
Abstract
Mesenchymal stem cells (MSCs) represent a promising therapeutic approach in nerve tissue engineering. To date, the local implantation of MSC in injured nerves has been the only route of administration used. In case of multiple sites of injury, the systemic administration of cells capable of reaching damaged nerves would be advisable. In this regard, we found that an intravenous administration of adipose-derived MSC (ASC) 1 week after sciatic nerve crush injury, a murine model of acute axonal damage, significantly accelerated the functional recovery. Sciatic nerves from ASC-treated mice showed the presence of a restricted number of undifferentiated ASC together with a significant improvement in fiber sprouting and the reduction of inflammatory infiltrates for up to 3 weeks. Besides the immune modulatory effect, our results show that ASC may contribute to peripheral nerve regeneration because of their ability to produce in culture neuroprotective factors such as insulin-like growth factor I, brain-derived neurotrophic factor, or basic fibroblast growth factor. In addition to this production in vitro, we interestingly found that the concentration of glial-derived neurotrophic factor (GDNF) was significantly increased in the sciatic nerves in mice treated with ASC. Since no detectable levels of GDNF were observed in ASC cultures, we hypothesize that ASC induced the local production of GDNF by Schwann cells. In conclusion, we show that systemically injected ASC have a clear therapeutic potential in an acute model of axonal damage. Among the possible mechanisms promoting nerve regeneration, our results rule out a process of trans-differentiation and rather suggest the relevance of a bystander effect, including the production of in situ molecules, which, directly or indirectly through a cross-talk with local glial cells, may modulate the local environment with the down-regulation of inflammation and the promotion of axonal regeneration.
Introduction
Previous studies aiming at restoring sciatic nerve function after injury have employed artificial nerve conduits and scaffolds or delivery of cells directly at the lesion site.9,10 The intrinsic limitation of this route of administration is the additional trauma necessary in reaching the injection site as well as the difficulty in treating multiple or diffuse sites of injury. In such circumstances, a systemic administration of cells capable of reaching the damaged PNS would be advisable.
In this regard, MSC display a full repertoire of adhesion molecules that enable their migration in damaged tissues. In the last years, much attention has been paid to adipose-derived MSC (ASC), because adipose tissue is an abundant, easily accessible, and appealing source of donor tissue for autologous cell transplantation.11,12 The comparative analysis of ASC and BM-MSC clearly showed that both populations shared a common morphology, immune phenotype, success rate of isolation, colony frequency, and differentiation potential.13,14 With regard to the cell surface marker expression, ASC showed a peculiar adhesion molecules profile with the expression of CD49d (α4 integrin),15,16 which forms a heterodimer with CD29 to create very late activation antigen 4 (VLA-4). This molecule plays a key role in leukocyte extravasation during inflammation, interacting with endothelial vascular cell adhesion molecule-1 (VCAM-1), which is up-regulated into the inflamed PNS. 3 Besides the ability to migrate into damaged tissues, 17 ASC exerted a relevant immune modulatory activity both in vivo and in vitro in a broad range of immune cells.18–20 Moreover, several research groups have found evidence for neural and glial trans differentiation of ASC in vitro and in vivo.12,21,22 Finally, ASC secrete a variety of neurotrophins and cytokines,15,23 which might positively impact neural cell survival and neuroregeneration. The present study was designed to evaluate the potential benefits of an intravenous administration of ASC in a murine model of acute axonal damage, the sciatic nerve crush injury. The effects of ASC treatment were evaluated in terms of functional recovery and nerve regeneration, assessing the ability of injected cells to integrate into the injured peripheral nerve, to modulate the immune response, and to influence the local micro environment through the release of neurotrophic molecules.
Materials and Methods
Human ASC cultures
Human (h)ASC were isolated from the lipoaspirates of subcutaneous abdominal fat obtained from five female donors during abdominoplastic procedures. All donors gave their informed consent. The isolation of stromal-vascular fraction was carried out on 5 mL of lipoaspirates, as previously described 21 ; briefly, adipose tissue was washed with sterile Hank's balanced salt solution (HBSS), and the extracellular matrix was digested at 37°C in HBSS with type I collagenase and bovine serum albumin (Sigma-Aldrich). The stromal-vascular fraction was then collected by centrifugation at 1200 g, and the cells were cultured in 25 cm2 flasks (EuroClone) in Dulbecco's modified Eagle's medium, with 15% heat-inactivated adult bovine serum, penicillin, and streptomycin (EuroClone) at 37°C in a 5% CO2 atmosphere. After 72 h, nonadherent cells were removed, and thereafter, the medium was changed twice a week. When 70%–80% adherent cells were confluent, they were trypsinized, harvested, and expanded in larger flasks. A homogenous cell population from all donors (in terms of morphological features and proliferation rate) was usually obtained after 3 to 5 weeks of culture. All the experiments were performed at passages 8 to 12. 21
hASC were characterized by the expression of CD105 (endoglin), CD73, CD29, CD44, CD90 class I HLA, and lack of hematopoietic (CD45, CD14, CD34) and endothelial (CD31) markers. In addition, the absence of hematopoietic markers (CD45, CD11c, and CD34) and endothelial markers (CD31) was assessed as previously described. 13 All the mAb just mentioned were purchased from Pharmingen/Becton Dickinson. For immunophenotypical analysis, 106 cells/mL ASC suspension was incubated in 15% of fetal bovine serum, followed by the specific mAb for 30 min. At least 10,000 events were analyzed by flow cytometry (FACScalibur; Becton Dickinson) using Cell Quest software.
Animals and surgical procedures
For sciatic nerve crush, 6–8 week-old female C57Bl/6 mice obtained from Charles River Laboratories underwent unilateral sciatic nerve crush. The animals were anesthetized with intraperitoneal injections of ketamine (100 mg/kg; Alcyon) and xylazine (10 mg/kg; Sigma-Aldrich). The right hindquarter was shaved; the skin was incised 1 mm posterior and parallel to the femur; and the biceps femoris was bluntly split to expose the sciatic nerve under 16×magnification. The crush injury was performed 5 mm proximal to its trifurcation using number 5 Jeweler's forceps for 30 s. 24 After the injury, the muscles were approximated, and the wound was closed by suturing the skin with 4/0 interrupted nylon sutures. Two×106 hASC were administered through the tail vein 7 days after the surgery in 1 mL of phosphate-buffered saline (PBS) without Ca2+ and Mg2+, while the control mice received i.v. 1 mL of PBS. The animals (nine animals per group per time point) were sacrificed by CO2 chamber euthanasia at different time points (14, 21, and 35 days after the crush), and the sciatic nerves were dissected from both sides. A 2-cm portion of each sciatic nerve (1-cm above and 1-cm below the crush site) was collected, snap frozen in liquid nitrogen, and stored at −80°C for histological and immunohistochemical analyses. The experiments received the authorization from the Italian Ministry of Health, and were conducted following the principles of the National Institutes of Health Guide for the Use and Care of Laboratory Animals, and the European Community Council (86/609/EEC) directive.
Functional assessment
Walking track analysis was performed the day before the surgery and then weekly on the 7th, 14th, 21st, 28th, and 35th days postlesion. Briefly, the plantar surface of the hind paws was dipped with black ink, and the mice were allowed to ambulate down a standard walking trace in which a strip of graph paper (30×7 cm) was displayed. Measurements were made according to the Sciatic Functional Index (SFI), 25 where 0 corresponds to normal function, and −100 corresponds to a total functional impairment of one side. Print length (PL) and toe spread (TS) were blindly measured from two different examiners (S.M. and E.T.). At least four footprints per animal were taken from each side, which were used for calculation of the SFI mathematical formula 25 : SFI=118.9×(ETS−NTS)/NTS−51.2×(EPL−NPL)/NPL−7.5, where E means experimental, and N is normal. According to the SFI value assessed at day 7 postcrush, the mice were divided into two functionally homogeneous groups. Results are expressed as mean±standard deviation (SD).
Histology and immunohistochemistry
Histological assessment of sciatic nerve regeneration and inflammatory infiltrates was blindly performed by two different examiners (S.M. and E.T.) on frozen serial sections (13 μm thick). Both cross and longitudinal sections were obtained from healthy and crushed sciatic nerve (at least six animals for each time point: 14, 21, and 35 days postcrush, for both treatment conditions) and processed for immunohistochemistry with antibodies for CD3 (T lymphocytes), CD11b (macrophages/monocytes, both 1:1500; R&D Systems), or growth associated protein-43 (GAP-43; 1:1000; Chemicon). The slices were incubated overnight at 4°C and, after washing with PBS, anti-rat-biotynilated IgG (for CD3 and CD11b) or anti-rabbit-biotinylated IgG (for GAP-43) secondary antibody (1:100; Vector Laboratories) was added for 1 h. After washing, the reaction was visualized with the ABC kit and Novared Substrate Kit (Vector) according to the manufacturer's instructions. Images of immune peroxidase were obtained with a Zeiss Axiophot microscope and Axiocam camera and analyzed with Axiovision software.
For the determination of CD3+ and CD11b+ infiltrates within nerves, their density was calculated by dividing the software counts of positive cells by the damaged area considered (cells/mm2), as previously described. 15 GAP-43 immunostaining was used to characterize the regenerative process, as it is highly induced in regenerating fibers in the injured sciatic nerve as early as 2 days after the injury. 1 The mean number of fibers per section was determined in the sciatic nerves from different experimental groups, quantifying the number of GAP-43+ fibers (fibers/section) 1.5 mm beyond the injury site, as previously described.1,26 The extent of sciatic nerve regeneration was also analyzed by measuring in each sample the sum of the length of GAP-43+ fibers from the centre of the crush site, as previously described.26,27 In order to evaluate the presence, distribution, and differentiation of hASC in the sciatic nerve, we injected hASC in nine animals, which were sacrificed at days 14, 21, and 35 postcrush; the sections were stained with anti-human nuclei (Chemicon; 1:100), 4',6-diamino-2-phenylindole (DAPI), and then with S-100 (Dako), peripheral myelin protein-22 (PMP-22; Chemicon), neurofilament, or glial fibrillary acidic protein (GFAP; Dako); the signal was then detected by appropriate secondary Alexa488-antibody (Molecular Probes, Life Technologies) or biotinylated antibodies and Streptavidin Texas Red (Vector). The slides were viewed under a Leica TCS SP5 tandem confocal scanner with the acquisition of images at different wavelengths (DAPI 455 nm, Alexa488 500 nm, Texas Red 615 nm). All the counts were performed in three sets (four sections 100 μm apart) for each animal, and all data are expressed as mean percentage±SD.
Enzyme-linked immunosorbent assay for neural growth factors
The quantification of neural growth factors was performed both on the supernatants of ASC and in the homogenates from the sciatic nerves of mice injected with ASC or PBS and healthy animals. To determine the production by ASC of glial-derived neurotrophic factor (GDNF), basic fibroblast growth factor (bFGF), brain-derived neurotrophic factor (BDNF), vascular endothelial growth factor (VEGF), and insulin-like growth factor I (IGF-I), supernatants were obtained from 1×105 ASC in basal condition for 24 h and analyzed by Quantikine® ELISA Immunoassay (R&D Systems) following the manufacturer's instructions. Briefly, the cells were grown in 24-well plates, and the supernatants were harvested and centrifuged for 10 min to remove cell debris. The samples were added in 96-well precoated plates and incubated for 2 h at RT. After washing, a specific polyclonal antibody followed by a substrate solution was added, and the color development was measured at 450 nm (BioRad Microplate Reader). The concentration of growth factors was calculated using the standard curve.
To determine the protein level of GDNF, IGF-I, BDNF, and bFGF in the sciatic nerves, proteins were extracted from the frozen tissue samples using the PARIS kit (Applied Biosystem). Briefly, the samples were cut into small pieces and then homogenized in cell disruption buffer according to the manufacturer's instructions. The concentration of proteins was assessed according to Bradford assay. 28 The same protein amount was subjected to the Quantikine ELISA Immunoassay (R&D Systems), in accordance with the manufacturer's instructions. The levels of expression of neural growth factors in the sciatic nerve from three experiments were expressed as ng/mg total protein (mean±SD).
Statistical analysis
Between-group (ASC vs. PBS) and within-group (ASC) comparisons were performed over time using a generalized linear model (GLM) with one of the nine variables of interest (CD3 cells, CD11b cells, SFI, GDNF, IGF-I, BDNF, hASC, fibers per section, and length of GAP-43+ fibers) as outcome, a categorical time predictor (0, 5, 14, 21, 28, and 35 days for SFI and 14, 21, and 35 days for all other outcomes), and an additional group X treatment interaction for between-group comparisons. 29 The GLM family was set to Gaussian, and a log link was used to reduce skewness and to improve the model fit for all outcomes with the exception of SFI, for which an identity link was used. Margins were calculated from the GLM, and prespecified contrasts were used to perform between-group comparisons at each time point using Bonferroni's correction. When repeated measures were involved (SFI), cluster confidence intervals were used. Robust confidence intervals were used in all other cases. Statistical analysis was performed using Stata version 12.0. All reported p-values were corrected for multiple comparisons, and a corrected p-value<0.05 was used to define statistical significance.
Results
ASC increased motor functional recovery
The SFI scores at different time points for each group are shown in Figure 1A. The mean SFI score 7 days after crush before the systemic injection was −88.53±9.16 for the experimental group and 88.93±8.45 for the control group, confirming the reproducibility of both functional assessment and surgical procedure. The injection of ASC exerted a significant improvement in SFI as compared with the control-crushed animals starting from 14 days after the injury (i.e., 7 days after ASC injection) (p<0.001) and was maintained at day 21 (p<0.001). The sciatic nerve deficit produced an elongated PL resulting from foot drop and lack of plantar flexion, and also the TS strongly decreased. Treatment with ASC was able to ameliorate both plantar flexion and TS as early as after 7 days postinjection (Fig. 1B). At day 35 postcrush, both control and ASC-treated animals reached almost a completely normal motor function.
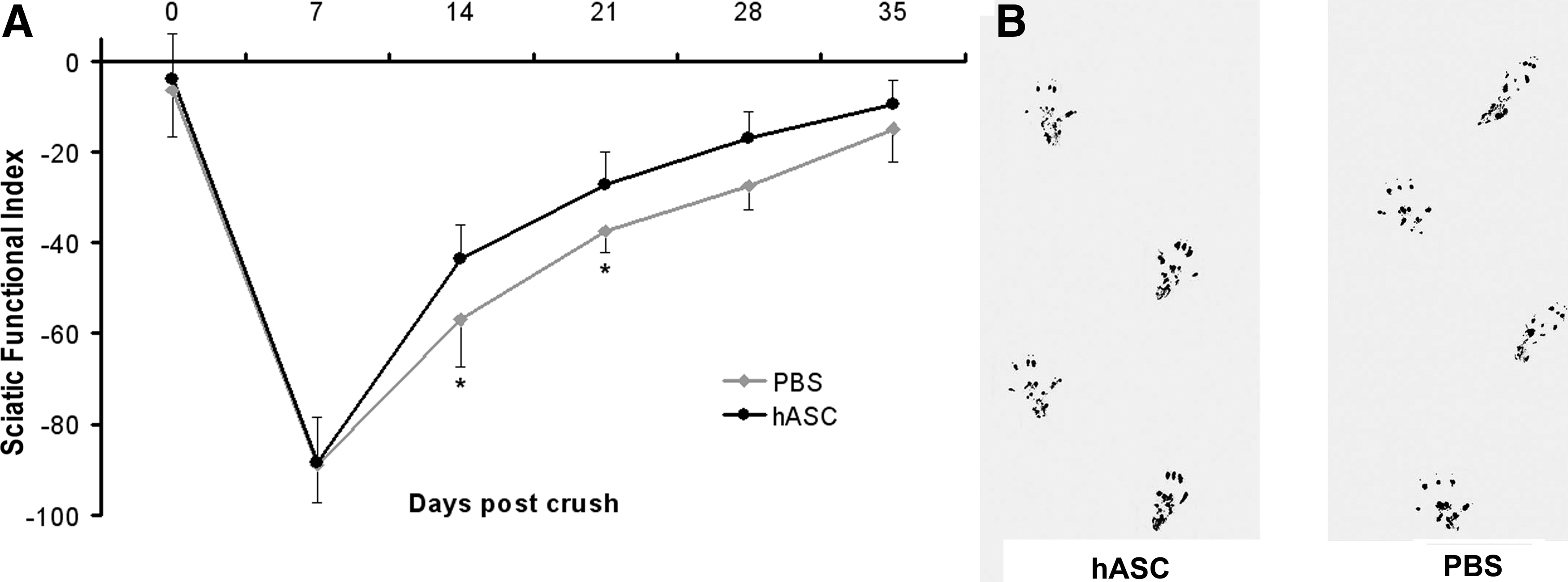
Motor functional recovery is accelerated after i.v. ASC treatment.
Treatment with ASC improves fiber sprouting and reduces inflammatory infiltrates
GAP-43 is the major protein kinase C substrate of growth cones and developing nerve terminals and is considered a good marker for evaluating the extent of axonal regeneration. 1 Immunostaining for the GAP-43 protein was performed on longitudinal sections of crushed sciatic nerves from animals treated or not with ASC at 14, 21, and 35 days after the crush (Fig. 2A, B). Fiber sprouting was quantified by counting the number of GAP-43 fibers per section (Fig. 2C, D). The number of regenerating fibers was significantly increased in the animals treated with ASC as compared with the PBS-treated controls at days 14 and 21 after the crush (Fig. 2E and Supplementary Fig. S1; Supplementary Data are available online at www.liebertonline.com/tea); in particular, the number of regenerating fibers after ASC treatment was almost 40% more than the controls. We also assessed the length of GAP-43+ fibers distal from the crush site, which was also significantly increased in the ASC-treated animals as compared with the controls (Fig. 2F and Supplementary Fig. S1). At day 35 after the crush, when the regenerative process had almost restored physiological conditions, no differences in GAP-43 expression (both number and length of positive fibers) were observed between the PBS- and ASC-treated mice.

Treatment with ASC increases neural regeneration. Immunohistochemistry for growth associated protein-43 (GAP-43) on longitudinal sciatic nerve sections showed an increase of regenerating fibers in the distal nerve stump in ASC-treated mice
The histological analysis of sciatic nerve sections in controls and ASC-treated mice showed the presence of extensive inflammatory infiltrates at the crush site 7 days after the crush, composed of monocyte/macrophages and T lymphocytes (data not shown). We found that mice treated with ASC showed a drastic reduction at the areas of inflammation, with a significant decrease of monocyte/macrophages compared with the PBS-treated controls at day 14 postcrush (Fig. 3A, B), and this effect was maintained until the end point (Fig. 3C and Supplementary Fig. S1). The number of CD3-positive lymphocytes was also significantly reduced after treatment with ASC, being statistically significant from 21 to 35 days postcrush (Fig. 3D and Supplementary Fig. S1).
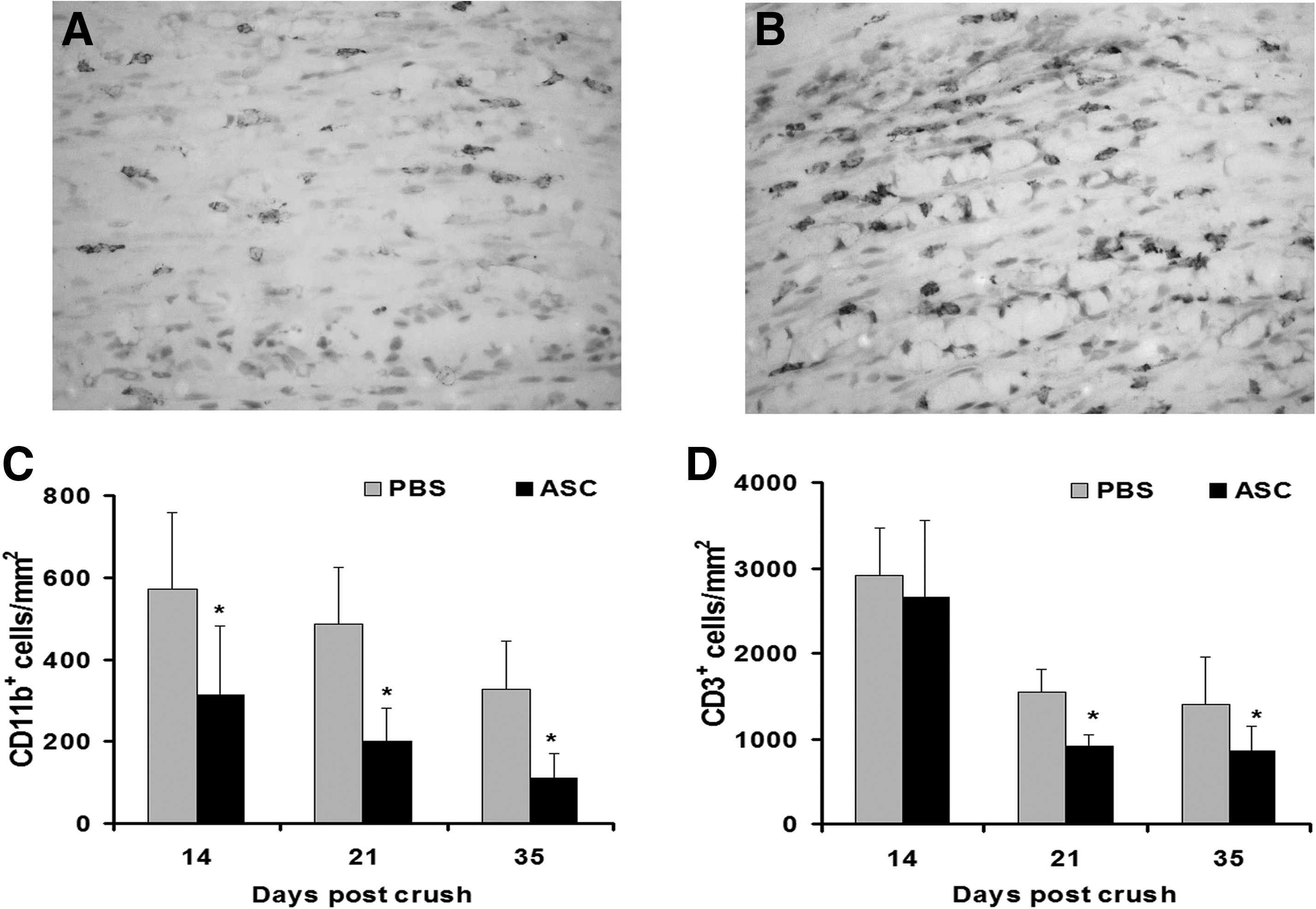
Treatment with ASC decreases neural inflammation. The analysis of inflammatory infiltrates in the nerves from ASC-treated mice showed a significant decrease in CD11b+ monocyte/macrophages
Distribution of ASC
The analysis of the distribution after a systemic injection of hASC by immunofluorescence with an anti-human-nuclei antibody confirmed the ability of ASC to migrate into lymphoid organs (data not shown), as seen in other animal models. 13 In addition to the lymphoid organs, a restricted number of hASC migrated into the inflamed PNS, being detectable after 7 days postinjection and persisting up to 35 days (Fig. 2G, H). The double staining with glial phenotypic markers showed sporadic hASC in the crushed nerves displaying Schwann cell markers (data not shown). No evidence of binucleated cells was observed in all the samples examined.
Production by ASC in vitro and modulation in vivo of neurotrophins
The presence of ASC with undifferentiated phenotypes in crushed sciatic nerves may suggest that these cells could promote the survival of Schwann cells and nerve regeneration and, at the same time, could modulate the immune response by by-stander effects. Among the possible mechanisms through which ASC may modulate neural cells, we assessed by enzyme-linked immunosorbent assay (ELISA) the production by ASC of a number of neural growth factors, known to influence their self-renewal as well as the process of nerve regeneration. In a previous study, 15 we found that ASC were able to secrete considerable amounts of BDNF, bFGF, and PDGF-AB in vitro. In the present study, the supernatants of ASC also showed the production of IGF-1 (991.3±63.3 pg/mL) and VEGF (1071.3±86.1 pg/mL), but not of GDNF or ciliary neurotrophic factor (CNTF). We then directly quantified these neurotrophins by ELISA in sciatic nerve homogenates to assess whether the ASC penetrated into the PNS were able to modulate their production in situ. The analysis was performed on the sciatic nerves of healthy and crushed animals treated or not with ASC and sacrificed at days 14, 21, and 35 after the crush. We found that ASC treatment significantly prevented the crush-induced decrease of GDNF at days 14 and 21 (Fig. 4A and Supplementary Fig. S1) and of IGF-I at day 14 (Fig. 4B) as compared with controls, whereas no significant difference was observed in the tissue levels of BDNF in both treatment groups (Fig. 4C). The quantification of tissue neurotrophins performed at the end stage, when the regenerative process was almost complete, did not show relevant differences between the ASC- and PBS-treated mice. We also observed that treatment with ASC slightly reduced the tissue levels of bFGF compared with the controls, while the tissue production of VEGF was not detectable in both the groups (data not shown).
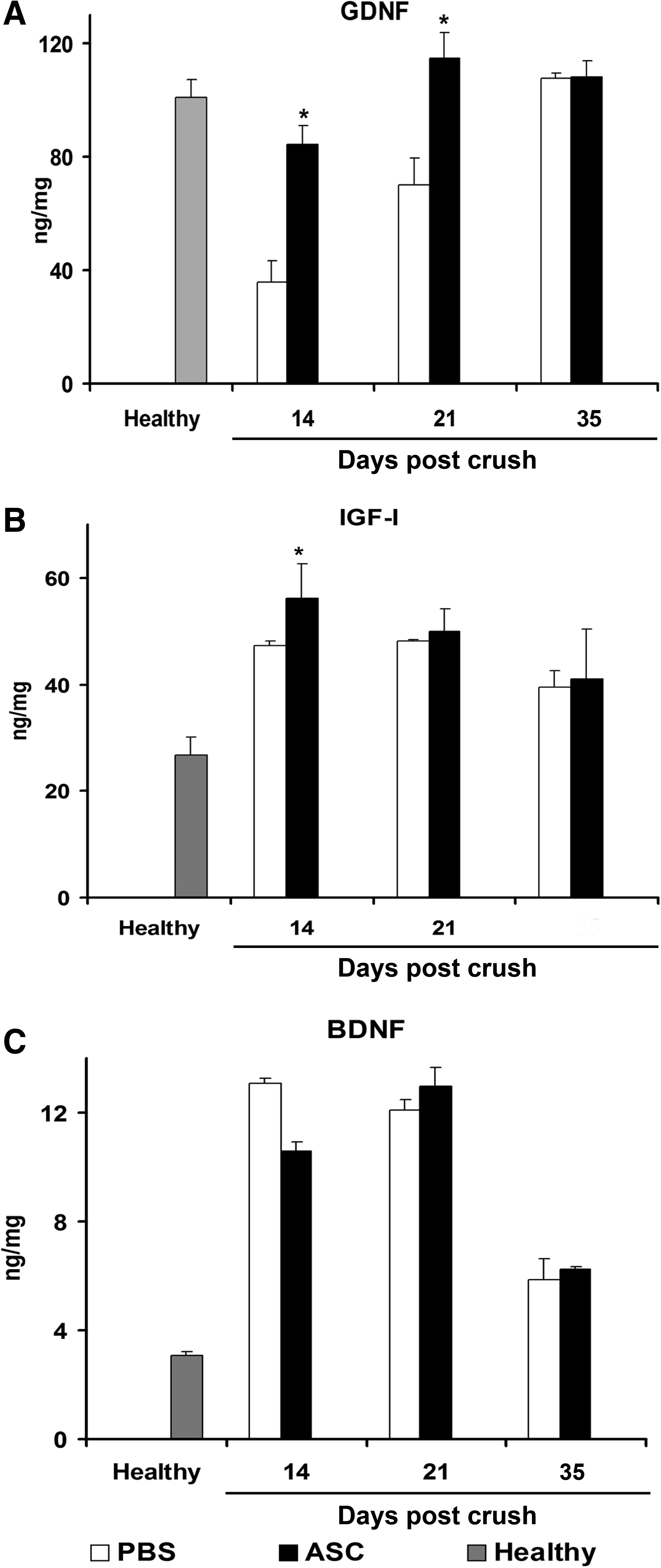
Modulation of nerve levels of neurotrophins after a systemic injection of ASC. A significant increase in the tissue concentration of GDNF has been detected in ASC-treated sciatic nerves at days 14 and 21 postcrush as compared with the PBS-treated samples; *p<0.001
Discussion
Stem cells represent a promising therapeutic approach in the treatment of neurological disorders affecting central and PNS. Stem cell-based therapy, with the local implantation of MSC in peripheral nerve injury, has been shown to promote nerve regeneration, with axonal regrowth and myelin formation.5,7,30–32 The intrinsic limitation of this approach is related to the invasiveness in reaching the injection site as well as to the difficulty in treating multiple injury sites. A major challenge in such cases is represented by the possibility to obtain nerve regeneration through a systemic administration of cells capable of reaching the damaged PNS. Several lines of evidence indicate that adipose tissue may represent an ideal source for stem cells: (1) it can be obtained in relatively large quantities with minimal risk or discomfort; (2) the frequency of ASC in digested adipose tissue is about 500-fold higher that in freshly isolated BM cells11,22; (3) ASC display a full repertoire of adhesion molecules that enable their extravasation into the damaged PNS. Moreover, hASC escape immune system surveillance, because they possess cell surface antigens that are poorly recognized by T cells; therefore, hASC can be transplanted as an autograft, allograft, and even xenograft.33,34 In the current study, we found that the systemic injection of undifferentiated ASC 1 week after the sciatic nerve crush injury induced a significant acceleration of the subsequent motor functional recovery. The SFI analysis showed that ASC treatment significantly improved motor performance as early as 1 week after the injection compared with the PBS-treated animals, and this improvement was maintained until the end of the experiments. Histopathological analysis confirmed this beneficial effect in terms of increased fiber sprouting and the reduction of inflammatory infiltrates. The extent of the regenerative process was assessed by GAP-43, which is usually expressed at low levels and after the injury is induced in the nerve cell body, rapidly transported along the axon, and inserted into lipid rafts in the growth cones. 1 Very interestingly, we found a significant improvement of GAP-43+ regenerating fibers in the mice treated with ASC compared with the controls. In addition to the regenerative effect, we observed a parallel reduction of inflammatory infiltrates (both lymphocytes and macrophages) after ASC treatment compared with the PBS-treated mice, and that persisted until the endpoint. This result further confirms the immune-modulatory effect of ASC18–20 also in a model of peripheral acute inflammation and degeneration. In addition to their systemic immune modulatory activity, our results suggest that the regenerative effect of ASC after sciatic nerve injury is mediated by by-stander effects exerted within the damaged PNS. After system injection, a restricted number of hASC, in fact, were able to cross the blood-nerve barrier and penetrate into the damaged PNS, probably due to the expression of activated α4β1 integrin, 15 a key adhesion molecule involved in leukocyte and stem cell migration into the inflamed central and PNS.3,35 The assessment of homing of hASC showed that these cells persisted in crushed nerves for up to 40 days, without the relevant expression of markers of Schwann cell lineage, thus ruling out a direct participation of ASC (i.e., glial trans-differentiation) in the regenerative process. It is rather conceivable that ASC may mediate their therapeutic effects in the target tissues by complex paracrine and autocrine mechanisms.
Peripheral nerve regeneration involves Schwann cells, neurotrophic factors, and inflammatory cells working synergistically to promote and maintain axonal sprouting and myelination of fibers. Several studies suggest that stem cells may contribute to the repair of injured tissue by releasing a large number of trophic factors in situ, which can profoundly influence the local micro environment.23,36,37 In our study, we demonstrated that ASC were able to secrete in vitro a vast panel of neural growth factors, such as BDNF, bFGF, PDGF-AB, 15 IGF-I, and VEGF, but not GDNF or CNTF. Among the neurotrophic molecules, GDNF, a member of the transforming growth factor β superfamily, is perhaps the most potent trophic factor influencing the development, survival, and maintenance of neural cells in the central and PNSs. 38 In peripheral nerves, Schwann cells are the main source of GDNF, which is needed for neuronal survival. 39 Chen and colleagues reported the effectiveness of GDNF-modified amniotic stem cells embedded in Matrigel and locally delivered at the moment of crush into the injured rat nerve, 40 while Shi and co-workers demonstrated that an intramuscular injection of adenovirus encoding the GDNF 7 days after the injury significantly increased the number of myelinated axons as well as axon fibers after sciatic nerve constriction. 38 Interestingly, we found that the systemic injection of undifferentiated ASC abolished the physiological decrease of GDNF in crushed nerves, maintaining a constant concentration of GDNF until the regenerative process was complete. This result is the best example of the cross-talk between ASC and Schwann cells, as ASC in culture produce no detectable levels of GDNF. In addition to the robust effect on GDNF, ASC treatment increased the tissue levels of IGF-I and BDNF, both involved in axonal growth and sprouting, and both required for peripheral nerve regeneration.36,41 It is conceivable that these trophic molecules work in a complementary way during peripheral nerve regeneration, in order to promote the survival and outgrowth of axons and Schwann cells.42,43 The cross-talk with local glial cells may explain how a limited number of ASC present within crushed nerves may amplify their biological effects, although such a hypothesis needs to be confirmed by additional experiments. From our experiments, we obviously cannot exclude that the beneficial effect observed after a systemic injection of ASC may be due to the additional mechanisms exerted at the CNS level (e.g., neurons in spinal cord and dorsal root ganglia, which undergo a robust response after any nerve injury) or systemically by peripheral immune suppression.
Our data show that ASC have relevant therapeutic potential in an animal model of acute axonal injury and might represent a valuable tool for stem cell-based therapy in inflammatory and degenerative diseases of the PNS. The persistence of ASC in the sciatic nerve, together with the profound changes in the local micro environment, not only via their direct release of growth factors, but also through the indirect modulation of neural cell behavior, may have important implications for the future therapeutic use of ASC. These results open the possibility of a novel form of treatment for patients suffering from different peripheral nerve diseases with an inflammatory and degenerative pathogenesis, from traumatic nerve injury to inflammatory polyradiculoneuritis.
Footnotes
Acknowledgments
This work was supported in part by the following grants: CariVerona 2008, PRIN 2009, European Research Council Grant Neurotrafficking, and the FP7 program.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.