
Abstract
This study investigated the hypothesis that timing and duration of dynamic compression are integral to regulating extracellular matrix (ECM) assembly of tissue-engineered (TE) menisci. The goal of this study was to examine the effects of varying load and static culture duration on structure, composition, and mechanical properties of TE menisci. We accomplished this by varying the duration of dynamic loading over 4 weeks of culture, and by examining increasing periods of static culture after 2 weeks of dynamic loading. Bovine meniscal fibrochondrocytes were seeded into 2% w/v alginate, crosslinked with CaSO4, injected into anatomical micro-computed tomography-based molds, and post-crosslinked with CaCl2. Meniscal constructs were dynamically compressed three times a week via a custom bioreactor for a total of 2 h, with an hour of rest between loading cycles, for 1, 2, or 4 weeks. They were then placed in static culture. After 4 weeks of culture, increased load duration was found to be beneficial to matrix formation and mechanical properties, with superior mechanical and biochemical properties in samples loaded for 2 or 4 weeks. Further, the mechanical properties of these constructs were similar, suggesting that the additional 2 weeks of loading may not be necessary. Samples loaded for 2 weeks followed by a 4-week static culture period yielded the most mature matrix with significant improvements in collagen bundle formation, 2.8-fold increase in the glycosaminoglycan content, 2-fold increase in the collagen content, and 4.3-fold increase in the compressive equilibrium modulus. Overall, this study demonstrated the importance of timing and duration of loading. By switching to prolonged static culture after 2 weeks of loading, we decreased the amount of ECM lost to the media, while significantly increasing biochemical and mechanical properties of TE menisci.
Introduction
Due to the challenges associated with allograft transplantation, the use of tissue engineering to develop a meniscal construct is of much interest. A tissue-engineered (TE) meniscus should be geometrically accurate to ensure the appropriate distribution of pressures across the joint, it must be able to withstand anatomical compressive and tensile loading conditions before implantation, and needs to achieve the heterogeneous mechanical and biochemical properties found in native menisci.2,7,8 For many years, research has concentrated on treatment of focal defects and extracellular matrix (ECM) production in plugs. Recently, several studies have attempted to create a whole TE meniscal replacement, whether anatomical or not, made from either synthetic or natural materials.9–21 None of these efforts have achieved a clinical use, as many lack the mechanical properties necessary to withstand implantation in vivo. Thus, mechanical conditioning may be necessary for engineered constructs before implantation.
Mechanical conditioning is widely used to enhance the in vitro formation of various types of TE cartilage, often resulting in increased mechanical and biochemical properties. Several studies have utilized dynamic compression to characterize the mechanical and biochemical properties of meniscal explants.22–32 Many of these studies22–25,30–32 and a few TE studies12,14,33 established that mechanical conditioning enhances ECM production and mechanical properties of scaffolds seeded with meniscal fibrochondrocytes.
Recently, we reported that dynamic compression of anatomically shaped TE alginate menisci significantly enhanced ECM production and mechanical properties after 2 weeks of loading. 14 However, prolonged loading subsequently decreased mechanical performance and matrix retention due to either increased alginate degradation 12 or induced catabolic cellular responses.28,32,34–36
These data are consistent with several other recent observations that prolonged mechanical stimulation is not optimal for engineered tissues. Several studies of meniscal tissue explants28,32 and articular chondrocytes seeded in poly(ethylene glycol) (PEG), 36 agarose, 35 and peptide hydrogels 34 have demonstrated that moderate dynamic strains of 5%–20% over extended periods (12–48 h) results in inhibited cell proliferation and proteoglycan synthesis, increased breakdown of proteoglycans, and increased expression of matrix metalloproteinases (MMP-1, -3, -9, and -13) and aggrecanases. Collectively, these studies demonstrate that prolonged dynamic compression induces a cellular catabolic response and may not be optimal for engineered tissues. While the catabolic response may be important to initiate proteoglycan turnover and stimulate matrix remodeling, sustained expression of the catabolic genes can lead to tissue degradation. 34 This may explain the decrease in mechanical and biochemical properties with prolonged loading of TE alginate menisci. 14
Nicodemus et al. 37 recently demonstrated that intermittent dynamic compression at 5%–20% strain over 1 week significantly increases expression of MMP-1, -3, and -13 and collagen II in articular chondrocytes encapsulated in PEG hydrogels. However, MMP levels dropped significantly once dynamic compression was removed. Furthermore, at 7 days postloading, MMP expression levels were lower than before loading was applied, while collagen and glycosaminoglycan (GAG) levels continued to increase. 37 This study is an excellent example of how the duration and timing of dynamic compression affect the catabolic and anabolic responses of TE articular cartilage; however, such effects have never been documented in scaffolds with meniscal fibrochondrocytes. We hypothesize that the timing and duration of loading (i.e., duty cycle) regulates ECM assembly of TE menisci. The specific goal of this study was to examine the effects of varying the duration of loading and postloading static culture duration on the structure, composition, and mechanical properties of seeded anatomical alginate menisci. We accomplished this by (I) varying the duration of dynamic loading over 4 weeks of culture and (II) altering the period of static culture after 2 weeks of dynamic loading.
Methods
Injection molding
Meniscal constructs were generated as previously described.11,38,39 Briefly, bovine menisci were isolated from freshly slaughtered 1–3-day-old calves and diced into 1-mm3 cubes. The tissue was digested overnight in 0.3% collagenase, 100 μg/mL penicillin, and 100 μg/mL streptomycin in Dulbecco's modified Eagle's medium (DMEM). The bovine meniscal fibrochondrocytes were seeded into sterile 2% w/v low-viscosity high G-content alginate at 50×106 cells/mL. The alginate-cell suspension was combined with 0.02 g/mL CaSO4 at a 2:1 ratio and injected into acrylonitrile butadiene styrene plastic molds of ovine menisci, designed as previously described from micro-computed tomography (μCT) scans.12,38 Molds were then allowed to further crosslink for 40 min in 60 mM CaCl2 before removal of the construct.
A total of 44 constructs were created and incubated in DMEM with 10% fetal bovine serum, 100 U/mL penicillin, 100 μg/mL streptomycin, 0.1 mM nonessential amino acids, 50 μg/mL ascorbate, and 0.4 mM
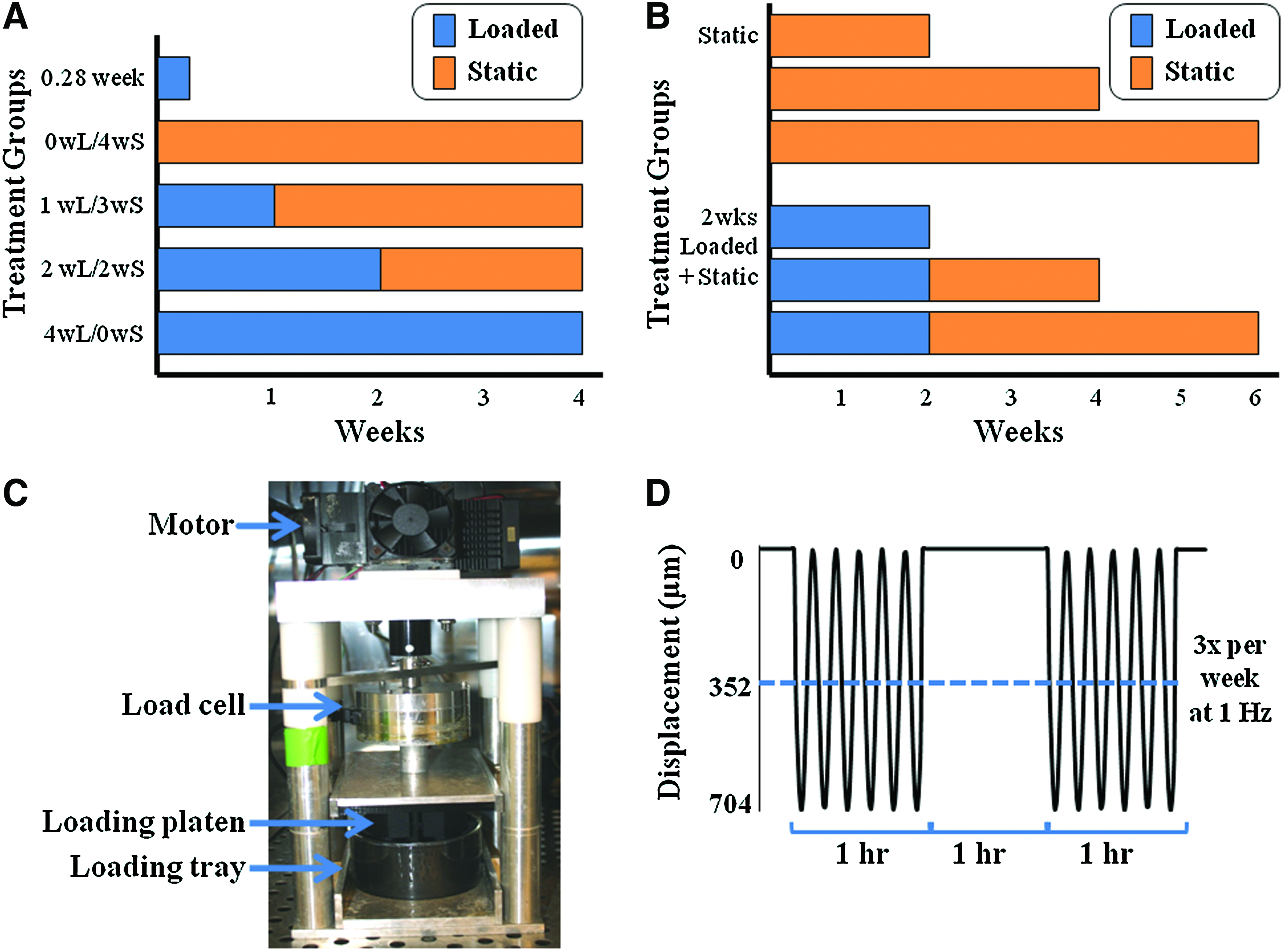
Depiction of experimental setup for part I
Dynamic loading
Dynamic compression was applied with a custom in-house-designed bioreactor outfitted with a loading platen and tray designed to match construct geometry and apply uniform deformation as previously described (Fig. 1C). 14 Briefly, the loading platen and tray were designed based on the same micro-CT scans used to design the menisci molds, thus allowing the loading platen to match surface geometry of the meniscus and the loading tray to have an impression that restricts motion of the construct under loading. 14
Loaded meniscal constructs were exposed to dynamic compression for an hour twice every other day with an hour of rest in between loading cycles. The constructs were exposed to a compressive sinusoidal displacement at 1 Hz with a 352 μm offset and 352 μm amplitude, resulting in a maximal displacement of 704 μm as previously described 14 (Fig. 1D). This loading condition was chosen due to work that identified it as the ideal loading frequency and most physiologically relevant loading regime to mimic the native meniscal environment.32,34,40 The linear poroelastic finite element analysis confirmed that this loading regime applies a compressive strain of 15% to the majority of the concave surface of the meniscus, 14 which is within the range of strain values used in other dynamic compressive studies.32,34,36
Postculture construct analysis
Upon removal from culture, meniscal constructs were photographed and visually inspected for changes in opacity and shape fidelity. Constructs were then processed for histology, biochemistry, and mechanical analyses as previously described.11,12,38 Briefly, cross sections of constructs were cut and fixed in 10% buffered formalin with 1 mM CaCl2 to prevent gel solubilization. Fixed sections were then sectioned and stained with Safranin-O and Picrosirius red to observe GAG and collagen localization, respectively. Twenty representative images of each treatment group stained with Picrosirius red were taken at 400× brightfield for image analysis of collagen bundle formation. Images were analyzed with a custom MATLAB program, which isolated collagen bundles based on color, size, and shape, and provided a collagen bundle count and area for each image. Data were collected and analyzed to determine the percentage of fibers per sample (measurement of the fraction of selected area occupied by fibers in each sample), the average number of collagen bundles per mm2, and the average size of collagen bundles.
The remainder of each meniscal construct had 4-mm-diameter by 2-mm-thick plugs cut from the face, center, and bottom of samples. Excess surrounding tissue from each spatial location was used for biochemical analysis, while plugs were used for mechanical analysis. A total of six plugs and six biochemical samples were obtained from each meniscal construct.
Biochemical samples were weighed to obtain the wet weight (WW), and the samples were frozen, lyophilized, and weighed again to obtain the dry weight (DW). The samples were then digested in 1.25 mg/mL papain solution overnight at 60°C and analyzed for DNA, GAG, and collagen content via a Hoechst dye assay, 41 a modified dimethylmethylene blue dye assay at pH 1.5, 42 and a hydroxyproline assay, 43 respectively. Media samples, obtained when media were changed three times a week, were analyzed biochemically for DNA, GAG, and collagen content using the same assays. Retention percentages for biochemical properties were calculated as the fraction of total accumulation within the scaffolds to the sum of accumulation within the scaffold plus accumulated lost to the media.
Plugs were tested in confined compression to determine the equilibrium modulus [Electroforce (ELF) 3200 System; EnduraTech, Minnetonka, MN] as previously described.39,44 Briefly, stress–relaxation tests were performed by imposing 10×50-μm steps on the plug with the resultant loads fit to a poroelastic model using a custom MATLAB program to calculate the equilibrium modulus.
Statistics
All data were analyzed by two-way analysis of variance using the Tukey's t-test for post-hoc analysis and p<0.05 as a threshold of statistical significance. All statistical analyses were performed using Sigmastat version 3.0 and all data expressed as mean±standard deviation.
Results
Meniscal construct shape fidelity and composition
Gross inspection of engineered constructs demonstrated that shape fidelity was maintained in all treatment groups for the entire length of culture (Figs. 2 and 3). Also, there was a noticeable increase in opacity with loading and time in culture, which suggests an increase in matrix accumulation (Fig. 2).

Photographs of part I engineered constructs after removal from culture (column 1), tissue sections stained with Safranin-O at 200×(column 2), and Picrosirius red at 400×under brightfield (column 3). Scale bars for constructs=5 mm and for stained sections=100 μm, arrows point to collagen bundles. Color images available online at www.liebertonline.com/tea

Photographs of part II engineered constructs after removal from culture (column 1 and 4), tissue sections stained with Safranin-O at 200× (column 2 and 5), and Picrosirius red at 400× under brightfield (column 3 and 6). Scale bars for constructs=5 mm, and for stained sections=100 μm; arrows point to collagen bundles. Color images available online at www.liebertonline.com/tea
Overall, histology for both parts of the experiment demonstrated an increase in tissue development with time in culture, with increased development in loaded samples. In part I, Safranin-O and Picrosirius red staining demonstrated increased GAG localization and collagen bundles in the 2wL/2wS and 4wL/0wS treatment groups, respectively, with a noticeable change to a more heterogeneous morphology in the 2wL/2wS and 4wL/0wS groups, similar to that of native menisci (Fig. 2; columns 2 and 3). In part II, Safranin-O staining exhibited increased GAG localization in static and loaded samples with time in culture, along with a more heterogeneous composition in loaded samples (Fig. 3; columns 2 and 5). Picrosirius red staining demonstrated increased collagen content with time in culture for static and loaded samples, with more prominent localization in the 2-week loaded samples. Collagen bundle formation was evident starting at week 2 and increased through week 6 in loaded samples, while only a few collagen bundles formed in static samples by week 6 (Fig. 3; columns 3 and 6).
Image analysis of Picrosirius red staining demonstrated that the percentage of fiber content, number of bundles per mm2, and average size of collagen bundles significantly increased with 2 weeks of loading (Fig. 4). Collagen bundles were present after just 1 day of loading as evident in the 0.28-week samples. In part I, the 2wL/2wS and 4wL/0wS samples had a >2.5-fold increase in fiber content (p<0.001), a 3-fold increase in number of collagen bundles (p<0.001), and a 2-fold increase in average size of individual collagen bundles (p<0.05) over all other samples (Fig. 4A–C). In part II, the percent fiber content and number of collagen bundles increased 13- and 9.5-fold, respectively, after just 2 weeks of loading (p<0.02). Loaded samples continued to significantly increase with prolonged static culture, with 6-week samples significantly greater than all other time points for loaded and static samples and threefold higher than 6-week static samples (p<0.001) (Fig. 4D, E). The average individual bundle area significantly increased with loading, and appeared to plateau for both static and loaded samples by week 6 (Fig. 4F). Notably, fiber sizes observed at early times in culture were quite large, ∼200 μm2 in area or ∼16 μm in diameter. This size may represent some critical nucleation size for fibers in this system, or may be a technical limitation of this technique due to the inability to accurately distinguish between extracellular and intracellular fibers.

Collagen bundle content determined by image analysis of 20 representative images of each treatment group stained with Picrosirius red. Percent fiber content of constructs, number of collagen bundles per mm2, and average area of individual collagen bundles for parts I
Biochemical properties
Biochemical analysis demonstrated an increase in GAG content with time in culture, with no effect or a decrease in GAG content for loaded constructs. In part I, where duration of loading was varied over 4 weeks of culture, GAG content increased with time for all groups, with the 4wL/0wS and 0wL/4wS treatment groups having significantly higher GAG concentration than the 1wL/3wS and 2wL/2wS groups (p<0.02) (Fig. 5A). In part II, where the period of static culture duration was varied after 2 weeks of dynamic loading, GAG content for static culture constructs significantly increased at 2 weeks by twofold, and plateaued for the remainder of culture. In contrast, constructs loaded 2 weeks and then static cultured had significantly lower GAG content than the static samples until week 6 when the GAG concentration increased 2.8-fold and became significantly greater than static constructs (p<0.001) and all other time points for loaded samples (p<0.02) (Fig. 5C).
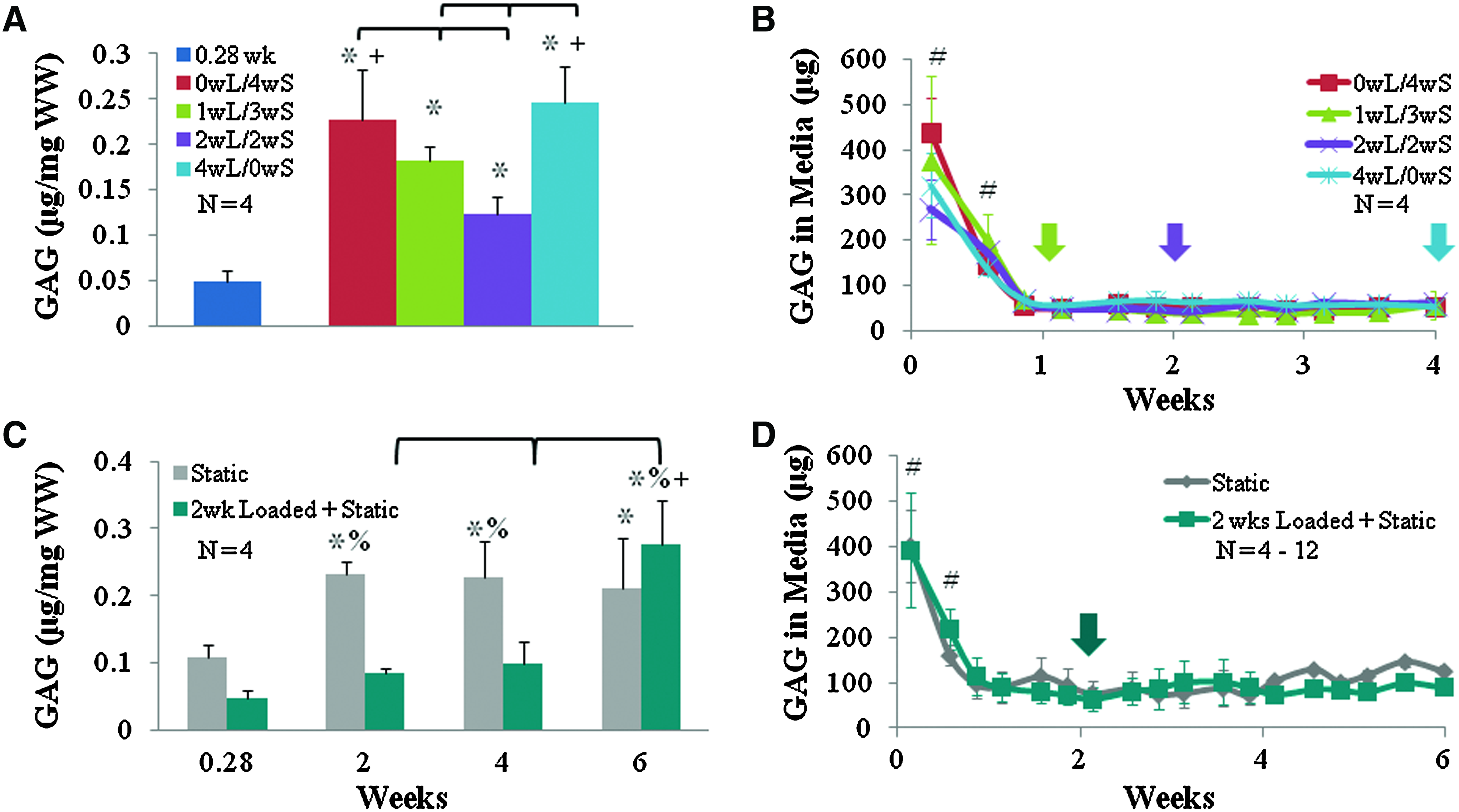
GAG content found in meniscal constructs
All treatment groups had an initial large loss of GAG to the media, which decreased to a significantly lower level throughout the remainder of culture (p<0.001) (Fig. 5B, D). Treatment had no apparent effect on release of GAG to the media. In part I, GAG retention remained relatively similar for 0wL/4wS, 1wL/3wS, and 4wL/0wS with retentions of 39%, 30%, and 36%, respectively, while 2wL/2ws had 20% retention. In part II, static samples began with a high 38% retention at 2 weeks, which decreased by week 6 to 15%. Samples loaded 2 weeks and then static cultured maintained 20% retention of GAG throughout culture.
Prolonged loading significantly increased the collagen content. In part I, collagen content was significantly higher in 2wL/2wS and 4wL/0wS groups than in all other treatment groups (p<0.001), while the 4wL/0wS group had a significantly higher concentration of collagen than the 2wL/2wS group (Fig. 6A). In part II, samples loaded for 2 weeks had significantly higher collagen content at 2, 4, and 6 weeks than the respective static and 0.28-week construct. By week 6, the 2-week loaded samples had a twofold increase in collagen content, and the collagen content was significantly higher than all other time points (p<0.002) (Fig. 6C).

Collagen content found in meniscal constructs
Collagen released to the media was abolished in part I and significantly reduced in part II for the week after removal of load, after which collagen loss returned to previous levels for the duration of culture (Fig. 6B, D). In part I, collagen retention increased with prolonged loading with 2wL/2wS and 4wL/0wS having 8% retention, and 0wL/4wS and 1wL/3wS having 5% retention. In part II, collagen retention decreased with time in culture with static and loaded samples having retentions of 46% and 27% at 2 weeks and 11% and 10% at 6 weeks.
Despite increased ECM content, DNA content demonstrated a significant drop after 2 days for all treatment groups in both part I and II (p<0.001; data not shown). Similarly, DNA released to the media demonstrated a large amount of DNA in the media over the first few days, which decreased and leveled off with time to a very low level (below 20 μg/day) in the media for all treatment groups (data not shown).
There was no consistent difference seen spatially for DNA, GAG, and collagen content in all treatment groups (data not shown).
Equilibrium modulus
The equilibrium modulus significantly increased with prolonged loading. In part I, the equilibrium modulus was significantly higher for 2wL/2wS and 4wL/0wS groups with a 4.3- and 3.4-fold increase over 0.28-week controls, respectively (p<0.012) (Fig. 7A). In part II, the equilibrium modulus for samples loaded 2 weeks and then static cultured significantly increased 2.0- to 2.8-fold over the static and 0.28-week controls by 2 weeks of culture and further significantly increased by 4 weeks of culture for a final 4.3-fold increase over 0.28-week controls (Fig. 7B). Loaded samples were significantly greater than static samples at 2, 4, and 6 weeks (p<0.05).
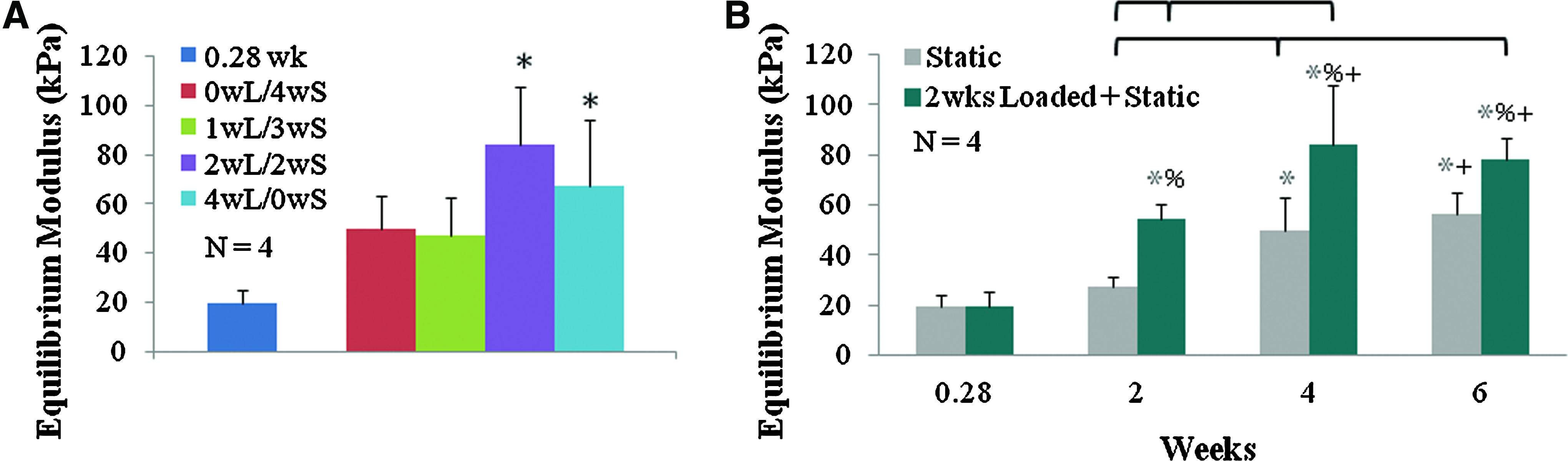
Compressive equilibrium modulus from part I
There were no consistent significant spatial differences in the equilibrium modulus; however, the bottom of the construct did trend toward being higher than the face and center of the constructs in samples loaded for 2 weeks and then static cultured (p<0.1; data not shown).
Discussion
This study investigated the hypothesis that altering the duration of loading and total culture time is key to regulation of ECM assembly. This was achieved in two parts: (I) the duration of dynamic loading was varied over 4 weeks of culture; (II) the period of static culture after 2 weeks of dynamic loading was varied. In part I, we demonstrated that the effect of dynamic stimulation on TE menisci was dependent on load duration. In general, increased load duration was beneficial to matrix formation, collagen accumulation, and mechanical properties, with 2wL/2wS and 4wL/0wS superior to static and 1wL/3wS. Loading decreased or had no effect on GAG accumulation. Further, the mechanical properties of 2wL/2wS and 4wL/0wS were similar, suggesting that the additional 2 weeks of loading may not be necessary. In part II, we demonstrated that static culture after 2 weeks of loading improves ECM properties, with further significant increases in GAG, collagen content, formation of collagen bundles, and equilibrium modulus by 6 weeks of culture. Overall, this study demonstrated that we were able to maintain and improve biochemical and mechanical properties with static culture after 2 weeks of loading.
In this study, meniscal fibrochondrocytes seeded in alginate responded to dynamic compression by differentially regulating collagen and GAG accumulation, which is contrary to how TE articular cartilage typically responds. We demonstrated that meniscal fibrochondrocytes seeded in alginate respond immediately to load by increasing collagen concentrations after 2 weeks of loading, while decreasing or not affecting GAG concentrations. GAG concentrations only increased with time in culture and/or accumulation of collagen content. Further, removal of loading demonstrated a significant change in release of collagen to the media, but had no effect on the release of GAG. In contrast, TE cartilage exposed to dynamic compression has been repeatedly shown to have increased GAG and collagen accumulation, often with a greater accumulation of GAG than collagen.45–49 This response of TE menisci and articular cartilage to dynamic compression is further supported by a study that has observed a differential response of loaded explants of articular cartilage and meniscus tissue, 22 and by another study that has observed articular chondrocytes and meniscal fibrochondrocytes to synthesis and accumulate aggrecan differently. 50 Further, meniscal fibrochondrocytes may differentially regulate collagen and GAG based on the composition of the native tissue. GAG concentration in fibrocartilage is eightfold less than that found in articular cartilage, 1 thus demonstrating the importance of collagen in fibrocartilage.
However, the lower concentration of GAG seen in this study could be due to other factors besides cellular response to loading. Multiple studies have demonstrated mechanical stimulation of cell-seed scaffolds to result in loss of GAG to the media and/or no effect on GAG accumulation.14,37,51,52 We have seen this previously with significant increases in release of GAG to the media with prolonged loading 14 and in this study with a 18% and 38% retention in 2-week loaded and static samples, respectively. It is believed that a collagen matrix must be present to maintain GAG within the scaffold; otherwise, GAG could be easily washed out of the sample by mechanical loading.51,52 This could also explain why at 6 weeks in our 2-week loaded and then static cultured samples we see significantly increased GAG accumulation coinciding with significantly increased collagen accumulation.
Additionally, in this study, we observed significant increases in collagen content and collagen bundle formation coincided with significant increases in the compressive modulus. This suggests that collagen has a more direct relationship to the compressive equilibrium modulus in TE fibrocartilage, contrary to the relationship between GAG and the compressive modulus often seen in TE articular cartilage.49,51,53 This relationship between collagen and the compressive modulus could be due to increased organization attributed to collagen fiber accumulation and not necessarily the properties of collagen.51,53 Once GAG accumulation increases, its contribution to the compressive equilibrium modulus may increase.
Previously, we reported that dynamic compression of anatomically shaped TE alginate menisci significantly enhanced ECM production and mechanical properties after only 2 weeks of loading. 14 However, we also reported that prolonged loading subsequently decreased mechanical performance and matrix retention. We believe that this decrease in properties could be caused by either a degradation of alginate 12 or a cellular catabolic response induced by prolonged loading resulting in increased expression of MMPs and aggrecanases, and increased loss of ECM to the media.28,32,34–36 In this experiment, loading for 2 weeks and then switching to prolonged static culture eliminated the decrease in mechanical and biochemical properties we previously saw with prolonged loading and resulted in significantly increased mechanical and biochemical properties. Further, we saw no change in release of DNA, GAG, or collagen content in the media, which we previously demonstrated to significantly increase with prolonged loading. 14 By eliminating loading after 2 weeks, we are able to decrease the amount of ECM lost to the media, while still inducing significant increase in ECM accumulation and mechanical properties. Removing load could be slowing alginate degradation, or it may be downregulating catabolic activities often induced by mechanical loading.
It has been increasingly suggested that catabolic activity induced by mechanical loading is necessary to initiate proteoglycan turnover and remodeling for a biomechanically functional tissue.34,37,54,55 Further, it has been demonstrated that catabolic activity is downregulated, while anabolic activity continues when mechanical stimulation is removed.37,55 Recently, De Croos et al. 55 demonstrated that a single application of dynamic compression for 30 min to chondrocytes in calcium polyphosphate substrates results in upregulation of MMPs within 2 h postapplication, release of proteoglycans and collagen by 6 h postapplication, and finally a rebuilding phase of collagen and proteoglycan accumulation within 24 h. Further, Nicodemus et al. 37 demonstrated that intermittent dynamic compression over 1 week significantly increased expression of MMPs and collagen type II in chondrocytes encapsulated in PEG hydrogels, and once dynamic compression was removed, MMP levels significantly dropped. Seven days after loading, MMPs were expressed at levels below those expressed before loading was applied, while collagen and GAG accumulation increased. Several other studies have suggested that mechanical stimulation's upregulation of MMPs and aggrecanases that results in degradation and release of GAGs is a necessary rebuilding process for turnover of ECM and continued development of tissue.34,54 This pattern of upregulation of an anabolic and catabolic response with mechanical stimulation, followed by downregulation of the catabolic response once loading is removed, could suggest why our constructs continue to increase in mechanical and biochemical properties after loading is removed at 2 weeks. It could also suggest why during loading we see a decrease in GAG accumulation, and after loading is removed we observe a significant increase in GAG accumulation. However, further evaluation of MMP and aggrecanase expression during culture would be required to verify.
We observed a significant temporary reduction in collagen released to the media for the week after removal of load. To our knowledge, this is the first study to observe such a delayed effect of loading, likely due to previous studies focusing on shorter culture times after loading.34,37,55 Further analysis will be required to determine whether the lack of loss of collagen after loading is due to a temporary downregulation in collagen synthesis (which seems unlikely, given the continued accumulation of collagen in the samples) or a downregulation in MMP expression or activity, as has been reported for mechanical stimulation of articular chondrocytes. 37
There are some limitations to this study that should be considered. Although we describe how combinations of loading and culture can enhance the assembly of ECM by meniscal fibrochondrocytes, the mechanisms behind these phenomena are not clear and will require a significant future study. Further, as these studies were conducted on healthy neonatal bovine fibrochondrocytes, it is unclear how the results here would translate to adult human fibrochondrocytes, particularly if obtained with a patient with joint disease. As such, understanding the catabolic components of matrix assembly in greater detail will be a focus of the future study.
Overall, in this study, we demonstrated that altered loading protocols significantly affect the mechanical properties and matrix production of TE menisci, providing an in vitro mechanism for conditioning TE menisci replacements. We demonstrated that we are able to maintain and improve mechanical and biochemical properties past 2 weeks of loading; however, these properties (3% collagen fiber concentration, 7.3–9.4 μg/mg DW GAG, 144–158 μg/mg DW collagen, and 84–108 kPa equilibrium modulus) are still not equivalent to native values (19.1% collagen fiber concentration, 56 20–30 μg/mg DW GAG,10,38 600–700 μg/mg DW collagen,10,38 and 150–220 kPa equilibrium modulus 1 ). Further investigations need to be performed to further improve the properties of our anatomically correct TE menisci. Prolonged culture past 6 weeks, treatment with growth factors, or use of a material that encourages cell attachment and better retention of ECM products could all serve to further improve the mechanical and biochemical properties of these menisci scaffolds.
Footnotes
Acknowledgments
The authors would like to thank the National Science Foundation's Graduate Research Fellowship Program, the Cornell BME NSF GK-12 program: DGE 0841291, Derin Sevenler, and the members of the Bonassar Lab for their support in this research.
Disclosure Statement
No competing financial interests exist.