
Abstract
Fibroblasts as many other cells are known to form, contract, and remodel the extracellular matrix (ECM). The presented study aims to gain an insight into how mechanical boundary conditions affect the production of ECM components, their remodeling, and the feedback of the altered mechanical cell environment on these processes. The influence of cyclic mechanical loading (f=1 Hz, 10% axial compression) and scaffold stiffness (E=1.2 and 8.5 kPa) on the mechanical properties of fibroblast-seeded scaffold constructs were investigated in an in vitro approach over 14 days of culture. To do so, a newly developed bioreactor system was employed. While mechanical loading resulted in a clear upregulation of procollagen-I and fibronectin production, scaffold stiffness showed to primarily influence matrix metalloproteinase-1 (MMP-1) secretion and cell-induced scaffold contraction. Higher stiffness of the collagen scaffolds resulted in an up to twofold higher production of collagen-degrading MMP-1. The changes of mechanical parameters like Young's modulus, maximum compression force, and elastic portion of compression force over time suggest that from initially distinct mechanical starting conditions (scaffold stiffness), the construct's mechanical properties converge over time. As a consequence of mechanical loading a shift toward higher construct stiffness was observed. The results suggest that scaffold stiffness has only a temporary effect on cell behavior, while the impact of mechanical loading is preserved over time. Thus, it is concluded that the mechanical environment of the cell after remodeling is depending on mechanical loading rather than on initial scaffold stiffness.
Introduction
Cells actively modify their mechanical microenvironment by means of forming, contracting, and degrading extracellular structures. This is based on their ability to sense this environment and to process the information into a specific response.3–5 In this context, substrate elasticity has gained more and more attention since it has been found to influence cell morphology, adhesion, motility, proliferation,6–8 and cell differentiation.9,10 A well-regulated force interplay between the cell and the extracellular matrix (ECM) is the basis for a physiological cell behavior, including many of the previously mentioned functional parameters. In this interplay, contracting (actin cytoskeleton) and stabilizing elements (e.g., microtubuli) inside the cell add up to a net contraction force that is transferred to the counterpart, the ECM, via focal adhesions. 11 The force that a cell is able to build up and the resulting ECM/substrate deformation are assumed to serve as an input parameter for cell behavior but also for cell force regulation.12,13
There are indications that substrate elasticity influences the production and organization of ECM components. 14 However, the consequences of these alterations for the local mechanical environment remain largely unknown. Also, an alteration of cell contraction forces with changes in substrate or boundary stiffness has been observed.7,15–17 The adaptation of ECM formation and cell contraction cannot be seen just as a one-way response to the cell's microenvironment, because they actively change its mechanical properties. In fact the cellular response seems to be part of a mechanical regulating mechanism between the cell and its microenvironment toward an equilibrium state.
Extrinsic mechanical loading is a further mechanical parameter and results in substrate and cell straining if applied to tissues or cell-seeded constructs. Loading interferes with the earlier mentioned cell–ECM force equilibrium and provokes cell responses. Especially in the field of bone healing, various in vitro and in vivo models have shown that mechanical loading regulates cell proliferation, 18 cytokine secretion, and angiogenesis19–21 and finally the healing outcome.22,23 On the basis of gene expression analysis the influence of mechanical stimulation on ECM synthesis has been demonstrated.24–27 This suggests that mechanical loading might lead to an increase in ECM stiffness and, consequently, a sustained alteration of the cell–ECM force balance.
A multitude of in vitro systems has been developed for the application of mechanical stimuli to cell constructs, mostly in the context of cartilage28–30 or bone.31,32 Just recently, an increasing number of systems have been presented that combine mechanical stimulation and online monitoring of changes in mechanical properties.33–38 These changes in mechanical properties are obtained to serve as a quality parameter for the evolution of mechanical competence during culture of tissue engineering constructs. However, a link between the cellular response to mechanical loading or altered substrate stiffness and changes in individual mechanical parameters (elastic modulus, relaxation of the compression force, and its elastic component) is missing. Thus, it was not yet possible to directly link cellular processes like ECM remodeling or traction-force-induced ECM compaction with changes in macroscopic material properties.
In this work we aim to investigate how cells respond to, and actively tune, the mechanical environment by ECM remodeling and contraction. To do so, we combined a variation in substrate elasticity with the application of cyclic mechanical loading. A newly developed bioreactor system was utilized that allows long-term culture of cell-seeded constructs in combination with the application of mechanical loading and monitoring of the construct's mechanical properties. With the new bioreactor system, mechanical characterization of cell-seeded constructs was performed by precise mechanical testing over the full 2 weeks period of culture. Macroporous collagen-I scaffolds of two different stiffnesses seeded with human primary fibroblasts were chosen as artificial tissue constructs. Changes in Young's modulus, relaxation force, and the elastic portion of compressive force were compared to the amounts of ECM components (collagen-I and fibronectin) produced and matrix-degrading metalloproteinase MMP-1 secreted to the medium. The aim of this study was to gain new insights into the cellular strategy of creating microenvironments based on ECM formation and remodeling processes.
Materials and Methods
Bioreactor
Each bioreactor unit (Fig. 1a) is divided into a cell culture unit and a mechanical unit for load application. The cell culture unit contains a reactor chamber, a medium reservoir with implemented gas exchange, and a micropump to provide medium circulation through the components (Fig. 1b). The mechanical unit applies displacement patterns to the samples inside the bioreactor chamber and measures the loads experienced by the samples.
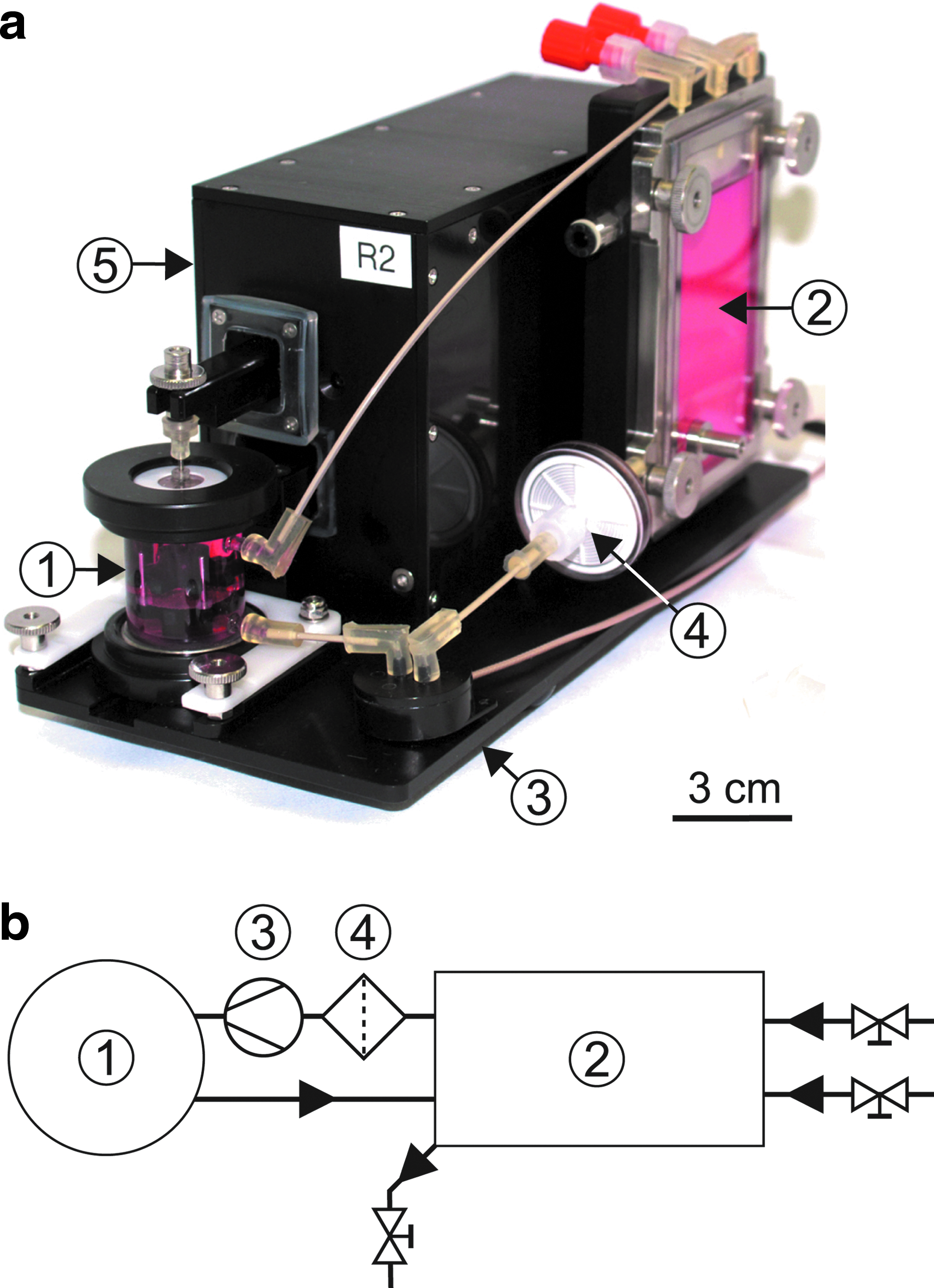
Bioreactor as completely mounted unit
Mechanical unit
The mechanical unit contains two mechanically active elements, a linear actuator (M-232.17; Physik Instrumente [PI] GmbH & Co.) and a piezo actuator (P-601.4SL; PI). The linear actuator provides a slow movement over a long travel range (vmax=2.5 mm/s, d=17 mm) while the piezo actuator provides a highly dynamic movement with high resolution, however, restricted by a small travel range (d=400 μm). By combining these two drives, a long travel range and a dynamic movement pattern were realized. Figure 2a shows how the movement of both drives is transferred to the sample: a wedge driven by the spindle of the linear drive leads to a vertical movement of a bolt. The pivoted lower arm of the bioreactor is resting on the bolt with a force sensor mounted inside the arm detecting the load applied. The force limit of the sensor is 15 N with a resolution of 1.5 mN. The displacement of the piezo actuator is increased to threefold by a pivoted cantilever. The cuboid housing of the mechanical unit includes all components. Mechanically moving parts are sealed by silicone gaskets to allow spray disinfection of the whole unit.
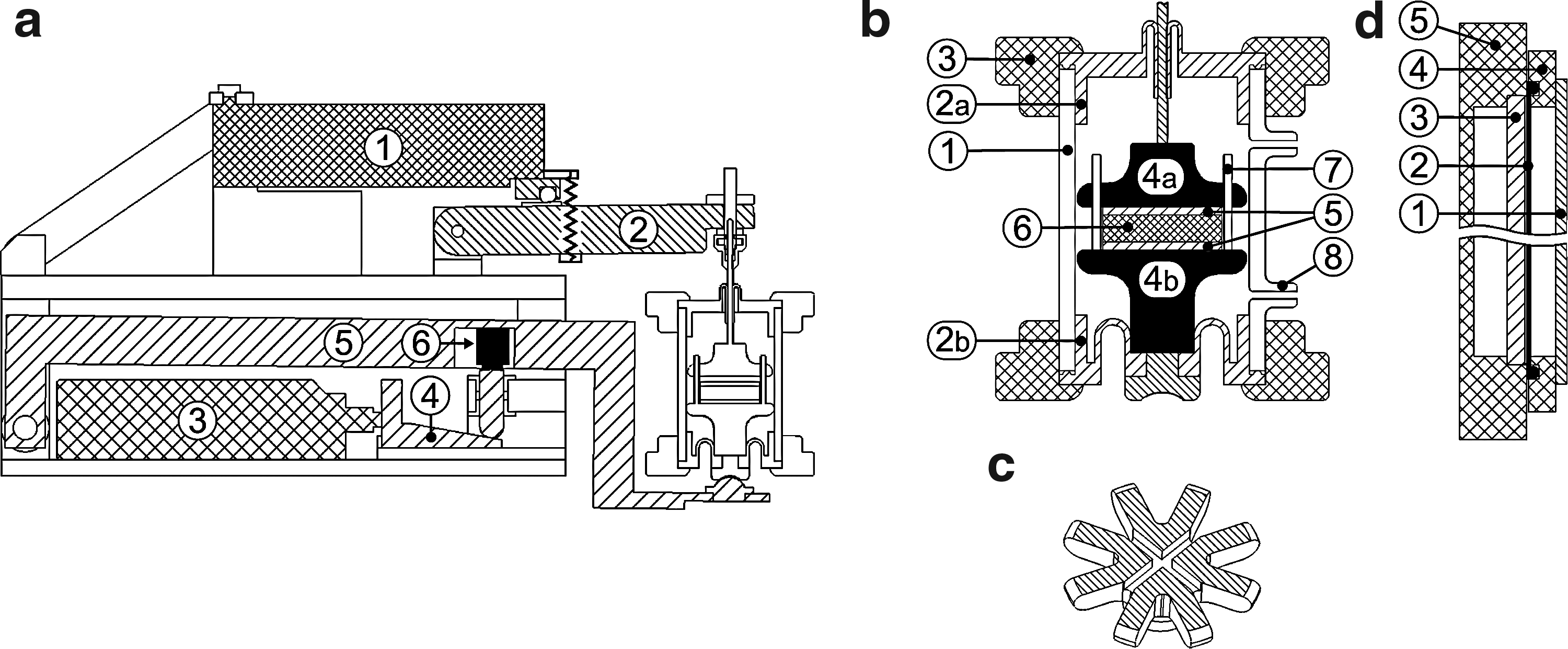
Technical details of the individual bioreactor components in cross-section and three-dimensional schematic view: mechanical unit
Culture unit
The cylindric glass housing of the reactor chamber shown in Figure 2b includes a medium inlet (lower) and outlet (upper adapter). The chamber is sealed by custom-made silicon rolling diaphragms (Elastosil RT625 two-component silicone) to allow displacement of the connected plungers (polyether ether ketone [PEEK]) and provide full sealing. Stainless steel pins keep the sample centered, while cylindrical mesh disks (PEEK) on the top and the bottom sides of the sample provide a flat bearing area with medium supply through mesh holes. The plungers have star-shaped crown and additionally offer two perpendicular grooves to support medium transport to the sample (Fig. 2c). The lower plunger is guided by the silicone membrane and the bearing to the lower arm of the mechanical unit only, avoiding friction between the plunger and the glass chamber. This allows the full load applied to the samples to be measured by the force sensor. Gas exchange was realized through a 200-μm-thick custom-made silicone membrane (Elastosil RT625) implemented between the gas side and the medium side of the medium reservoir (Fig. 2d).
Multiple connections are implemented in the reservoir for medium exchange, sampling, or introduction of supplements. A sterile 0.2-μm filter is mounted via a disposable Luer-Lock polyethylene tube to one of the connectors to balance pressure changes in the medium circuit (not shown in Fig. 1a). An oxygen sensor spot (PreSens—Precision Sensing GmbH) is glued to the glass window of the reservoir to optically detect oxygen partial pressure of the medium by a fiber optic attached from the outside.
The three components of the biological unit (chamber, reservoir, and micropump) are connected by PEEK tubes (low gas diffusion coefficient). A sterile 5-μm filter (MinisartCA; Sartorius Stedium Biotech S.A.) is protecting the micropump from particles. The pH value of the medium can be detected by mounting a flow-through minisensor into the circuit.
All components in contact with cell culture medium (materials: glass, stainless steel, PEEK, silicone, and polypropylene) can be sterilized in an autoclave for repeated usage. All other components can be spray disinfected. The bioreactor units are located in a dry incubator (INE 600; Memmert GmbH & Co.) at 37°C.
Besides the eight bioreactor units, the system includes motor controllers for piezo (E-621.SR; PI) and linear drives (C-863 Mercury™; PI), two gas-mixing units (MX 4/1; DASGIP AG), a micropump controller (in house development), a measurement data acquisition unit (Spider8; Hottinger Baldwin Messtechnik GmbH) and the transmitter unit for optical oxygen/pH detection (OXY-4 mini/pH-4 mini; PreSens). The bioreactor system is fully computer controlled via a LabView interface. Complex protocols, including all motor movements, pump switching on/off, and change of gassing, can be build to run the bioreactor automatically over days and weeks while data (piezo and linear drive positions, force sensor data, pH, and pO2 values) are recorded.
Scaffolds
Macroporous scaffolds (Optimaix®; Matricel GmbH) were used as cell carrier materials. The scaffold offers an ideal matrix for cell attachment and tolerates long-term cyclic compression without major changes in its dimensions. The free space in the porous scaffold enables cell spreading, migration, and the formation of the cell's own ECM (see Fig. 9a–c). Scaffolds consisted of porcine collagen-I in a highly orientated structure with a mean pore diameter of 84 μm. Two groups of scaffolds with 1.5 (=Shigh) and 1.1 vol-% collagen (=Slow) were used with elastic moduli of 8.5±0.9 kPa and 1.2±0.4 kPa, respectively, measured in buffer solution using the bioreactor. No differences in pore structure and pore size were observed between both scaffold types (see Fig. 9a, b). Cylindrical scaffolds with a diameter of Ø=13 mm and a height of h0=3 mm were used. For bioreactor biocompatibility testing, scaffolds with Ø=5 mm were chosen.
Cells and culture medium
Primary dermal human fibroblasts were used in passages 4–6. Cells were expanded in Dulbecco's modified Eagle's medium (DMEM, catalog # 41965; Gibco, Invitrogen) supplemented with 10 vol-% fetal bovine serum (# S 0115; Biochrom AG), 1 vol-% penicillin/streptomycin (# A 2213; Biochrom AG), and 1 vol-% nonessential amino acids (# K 0293; Biochrom AG) in a humid incubator with 5% CO2. For bioreactor experiments additional 1.36 mM ascorbic acid was added to the medium to enhance ECM formation.
Cell seeding into scaffolds
Cells were trypsinized and brought into suspension in a concentration of 7.5×103 cells/μL in DMEM. Scaffolds were dipped into the cell suspension which they immediately absorbed. They were subsequently transferred into a 12-well culture plate without additional medium. Cell-seeded constructs Chigh and Clow obtained from scaffolds Shigh and Slow were kept in the humid incubator for 40–60 min to allow cell adherence. Before being transferred into the bioreactors, cell constructs were washed in DMEM to remove unattached cells.
Immunoassays
ECM-related proteins and peptides were analyzed from conditioned medium using ELISAs for Procollagen Type I C-Peptide (# MK101; Takara Bio, Inc.), fibronectin (# 607; American Diagnostica), and pro-matrix metalloproteinase-1 (pro-MMP-1) (Quantikin® DMP100; R&D Systems, Inc.). Tests were performed according to manufacturer's guidelines.
Glucose and lactate analyses
Five hundred microliters of conditioned medium from each bioreactor was given to the Interdisziplinäres Liquorlabor of the Charité—Universitätsmedizin Berlin for routine analysis of glucose and lactate concentrations.
Immunostainings
At the end of the bioreactor experiments, cell constructs were fixated in 4% paraformaldehyde overnight, quenched in 25 mM ammonium chloride solution (to block free aldehyde groups), placed overnight in excessive amount of cryoblock (# 41302000; Medite GmbH), and frozen in cryomolds on a liquid nitrogen cold aluminum block. Cryosections of 50 μm were prepared (LEICA CM3050S cryostat) and stained for actin filaments (Alexa fluor 488 Phalloidin, # A12379; Invitrogen), fibronectin (anti-fibronectin antibody, # ab23750; Abcam plc), and cell nuclei (DAPI, # D3571; Invitrogen).
Analysis of cell number
For each sample, six representative images of DAPI-stained cryosections were taken at 10-fold magnification across a section. The cell density was obtained by image analysis using image J. Based on the construct's outer dimensions at the end of the experiment, the total cell number per construct was calculated.
Analysis of construct's material properties
Material parameters of compressive force Fc,max, elastic component of the compressive force Felast, and force relaxation Frelax=Fc,max − Felast were derived from the force–time curves Fc(t) of scaffold material testing as illustrated in Figure 3. Felast is created by the elastic structures in the construct, for example, the scaffold walls and the ECM. Frelax represents the portion of the compressive force that is created by the resistance of fluid flow in the scaffold pores upon compression. It might also contain nonelastic components of scaffold and ECM. Young's modulus E (stiffness) was calculated as E=(ΔF/Δt)·h/(A·vc), with h denoting scaffold height (obtained from the position of the bioreactor's lower plunger), A the cross-sectional area, and vc the compression velocity. ΔF/Δt was analyzed from the slope of the curves Fc(t) within the last 2 s of linear compression. Construct diameter, necessary for the calculation of A, was assumed to decrease proportionally with scaffold height. Additional details can be found in the experimental procedure.

Sketch of plunger displacement s(t) (dot-dash line) and resulting mechanical testing curve Fc(t) (solid line) illustrating the analyzed parameters E, Fc,max, Frelax, and Felast (see text).
Statistical analysis
Data are presented as mean values with standard error of the mean if not indicated otherwise. For boxplots, 25% and 75% of data points were chosen as lower and upper limits. The median is shown as horizontal line, mean value as filled diamond, and outliers are indicated by diagonal crosses. Statistical significance was analyzed using Student's t-test with normal distribution proved for all data sets. For data from immunoassays (Fig. 8) a two-way ANOVA, including a Holm-Bonferroni correction, was performed to analyze the influence of the factors load and scaffold stiffness. OriginPro 8G SR4 by OriginLab Corporation was used for all statistical analyses.
Bioreactor validation
Culture unit
Biological compatibility of the bioreactor components was assessed by (1) exposure of two-dimensional (2D) cell layers to medium conditioned in the bioreactor and (2) culture of cell-seeded scaffolds inside the bioreactor.
(1) Medium was conditioned over 3 days inside the bioreactor including all components used for later experiments. Fibroblasts were seeded in a 96-well plate at a density of 2×103 cells per well and exposed to conditioned medium 1 day after seeding. MTS-based CellTiter 96® AQueous cell proliferation assay (Promega) was performed at day 1 (value of initial cell number) and 3 days after application of conditioned medium (number of cells after proliferation). Increase in absorbance (Tecan Infinite M200PRO microplate reader) was compared with cells cultured in unconditioned medium according to the same protocol.
(2) Collagen scaffolds Shigh (Ø=5 mm) were seeded with fibroblasts as described before, kept in static culture without additional medium for 40 min to allow cell adherence, and subsequently transferred either into bioreactor units or placed into wells of a six-well plate with 4 mL medium per well added. Cell-seeded scaffolds were cultured for 3 days while medium was exchanged every day for constructs in static culture to account for the smaller medium volume in comparison to the bioreactors. Number of cell nuclei was counted from DAPI-stained cryosections prepared along the symmetry axis of the scaffold and compared with values of scaffolds kept for 3 h in static culture only (control).
Mechanical unit
The mechanical unit of each bioreactor unit was validated against a BOSE ElectroForce® TestBench equipped with a 225 N force sensor. For comparability reasons, identical plungers as used in the bioreactor were mounted onto the BOSE TestBench system. A cylindrical dummy prepared from silicone (Ø=4 mm, h=3 mm) was used to ensure elastic material properties and comparability between subsequently performed experiments in both systems. The dummy was glued to PEEK mesh discs on the top and on the bottom sides, identical to the ones used in the bioreactor experiments. A preload of 50 mN was defined in all tests. Linear axial compression of the dummy was performed with a speed of vc=50 μm/s until displacements of 50, 100, 150, 300, 500, 700, and 1000 μm, respectively, were reached. The testing for each displacement was repeated three times starting from zero position. Data were analyzed for compressive force Fc,max. Additionally, the Young's modulus E was analyzed by calculating the linear curve fit for three displacement intervals 25–100, 100–300, and 300–500 μm of curves with 500 μm total displacement exemplarily.
Experimental procedure
Before each experiment, force sensors were calibrated by placing a reference weight on the lower arm of the mechanical units. Culture units of the bioreactors were built up under sterile conditions in a laminar flow bench and filled with culture medium. Cell-seeded scaffolds were transferred into the individual reactor chambers, chambers were closed, and culture units were mounted onto the mechanical units inside the incubator. To bring samples into contact with the upper plunger, a sinusoidal oscillation of the upper arm was applied (f=0.2 Hz, 2A=100 μm) while the lower plunger was moved up at a constant speed of 10 μm/s until a force magnitude of 6 and 10 mN for Slow and Shigh, respectively, was detected by the force sensor. From this starting position, the experimental protocol was executed.
Starting with a 15-h resting phase, a sequence of 3-h cyclic compression (f=1.0 Hz, 2A=300 μm=10% scaffold height) and 5 h of resting and mechanical testing was repeated consistently over the 2 weeks of experimental period. Mechanical testing was performed by a linear compression (300 μm, vc=50 μm/s), a holding period of 20 s and a linear retraction to zero position at the same speed. Testing was repeated every 30 min during the resting phase. All movements for mechanical testing and cyclic compression were performed by the upper plunger. In the middle of the resting phase (at 2.5 h), sample position was rechecked by applying the same protocol sequence as for initial sample positioning described previously. This repositioning assured that samples were always in contact with both plungers even if the sample contracted over time. The change of the lower arm's position thus provided information about the change of sample height. The protocol for the control group without mechanical loading was identical just without cyclic compression during the 3-h period (2A=0 μm). For this group the term “unloaded” was used. It indicates that no additional cyclic compression is applied, even though mechanical perturbations are induced by mechanical testing and sample repositioning.
At day 2, 2 mL of medium was withdrawn from the reservoir for analysis. Medium in the reservoir was exchanged at day 7 and again at day 11. Medium collected at days 2 and 7 was cryopreserved for later analysis.
Results
System validation
Biocompatibility of the culture unit
The 2D MTT assay did not show a cytotoxic effect of the medium conditioned in the bioreactor (see Table 1, top). For three-dimensional (3D) scaffolds cultured in the bioreactor, the increase of cell number was slightly higher than for those ones cultured under static conditions (see Table 1, bottom). This can be explained by a better supply with oxygen and nutrients as a consequence of the medium circulation compared with static culture. Due to the small dimensions and the open porous structure of the scaffolds, the effect, however, was not very pronounced. Both tests clearly proved the biocompatibility of the bioreactor's culture unit.
Two-dimensional MTS test with conditioned medium (top) and cell counting of three-dimensional cell-seeded constructs (bottom). The given errors represent the standard deviation.
Material testing and mechanical loading
Mechanical testing performed with a silicone dummy in the bioreactor versus the BOSE TestBench showed good agreement between the systems. Data points of Fc,max showed a linear arrangement on or very close to the diagonal line of Fc,max BR=Fc,max BOSE (Fig. 4a) with a regression coefficient of R 2 =0.994. The data point for highest compression (1000 μm) shows a slightly lower value for the bioreactor since the force limit of the piezo actuator was reached. Also, values for Young's modulus E showed good agreement with R 2 =0.986 for a linear regression based on EBR=EBOSE (Fig. 4b).
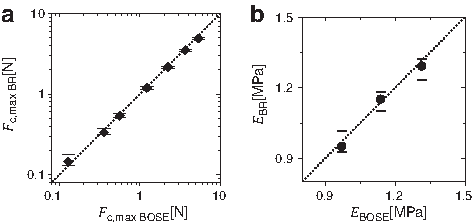
Mechanical validation of the bioreactor units. mean values of compressive forces measured with the bioreactors Fc,max BR plotted against Fc,max BOSE of the BOSE TestBench
Compressive force and scaffold height
During the 2-week culture period the mechanical properties of cell-seeded constructs showed characteristic changes for the individual experimental groups. The compressive force Fc,max, shown in Figure 5a, increased clearly for constructs Clow in an almost linear way.
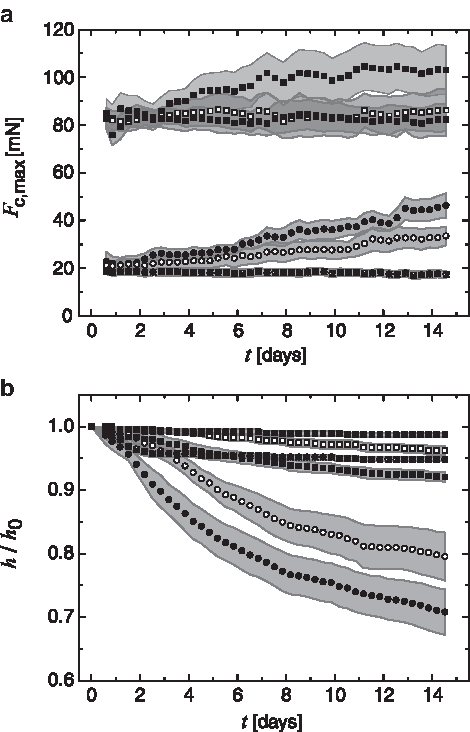
Online monitoring of construct properties. Change of Fc,max over the cultivation time
Loaded constructs showed a more pronounced increase (higher slope) compared with the unloaded group. For constructs Chigh, an increase of Fc,max was observed only for the loaded group and only up to day 7. Subsequently Fc,max was stable up to day 14. For the unloaded group, no pronounced changes could be observed over the culture period from day 0 to 14. Cell-free scaffolds Slow and Shigh did not show any significant change in Fc,max over time in the loaded and nonloaded groups. Only the data for loaded cell-free scaffolds are shown in Figure 5 for easier reading. At day 7 and 11 perturbations in the curves are visible. These are caused by potential cellular responses to the medium exchange performed at the two time points.
For soft constructs Clow, a pronounced contraction was observed, leading to a reduction of scaffold height of h/h0=0.79 and 0.71 at day 14 for the unloaded and loaded groups, respectively (Fig. 5b). The increased reduction in height observed in the loaded group is taking place predominantly between day 0 and 7 (h/h0=0.86-fold for the unloaded vs. 0.79-fold for loaded group at day 7), while the additional change from day 7 to 14 was almost comparable.
The change of sample height for Chigh was dramatically lower than for Clow with a decrease to h/h0=0.96 and 0.92 at day 14 for the unloaded and loaded groups, respectively. Thus, as for Clow, loading resulted in a stronger decrease of scaffold height. The height decrease for all cell-seeded groups was observed to be decelerating over the 2-week culture period, indicating a possible limit to be reached if cultivation time would be extended (saturation behavior). Following a slight reduction during the first 48 h (h/h0=0.96 and 0.99), sample height of cell-free constructs Slow and Shigh was almost constant (h/h0=0.95 and 0.98 at day 14).
Young's modulus and poroelastic material properties
Even though mechanical testing is performed continuously during the experiment, values for Young's modulus can only be calculated if the cross-sectional area of the scaffolds is known (see the Materials and Methods section). For day 0 the initial scaffold diameter as provided by manufacturer and for day 14 the diameter analyzed from images taken at the end of the experiment were taken into account. Scaffold diameter at day 7 was calculated based on the values of day 0 and 14 and the data recorded for the change in scaffold height. To do so, it was assumed that scaffold height and diameter decrease proportionally.
Unloaded cell constructs Clow showed a steady increase of Young's modulus (E) leading to a 2.8-fold higher value at day 14 compared with day 0 (Fig. 6a). This increase was significant (p=0.01). Constructs cultured under cyclic mechanical loading showed a 2.6-fold, also significant, increase (p=0.01). Difference between mean values EClow, load=4.38 kPa and EClow, no load=3.75 kPa at day 14 was obvious, however, not statistically significant due to the large data point variations. In unloaded cell constructs Chigh, such a clear trend could not be observed. While an increase of Young's modulus from day 0 to 7 is noticeable, construct stiffness seems to decrease again from day 7 to 14. For loaded constructs Chigh, a trend for an increase in Young's modulus can be observed from day 0 to day 7 and 14. However, due to the large variation of the measurement points, this difference was not significant. In contrast to cell-seeded scaffolds, cell-free scaffolds of both groups Slow and Shigh did not show an increase in Young's modulus over time. For easier reading, only data of mechanically loaded scaffolds with larger expected changes are shown in Figure 6a. Unloaded cell-free scaffolds as well did not show a change over time.
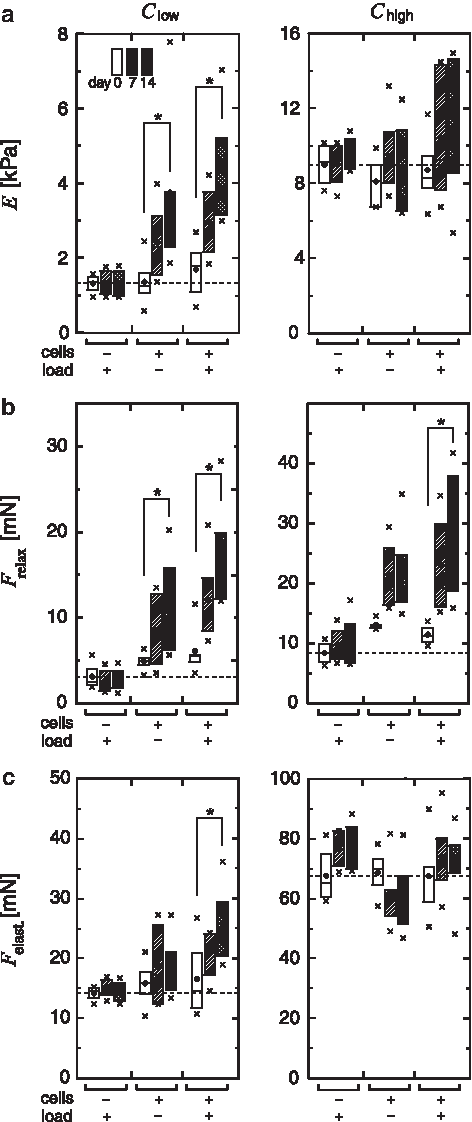
Boxplots of the parameters E
Values for the force relaxation Frelax are shown in Figure 6b. For constructs Clow, the change of Frelax over the time of culture was very similar to the change of Fc,max in Figure 5a; a significant increase from day 0 to 14 was observed, which is even more pronounced in the loaded (2.9-fold, p=0.01) versus the unloaded (2.5-fold, p=0.02) group. Also for Chigh, an increase in Frelax was observed. A 1.7-fold higher value in the unloaded and a significantly 2.4-fold higher value (p=0.04) in the loaded group was seen at day 14 compared with day 0. These values are high compared with the relatively mild increases in Fc,max and E (see Figures 5a and 6a). For cell-free constructs Slow and Shigh, Frelax did not change over time.
The elastic component of the compressive force Felast showed changes comparable to those in E for Clow (see Fig. 6c) with a 1.2-fold increase from day 0 to 14 in the unloaded, and a significant 1.6-fold increase in the loaded group (p=0.04). For constructs Chigh, a decrease of Felast in the unloaded group is visible from day 0 to 14 in contrast to a slight increase of E. The increase of Felast in the loaded group is again comparable to the trend in E.
The change of the individual mechanical parameters over time is additionally illustrated in Figure 7, where mean representative curves of the force Fc(t) recorded during mechanical testing are shown for each individual group. It is obvious that for cell-seeded constructs Clow parameters Fc,max, E, Frelax, and Felast all increased over the culture period, most distinct in the loaded group (see Fig. 3 for parameter reading from curves). In cell constructs Chigh, no changes of Fc,max and E were visible for the unloaded group. At the same time, Frelax increased from day 0 to 7 (Felast decreased) and slightly decreased again from day 7 to 14 (Felast increased). In the loaded group, Fc,max, E, Frelax, and Felast all increased from day 0 to 7 while from day 7 to 14 Fc,max and E were constant and only Frelax increased slightly (Felast decreased).
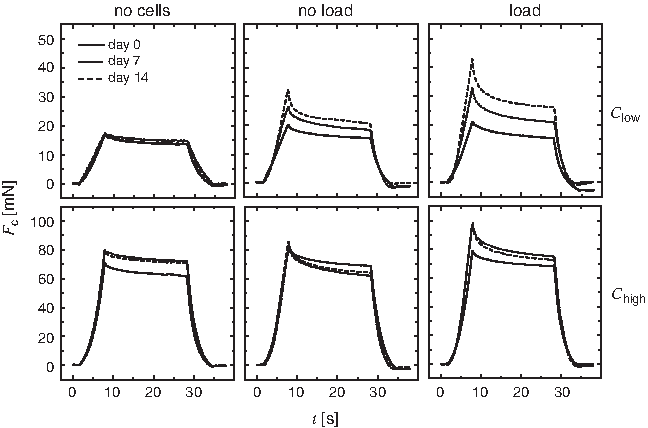
Representative force–time curves Fc(t) for Clow and Chigh in the unloaded and loaded groups compared with cell-free scaffolds. The depicted curves are the averages of Fc(t) of all constructs in the according group at the time points day 0, 7, and 14.
Cell proliferation
The analysis of the cell number from DAPI-stained histological sections revealed fundamental differences in cell proliferation between Clow and Chigh (Table 2). In constructs Clow, a relatively low 1.6 and 1.5-fold increase in total cell number from day 0 to 14 was observed in the unloaded and the loaded groups, respectively. In contrast, total cell number in Chigh increased to 2.5 and 2.3-fold for the unloaded and loaded groups, respectively. Lower cell numbers in Clow are, however, not a direct consequence of substrate stiffness on cell proliferation. The increase in cell number from day 0 to 2 was analyzed from additional samples in static culture and showed no significant differences between Clow and Chigh (see Table 2, cell proliferation). Also seeding density (cell density at day 0) was comparable. Additionally, Table 2 shows that the cell density inside the constructs was almost identical for all groups at day 14. This suggests that a maximum value triggering cell apoptosis was reached. 39 Thus, cell number was assumed to be comparable until cell density goes into saturation and from this point on to be governed by construct volume with higher rates of apoptosis and lower cell numbers in fast-contracting constructs Clow. Analysis of glucose consumption and lactate production at day 7 (before the first medium exchange) confirmed a higher cell number in constructs Chigh compared with Clow by a 1.3/1.2-fold higher glucose consumption and a 1.3/1.3-fold higher lactate production for the unloaded/loaded group. In agreement with the analysis of the cell number, no significant differences between the unloaded and the loaded groups were observed.
Cell proliferation until day 2 was calculated as number of cells (day 2)/number of cells (day 0), cell densities inside the scaffolds were counted from DAPI-stained cryosections at day 0 and 14, and cell numbers at day 14 were calculated based on the construct's volume. The mean seeding cell number was 2.2×106 cells/scaffold. Additionally shown are glucose consumption Δc=cday 0 − cday 7 and lactate accumulation up to day 7 as analyzed from the medium.
Values were obtained from experiments in static culture in scaffolds with Ø=5 mm.
Protein secretion
The analysis of culture medium conditioned for 2 and 7 days showed that at both time points mechanical loading leads to a significantly higher production of procollagen-I compared with unloaded constructs while no influence of scaffold stiffness was observed (Fig. 8a). Similar to the trend observed for procollagen-I, higher amounts of fibronectin were found in medium from loaded compared with unloaded constructs with a significant difference at day 7 (Fig. 8b). In contrast to procollagen-I, higher values were also found in constructs Chigh compared with Clow. Statistical analysis at day 7 confirmed that the influence of scaffold stiffness was significant.
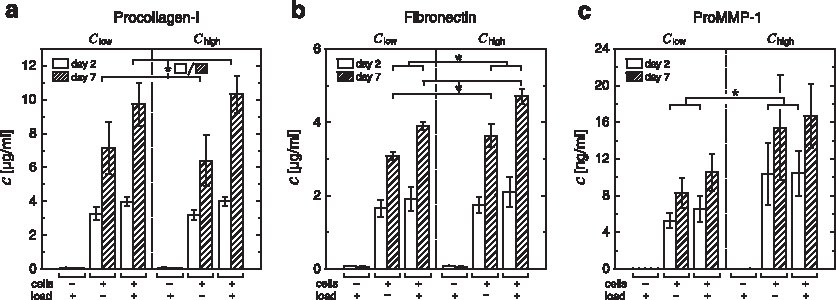
Immunoassay (ELISA) analysis of procollagen-I
Proform of collagenase-1 (pro-MMP-1) was found in a higher concentration for loaded versus unloaded constructs Clow. For stiffer constructs Chigh, this trend could not be confirmed due to the higher deviation of the measured values. Noteworthy, the comparison between Clow and Chigh shows a substantially higher amount of pro-MMP-1 for cells seeded in stiffer scaffolds. The values were found to be 2.0 and 1.6-fold higher at day 2 and 1.9 and 1.6-fold higher at day 7 for unloaded and loaded constructs, respectively. For day 2 the influence of scaffold stiffness on MMP-1 secretion was significant.
In the medium exposed to cell-free scaffolds Slow and Shigh for 2 and 7 days, only minimal amounts of procollagen-I, fibronectin, and pro-MMP-1 were found (see Fig. 8a–c). This indicated that these molecules are produced as a response of the cells to the matrix environment after scaffold seeding.
Histology
The formation of ECM inside the scaffold pores of all groups could be documented histologically. Collagen and fibronectin fibers stretched between scaffold walls and created a dense ECM network (compare Fig. 9d and 9e). Cells are fully attached to the matrix in all groups and the well-developed actin cytoskeleton arrangement indicates tension applied by the cells to the ECM network (Fig. 9f). Cells are visible inside the self-created ECM network (day 14) rather than on the scaffold walls only, where they were initially located at day 0 (Fig. 9g). ECM density appears to be slightly higher in constructs Clow, resulting from ECM compaction due to scaffold contraction (Fig. 9e). This contraction can clearly be seen in the sirius red staining of Clow in contrast to Chigh.

Representative scanning electron microscopy (SEM) and histological images of empty scaffolds and cell-seeded constructs at day 14. SEM images of the pore structure cut perpendicular to the main pore orientation for scaffolds Slow
Discussion
A new bioreactor system has been presented that was built for culturing cell-seeded constructs under defined mechanical loading conditions and that allowed online monitoring of their mechanical properties. The bioreactor is a further development of a previously published system 40 that has been used extensively in our labs to investigate the biochemical cellular response upon mechanical loading. In contrast to existing systems,28,33–38 the bioreactor system presented here provides the possibility for precise online mechanical testing of very soft samples (≈1 kPa) comparable to established offline systems like the BOSE TestBench. Additionally, the individual bioreactor units contain closed medium circuits with implemented gas exchange and thus provide a closed loop system except the necessity for a temperature-controlled environment. They are not bound to humidified CO2 incubators. Mechanical validation and biocompatibility testing proved the full functionality of the bioreactor system.
For our investigations we chose fibroblasts since they are known to be the key player in synthesizing ECM components, like collagen, fibronectin, and elastin, 41 in connective tissue. Together with other components these polymerized molecules give mechanical strength to the matrix and thus enable tissue functionality. Further, the ability of fibroblasts to contract existing ECM structures is essential for tissue healing and wound closure.42–45 Tissue contraction is a result of mechanical forces46,47 applied by the cell cytoskeleton being attached to ECM fibers via focal adhesions.11,48 Fibroblasts thus play a central role in ECM and tissue remodeling and are essential for wound healing, also in respect to bone regeneration.
The utilization of an appropriate carrier material is a premise for the investigation of the cell's contribution to ECM synthesis and contraction. A variety of polymer gels and macroporous scaffolds have been used for 3D cell culture. Collagen gels are widely used to study cell-based gel contraction 49 since they provide a tissue-like biochemical environment fostering cell attachment. Synthetic gels, for example, from polyethylene glycol 50 or alginate 51 offer the possibility to tune the mechanical properties over a wide range. Cell adhesion and the application of contractile forces might, however, be altered in gels compared with the physiological situation. Macroporous scaffolds are advantageous for 3D culture at high cell densities as they provide enhanced nutrient and oxygen supply by medium flow through scaffold pores. Stiffness of scaffolds can be tuned by variations in crosslinking (e.g., collagen-GAG-scaffolds 52 ) or by changing the solid content to modify scaffold wall thickness. An advantage of the latter approach is that the surface the cells see is chemically identical. For the earlier mentioned reasons a macroporous collagen-I scaffold with a variation in collagen solid content was chosen.
Using the newly developed bioreactor, we performed a study with the goal to gain an insight into the cellular processes involved in the remodeling of the cell's mechanical environment. It has been shown that substrate stiffness is one key regulator of cell behavior and cell fate decision.10,51,53 Thus, understanding the way in which cells respond to and remodel the mechanical microenvironment has gained more and more attention.16,54 Deliberately, two distinctly different mechanical environments were defined by scaffold elasticities with Young's moduli of 1.2 and 8.5 kPa. This allowed a study of cell response and cell-induced changes in construct mechanical properties by ECM formation.
The increase in mechanical stiffness (E) over the time of culture showed to be a direct consequence of the formation of ECM together with the cell-induced contraction of scaffold and ECM and not of one of the two alone. The dense network of ECM fibers forming inside the scaffold pores, as visible in Figure 9, gives additional stability to the scaffold walls, leading to an increase in Felast for constructs Clow over time (Fig. 6c). This is accompanied by an increasing resistance for liquid movement in the pores that results in higher values for Frelax (Fig. 6b), observed in constructs Clow and Chigh. Additionally to the formation of ECM, the findings suggest that cell-induced construct contraction increases the mechanical stiffness. In accordance with this assumption, the increase in Felast from day 0 to 14 is higher for highly contracting constructs Clow than for less-contracting constructs Chigh in the unloaded and the loaded groups. This is true even though the amount of fibronectin produced is higher for stiffer scaffolds Chigh, while procollagen-I production is comparable.
It has been shown that the mechanical environment regulates cellular production of ECM components.16,24,25,55,56 We found a significant upregulation of procollagen-I and fibronectin as a consequence of cyclic mechanical compression of 10% sample height (Fig. 8a, b). Also, matrix-degrading MMP-1 secretion was enhanced by loading (for constructs Clow) consistent with earlier findings for MMP-2 secreted by mesenchymal stromal cells embedded in fibrin gels.19–21 Altogether this indicates enhanced ECM remodeling as a response to mechanical loading.
It is noteworthy that the production of procollagen-I and fibronectin on the one hand and MMP-1 on the other hand showed distinct time dependencies (Fig. 8). Procollagen-I and fibronectin production slows down within the first week as indicated by a ratio Rc=(cday 7 − cday 2)/cday 2 between 1.2 (Clow, unloaded) and 1.6 (Chigh, loaded) for procollagen and between 0.9 (Clow, unloaded) and 1.3 (Chigh, loaded) for fibronectin (minimum and maximum values of all four groups). A value of Rc=2.5 would represent a linear increase. The reduction in production of ECM components over time might indicate that the microenvironment is slowly being adapted to the cell's (e.g., mechanical) target value. From this perspective, a saturation behavior would be observed. MMP-1 release in contrast is almost restricted to the first 2 days with very low Rc values between 0.5 (Chigh, unloaded) and 0.6 (Chigh, loaded). Analysis of immunohistological sections at different time points revealed that the cells, initially only attached to the scaffold, are found connected to newly formed ECM (mostly fibronectin) already after a few days (data not shown). Together these observations indicated that MMP-1 is produced primarily as a consequence of direct interaction between the cells and the collagen scaffold (see also Supplementary Data; Supplementary Data are available online at www.libertpub.com/tea). The details of that interaction remain so far, however, unclear and are subject of further investigation.
Interestingly, the production of matrix-degrading MMP-1 is significantly higher in constructs Chigh than in Clow, in contrast to procollagen-I (see Fig. 8c). The values are 2.0 and 1.6-fold higher in the unloaded and loaded groups at day 2, respectively. This might be a consequence of the higher mechanical stiffness of the cells' initial microenvironment resulting from thicker scaffold walls; even though the amount of collagen is higher in scaffolds Shigh, the mean pore size is comparable. Thus, the surface the cells attach to is expected to be comparable in structure and dimension, only the bulk of collagen below it differs. Cells seem to sense this difference via the application of mechanical, contractile forces. In the one case (Clow), cellular forces are high enough to deform the scaffold walls in a way that initially straight plains become wavy (Fig. 9e, left). In the other case (Chigh), the walls are too stiff to be deformed to such an extend (Fig. 9e, right). The higher production of MMP-1 in the latter case suggests that fibroblasts try to remodel the stiffer collagen substrate by enzymatic degradation, supposedly to reduce matrix stiffness. For other MMPs, a similar behavior has been reported. 57 The goal of the MMP secretion by the cells might be to relax the substrate and introduce deformations by the cells' own forces. However, the crosslinked collagen scaffold is not expected to be degraded significantly in the investigated time frame, as in vivo experiments show almost intact scaffolds 3 weeks after implantation. 58 Constructs Chigh show higher total cell numbers at day 14 as a result of less contraction. At day 2, however, cell numbers were shown to be comparable (Table 2) such that MMP-1 production in stiff scaffolds is higher even if referred to the individual cell.
It has been shown that cell contractility is regulated by cellular fibronectin, 59 presumably in its polymerized form. 60 Further, the network of polymerized fibronectin is a key regulator of the composition and stability of the ECM (as fibronectin polymerization regulates the remodeling of ECM collagen-I) and cell matrix adhesions.61–63 Fibers of polymerized fibronectin are predominantly visible in our immunohistological stainings. This indicates that the fibronectin-rich environment produced by the fibroblasts themselves enhances matrix contraction. The production of a fibronectin matrix and a sustained matrix contraction seem to represent the cell's primary response to tune their local mechanical microenvironment.
If the change in Young's modulus E and the elastic force component Felast is compared between the softer and stiffer scaffold groups Clow and Chigh in Figure 6, it becomes obvious that the cells actively remodel the matrix around them, potentially to achieve a certain “optimal” mechanical environment. The starting conditions are defined by the two groups of scaffolds with different material properties. Over the 2 weeks of culture, however, the mechanical properties seem to converge (Fig. 5) and might finally stabilize at a comparable level for longer culture time. Being a result of excessive scaffold and ECM remodeling, this level seems to be shifted by mechanical loading toward a higher value; while E increases in Clow and is rather constant in Chigh to potentially meet at a level between 4 and 8 kPa in the unloaded group, a stronger increase in Clow and a still visible increase in Chigh indicate a trend toward a level above 8 kPa for the loaded group. Taken together, mechanical stimulation results in a higher mechanical stiffness of cell-seeded constructs as a consequence of enhanced ECM production and remodeling in combination with an increased compaction due to cell contraction.
Obviously there are limitations to our in vitro experiments that keep the interpretation to a certain extend speculative. Large data variations for mechanical testing of constructs Chigh are possibly due to the highly orientated structure of the scaffold. The deformation under compression seems to be variable to a certain extent leading also to variations in compressive force and E. Since cell-seeded constructs show higher variation compared with cell-free scaffolds, the stepwise detachment of the construct from the PEEK mesh discs, caused by lateral contraction, might also contribute to large variations. Soft scaffolds are expected to have a more continuous shape change due to a much stronger lateral contraction. To prove that mechanical properties of Chigh and Clow would equalize, prolonged culturing over more than 2 weeks would be necessary. The compression of the sample for mechanical testing was performed with a constant displacement of 300 μm according to 10% sample height of noncontracted constructs. The reduction of scaffold height over the 2 weeks of culture leads to higher strainings. However, the increase of Fc(t) during compression showed to be highly linear (see Fig. 7) and the resulting error for E was below 5%. As a further point, the type of mechanical stimulation that the cells experience is unknown. It is expected that fluid flow through the scaffold pores (shear force) and the deformation of the scaffold walls contribute to cell straining upon compression. The contribution of both might even change over the culture period as shear forces increase with the blockage of pores due to ECM formation (Fig. 6b). The possibility of an insufficient supply with oxygen and nutrients inside the 3D constructs was not evaluated in this study. However, mechanical stimulation, known to enhance solute transport,64,65 did not lead to a higher cell density at day 14 and no differences were found in glucose consumption and lactate production at day 7 between the loaded and nonloaded groups. Thus, the observed differences in ECM remodeling between the groups were obviously not caused by differences in solute transport and supply. As a further point, applying a wider range of scaffold stiffnesses would allow a more precise identification of the “optimal” mechanical environment. Finally, it is known that besides the mechanical stiffness of the material also the substrate's surface chemistry and topography have an impact on cellular behavior. Thus, the results of this study in collagen-I scaffolds cannot be transferred to other polymer scaffolds without further investigation.
In conclusion, we presented a new bioreactor system that is capable of culturing, mechanical stimulation, and precise mechanical testing of cell-seeded constructs. Compared with existing systems, this allows a deeper insight into how cells remodel their local mechanical environment and how this affects the macroscopical (global) mechanical properties. Our results show that by combining the information gained from the analysis of Young's modulus, relaxation force, and the elastic portion of compressive force (equilibrium state), a much deeper insight into cellular processes involved in tissue maturation and remodeling can be gained than from individual parameters alone. The production of ECM components and the remodeling of the ECM were shown to be altered as a consequence of changed mechanical boundary conditions, namely, by the application of mechanical loading and the change in scaffold stiffness (Fig. 10). Mechanical loading, on the one hand, resulted in an upregulation of ECM production and its impact was preserved over the full experimental time frame. The stiffness of the collagen scaffold, on the other hand, showed to influence ECM remodeling via MMP-1 secretion and ECM compaction (resulting from scaffold contraction). The material's impact, however, was found to be highest in the early phase of remodeling when cells were in direct contact with the scaffold walls. According to our interpretation, mechanical loading rather than initial substrate stiffness is decisive for the cells' mechanical environment at the end of a complex process of scaffold and ECM remodeling. Taking into account that the differentiation of pluripotent cells, like mesenchymal stem cells, has been shown to be sensitively influenced by the substrate stiffness, 10 the shift of mechanical stiffness induced by mechanical loading might influence the fate of tissue regeneration at a very early time point. For bone healing this implies that not only the direct influence of mechanical signals on, for example, fibroblast, MSC, and osteoblast behavior, but also the indirect influence via a change in matrix stiffness might contribute to altered healing.

Illustration of the changes in construct stiffness E over the time of culture caused by cell-induced remodeling processes of scaffold contraction, ECM formation (collagen-I and fibronectin secretion), and ECM degradation (MMP-1 secretion). Only the most relevant processes are shown with bolt text indicating a higher level. The balance of these processes ultimately leads to a specific construct stiffness.
Footnotes
Acknowledgments
This work has been funded by the Berlin-Brandenburg Center for Regenerative Therapies (BCRT) and the Berlin-Brandenburg School for Regenerative Therapies (BSRT). The authors would like to thank the research group of Professor Mundlos at the Max Planck Institute for Molecular Genetics for providing human fibroblasts and Matricel GmbH for the production of Optimaix scaffolds with adapted mechanical properties for this study.
Disclosure Statement
The authors declare that no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.