured human chondrocytes, and to investigate if collagen-mediated cartilage degradation occurs.
Methods:
Human chondrocytes of eight healthy cartilage donors were isolated, expanded, and cultured on culture well inserts coated with either collagen type I, type II, or no coating (control). After 28 days of redifferentiation culture, safranin O and immunohistochemical staining for collagen types I, II, X, and Runx2/Cbfa1 were performed and glycosaminoglycan (GAG) and DNA content and release were examined. Further, expression of collagen type I, type II, type X, MMP13, Runx2/Cbfa1, DDR2, α2 and β1 integrin were examined by reverse transcriptase-polymerase chain reaction.
Results:
The matrix, created by chondrocytes grown on collagen type I- and II-coated membranes, resembled cartilage more than when grown on noncoated membranes as reflected by histological scoring. Immunohistochemical staining did not differ between the conditions. GAG content as well as GAG/DNA were higher for collagen type II-coated cartilage constructs than control. GAG release was also higher on collagen type I- and II-coated constructs. Expression of collagen type X was higher of chondrocytes grown on collagen type II compared to controls, but no collagen X protein could be demonstrated by immunohistochemistry. No effects of collagen coating on DDR2 nor MMP-13 gene expression were found. No differences were observed between collagen types I and II.
Conclusion:
Chondrocyte culture on collagen type I or II promotes more active matrix production and turnover. No significant differences between collagen types I and II were observed, nor were hypertrophic changes more evident in either condition. The use of collagen type I or II coating for in vitro models, thus, seems a sound basis for in vivo repair procedures.
Introduction
However, recently, a host of literature has suggested that the presence of native fibrillar collagens in the vicinity of chondrocytes may induce a catabolic response leading to cartilage degradation. Collagen II coating of cell culture surfaces induced upregulation of the discoidin domain receptor 2 (DDR 2) and matrix metalloproteinase 13 (MMP 13) expression in human chondrocytes, 9 which are thought to induce a downward spiral of matrix degradation and hypertrophy.10,11 Denatured collagen was not capable of inducing a suchlike response. 12
To further elucidate the role of collagen as chondrocyte carrier, the aim of the current study was to evaluate the effects of collagen type I and II on overall chondrogenesis, DDR2 activation, and hypertrophic differentiation.
Methods
Chondrocyte isolation and expansion
Healthy human femoral cartilage was obtained postmortem of four male donors and four female donors. Five cartilage samples were Collins grade 0, and 3 samples were grade 1. The mean age of the patients was 67 years (range: 47 years–83 years). Tissue samples were used with patients' informed consent and through institutional approval (University Medical Center Utrecht). The cartilage was digested overnight in the Dulbecco's modified Eagle's medium (DMEM) (42430; Invitrogen) containing 0.1% collagenase (CLS2; Worthington) and 1% penicillin/streptomycin (15140; Invitrogen). Isolated chondrocytes were washed three times in phosphate-buffered saline (PBS) and expanded at 5000 cells/cm2 in monolayer in two passages in DMEM containing 1% penicillin/streptomycin, 10% fetal bovine serum (FBS, DE14-801F; Cambrex Bio Science Verviers), and 10 ng/mL bFGF (223-FB; R&D Systems). At 80% confluency, cells were trpysinized and passaged, and used at passage 2 for redifferentiation.
Redifferentiation culture
As an in vitro model for regeneration, a 24-well transwell system was used as described previously. 13 After two passages, chondrocytes were washed three times in PBS and seeded on Millipore culture inserts (Millipore BV) with hydrophilic poly-tetrafluoroethylene membrane at 720.000 cells/filter (corresponding with a total DNA amount of 8.4×106 pg DNA/filter) 14 and cultured in DMEM supplemented with 2% ITSx (51500; Invitrogen), 2% ascorbic acid (A8960; Sigma-Aldrich), 2% human serum albumin (HS-440; Seracare Life Sciences), 1% penicillin/streptomycin, and 10 ng/mL TGF-beta2 (302-B2; R&D Systems). Cells were cultured on the inserts for 28 days in duplo as described earlier.13,15 The insert membranes were coated with 0.125 mg/mL collagen type I in 0.1 M acetic acid (C9791; Sigma-Aldrich) or 0.125 mg/mL collagen type II in 0.1 M acetic acid (C9301; Sigma-Aldrich). Control inserts were treated with 0.1 M acetic acid (control). Efficient collagen coating was verified by immunohistochemistry for collagen types I and II (Supplementary Figs. S1 and S2; Supplementary Data are available online at www.liebertpub.com/tea). Before seeding with chondrocytes, all filters were washed for 30 min with sterile PBS. At 28 days of culture, regenerated tissues were washed three times in PBS and fixed in 10% neutral buffered formalin or snap-frozen in liquid nitrogen and stored at −20°C for GAG analysis or at −80°C for RNA analysis.
Histological analysis
Histological evaluation was performed on 5-μm deparaffinised sections using safranin O/fast green staining as described by Rosenberg. 16 Histological quality was assessed using the Bern Score, 17 a score designed specifically for tissue-engineered cartilage, assessing darkness and uniformity of Safranin O staining, the amount and organization of the extracellular matrix and cell morphology. The maximum score is 9, representing healthy hyaline cartilage.
Immunohistochemistry
Immunohistochemical staining was performed for collagen types I, II, and X, and core-binding factor α1 (Cbfa1/Runx-2) as markers of hypertrophic- and re-and dedifferentiation. To this end, sections were incubated with PBS/BSA 5% (collagen type I, type II, and X) and incubated in 10% egg white, rinsed, and then blocked with 5% skim milk in TBS-T (Cbfa1) to correct for a specific binding, then incubated overnight at 4°C with monoclonal antibodies against collagen type I (20 μg/mL, CP-17; Calbiochem), collagen type II (1:100, ascites II-II6B3, Developmental Studies Hybridoma Bank (DSHB)), collagen type X (1:50×53; cell culture supernatant, Quartett, Germany), and Cbfa 1 (1.4 μg/mL, D130-3; MBL, Woburn, USA in 5%PBS/BSA. Subsequently, the samples were incubated with a biotinylated sheep anti-Mouse (1:200, RPN1001V; GE Healthcare) followed by peroxidase-labeled streptavidin (1:500; IM0309 Beckman Coulter) (Collagen I and X) or with goat anti-Mouse HRP (1:100; P0447 DAKO)(Collagen II and Runx2/Cbfa1) all for 1 h at room temperature in 5% PBS/BSA. Diaminobenzidine was used to obtain a signal. Juvenile growth plate cartilage was used as a positive control for type X and cbfa1 production. Sections were incubated with isotype antibodies as negative controls. Finally, the sections were counterstained with hematoxylin.
Alcian blue precipitation and DNA assay
In vitro constructs were digested in 2% papain (P3125; Sigma-Aldrich), in 50 mM phosphate buffer, 2 mM N-acetylcysteine, and 2 mM Na2-EDTA (pH 6.5) at 65°C for 2 h. Part of the digest was used to measure DNA content and part was used for the quantification of glycosaminoglycan (GAG) content as a measure of proteoglycan content, using an Alcian Blue precipitation assay (described below).
GAGs were precipitated from the chondrocyte digests as well as from the culture medium (at each medium change) and stained with an Alcian blue dye solution (Alcian blue 8GX, 05500; Sigma-Aldrich), saturated in 0.1 M sodium acetate buffer, containing 0.3 M MgCl2 (pH 6.2) for 30 min at 37°C. 18 The blue staining of the medium was quantified photospectrometrically from the change in absorbance at 620 nm, using chondroitin sulfate (Sigma) as a reference. DNA was stained with the fluorescent dye Hoechst 33258 (Sigma) and fluorescence was measured on the Cytofluor (Bio-Rad), 19 using calf thymus DNA (Sigma) as a reference.
Polymerase chain reaction
For RNA isolation, all steps were performed on dry ice using instruments pretreated with RNAzap and distilled water. After separation of the cartilage from the transwell membranes using a scalpel (Swann-Morton), the cartilage samples were crushed in a 2-mL Eppendorf tube and pestled using liquid nitrogen with RLT-mercapto-ethanol buffer and a 21G syringe until a homogenous mass was obtained. RNA isolation was done using the RNEasy Micro Kit (74004; Qiagen) according to the manufacturer's protocol using DNAse treatment. RNA concentration was measured spectrophotometrically using the NanoDrop® ND-1000 spectrophotometer (NanoDrop Technologies), sufficient RNA purity was assumed if the 260/280 nm ratio was higher than 1.9. Subsequently, 500 ng of RNA was reverse transcribed using the iScript cDNA synthesis Kit (Bio-Rad). Reverse transcriptase-polymerase chain reaction (PCR) was performed according to the manufacturer's protocol using the following TaqMan® Gene Expression Assays (Applied Biosystems): Collagen type I (Hs00164004_m1), Collagen type II (Hs00264051_m1), Collagen type X (Hs00166657_m1), Runx2/Cbfa1 (Hs00231692_m1, recognizes all three isoforms of Runx2), MMP 13 (Hs00942589_m1), DDR2 (Hs00178815_m1), α2 integrin (ITGA2, Hs00158127_m1), and β1 integrin (ITGB1, Hs00559595_m1), corrected for beta-actin (Hs99999903_m1). All analysis was performed in technical duplicates except for RNA quantification, which was performed in triplicate.
Statistics
Statistical analyses were performed using the statistical software SPSS 15.0 for Windows (SPSS, Inc.). The comparison between the three types of coatings was performed using repeated measurement analysis. Gene expression comparison for collagen type X was compared using the Friedmans test. After performing a Bonferroni correction, p≤0.05 (*) was considered statistically significant. All figures show mean values±standard error of the mean. Histological slides shown are representative of all donors.
Results
Bern score
Macroscopically, chondrocytes grown on noncoated membranes formed a more irregularly shaped cartilage matrix than cartilage generated on collagen type I- or type II-coated inserts. Histologically, 6 out of 8 tissue cultures grown on noncoated inserts demonstrated a more aggregated appearance.
Histological quality of in vitro cultured cartilage (Bern score) was higher for collagen type I and collagen II versus control (p<0.01 and p=0.05, respectively) (Fig. 1a, b).
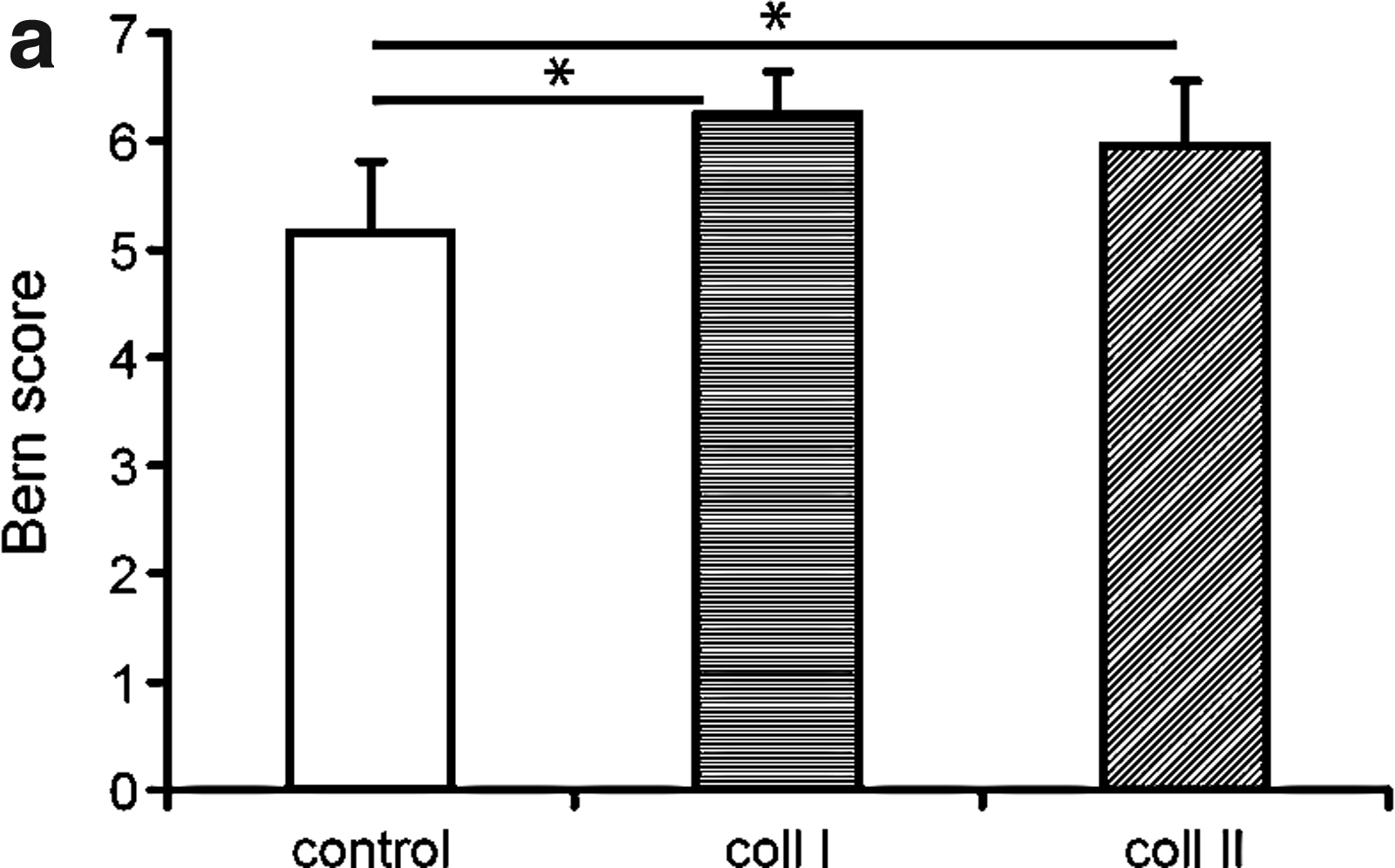
Histological scoring of the in vitro generated cartilage using the Bern score (16).
Immunohistochemistry
Immunohistochemistry for collagen type I and II demonstrated a variable strong staining in all conditions; however, this seemed donor-specific rather than dependent on coating and no significant differences were observed. No collagen type X production was observed in any of the conditions. Low Cbfa1 staining was observed under all conditions (Fig. 2).
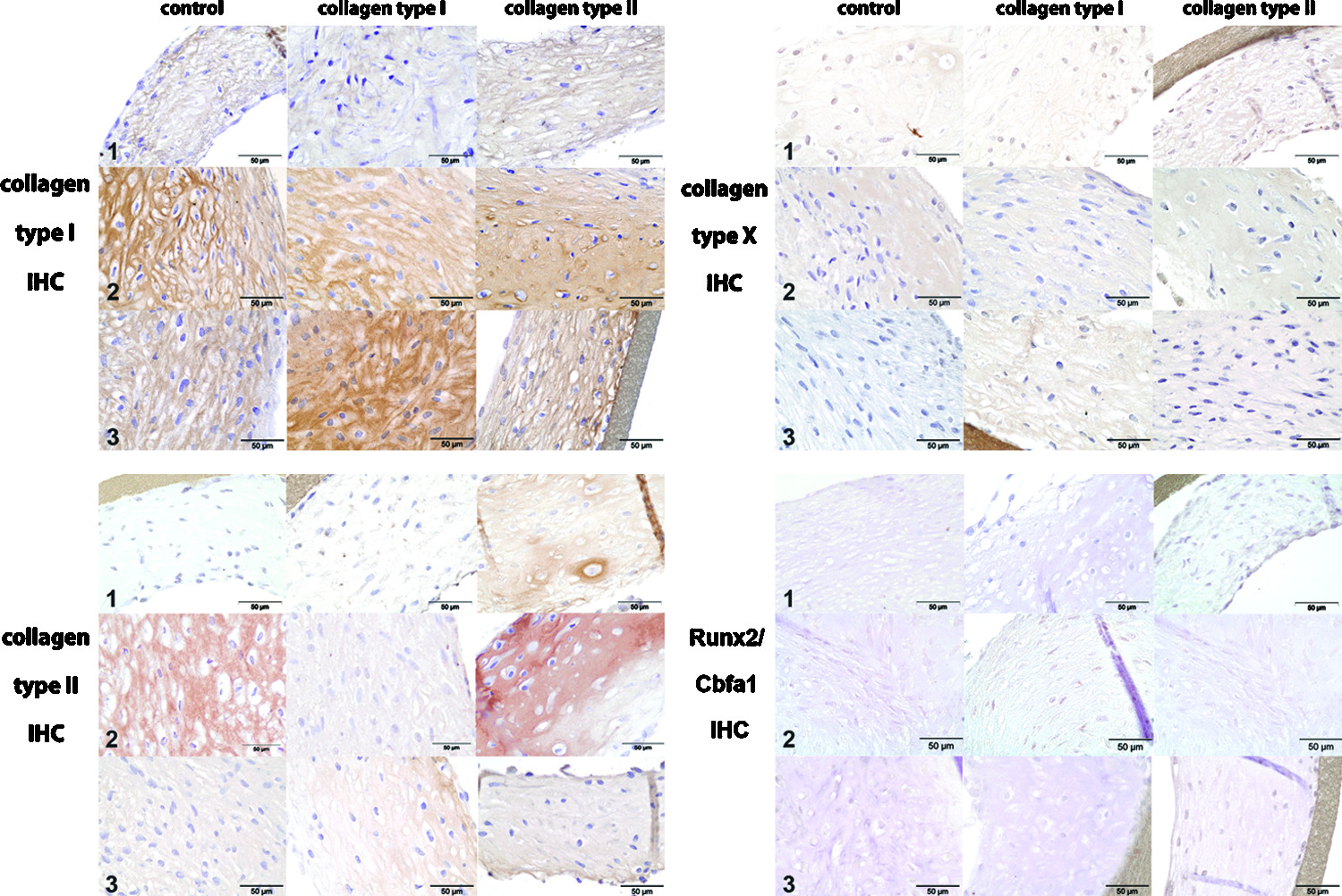
Immunohistochemical staining for collagen types I, II, collagen type X, and Runx2 at the end of the 28-day in vitro culture for 3 human chondrocyte donors at 400× magnification. Collagen type I and II staining were more donor-dependent than dependent of the coating conditions and no differences between either condition was detected. Immunohistochemical staining for collagen type X and Runx2/Cbfa1 was low or absent for all conditions in all donors. Color images available online at www.liebertpub.com/tea
GAG and DNA content
No visible loss of cells was noted after seeding of the inserts immediately after plating, nor after the first medium change. Upon comparison of the separate coating procedures after 28 days, DNA content did not differ significantly between the conditions (10.5×106 pg DNA/filter for collagen type I-coated filters (corresponding with 910.000 cells), 10.9×106 pg DNA/filter (corresponding with 940.000 cells) for collagen type II-coated filters, and 8.2×106 pg DNA/filter (corresponding with 710.000 cells) for control filters; p=0.16). Total GAG content was higher in tissues grown on collagen type II (80 μg±11 μg) compared to cells cultured on uncoated scaffolds (50 μg±11 μg; p<0.01). The amount of GAG normalised per DNA was higher for collagen type II-coated scaffolds compared to controls (p=0.02) (Fig. 3a). Total GAG release was higher on collagen type I (139 μg±33 μg; p<0.01)- and II-coated filters (116 μg±26 μg; p=0.05) versus control. Total proteoglycan production (final GAG content+total amount of GAG released during culture) was higher for collagen type I versus control (p=0.01) and collagen II versus control (p=0.02). When release was expressed as percentage of total production, no differences were observed between collagen type I or II and controls (p=0.60) (Fig. 3b).
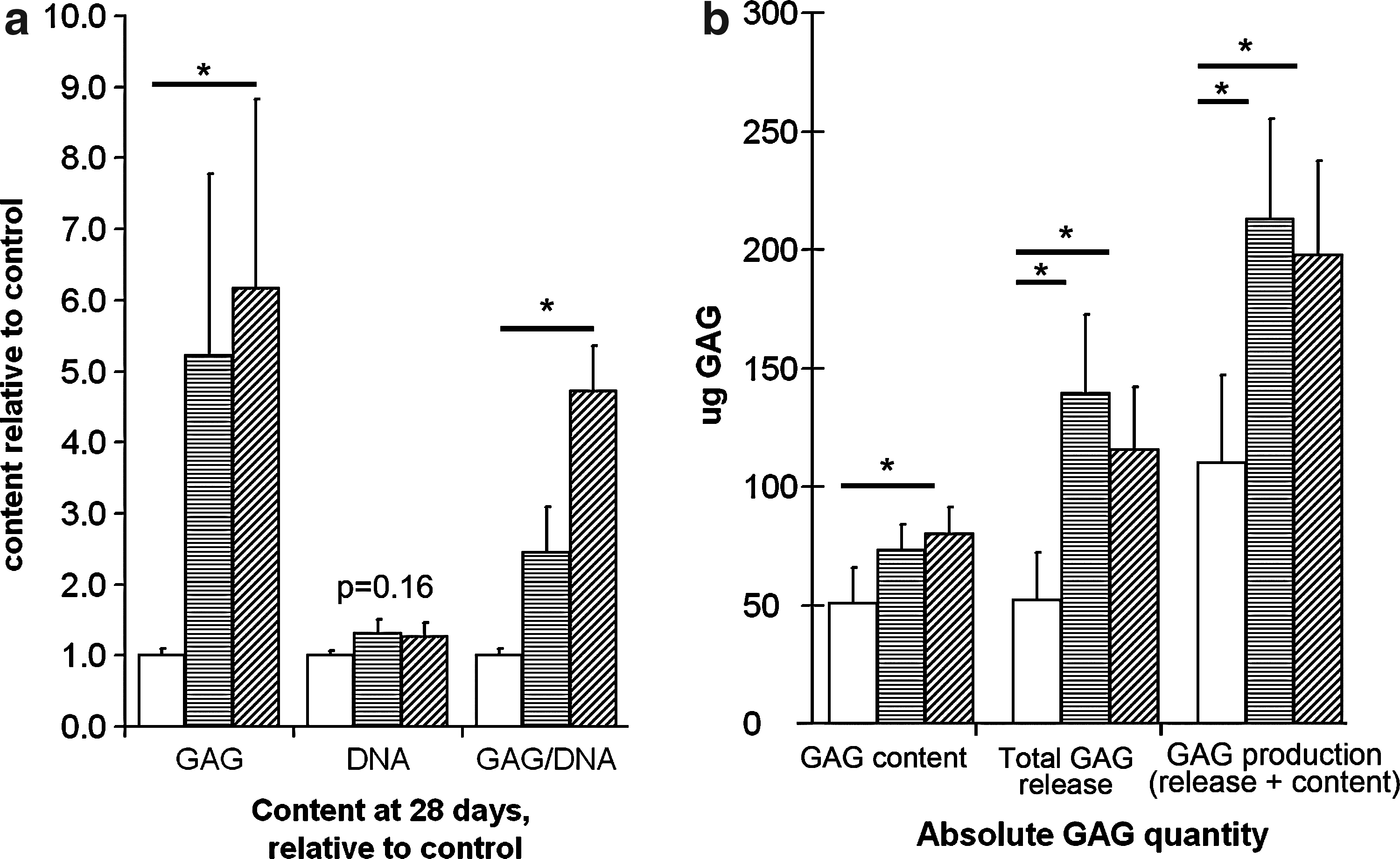
Glycosaminoglycan (GAG) and DNA content of the in vitro engineered cartilage grown on collagen type I- or II-coated inserts.
PCR extracellular matrix and hypertrophic markers
Collagen type X expression was higher for collagen type II versus control (p=0.03). Collagen type II expression was nonsignificantly higher in type II-coated constructs (p=0.09). DDR2 expression appeared higher for collagen types I and II, although this did not reach statistically significant levels. No differences were observed in expression of collagen type I, collagen type II, MMP-13, Runx2/Cbfa1, α2 integrin and β1 integrin expression (Fig. 4).
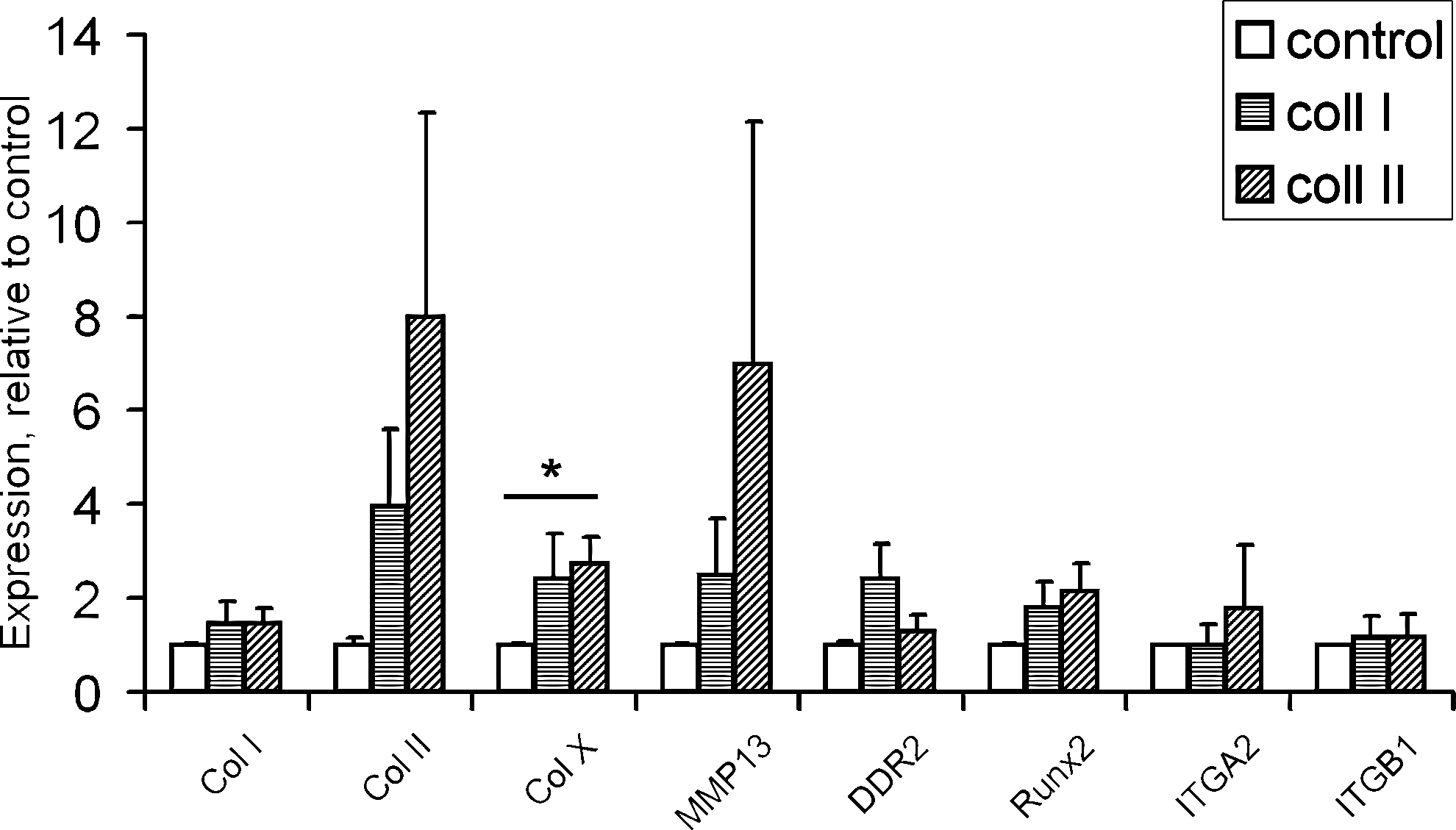
RNA expression of collagen type I, type II, and type X, MMP13, DDR2, Runx2/Cbfa1, α2 integrin (ITGA2), and β1 integrin (ITGB1) at the end of the 28-day redifferentiation culture. Collagen type X expression was higher in chondrocytes cultured on type II-coated constructs (p=0.03), than in chondrocytes cultured on noncoated constructs. For none of the other genes, significant differences in expression were found between control, collagen type I, or collagen type II coating. Asterisk refers to p-values.
Discussion
The current study aimed at determining the effect of collagen I and II on chondrocyte regeneration, with respect to extracellular matrix production and degradation, but also to examine possible activation of DDR2-mediated degradation. Cartilage regenerated on collagen type II was characterized by a higher final GAG content, a higher GAG/DNA ratio and morphologically resembled hyaline-like cartilage more than cartilage generated on noncoated constructs, although no clear difference was observed between collagen types I and II. Further, total GAG production was higher in the collagen-coated scaffolds compared to controls. Although the release of GAGs into the culture medium was also higher, the release expressed as percentage of total production did not differ significantly, thus suggesting that the increased release into the medium was partly due to the release of newly produced GAGs that could not be retained in the matrix, rather than degraded GAGs. At the protein level, collagen production appeared to vary between the donors, which was reflected by the mRNA levels. Hypertrophic differentiation was not evident at the protein level either, although at the mRNA level, collagen type X gene expression was increased on coated scaffolds. However, MMP-13 expression was not increased, which was in line with the lack of a clear upregulation of DDR2 in the current study. MRNA for α2 and β1 integrins was clearly detected, but was not affected by culture on the different coatings.
To what extent the collagen-mediated stimulatory effect on GAG production was collagen-specific cannot be stated with full certainty. The effect may be related to mere cell attachment to collagen as proteinaceous substrate. Integrin-mediated attachment and signaling could also be responsible 7 as α2 and β1 integrin were clearly shown to be expressed in the current study. However, even if these integrins would have been present at the protein level, inhibition of their expression would have provided firm evidence for this explanation. The differential morphology of cartilage grown on noncoated inserts suggested that a lack of cell–matrix or cell–protein adherence may have led to some aggregation of cells, a principle used in pellet culture models of chondrogenic regeneration. In a previous study, however, no differences in cartilage matrix production were observed 20 between these two culture models, suggesting the collagen coatings as such had played a role in improving cartilage regeneration rather than the inhibition of aggregation. The collagen types I and II used in the current study were from bovine and chicken origin, respectively. Although ideally, collagens from the same animal source would have been used, the currently used collagens both have a long history in in vitro cell culture use.15,20–24 However, differential effects using collagens from other species cannot be excluded.
Despite the recently postulated effects of fibrillar collagens on cartilage degradation, no significant upregulation of DDR2 or its downstream target gene MMP-13 was observed, nor did GAG degradation during culturing seem to be affected by the use of fibrillar collagens for coating. Although collagen release was not determined in the current study, the lack of any difference in MMP-13 levels and in GAG content suggests the absence of an effect on collagen degradation. Although no DDR2 activation was noted in the current study, this may have occurred during the early culture periods, its expression wearing off by the end of the period. Even so, apparently, these effects are not strong enough to affect the net production of cartilaginous tissue, which was actually highest in collagen type I- and II-coated constructs. While the increase in collagen type X expression on collagen type II-coated scaffolds suggested some hypertrophic differentiation, at the histological level this was difficult to confirm. However, a discrepancy between mRNA levels and protein production frequently occurs,25,26 with sometimes large effects on mRNA levels being translated into smaller or even opposite effects on protein production.25,27,28 Overall, the high interdonor variability in immunohistochemical staining for collagen I and II is in accordance rather than in contrast with the mRNA levels found.
Why, in contrast with previous results, 4 no clear difference in matrix production was observed between the two collagen types is not clear, but was reflected in the α2β1 integrin receptor expression, which was similar in all conditions at 28 days. However, expression may have been different at earlier stages of culture, or when the chondrocytes first came into contact with the collagens. The common drawback of in vitro studies is that their outcome is usually based on the results of one or a limited number of time points, which for cartilage regeneration is usually limited to a period of 21–28 days. It is not known at which time point in vitro regeneration reflects the final effects found in vivo, typically analyzed several months or years after implantation. It would be worthwhile to study the relation between regeneration in vitro and in vivo and further define which stage of in vitro regeneration best predicts in vivo cartilage formation. In addition, the lack of more time points in these and other studies also precludes a clear picture of the course of the redifferentiation process, which may be suboptimal and reflect fibrocartilage formation. 29 However, given the identical starting point of this study, that is, dedifferentiated chondrocytes, it can be concluded that collagen coating positively affected their phenotype at least in the course of the 4 weeks studied.
Another explanation for the difference observed between collagen I and II in the study by Nehrer et al., 4 may be the more extensive cell–matrix contact area between the 3D collagen gel and the canine chondrocytes in this particular study. This may have been responsible for a more cell surface receptor–matrix interaction, thereby maintaining specific cell characteristics.
Although the currently used filter culture model is a 2D rather than a 3D model, also MACI procedures are in essence 2D culture conditions, as commonly cells seeded on carriers attach to the surface of carrier pores rather than being surrounded by the materials as is the case with, for example, hydrogel encapsulation. As on MACI membranes, a multilayered cell sheet is formed in the current culture model.30–34 However, most MACI procedures employ a collagen type I/III membrane, and therefore the current culture setup is not completely comparable.
In conclusion, fibrillar collagens type I and II do not seem to have any lasting negative influences on in vitro cultured human chondrocytes with regard to synthesis of tissue-engineered cartilage. Collagen type II coating seems slightly preferable to collagen type I, although both collagen types are preferable to no collagen coating. Future studies may include analysis of the effects of collagen type III, which is currently also used in MACI constructs.
Footnotes
Acknowledgments
The authors wish to acknowledge the Anna Foundation for Musculoskeletal Research in The Netherlands, the Netherlands Organisation for Health Research and Development (NWO) and the Dutch Arthritis Association (Reumafonds) for their continuous support. The authors further wish to thank the Pathology department of the UMC in Utrecht for their assistance during acquisition of cartilage samples as well as P. Westers, Ph.D., for his statistical advice.
Disclosure Statement
The authors, hereby declare that they have no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.