
Abstract
The utilization of growth factors for bone regeneration is a widely studied field. Since the approval of bone morphogenetic protein-2 (BMP-2) for therapeutic use in humans, the concept of utilizing growth factors for bone regeneration in translational medicine has become even more attractive. Despite many studies published on individual growth factors in various bone models, comparative analysis is largely lacking. The aim of our study was to compare three different proosteogenic factors under identical in vivo conditions. Thus, we tested the bone regeneration capacity of the three different growth factors BMP-2, fibroblast growth factor-2 (FGF-2), and vascular endothelial growth factor A (VEGFA) in a calvarial defect model. We demonstrated that BMP-2 and VEGFA had similar bone healing capacities, resulting in complete calvarial healing as early as week 3. FGF-2 also showed a significantly higher bone regeneration capacity; however, the healing rate was lower than with BMP-2 and VEGFA. Interestingly, these findings were paralleled by an increased angiogenic response upon healing in BMP-2– and VEGFA-treated calvarial defects as compared with FGF-2. Immunohistochemistry for proliferating and osteoprogenitor cells revealed activity at different points after surgery among the groups. In conclusion, we demonstrated an efficient bone regeneration capacity of both BMP-2 and VEGFA, which was superior to FGF-2. Moreover, this study highlights the efficient bone regeneration of VEGFA, which was comparable with BMP-2. These data provide a valuable comparative analysis, which can be used to further optimize growth factor–based strategies in skeletal tissue engineering.
Introduction
The calvarial defect model is a suitable system to study growth factors that augment bone regeneration, since it allows the creation of critical-sized defects and reliable in vivo monitoring of bone healing with micro-computed tomography (μCT) scanning.12,13 Furthermore, the osteogenic response of osteoblasts and dura mater cells can be monitored by immunohistochemistry in vivo. 13 We have previously demonstrated a superior osteogenic healing potential of neural-crest–derived frontal bones as compared with mesoderm-derived parietal bones. 14 We could further demonstrate that FGF ligands −2, −9, and −18 were at least in part responsible for differences in calvarial healing of frontal and parietal bones. 13 Importantly, application of FGF-2, −9, and −18 to parietal bone defects could overcome the inferior healing potential of parietal bones.
In a different independent study we have also demonstrated a proosteogenic and angiogenic effect of VEGFA on human adipose–derived stem cells (hASCs) in vitro and in vivo. 15 Remarkably, by utilizing the calvarial defect model we could see a striking increase in paracrine and cell-autonomous angiogenesis upon treating hASCs with VEGFA. These data suggested that an increase in angiogenesis augments bone formation, since bone formation requires an efficient crosstalk between osteoblasts and endothelial cells. 16
Despite many studies highlighting the osteogenic potential of a certain growth factor, comparative analyses have been largely missing. This hinders a direct comparison of these growth factors, because they were used in different experimental settings and research questions. Therefore, we analyzed the three growth factors BMP-2, FGF-2, and VEGFA in identical experimental conditions to investigate for differences in their bone regeneration capacity.
Materials and Methods
Animal experiments
All animal experiments were performed in accordance with the Stanford University Animal Care and Use Committee Guidelines; 7-day-old CD-1 mice (Charles Rivers Laboratories, Inc.) were utilized for this study. We have previously demonstrated that untreated 2-mm defects in parietal bone of CD-1 mice fulfill the definition of critical-sized calvarial defects (i.e., <10% healing). 13 Each treatment group consisted of at least five animals. Mice underwent a modified calvarial defect procedure as previously described. 13 Briefly, after anesthesia, disinfection and longitudinal incision of the skin was performed. The pericranium was removed and 2-mm calvarial defects were created in the left parietal bone with a trephine drill and meticulous care to avoid damaging of underlying dura mater. Parietal bone defects were treated with a 1.5-mm-diameter collagen sponge (Helistat; Integra Lifesciences Cooperation) soaked in 200 ng recombinant FGF-2 (Santa Cruz Biotechnology), BMP-2, VEGFA (R&D Systems), or phosphate buffered saline (PBS) as a control. The chosen comparative dose of 200 ng of growth factor in the current study was adjusted to what has shown success clinically. Assuming an average body weight of 70 kg of a standard person, the minimal dose in relation to the body weight for successfully treating a patient was 11 ng/g for FGF-2, 17 while it was 171 ng/g for BMP-2. 3 Clinical data for VEGFA treatment were not available. Given the average weight of a p7 mouse (4–5 gram), we decided to define the comparative dose to 200 ng/p7 mouse. The skin incision was then closed and mice were allowed to recover.
μCT imaging
μCT imaging was performed as previously described. 13 Mice underwent μCT scanning at weeks 2, 4, 8, and 12. Microcomputed tomography was performed, using a high-resolution MicroCAT II™ (ImTek, Inc.) small animal imaging system, with previously described settings. Two-dimensional projection images were used to reconstruct tomograms with a commercial software package (Cobra EXXIM; EXXIM Computing Corp.), resulting into a resolution of 80 μm. Three-dimensional reconstructions were generated by MicroView software (GE Healthcare). Each mouse was scanned with a CT-phantom, which was used to calibrate each scan. The precise threshold for regenerating calvarial bone was previously determined as equivalent to 510 Houndsfield Units. The rest-defect area was then determined with the Magic Wand Tool in Photoshop (Adobe). Percentage healing was determined by dividing the rest-defect area by the mean of the defect size one day postoperatively. Mann-Whitney test was used for statistical analyses. A p-value<0.05 was considered statistically significant.
Histology and immunohistochemistry
Mice were harvested 24 h, 48 h, 72 h, 7 days, 3 weeks, and 12 weeks after surgery and processed for histology. Fixation was performed in 10% neutral buffered formalin overnight, followed by decalcification in 19% ethylenediaminetetraacetic acid. Calvaria were then dehydrated, paraffin embedded, and sectioned at 10 μm. For pentachrome staining, standard methods were applied. 14 Bone volume (BV) and BV/tissue volume (BV/TV) analysis were performed as follows. Pentachrome-stained sections were photographed at 10× and evaluated with Photoshop (Adobe). For determination of BV, the selection of pentachrome-stained yellow-positive pixels was partially automated with the magic wand tool (tolerance: 60, no contiguous) within the defect area. TV was determined by selecting the entire defect area with the lasso tool. BV pixels were divided by TV pixels and are presented as percentages. Immunohistochemistry was performed as previously described. 13 Briefly, sections were rehydrated and treated with Proteinase K (Roche Diagnostics) for 15 min. Primary antibodies used were Runx2 (#SC-10758; Santa Cruz Biotechnologies; 1:50) and platelet endothelial cell adhesion molecule 1 (PECAM-1) (#553370; BD-Pharmingen; 1:400). For proliferating cell nuclear antigen (PCNA), immunohistochemistry was performed using a commercially available kit (#93-1143; Invitrogen) according to the manufacturer's instruction. Tartrate resistant acid phosphatase (TRAP) staining was performed with the Leukocyte Acid Phosphatase Kit and Fast Red Violet (both Sigma Aldrich) as previously described. 18 All experiments were performed twice on at least three different mice.
Results
Bone regeneration capacities of BMP-2 and VEGFA are superior to FGF-2
The bone regeneration capacity of different growth factors was assessed using a calvarial healing model. Following surgery, μCT scanning was performed to analyze the healing capacity of FGF-2, BMP-2, and VEGFA over the time lapse of 12 weeks (Fig. 1A). PBS-soaked collagen sponges served as controls. Already at postoperative week 2, high healing rates were observed in BMP-2– (88.9%±12.6%) and VEGFA- (81.0%±23.3%) treated defects, whereas the healing rate of FGF-2–treated defects was lower (61.2%±18.4%; Fig. 1B). Healing in the PBS control group was marginal (1.2%±20.3%). This trend continued to week 12, when healing was found to be 95.1%±8.3% in VEGFA- and 95.2%±9.0% in BMP-2–treated defects. In contrast, the healing rate was lower in FGF-2–treated calvarial defects (72.0%±14.2%). Minor healing was observed in the PBS control group (18.3%±13.1%; Fig. 1B). In a direct comparison between the different groups, the healing capacity of VEGFA-treated defects was significantly better at weeks 4, 8, and 12 as compared with FGF-2. At week 12, differences between BMP-2– and FGF-2–treated defects were likewise significant. No significant differences were found between VEGFA- and BMP-2–treated defects at all time points. Pentachrome staining of defects, harvested 3 and 12 weeks after surgery, confirmed the superior healing capacity of BMP-2 and VEGFA as compared with FGF-2 and PBS control (Fig. 2A). Pentachrome staining of BMP-2– or VEGFA-treated defects revealed complete healing at week 3 after treatment. The thickness of the regenerated bones in the defects was further augmented at week 12. In contrast, FGF-2 treatment showed reduced bone regeneration in the defect compared with BMP-2 and VEGFA. Thus, VEGFA and BMP-2 both had remarkable healing capacities, which were superior to FGF-2. BV measurements at week 3 and 12 revealed significant more BV in the BMP-2– and VEGFA-treated groups as compared with FGF-2 and PBS (Fig. 2B). Moreover there was a trend that BV at week 12 was 25.3% (BMP-2) and 11.1% (VEGFA) increased as compared with the uninjured sides. However when analyzing the bone density (BV/TV), uninjured sides showed significantly higher values (86.3%) as compared with the BMP-2– (67.2%) and VEGFA- (65.4%) treated groups at week 12 (Fig. 2C). This indicated that in both BMP-2– and VEGFA-treated groups, BV exceeded the uninjured side, whereas bone density was higher in uninjured bone. Of note, TRAP staining for osteoclast activity at week 12 did not show significant differences in the regenerated bone between the BMP-2 and vascular endothelial growth factor (VEGF) groups in relation to BV (Supplementary Fig. S1; Supplementary Data are available online at www.liebertonline.com/tea). In contrast, no TRAP staining could be detected in FGF-2–treated group due perhaps to poor tissue regeneration.
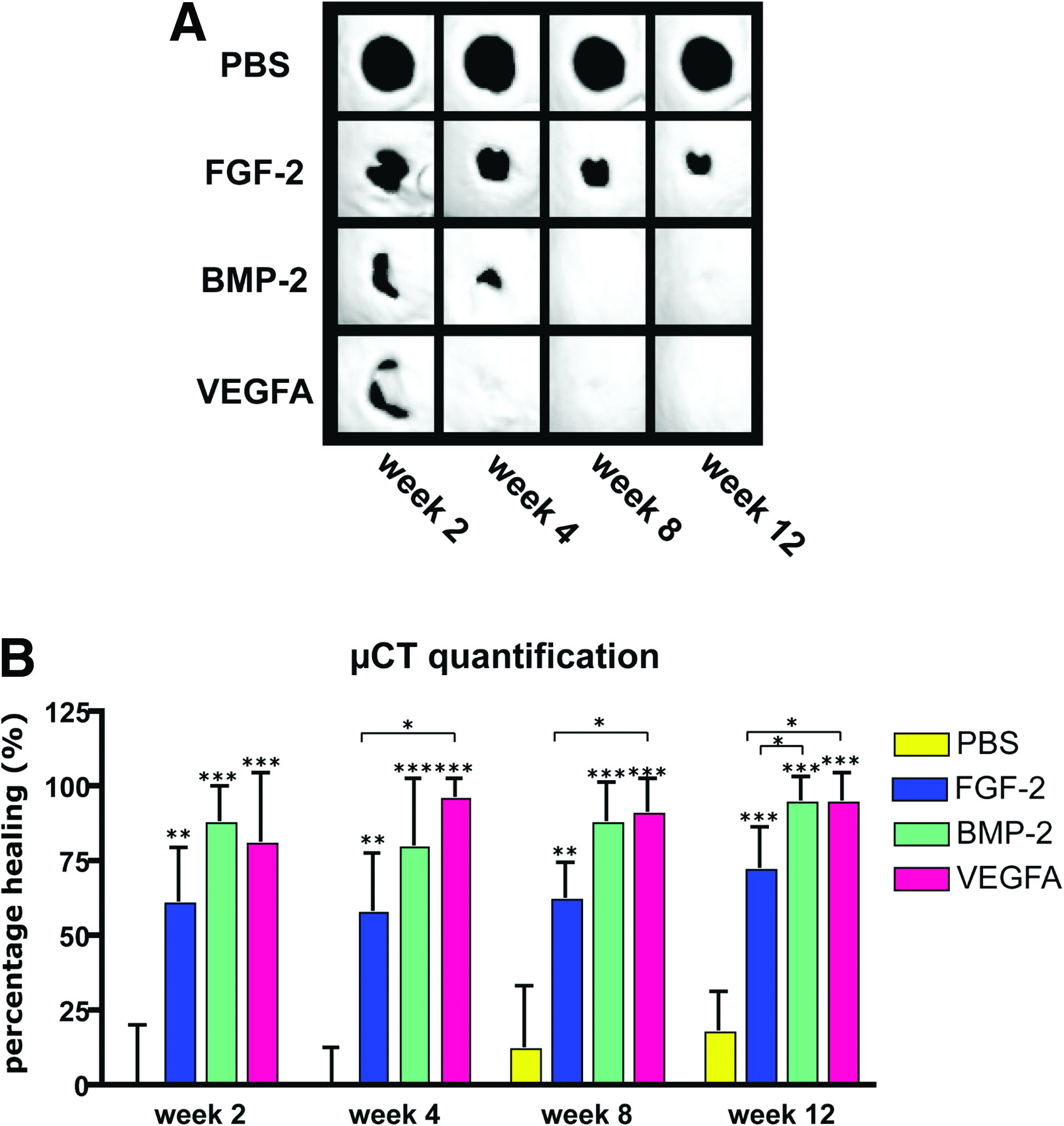
μCT time course analysis of calvarial defects over 12 weeks.
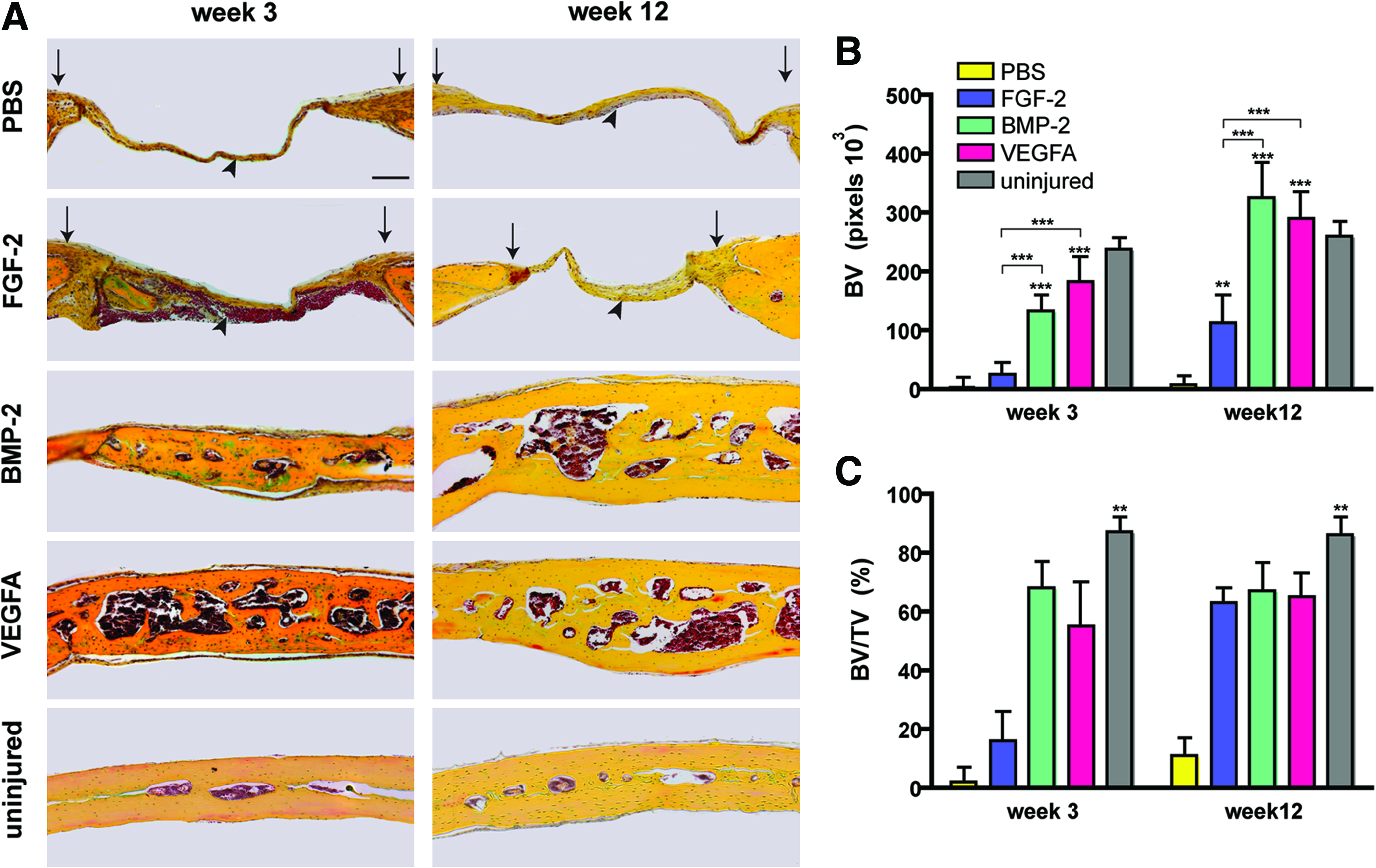
Cell proliferation and osteogenic differentiation
The different healing capacity of BMP-2– and VEGFA-treated calvarial defects as compared with FGF-2 opened the question: Which mechanism could account for this difference? To address this question, we performed immunohistochemistry to assess cell proliferation and osteogenic differentiation in an early postoperative time frame (24 h, 48 h, and 72 h). Cell proliferation was assessed by immunohistochemistry for PCNA (Fig. 3). Most of the proliferating cells were found in the dura mater under the bone defect. The areas presented in Figure 3 show the dura mater and one bony edge of the defect. At first, proliferating cells were noted in the dura mater of VEGFA-treated defects at 24 h. At 48 h, cell proliferation was noted in all treated groups, as well as in the control. This proliferation increased at 72 h in all treated groups, whereas in the PBS control, proliferation was no longer detected. Despite an earlier detection of PCNA-positive cells in VEGFA-treated defects, no substantial differences could be observed among the different treatment groups.
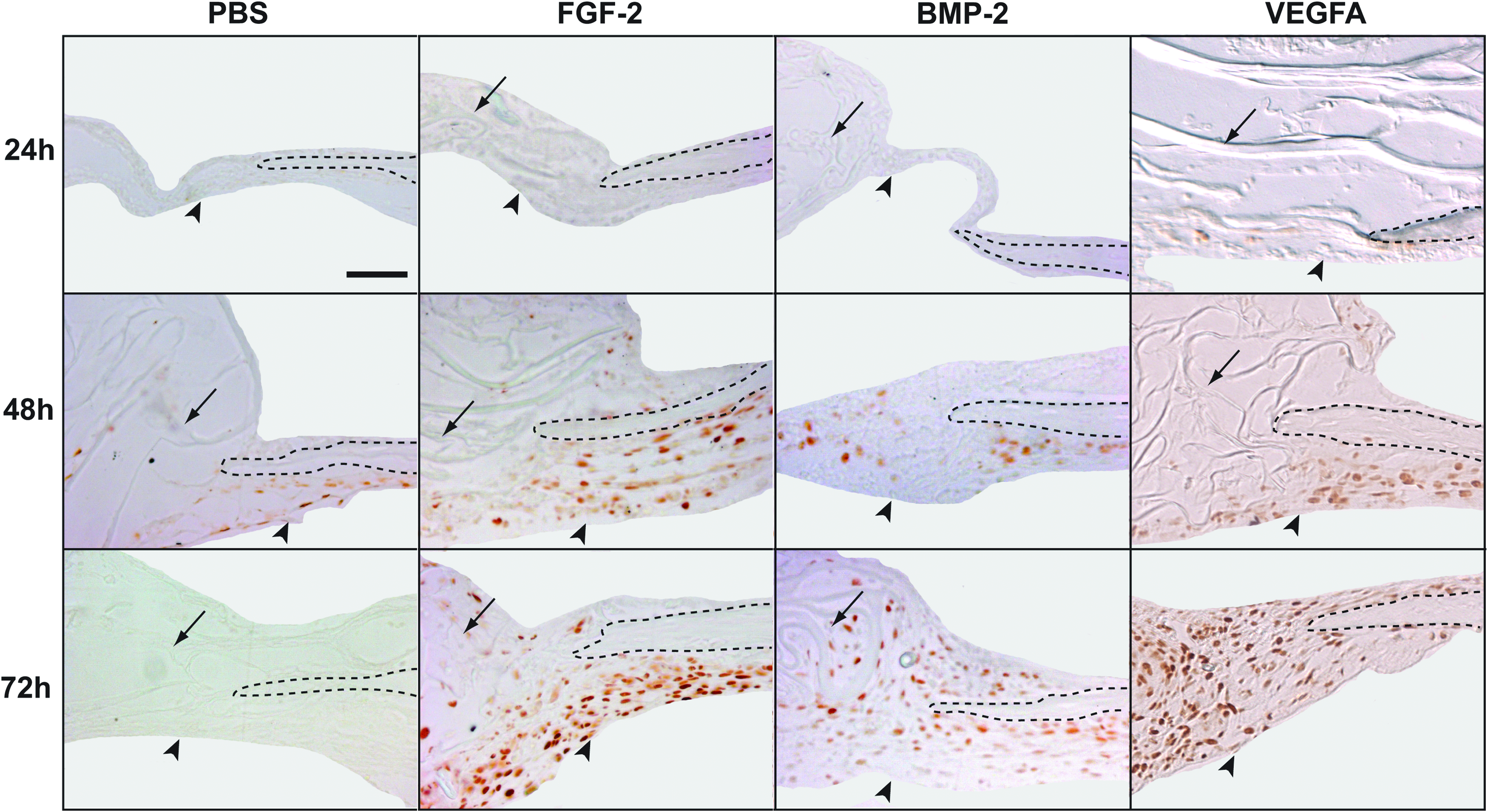
Cell proliferation in calvarial defects at 24 h, 48 h, and 72 h postoperatively. Immunohistochemistry for PCNA revealed PCNA-positive cells in VEGFA-treated defects 24 h after surgery. Similar levels of staining were found between the treatment groups at 48 h; hence staining was also evident in the PBS control group. At 72 h, PCNA-positive cells were detected in all groups, while no staining was observed in PBS control. Arrowheads indicate the dura mater; arrows indicate collagen sponges; dashed lines indicate the bony edges of the defects. Scale bar: 50 μm. PCNA, proliferating cell nuclear antigen. Color images available online at www.liebertonline.com/tea
As a next step, we performed immunohistochemistry for Runx2, a marker for osteoprogenitor cells (Fig. 4). At 24 h, no Runx2-positive cells were found in the PBS control, and few cells stained positive for Runx2 in FGF-2–treated calvarial defects, whereas many Runx2-positive cells were found in BMP-2– and most notably VEGFA-treated defects. At 48 h, Runx2-positive cells were found in all treated groups, while no Runx2-positive cells were found in the PBS control. This trend continued at 72 h. Thus, in BMP-2– and VEGFA-treated defects more Runx2-positive cells were detected at earlier time than FGF-2–treated defects.

Osteoprogenitor cells in calvarial defects at 24 h, 48 h, and 72 h postoperatively. Immunohistochemistry for Runx2, a marker for osteoprogenitor cells, revealed staining in BMP-2– and VEGFA-treated defects at 24 h, while minor staining was found in FGF-2–treated defects. In PBS controls, no staining was observed at any time. At 48 h and 72 h, strong staining was observed in all treated groups. Arrowheads indicate the dura mater; arrows indicate collagen sponges; dashed lines indicate the bony edges of the defects. Scale bar: 50 μm. Color images available online at www.liebertonline.com/tea
Angiogenesis was increased in BMP-2– and VEGFA-treated defects
Immunohistochemistry against PCNA and Runx2 disclosed some clues about the superior healing capacity of BMP-2 and VEGFA. Since all tested cytokines are also known to induce angiogenesis, we analyzed their angiogenic potential in the calvarial healing model. After 7 days, skulls were harvested and immunohistochemistry for PECAM-1 was performed (Fig. 5). Round PECAM-1–positive structures, which indicate vessel formation, were observed in areas of regenerating bone of BMP-2– and VEGFA-treated defects. In contrast, PECAM-1–positive cells were found in FGF-2–treated parietal defects but vessel formation was largely missing. Thus, in this model angiogenesis was observed upon treatment with BMP-2 and VEGFA, but not FGF-2.

Angiogenesis in calvarial defects at postoperative day 7. Immunohistochemistry for PECAM-1 revealed vessel formation in BMP-2– and VEGFA-treated defects. Arrowheads indicate vessels; arrow indicates the collagen sponge; dashed lines indicate regenerating bone in the defects. Scale bar: 50 μm. PECAM-1, platelet endothelial cell adhesion molecule 1. Color images available online at www.liebertonline.com/tea
Discussion
The current comparative analysis indicated significant increases in bone regeneration of parietal calvarial defects by treatment with BMP-2, FGF-2, and VEGFA. The most efficient healing of calvarial defects was obtained with BMP-2 and VEGFA, which was superior to treatment with FGF-2. These differences correlated with an earlier presence of osteoprogenitors and an increase in angiogenesis of BMP-2– and VEGFA-treated defects.
Many studies have demonstrated the potential to induce bone formation by BMP-2 through various carriers in calvarial defects.19–24 Thus, we included BMP-2 as a reference in our study and confirmed robust bone formation. Similar to the prolonged augmentation of bone formation in BMP-2–treated defects from 3 to 12 weeks observed in our study, Lee and colleagues also reported an advancement of bone formation from 2 to 8 weeks upon treatment of calvarial defects with BMP-2–loaded collagen sponges. 19 Interestingly, BMP-2 itself has the potential to induce angiogenesis.25–27 Our data showing that BMP-2 can cause angiogenic effects in a calvarial defect model are supported by a recent report investigating the effects of BMP-2 in nonstabilized fractures, 28 where BMP-2 affected angiogenesis in the hard callus phase. Moreover, BMP-2 was shown to induce an activation of endothelial progenitor cell chemotaxis. 29 In contrast, Cowan and colleagues reported no difference in the number of blood vessels between rat calvarial defects treated with poly(lactic-co-glycolic acid) scaffolds and BMP-2 and controls 12 weeks after surgery. 22 This discrepancy can be explained by a different species and carrier material and most importantly by a considerably earlier time point (7 days) to analyze differences in angiogenesis in our study.
Given the expected robust bone regeneration of BMP-2, we were surprised about the similar remarkable bone regeneration capacity of VEGFA. This effect was accompanied by angiogenesis. The beneficial effects of recombinant VEGFA on bone healing have been previously studied with bioerodible polylactic acid depot scaffolds. 5 Similarly, VEGFA, delivered by gene-activated matrices in radial critical-sized defects of rabbits, led to an increase of vascularization and bone regeneration. 30 In our current study, we provide all growth factors on collagen sponges, which represent a potential translational scenario, since BMP-2 delivery on collagen sponges is already FDA approved. This approach is supported by a previous study, which revealed that application of VEGFA-soaked collagen sponges in rabbit calvarial defects increased angiogenesis and bone formation. 31 However the authors did not attempt to test these effects in a critical-sized defect, to evaluate the remarkable healing capacity of this growth factor. It is worth mentioning that VEGF acts synergistically with BMP-2 and BMP-4.32,33 Bone formation was inhibited in BMP-2–expressing muscle-derived stem cells under treatment with the VEGF antagonist s-Flt1, suggesting that angiogenesis is important in BMP-2–induced bone formation. 33 Conversely, exogenous VEGF enhanced BMP-2–induced bone formation, presumably by promoting angiogenesis. 33 An additional study of the same group examining the dose-ratio of VEGF and BMP-4 revealed that high concentrations of VEGF lead to less mineralization potential. 34 Congruent with our results it might be possible to reduce the dose of BMP-2, when combined with VEGFA.
Application of BMP-2 and VEGFA did result in closure of the critical-sized defects as observed with μCT analysis and histology. Though, when examining the morphology of the regenerated bone, it was evident that BMP-2– and VEGFA-treated defects showed a trend to higher BV as compared with the contralateral uninjured bone, while the bone density was significantly lower. Calvaria belong to nonweight-bearing bones, which might explain the lower density in the regenerated bones. Certainly, it has to be stated that more bone does not necessarily equal quality and functional bone. Thus, further biomechanical studies are needed to faithfully answer these questions.
Interactions between the tested growth factors BMP-2, FGF-2, and VEGF in bone biology have been attributed to their signaling pathways. For instance, it has been previously reported that FGF-2 stimulates VEGF release in osteoblast-like MC3T3-E1 cells through the p42/44 MAP kinase pathway, whereas activation of the p38 mitrogen-activated protein (MAP) kinase pathway blocks VEGF release. 35 An additional pathway important for FGF-2–induced VEGF release described subsequently is stress-activated protein kinase/c-Jun NH2-terminal kinase (SAPK/JNK). 36 An explanation for the inferior bone regeneration capacity of FGF-2 as opposed to VEGFA in our model could originate from the fact that FGF-2 is a highly basic protein and release is less efficient than for other growth factors.
Moreover, it has been proposed that low doses of FGF-2 enhance BMP-2–induced ectopic bone formation in mice. 37 Interestingly, Bmpr1-B receptor expression was increased 10–30-folds upon treatment with low-dose FGF-2. The authors did not investigate the impact of specific FGF pathways; however, it is known that high doses of the Erk1/2 pathway negatively regulate BMP-2 stimulation of alkaline phosphatase, osteopontin, and Msx2 expression in mesenchymal stem cells, probably because SMAD levels are decreased. 38 In contrast, PI3-K signaling positively regulates BMP-2–induced alkaline phosphatase and osteopontin expression. 38 Although the BMP-2–induced SMAD signaling pathway is considered to be the main pathway for regulating Runx2 expression levels and stability, the BMP-2–activated Erk/MAP kinase pathway also increases Runx2 stabilization and transcriptional activity. 39
Several groups, including our own, have previously described the close relationship between angiogenesis and osteogenesis in bone regeneration.15,40–43 Localization of vessels surrounded by regenerated bone in BMP-2– and VEGFA-treated defects further supports the importance of a crosstalk between osteoblasts and endothelial cells during bone formation. 16 Thus, the relevance of a concurrence of angiogenesis and osteogenesis to allow sufficient bone regeneration may be further underlined by the correlation of increased angiogenesis and bone regeneration in BMP-2– and VEGFA-treated defects. In our direct in vivo comparison, both the angiogenic potential and bone regeneration capacity were inferior in FGF-2–treated calvarial defects as compared with BMP-2 and VEGFA. A question that remains is the origin of the cell source for angiogenesis in the calvarial defect model. A potential candidate could be the dura mater, which has been previously reported to display angiogenic activity when exposed to hypertension. 44 However, circulating vascular progenitor cells that are attracted to the skeletal injury area could also be involved.
It has been previously described that the factors BMP-2, VEGFA, and FGF-2 tested in the current study displayed chemoattractive effects on osteoprogenitor cells.13,45,46 Of note, that all tested growth factors exhibited effects on proliferation and osteogenic differentiation on dura mater and pericranium cells, similar to what we have previously observed upon treatment of parietal defects with FGF-2, −9, and −18. 13 In the light of the results presented herein, recruitment of dura mater and pericranial cells was also achieved by BMP-2 and VEGFA, suggesting that this observation is independent of the proosteogenic factor under study and may therefore be a general principle in the calvarial defect model. It has to be stated though, that the strong effects of FGF-2 on proliferation and osteogenic differentiation were not sufficient to gain the bone healing capacity observed with BMP-2 and VEGFA at weeks 3 and 12. Thus, it is possible that important steps such as induction of angiogenesis and maturation of osteoprogenitor cells are not elicited by the application of FGF-2.
Despite the tremendous amount of studies focusing on single growth factor or combinations of growth factors for bone regeneration, comparative analysis is largely lacking. Nonetheless in one study, glass ceramic scaffolds loaded with BMP-2, FGF-2, and VEGFA have been implanted in sheep tibia bone defects 47 ; however, the doses were not kept consistent among treatment groups. Four weeks postoperatively, BMP-2–loaded scaffolds exhibited an increase in bone formation, whereas these effects were not found for FGF-2 and VEGFA.
In conclusion, the growth factors BMP-2 and VEGFA showed similar efficient bone regeneration rates, which were superior to the regenerative capacity of FGF-2. Based on a side-by-side comparison, differences in proliferation, recruitment of osteoprogenitor cells, and angiogenesis and bone regeneration rates revealed distinct profiles for the three growth factors. Finally, the work presented herein will aid in selecting candidate growth factors for translational approaches and suggests the potential for VEGFA in bone tissue engineering as an alternative to BMP-2 treatment.
Footnotes
Acknowledgments
This work was supported by the Oak Foundation, the Hagey Laboratory for Pediatric Regenerative Medicine, and NIH R21DE019274 and NIH R01DE019434 to M.T.L. and the German Research Foundation (DFG BE 4169-1) to B.B.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.