
Abstract
Mesenchymal stromal cells (MSCs) can be isolated not only from bone marrow (BM) but also from other tissues, including adipose tissue (AT) and umbilical cord Wharton's Jelly (WJ). Thanks to their ability to differentiate into various cell types, MSC are considered attractive candidates for cell-based regenerative therapy. In degenerative clinical settings, inflammation or infection is often involved. In the present work, we hypothesized that an inflammatory environment and/or Toll-like receptor (TLR) ligation could affect the MSC differentiation potential. MSC were isolated from BM, AT, and WJ. Inflammation was mimicked by a cytokine cocktail, and TLR activation was induced through TLR3 and TLR4 ligation. Osteogenesis was chosen as a model for differentiation. Osteogenic parameters were evaluated by measuring Ca2+ deposits and alkaline phosphatase (ALP) activity at day 7, 14, and 21 of the culture in an osteogenic medium. Our results show that WJ-MSC exhibit a much lower osteogenic potential than the other two MSC types. However, inflammation was able to strongly increase the osteogenic differentiation of WJ-MSC as calcification, and ALP activity appeared as early as day 7. However, this latter enzymatic activity remained much lower than that disclosed by BM-MSC. TLR3 or TLR4 triggering increased the osteogenesis in AT- and, to lesser extent, in BM-MSC. In conclusion, WJ-MSC constitutively disclose a lower osteogenic potential as compared with BM and AT-MSC, which is not affected by TLR triggering but is strongly increased by inflammation, then reaching the level of BM-MSC. These observations suggest that WJ-MSC could constitute an alternative of BM-MSC for bone regenerative applications, as WJ is an easy access source of large amounts of MSC that can effectively differentiate into osteoblasts in an inflammatory setting.
Introduction
BM as a source of MSC is endowed with some drawbacks, including the intrusive method of BM aspiration, the limited in vitro expansion capacity, the high degree of viral infection, and the decreased cell number and proliferative/differentiation capacity with age.8,9 Therefore, alternative MSC sources are of great interest. Adipose tissue (AT) and umbilical cord matrix or Wharton's jelly (WJ) represent the main alternative sources due to a greater accessibility and a higher expansion potential. The MSC generated from both these sources fulfill the International Society for Cellular Therapy (ISCT) criteria for MSC, that is, negativity for hematopoietic and endothelial marker expression (such as CD11b, CD14, CD31, CD34, CD45, and human leukocyte antigen [HLA]-DR) and positivity for other markers, including CD105, CD73, CD29, and CD9. 10 AT- and WJ-MSC are able to differentiate into adipocytes, chondrocytes, and osteoblasts and present immunosuppressive properties.11,12
MSC express active Toll-like receptors (TLRs).13–15 TLR compose a family of germline-encoded pattern-recognition receptors that have evolved to detect different components of foreign pathogens, referred to as pathogen-associated molecular patterns, and to promote the activation of immune cells. 16 Exogenous danger signals released after microbial infections activate the TLR in innate immune cells, and mount an appropriate immune response that re-establishes homeostasis.16,17 MSC from different sources display different patterns of TLR expression. 18 Moreover, the TLR triggering of MSC differentially affects their immunosuppressive function according to the source from where they were isolated. 18 Besides TLR triggering, inflammation also affects both the TLR pattern expressed by MSC and their immunosuppressive function. Interestingly, in an inflammatory or infectious environment, the expression of adhesion molecule activity and galectin-1, lectin involved in the regulation of T-cell proliferation, by MSC was differently modulated. 19 Recently, we observed that neither TLR3/TLR4 ligation nor inflammation changed the WJ-MSC ability to suppress T-cell proliferation contrary to BM-MSC. 18
In the present work, we speculated that inflammation and/or TLR triggering could differentially affect the differentiation potential of the MSC isolated from different sources. Consequently, we focused our work on the study of the osteogenic potential of the MSC isolated from BM as compared with AT and WJ. We further tested the influence of inflammation and TLR3 or TLR4 triggering on the osteogenic differentiation of the MSC isolated from these three sources.
Materials and Methods
Ethics statement
This study has been specifically approved by the Bordet Institute Ethics Committee and conducted according to the principles expressed in the Declaration of Helsinki. All samples were collected after written informed consent had been obtained.
Isolation and culture of BM-MSC
BM was harvested from the sternum or iliac crest of healthy volunteers (n=3). Mononuclear cells were isolated by density gradient centrifugation (LinfoSep; Biomedics), washed in Hank's buffered salt solution (Lonza Europe), and seeded at 2×104 cells/cm2 in Dulbecco's modified Eagle's medium-low glucose (DMEM-LG, Lonza) supplemented with 15% of fetal bovine serum (Sigma-Aldrich), 2 mM of
Isolation of WJ-MSC
Umbilical cords from full-term deliveries were collected and processed as previously described (n=3). 20 Immediately after collection, the umbilical cords were transferred to an aseptic saline buffer. The umbilical cord segments (5–10 cm) were sectioned longitudinally to expose the WJ. Some incisions were made on the matrix with a sterile scalpel to expose a wider area of tissue contact with the plastic surface. The MSC isolation method is based on their migratory and adhesive properties. The WJ matrix was completely immersed in DMEM-LG culture medium for 5 days to allow the MSC to spread out and attach to the plastic surface. At the end of the culture period, the umbilical cord fragments were removed from the plates, and the cells were cultured until confluence.
Isolation of AT-MSC
AT was obtained from patients undergoing liposuction procedure (n=3). Fresh lipoaspirates used to isolate AT-MSC were processed according to a previous protocol. 21 Briefly, lipoaspirates were intensively washed with an equal volume of Dulbecco's phosphate-buffered saline (Lonza), and the extracellular matrix was digested with 0.075% collagenase A (Roche Applied Science) at 37°C for 30 min. The samples were centrifuged at 1200 g for 10 min in complete culture medium. After discarding the supernatant containing oil, primary adipocytes, and collagenase solution, the stromal-vascular fraction pellet was cultured.
Flow cytometric analysis
The MSC phenotype was defined using the following monoclonal antibodies: anti-CD166-FITC (DakoCytomation), anti-CD45-CY5 and anti-CD146-Cy5 (BD Biosciences Pharmingen), anti-HLA-DR-CY5 (Immunotech), anti-CD34-PE, anti-CD73-PE (BD Biosciences Pharmingen), anti-CD31-PE, antiCD29-PE (Immunotech), anti-CD105-FITC (R&D Systems), anti-CD40-PE (Miltenyi Biotec), anti-CD80-FITC (Immunosource), anti-CD86-APC (Miltenyi), and HLA-ABC-Cy5 (eBioscience). Data were acquired and analyzed on an MacsQuant analyzer (Miltenyi Biotec).
TLR triggering and inflammation
To assess the influence of TLR3 and TLR4 ligation, the MSC were activated for overnight by polyriboinosinic polyribocytidylic acid [poly(I:C)] at 30 μg/mL (Sigma-Aldrich) and lipopolysaccharide (LPS) at 10 μg/mL (Sigma-Aldrich) respectively. 22 The inflammatory environment was mimicked by incubating the MSC for overnight in a cytokine cocktail combining interleukin (IL)-1β 25 ng/mL (Peprotech), interferon (IFN)-γ 103 U/mL, tumor necrosis factor (TNF)-α 50 ng/mL, and IFN-α 3×103 U/mL (all from Prospec, Inc.) as previously described.23–25
Induction of osteogenic differentiation in vitro of BM-, AT-, and WJ-MSC
The osteogenic potential of different MSC was tested by differentiating the cells in nonhematopoietic OsteoDiff Medium for human MSCs (Miltenyi). The MSC were seeded in a 24-well plate at a density of 2000 cells/well (for Ca and alkaline phosphatase [ALP] assays) or in 6 wells at a density 20,000/well (for quantitative polymerase chain reaction [qPCR]) in this medium up to 3 weeks. Osteogenic medium was changed weekly, and experiments were stopped after 7, 14, and 21 days to assess the osteoblastic features of MSC. Calcium mineralization was assessed by coloration with Alizarin Red (Sigma). The cells were washed in phosphate-buffered saline (PBS) and fixed in 70% ethanol at room temperature for 5 min followed by several washes in H2O. The cells were stained in 40 mM Alizarin Red (Sigma) pH 4.2 for 15 min at room temperature, rinsed in H2O, and then air dried. Red staining was examined by light microscopy. To evaluate the calcium deposits, the matrix was demineralized by the addition of 500 μL of 0.6 N HCl and incubated overnight at 37°C. Solutions were then collected and centrifuged for 5 min at 2000 rpm. The calcium concentrations in the supernatants were determined by colorimetry (QuantiChrom calcium assay Kit; BioAssay Systems) according to the manufacturer's instructions by measuring the absorbance at 610 nm using a plate reader (Organon Teknika Cappel Products).
Quantitative analysis of ALP activity
ALP activity was determined using the LabAssay™ ALP (Wako Chemicals Gmbh), according to the manufacturer's recommendations. This assay measures the quantity of p-nitrophenol released from a chromogenic substrate. The cell layers were lysed with 100 μL of ice-cold 0.1% Triton X-100 in PBS. Samples were then frozen and thawed twice, and the cell lysates were collected. Samples (20 μL) were combined with 100 μL ALP reagent, and the activity was measured after an incubation of 15 min at 37°C. The absorbance was measured immediately at 405 nm, and the amount of p-nitrophenol was determined by comparison with a standard curve. The results were expressed as U/μL, one unit of the enzyme activity defined as a release of 1 nmol p-Nitrophenol per minute at pH 9.8, 37°C.
Real-time quantitative PCR
Total mRNA was isolated from BM-, WJ-, and AT-MSC using the TriPure Isolation Reagent (Roche Applied Science). cDNA was obtained by the reverse transcription of 1 μg mRNA using qScript™ cDNA SuperMix (QUANTA bioscience) for 5 min at 25°C, 30 min at 42°C, and 5 min at 85°C. qPCR was performed on an ABI Prism® 7900HT Sequence Detection System (Applied Biosystem Rotterdam). We used 25 ng of cDNA in a qPCR with SYBR® Green PCR Master Mix (Applied Biosystems) and 0.32 μM of gene-specific forward and reverse primers for ALP as described elsewhere. 26 Relative quantification was calculated by normalizing the test crossing thresholds (Ct) with the glyceraldehyde 3-phosphate dehydrogenase amplified control Ct. As a positive control for osteogenic differentiation, a human epithelial-like osteosarcoma cell line (SAOS2) was used.
Statistical analysis
Results are expressed as the mean±the standard error of the mean (SEM). For the direct comparison between the different culture conditions (inflammation and TLR ligation), the paired t-test was used. All tests were two sided. An effect was considered significant at p<0.05.
All analysis were performed using GraphPad Prism version 5.00 for Windows (GraphPad Software; www.graphpad.com).
Results
Characterization of MSC from BM, AT, and WJ
The evaluation of cell surface marker expression demonstrated among MSC sources the reproducible expression of stem cell markers such as CD73, CD105, CD146, and CD166 and the absence of the hematopoietic marker CD45 and the endothelial marker CD31 (Fig. 1). We found that all MSC cell types express CD29, MHC class I but are negative for MHC class II (data not shown). In contrast to BM- and WJ-MSC, AT-MSC expressed the CD34 antigen, as previously reported. 27 However, BM- and WJ-MSC expressed higher levels of CD146 (98%±2% and 95%±13%, respectively) compared with AT-MSC (18%±6%). In comparison with BM- and AT-MSC, WJ-MSC expressed lower levels of HLA class I (data not shown).
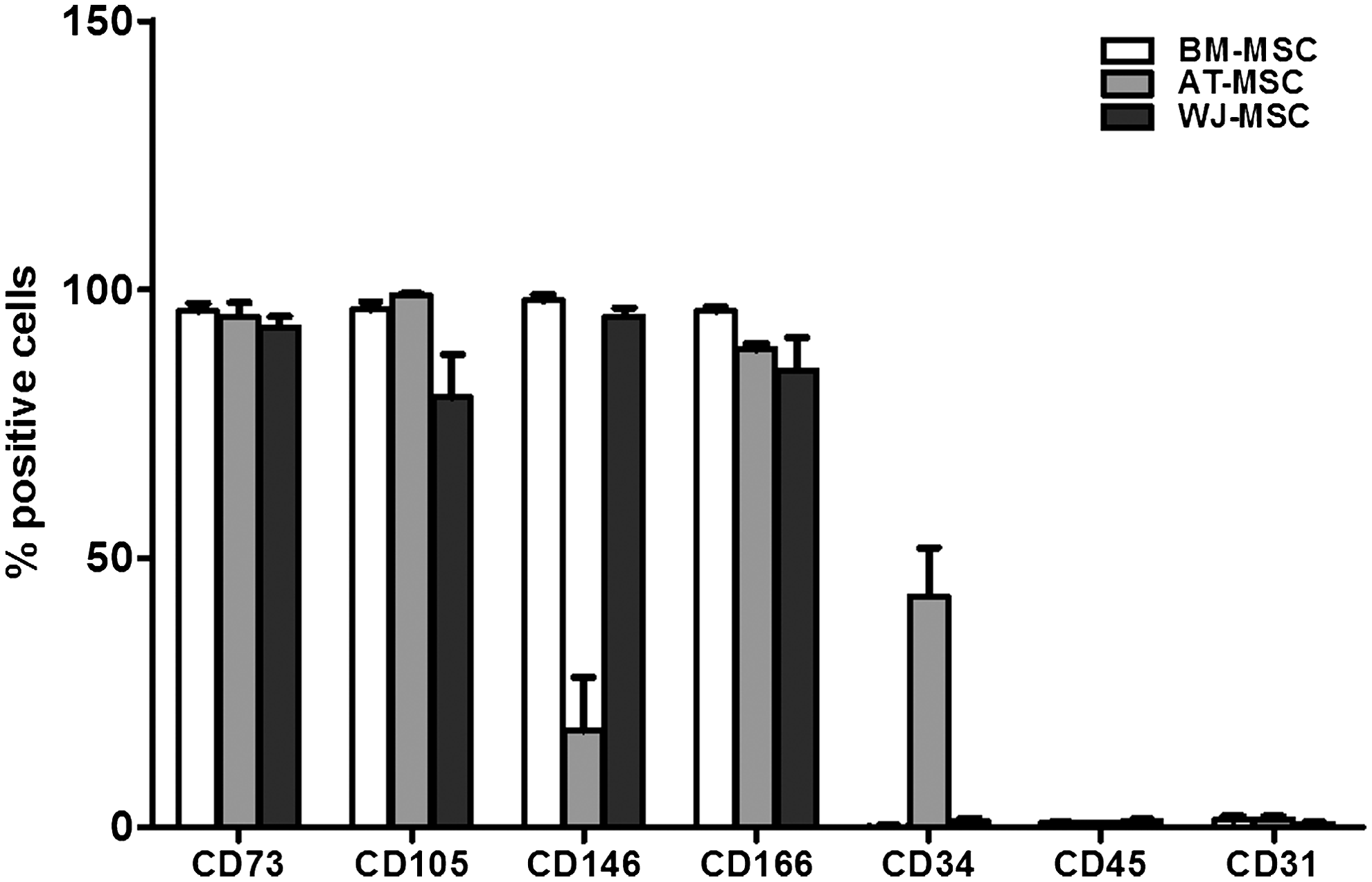
Expression of cell surface markers by BM-, AT-, and WJ-MSC. MSC isolated from three different sources. MSC were harvested at passage 2, labeled with monoclonal antibodies against CD73, CD105, CD146, CD166, CD34, CD45, and CD31, and analyzed by flow cytometry. Data represent mean±SEM of three different experiments. BM, bone marrow; AT, adipose tissue; WJ, Wharton's Jelly; MSC, mesenchymal stem cell; SEM, standard error of the mean.
The effect of TLR3 or TLR4 triggering on MSC calcification varies according to MSC source
Among the three different MSC types (BM, AT, and WJ) tested, BM-MSC presented constitutively the highest potential to differentiate into osteoblasts, and WJ-MSC expressed the lowest one. The increase in the Ca2+ deposit in BM-MSC, and, to a lesser extent, in AT-MSC, was already significant at day 14, while for WJ-MSC, a weak increase was only seen at day 21 of osteogenic differentiation (Fig. 2).
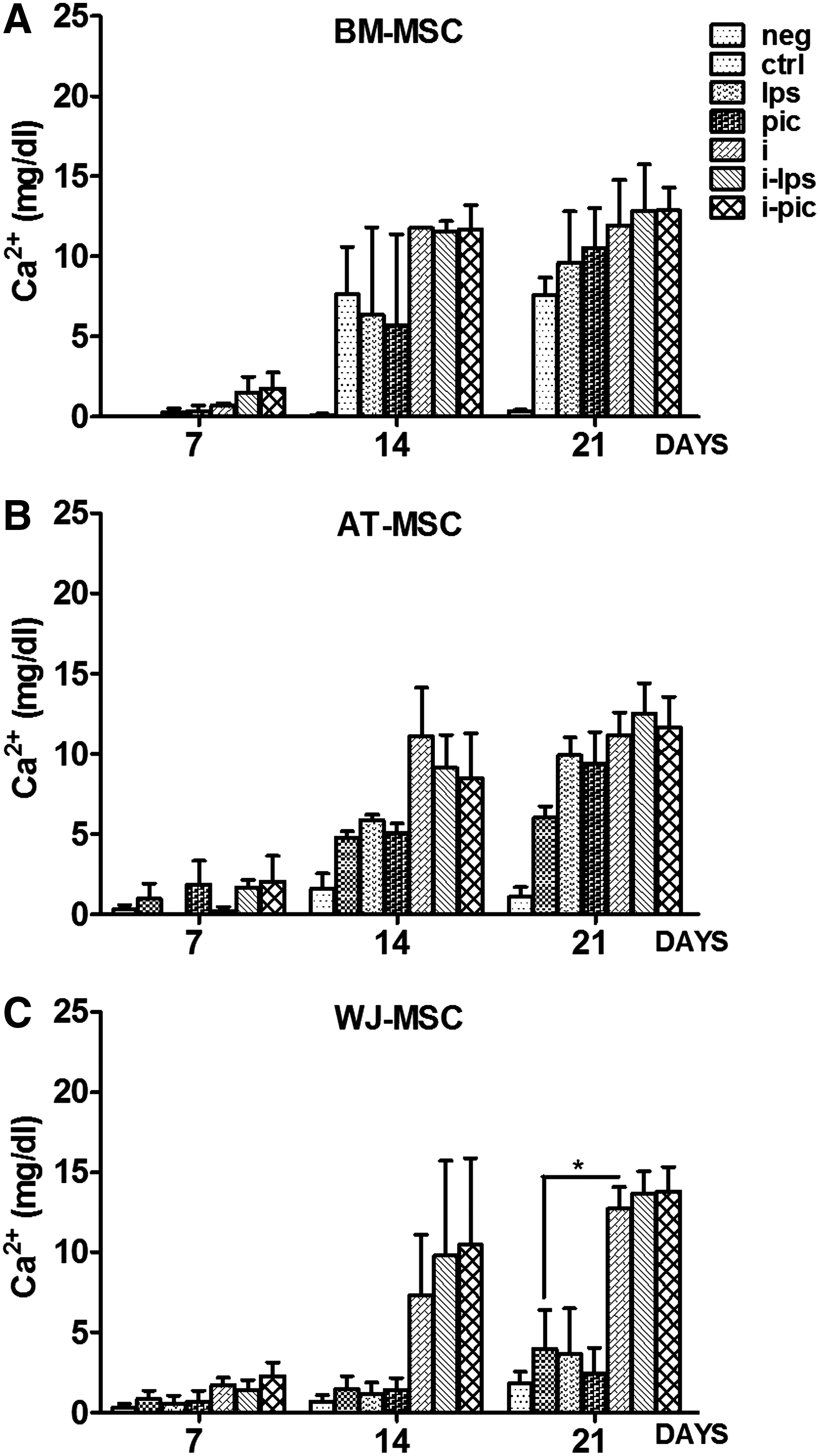
Influence of TLR ligation and/or inflammation on matrix calcification during the osteogenic differentiation of different MSC types. MSC isolated from
TLR3 or TLR4 triggering differentially affected MSC calcification depending on the source from which they have been isolated. TLR3 and TLR4 activation increased Ca2+ deposits in AT-MSC (Figs. 2B and 3B). We also noted a donor-dependent difference in the capacity of BM-MSC to differentiate into osteoblasts (Fig. 3A). BM-MSC isolated from young donors (<20 years) have higher osteogenic potential than those isolated from middle-aged (21–40 years) and older (>41 years) ones as observed by measuring the Ca2+ deposits (data not shown).

The effect of inflammation and/or TLR ligation on the osteogenic capacities of different MSC types. Mineralization abilities of MSC cells isolated from
Generally, TLR triggering did not affect the osteogenic potential of BM-MSC, but in some cases, an increased calcification was observed after treatment by Poly(I:C) or LPS. In accordance with our previous observations, 18 TLR4 activation did not affect the osteogenic potential of WJ due to its lack of expression. On the other hand, we also confirmed the nonactivity of TLR3 although expressed at the protein level.
Inflammation unveils the osteogenic potential of WJ-MSC
By measuring Ca2+ deposits after overnight MSC incubation in an inflammatory environment before adding the osteogenic medium, we observed an increase of the osteogenic potential of all three MSC types (Figs. 2 and 3). Calcification in WJ-MSC was already observed at day 14 (Figs. 2C and 3C). In the inflammatory setting, the values of Ca2+ measured at day 21 in WJ-MSC were comparable to those observed in BM-MSC or AT-MSC (Fig. 2A–C).
TLR3 or TLR4 triggering as well as inflammation affects ALP activity during the osteogenic differentiation process of the different MSC types
During osteogenic differentiation, ALP activity reached its peak value at day 14 in BM-MSC and at day 21 in AT-MSC. ALP activity was always very low in WJ-MSC even at day 21. TLR3 and TLR4 activation increased ALP activity in AT-MSC as well as accelerated it, thereby rendering this activity maximal already at day 14. TLR triggering did not have any influence on the ALP activity in BM- or WJ-MSC (Fig. 4). Similar to Ca2+, inflammation significantly boosted ALP activity in WJ-MSC with a peak value already reached at day 7 (Fig. 4C). The result of qPCR experiments confirmed these data. Inflammation up-regulated the expression of ALP mRNA, as witnessed by an appearance of ALP activity at day 7 and a peak value of mRNA copies at day 14 (Fig. 4D). Comparing the ALP activity in all three MSC types, it was clear that even when ALP activity reached its peak value in AT- and WJ-MSC (0.033±0.012 U/μL and 0.031±0.005 U/μL, respectively) after their preincubation in an inflammatory setting, it was almost ten times less than the one measured in BM-MSC (0.22±0.14 U/μL) in the same setting at day 14.
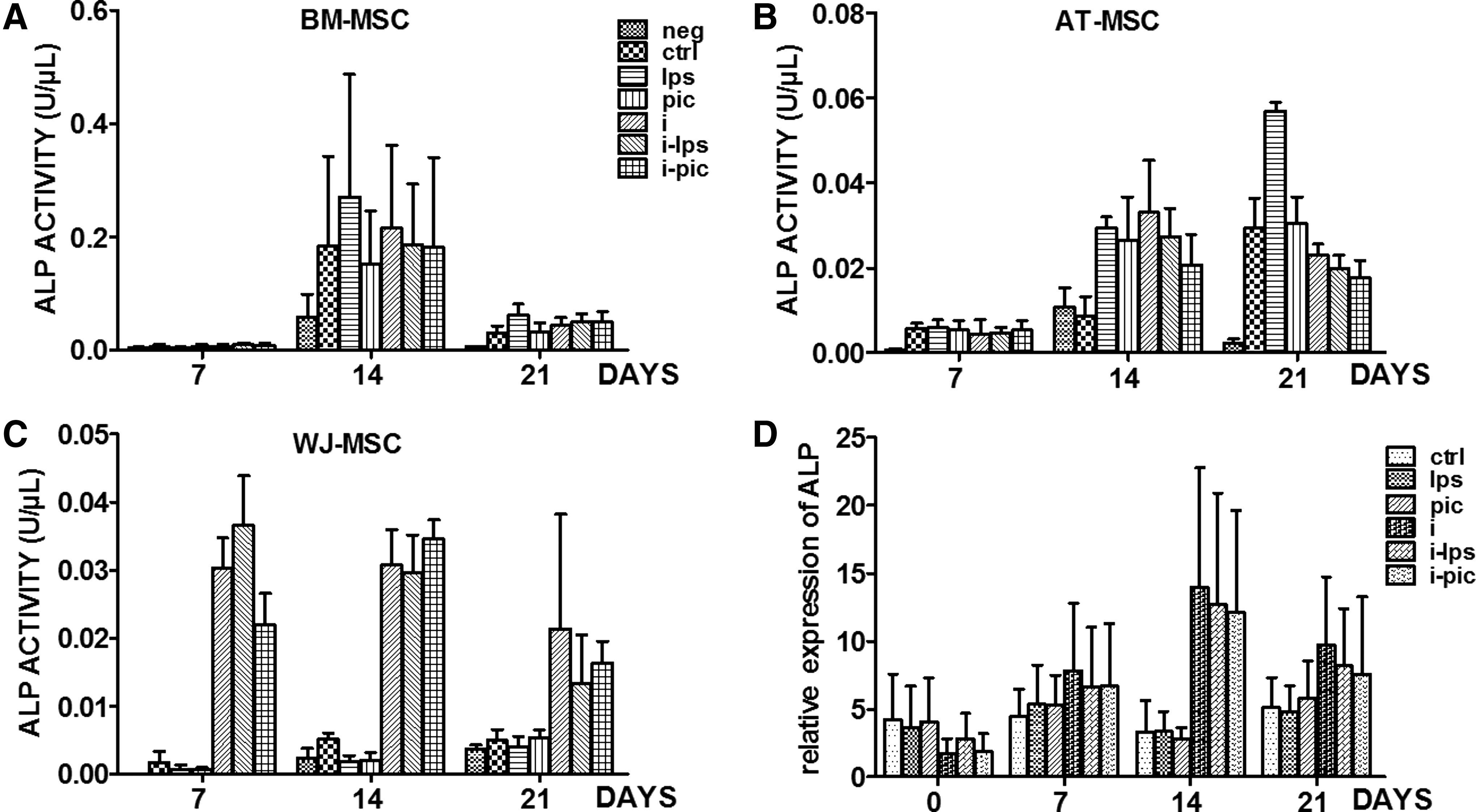
Influence of TLR ligation and/or inflammation on the ALP activity of MSC isolated from different sources. MSC isolated from
Discussion
In this study, we show, through a direct comparison of the different MSC types, that WJ-MSC have a weaker osteogenic capacity as compared with BM- and AT-MSC. Our work further reveals that the pretreatment of WJ-MSC with inflammatory cytokines significantly increases their osteogenic properties as measured by ALP activity and extracellular calcification. WJ-MSC osteogenesis is not influenced by TLR ligation, contrary to AT-MSC. BM-MSC osteogeneis was variably influenced depending on the donor.
Under adequate stimulation, MSC undergo osteogenic differentiation through a well-defined pathway involving osteoblastic gene expression and calcium crystals secretion.2,28 BM-MSC possess a very high potential to differentiate into osteoblasts that we confirmed by measuring ALP activity and Ca2+ deposits. The peak of ALP activity was reached after 14 days of MSC incubation in osteogenic medium. The comparison with WJ-MSC unveiled that ALP activity in these cells was very low even at day 21, suggesting that these cells present constitutively a very weak osteogenic capacity. This feature was confirmed by measuring extracellular calcification. Recently, Ciavarella et al. reported a predominant expression of the genes involved in osteogenesis by WJ-MSC as well as a trend to differentiate into osteoblasts similar to BM-MSC. 29 These data are apparently in contradiction with our observations. However, Ciavarella's work was performed on WJ-MSC without conducting a side-by-side functional comparison between WJ and BM-MSC in the same experiments, which could be of importance while taking into account the intra-experiment variability of such studies. Moreover, it has been already reported that WJ-MSC require more time to differentiate than other MSC types. 30 Two other reports also showed paradoxical data, suggesting that MSC purified from human umbilical cord disclose a better osteogenic potential than BM-MSC.8,31 Actually, these works studied MSC purified from the perivascular zone of the umbilical cord and not from the WJ matrix. By studying the global transcriptome as well as the functional differences between WJ and BM-MSC, Hseih et al. found that MSC from BM are a better source for osteogenesis, whereas WJ-MSC are more primitive and more similar to embryonic stem cells (ESCs), featuring a more multipotent status and a preferential growth advantage over BM-MSC. 32 It is worth noting that BM-MSC express more mesenchymal structural proteins such as vimentin or transgelin, which could partly explain that BM-MSC differentiate into bone nodules more rapidly than WJ-MSC. 33 Altogether, these data suggest that BM-MSC constitute the natural candidate for bone repair therapy, as they differentiate easily into osteoblasts.
Previous studies addressing the influence of inflammation on MSC have investigated the in vitro effects of different cytokines, including proinflammatory ones, on these cells. Readouts included bone formation. In mice, these studies showed that IL-1β and TNF-α significantly suppressed osteoblast development from MSC. 34 Another recent study showed that the osteogenic capacity of BM and WJ-MSC was not compromised on exposure to IFN-γ and TNF-α. 35 Rifas demonstrated that four cytokines released by activated T-cells, that is, TNF-α, TGF-β, IFN-γ, and IL-17, synergize to induce bone morphogenetic protein 2 (BMP-2) in MSC and induce ALP and extracellular matrix mineralization. 36 In our work, which aimed at studying the effect of inflammation on the osteogenic potential of MSC originating from different sources, we incubated the MSC in a medium supplemented with a pro-inflammatory cytokine cocktail consisting of IL-1β, TNF-α, IFN-α, and IFN-γ as previously described. 13 We observed not only that inflammation increases the osteogenic potential of BM and AT-MSC but also that it triggers WJ-MSC to differentiate into osteoblasts much earlier, as Ca2+ deposits were already measured at day 7 of osteogenic differentiation. After 14 days of culturing in an osteogenic medium, the levels of Ca2+ measured in WJ-MSC cultures supernatants were similar to those of BM- and AT-MSC. Even though the ALP activity reached its peak already at day 7 in WJ-MSC during the osteogenic incubation, this activity was almost 10 times weaker than in the case of BM-MSC, where the peak of ALP activity was reached at day 14. However, the ALP activity in WJ and AT-MSC was comparable with a peak activity of osteogenic differentiation at day 21 for AT-MSC. One of the earliest events in MSC commitment to osteoblasts is the expression of ALP activity. 37 The physiological substrate for this enzyme is inorganic pyrophosphate (PPi), which is hydrolyzed during the process of mineralization. 38 ALP is considered a major actor for the mineralization process. In our experiments, WJ-MSC disclosed a comparable calcification potential as compared with BM-MSC, although expressing almost ten times lower ALP activity. These data suggest that mechanisms other than ALP are involved in the osteogenic activity of WJ-MSC. Bigdeli et al. reported a superior osteogenic ability of ESCs compared with BM-MSC. Interestingly, despite the low ALP activity detected in ESC during osteogenic differentiation, these cells displayed a superior ability to form a mineralized matrix, indicating that high ALP activity is not crucial for the mineralization step. 39 All available data suggest that MSC from different species (human or mice) may behave differentially on the same stimulation. In addition, the influence of different proinflammatory cytokines on the osteogenic capacity of MSC depends on the experimental approach. The MSC preincubated with proinflammatory cytokines before the addition of the osteogenic medium, or the addition of an osteogenic medium along with different cytokines, will not have the same impact on the ossification of MSC. Moreover, it seems that the proinflammatory cytokine cocktail we used had a more potent influence on the osteogenic properties of all three MSC types than when individually used. 35 In conclusion, our data show that WJ-MSC, even though constitutively less osteogenic than BM-MSC, could be induced to differentiate into osteoblasts by inflammation in a manner similar to what is observed for BM-MSC or AT-MSC.
MSC express TLR. TLR signaling has been shown to be associated with the perpetuation of chronic inflammatory and autoimmune diseases.40,41 A better understanding of the effect of TLR activation on MSC function is pivotal, as these cells are now used in such clinical settings. In a previous study, our group showed that AT, BM, and WJ-MSC express different TLR patterns. WJ-MSC do not express TLR4 and express an inactive TLR3. 19 The data on the influence of TLR triggering on the suppressive properties of different MSC reported in the literature are somehow conflicting, probably reflecting a lack of standardization of the different experimental approaches.11–15,42 With regard to the influence of TLR ligation on the osteogenic potential of MSC, we observe that TLR3 and TLR4 triggering increases only the osteogenic potential of AT-MSC without affecting WJ-MSC. These data are in accordance with those of Lombardo et al. 15 BM-MSC had a variable capacity to differentiate into osteoblasts. Age-related changes in BM-MSC properties have been previously reported. They include a loss of differentiation and proliferation potential, as well as a loss in the capacity to generate bone in vivo. 43 TLR3 and TLR4 triggering on BM-MSC was associated with a trend to increase the osteogenesis of MSC with a lower differentiating capacity (isolated from older donors) without affecting the osteogenic potential of MSC with an already high differentiation potential (isolated from younger donors).
MSC express various TLR, and their activation induces various types of responses that affect the therapeutic efficacy of stem cells. TLR agonist engagement affects several functions of MSC: migration, invasion, and secretion of immune-modulating factors. Inflammatory conditions are associated with a significant up-regulation of TLR and can also modulate the immunosuppressive and differentiation capacities of MSC. Some of the molecules that modulate MSC immunological properties also affect the ability to differentiate along the mesenchymal lineage.
In conclusion, our work shows that WJ-MSC disclose original characteristics as compared with other MSC types; although being constitutively weakly osteogenic, they can become effectively osteogenic in an inflammatory environment. Furthermore, their properties are not affected by TLR3 and TLR4 ligation. Taking into account the easy accessibility as well as their huge proliferative potential, these data open an avenue for using these cells in regenerative medicine, particularly in clinical settings were chronic inflammation or infection is present.
Footnotes
Acknowledgments
G. Raicevic and M. Najar are Télévie research fellows of the Fonds National de la Recherche Scientifique (FRS-FNRS 7.4564.10-7.4638.10). They were supported by BRUSTEM, an impulse program of the Institute for the Encouragement of Scientific Research and Innovation of Brussels (ISRIB) and the “Fondation Lambeau-Marteau.” The authors wish to express their thanks to the team of the obstetric department of Edith Cavell Clinic for providing human umbilical cord samples.
Disclosure Statement
No competing financial interests exist.