
Abstract
Despite promising results from the therapeutic use of stem cells for treating ischemic diseases, the poor survival of cells transplanted into ischemic regions is one of the major problems that undermine the efficacy of stem cell therapy. Cord blood mononuclear cells (CBMNCs) are an alternative source of mesenchymal stem cells (MSCs) without disadvantages, such as the painful and invasive harvesting procedure, of MSCs derived from bone marrow or adipose tissue. In the present study, we investigated whether the angiogenic efficacy of cord blood mesenchymal stem cells (CBMSCs) can be enhanced by grafting as spheroids in a mouse hindlimb ischemia model. Human CBMSC (hCBMSC) spheroids were prepared by using the hanging-drop method. Mouse hindlimb ischemia was induced by excising the femoral artery and its branches. After surgery, the animals were divided into no-treatment, dissociated hCBMSC, and spheroid hCBMSC groups (n=8 per group) and received corresponding hCBMSC treatments. After surgery, the ischemic hindlimbs were monitored for 4 weeks, and then, the ischemic hindlimb muscles were harvested for histological analysis. Apoptotic signaling, angiogenesis-related signal pathways, and blood vessel formation were investigated in vitro and/or in vivo. The transplantation of hCBMSCs as spheroids into mouse ischemic hindlimbs significantly improved the survival of the transplanted cells by suppressing apoptotic signaling while activating antiapoptotic signaling. Furthermore, the transplantation of hCBMSCs as spheroids significantly increased the number of microvessels and smooth muscle α-actin-positive vessels in the ischemic limbs of mice, and attenuated limb loss and necrosis. Human CBMNC can be considered an alternative source of MSC, and spheroid-based hCBMSC delivery can be considered a simple and effective strategy for enhancing the therapeutic efficacy of hCBMSCs.
Introduction
To overcome such limitations, several approaches that improve engrafted cell survival, including the use of synthetic scaffolds, genetic modifications, and the codelivery of growth factors, have been attempted.11–13 Nevertheless, these approaches have limitations, such as the spatiotemporal release control for the growth factor codelivery, 14 and a potential risk of immunogenicity or mutagenesis in case of the genetic modifications. 15 As an alternative approach, we recently reported that grafting adipose-derived mesenchymal stem cells (MSCs) as spheroids can be a promising approach to improve the efficacy of stem cell therapy for ischemia. 16 In a spheroid, the mild hypoxic condition established in the inner core due to the limited nutrient diffusion can precondition the cells to an ischemic environment. 17 This preconditioning could make the cells resistant to the hypoxic condition of the target tissue. Furthermore, the spheroid culture system has been reported to increase the production of various growth factors that are necessary for cell survival. 18 Consequently, spheroid grafting could be a useful method for improving the angiogenic efficacy of the stem cells, allowing cells to be preconditioned to ischemic environments.
Although MSCs are usually obtained from bone marrow,19,20 it might be necessary to find an alternative source of MSCs due to some of the disadvantages of using bone marrow-derived stem cells, such as the painful invasive harvesting procedure. As an alternative source of MSCs, cord blood mononuclear cells (CBMNCs) that contain multi-potent MSCs have been suggested.21,22 Cord blood (CB)-derived MSCs (CBMSCs) do not differ from the MSCs obtained from other tissues in their differentiation ability. 23 Based on such reports, we investigated whether the transplantation of human CBMSC (hCBMSC) spheroids would improve the angiogenic efficacy of CBMSCs in a mouse hindlimb ischemia model. Apoptotic gene expression of hCBMSCs cultured as spheroids or as a monolayer was examined in vitro. Next, the survival and angiogenic growth factor secretion of hCBMSCs transplanted as spheroids or dissociated cells were evaluated. Finally, neovascularization in the ischemic tissues and limb salvage was evaluated.
Materials and Methods
Isolation and nomoxic culture of hCBMSC
Human CBMNCs (hCBMNCs) were purchased from (Lonza Walkersville, Inc.). hCBMSCs were isolated from hCBMNCs and, subsequently, cultivated as previously described. 24 Briefly, hCBMNCs were cultured in alpha-minimum essential medium (α-MEM; Gibco BRL), supplemented with 10% fetal bovine serum (FBS; Hyclone) at 37°C in a humidified atmosphere containing 5% CO2 with a change of culture medium twice a week. Unbound nonadherent cells were discarded by changing the medium. After 3 weeks, the cells reached ∼80% confluence and were sub-cultured to expand. The morphology of the cells was examined under a phase-contrast microscope. Neither recombinant cytokines nor growth factors were used in the culture medium. The cells of passage number 5–8 were used in this study.
To detect the possible presence of human endothelial lineage cells during cell maintenance, a flow cytometric analysis using human-specific PECAM (CD31) antibody was performed for the cells with passage number 5–8. Briefly, hCBMSCs were treated with trypsin and dissociated by gentle pipetting. The dissociated cells were washed with phosphate-buffered saline (PBS), and single-cell suspensions were then analyzed with an FACScan (Becton Dickinson) using the CellQuest software (Becton Dickinson). Both hCBMSCs cultured as spheroids and monolayers were analyzed using human-specific PECAM antibody (Sigma-Aldrich Co. LLC.).
Culture of hCBMSC spheroids
The hanging-drop method was used to form spheroids with modifications. 25 Briefly, 30 μL drops of the culture medium containing cells (1×106 cells/mL) were applied onto the inside of the lid of a Petri dish that contained PBS to prevent drying out. After 24 h of incubation in an incubator, the spheroids formed in the drops were retrieved using a Pasteur pipette. The hCBMSC spheroids (4.2×107 cells) were cultured in a siliconized spinner flask (Bellco) containing 70 mL of the culture medium (6.0×105 cells/mL) with stirring at 70 rpm for 2 days. To produce spheroids with varying sizes, the hanging-drop procedure was used with varying densities of cell suspensions (1.0×104–2.0×106 cells/mL). Spheroids were then sieved through 200 and 400 μm sieves (Becton Dickinson Labware) to separate the spheroids according to their sizes.
Hypoxic culture of hCBMSCs
To generate a hypoxic culture condition, culture dishes or spinner flasks were placed in a hypoxia chamber (MCO-18M; Sanyo) containing 1% oxygen and 5% CO2 at 37°C. The hCBMSCs were cultured with α-MEM with 100 units/mL of penicillin and 100 μg/mL of streptomycin, without an FBS supplement. For reverse transcription–polymerase chain reaction (RT-PCR), the cells were collected at 3 and 12 h after incubation under the hypoxic condition.
Overall cell death evaluation
For the evaluation of overall cell death, the same number (7.5×103 cells) of cells were used for monolayer culture and spheroid culture. The viability of hCBMSCs cultured in spheroids and monolayers under hypoxic conditions was determined by double staining with fluorescein diacetate (FDA; Sigma) and ethidium bromide (EB; Sigma). In this staining, FDA stains the cytoplasm of viable cells green, whereas EB stains the nuclei of nonviable cells red. The staining solution was freshly prepared by mixing 10 mL of FDA stock solution (1.5 mg/mL of FDA in dimethyl sulfoxide), 5 mL of EB stock solution (1 mg/mL of EB in PBS), and 3 mL of PBS (pH=8.0). The cells were incubated in the staining solution for 3–5 min at 37°C. After staining, the samples were examined using a fluorescence microscope (IX71 inverted microscope; Olympus). For 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay, MTT solution was added to the culture medium (10% v/v of culture medium), and the cells were incubated for 4 h. After incubation in the MTT solution, formed purple formazan crystals were dissolved in dimethyl sulfoxide, and the absorbance at 570 nm was measured.
Reverse transcription–polymerase chain reaction
The collected cells were lysed in TRIzol reagent (Invitrogen). Reverse transcription was performed using 5 μg of total RNA and SuperScriptTM II reverse transcriptase (Invitrogen). The synthesized cDNA was amplified by PCR, and the primers used for RT-PCR are as follow: BAX (forward 5′-GTG CAC CAA GGT GCC GGA AC-3′, reverse 5′-TCA GCC CAT CTT CTT CCA GA-3′), Bcl-2 (forward 5′-AGA TGT CCA GCC AGC TGC ACC TGA C-3′, reverse 5′-AGA TAG GCA CCA GGG TGA GCA AGC T-3′), and β-actin (forward 5′-GCA CTC TTC CAG CCT TCC TTC C-3′, reverse 5′-TCA CCT TCA CCG TTC CAG TTT TT-3′).
Real-time PCR
Real-time PCR was used to quantify the relative gene expression level of BAX and Bcl-2 (n=6 per group). Total RNA was extracted from samples using 1 mL Trizol reagent (Invitrogen) and 200 μL of chloroform. The lysated samples were centrifuged at 12,000 rpm for 10 min at 4°C. RNA pellet was washed with 75% (v/v) ethanol and dried. After the drying procedure, the samples were dissolved in RNase-free water. For real-time quantitative PCR, the iQ™ SYBR Green Supermix kit (Bio-Rad Laboratories) with the MyiQ™ single color Real-Time PCR Detection System (Bio-Rad Laboratories) were used.
Mouse hindlimb ischemia model
Mouse hindlimb ischemia was induced as previously described. 26 All the animal treatments and experimental procedures were approved by the Institutional Animal Care and Use Committee of the Seoul National University (SNU-110630-2).
Treatment of limb ischemia
One day after the surgery, the mice were randomly divided into three groups (n=8 for each group). The hCBMSCs for transplantation were cultured in a monolayer or as spheroids using culture media containing 10% (v/v) FBS. Dissociated hCBMSCs were prepared by harvesting the monolayer-cultured hCBMSCs using trypsin. The dissociated cells or spheroids of hCBMSCs (1.0×107 cell/200 μL PBS) were intramuscularly injected into the gracilis muscle of the medial thigh. The morphological status of the ischemic limb was monitored for 4 weeks. Mice with surgery but without treatment served as a negative control. Normal mice that did not undergo surgery served as a positive control.
Immunohistochemistry
The ischemic limb muscles were harvested 4 weeks after the surgery and treatment. The samples were embedded in Optimal Cutting Temperature compound (TISSUE-TEK® 4583; Sakura Finetek USA. Inc.) and then cryo-sectioned at a thickness of 10 μm at −22°C. For quantification of the blood vessels and incorporation of the engrafted human cells in the ischemic region, the slides were subjected to immunofluorescence staining with anti-von Willebrand factor (vWF) antibody (Abcam), antismooth muscle (SM) α-actin antibody (Abcam), and anti-human nuclear antigen (HNA) antibody (Abcam). FITC-conjugated secondary antibodies (Jackson ImmunoResearch Laboratories) were used. The sections were counterstained with DAPI and examined using a fluorescence microscope (Nikon TE2000). For quantification, twenty images per slide from three different samples per group were randomly analyzed.
Western blot analysis
Ischemic hindlimb muscles (three samples per group) were homogenized using a Dounce homogenizer (50 strokes, 4°C) in an ice-cold lysis buffer. Western blot analysis was carried out by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis using antibodies against human vascular endothelial growth factor (hVEGF; Abcam), human fibroblast growth factor 2 (hFGF2; Abcam), hepatocyte growth factor (HGF; Abcam), caspase-3 (Abcam), HNA (Abcam), intercellular adhesion molecule 1 (ICAM-1; Abcam), vascular cell adhesion molecule 1 (VCAM-1; Abcam), and NG2 (Abcam). Immunopositive bands were developed with an enhanced chemiluminescence detection kit (Amersham Bioscience).
Histological examination
The ischemic limb muscles were harvested 28 days after treatment. The specimens were fixed in 10% (v/v) buffered formaldehyde, dehydrated in a graded ethanol series, and embedded in paraffin. Specimens were sliced into 4-μm sections and stained with Masson's trichrome staining to assess tissue fibrosis.
Statistical analysis
For the quantification analysis, at least three independent experiments were performed. The results are shown as mean value±standard deviation For two-group comparisons, a t-test was used. For the comparison of more than three groups, a one-way analysis of variance test with the Bonferroni correction was performed (OriginPro 8 SR4 software, ver. 8.0951; OriginLab Corporation Northampton). p-Values of <0.05 were considered as indicating statistical significance.
Results
Morphological examination of CBMSCs and spheroid
Under a phase-contrast microscope, 4 day-cultured adherent hCBMSCs with passage number 2 showed a fibroblast-like morphology with long and thin cell processes, which is typical of MSCs (Fig. 1A). The typical morphology and size of spheroids used for the experiments is shown in Figure 1B.

Characterization of cord blood mesenchymal stem cells (CBMSCs) and CBMSC spheroids.
Effects of spheroid diameter
After being exposed to hypoxia for 24 h, spheroids with a diameter of 200–400 μm showed an obvious increase in angiogenic factor production (VEGF, FGF2, and HGF; Fig. 1C) and antiapoptotic Bcl-2 expression (Fig. 1D) compared with spheroids with different diameters (<200 μm or >400 μm).
Overall cell death in monolayer-cultured cells and spheroids
Cells grown in monolayers and spheroids were exposed to hypoxia up to 12 h. Compared with a monolayer culture where most cells were stained red (dead), the spheroid group showed more green (live) color (Fig. 2A). The MTT assay result indicated that cell viability is higher in the spheroid group compared with the monolayer group (Fig. 2B).

Overall cell death compared between monolayer-cultured hCBMSC and spheroid-cultured hCBMSCs. The same number of cells in either monolayer culture or spheroid culture was exposed to hypoxia up to 12 h.
Antiapoptotic effect of three-dimensional spheroid culture with or without hypoxia
The RT-PCR results indicated that mRNA expressions of apoptotic factor BAX and antiapoptotic factor Bcl-2 were barely detectable in both monolayer-cultured cells and spheroid-cultured cells under normoxic condition up to 72 h (Fig. 3). However, under hypoxic condition, BAX expression was significantly increased in the monolayer-cultured cells, while Bcl-2 expression was significantly increased in the spheroid-cultured (Fig. 3A). According to the results of real-time PCR, at 72 h, the BAX expression of the monolayer-cultured cells under hypoxia was >100-fold higher compared with that of the cells under normoxia (138.4-fold against the monolayer-cultured cells and 178.2-fold against the spheroid-cultured cells), and even compared with the spheroid-cultured cells under hypoxia, it was approximately ninefold higher (Fig. 3B). On the other hand, in case of the Bcl-2 expression, the spheroid-cultured cells under hypoxia showed highest expression compared with all other groups (25.1, 24.2, and 11.5-fold higher against the monolayer-cultured cells under normoxia, the spheroid-cultured cells under normoxia, and the monolayer-cultured cells under hypoxia, respectively) (Fig. 3C).
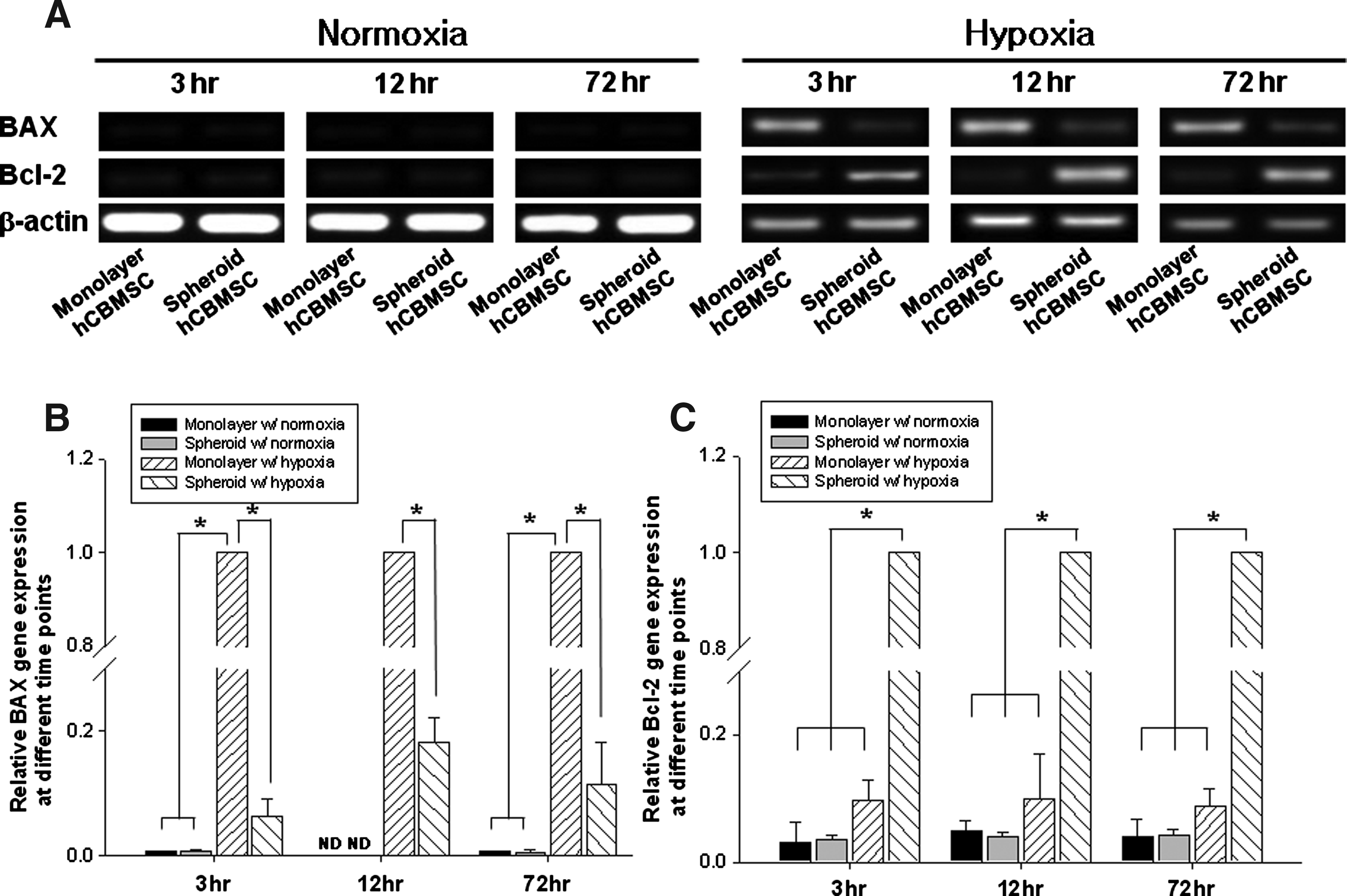
Antiapoptotic effects of spheroid culture with or without hypoxia. hCBMSCs were cultured as either a two-dimensional monolayer or three-dimensional spheroids under either normoxic or hypoxic conditions. The cells were collected at 3, 12, and 72 h, and were subjected to an RT-PCR using apoptotic BAX primers and antiapoptotic Bcl-2 primers.
Enhanced cell survival and growth factor secretion by spheroid transplantation
The results of western blot using cell-transplanted tissue samples showed that the amount of HNA was approximately sixfold higher in the spheroid transplantation group compared with the monolayer cell transplantation group, while the amount of caspase-3 was approximately fourfold higher in the monolayer transplantation group (Fig. 4A). The amounts of hVEGF and hFGF2 were significantly higher (5.6 and 3.3-fold higher, respectively) in the spheroid transplantation group compared with the monolayer cell transplantation group (Fig. 4B).
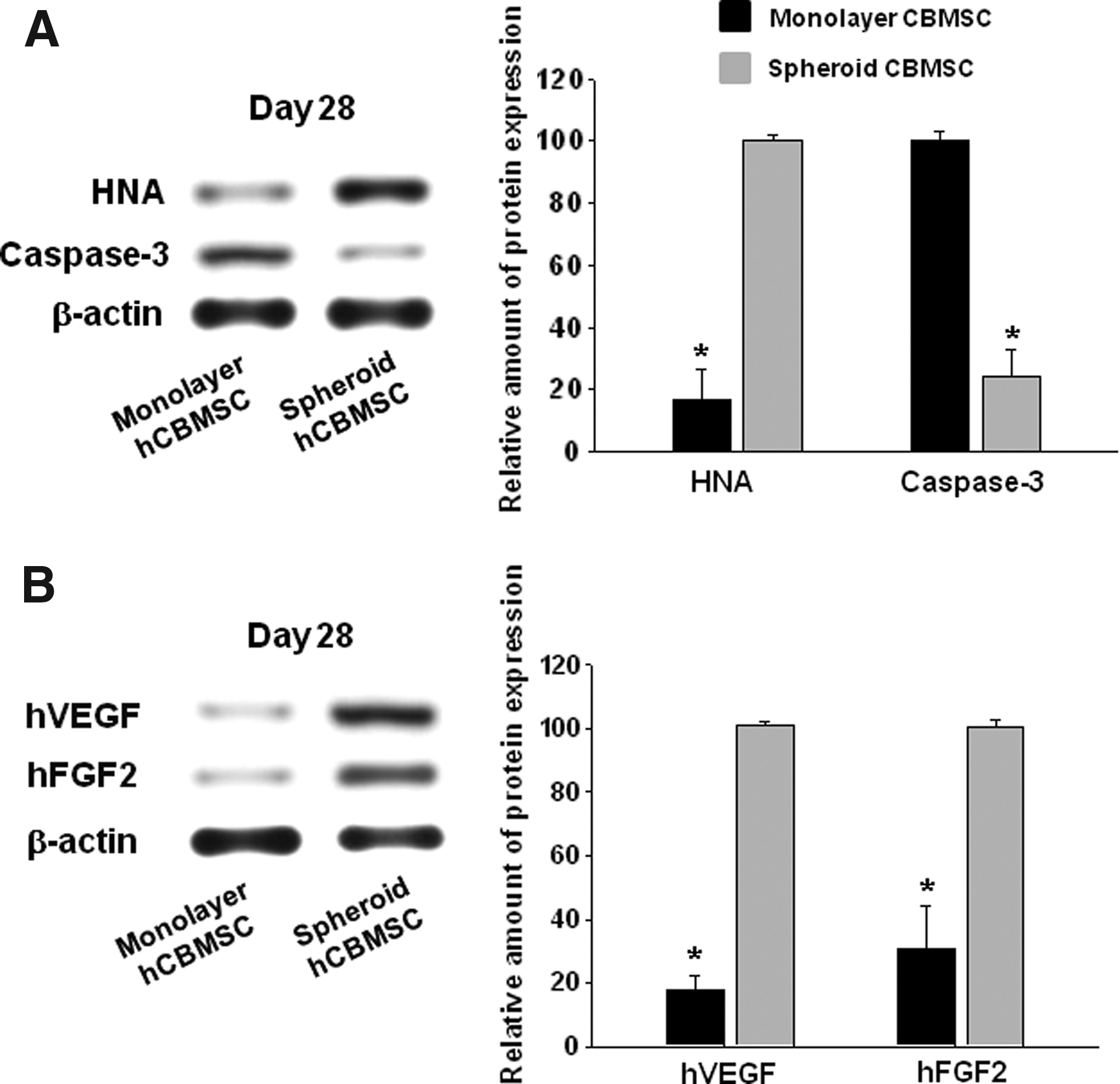
hCBMSCs transplanted as spheroids into mouse ischemic hindlimbs showed improved cell survival and angiogenic growth factor secretion.
Enhanced angiogenesis by spheroid transplantation
According to the result of flow cytometry analysis using an endothelial cell marker human-specific PECAM antibody, the portion of the human-PECAM positive cells among cultured CBMSCs was about 1% or less (Table 1). To detect blood vessels in the ischemic hindlimbs, the sectioned tissue samples were immunostained for another endothelial cell marker vWF and an SM cell marker SM α-actin. Compared with both the no-treatment group (1.7±1.5 vessels/mm2) and the monolayer cell transplantation group (31.7±14.6 vessels/mm2), the number of vWF-positive microvessels detected in the ischemic region significantly increased in the spheroid transplantation group (99.0±23.6 vessels/mm2) (Fig. 5). No detectable HNA/vWF double-positive cells were observed. Similar to this result, the number of SM α-actin-positive vessels in the ischemic region also increased in the spheroid transplantation group (11.7±4.9 vessels/mm2) compared with both the no-treatment group (0.33±0.6 vessels/mm2) and the monolayer cell transplantation group (3.0±2.0 vessels/mm2) (Fig. 6A, B). Furthermore, the number of HNA/SM α-actin double-positive cells was significantly higher in the spheroid hCBMSC group (6.3±2.3×104 cells/mm2) compared with the monolayer hCBMSC group (1.0±1.0×104 cells/mm2) (Fig. 6C, D).
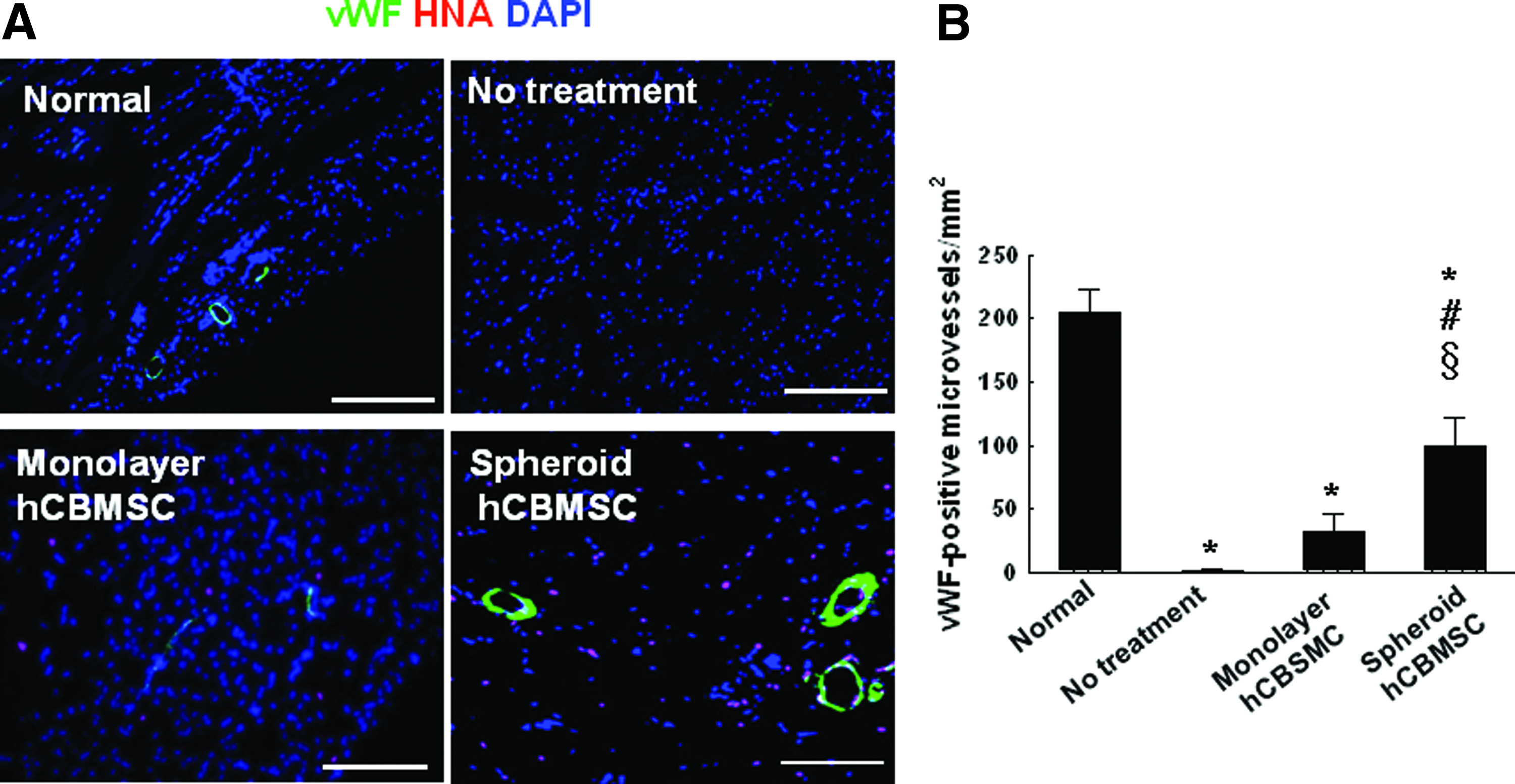
von Willebrand factor (vWF)-positive microvessels in the ischemic region after 28 days of treatment.
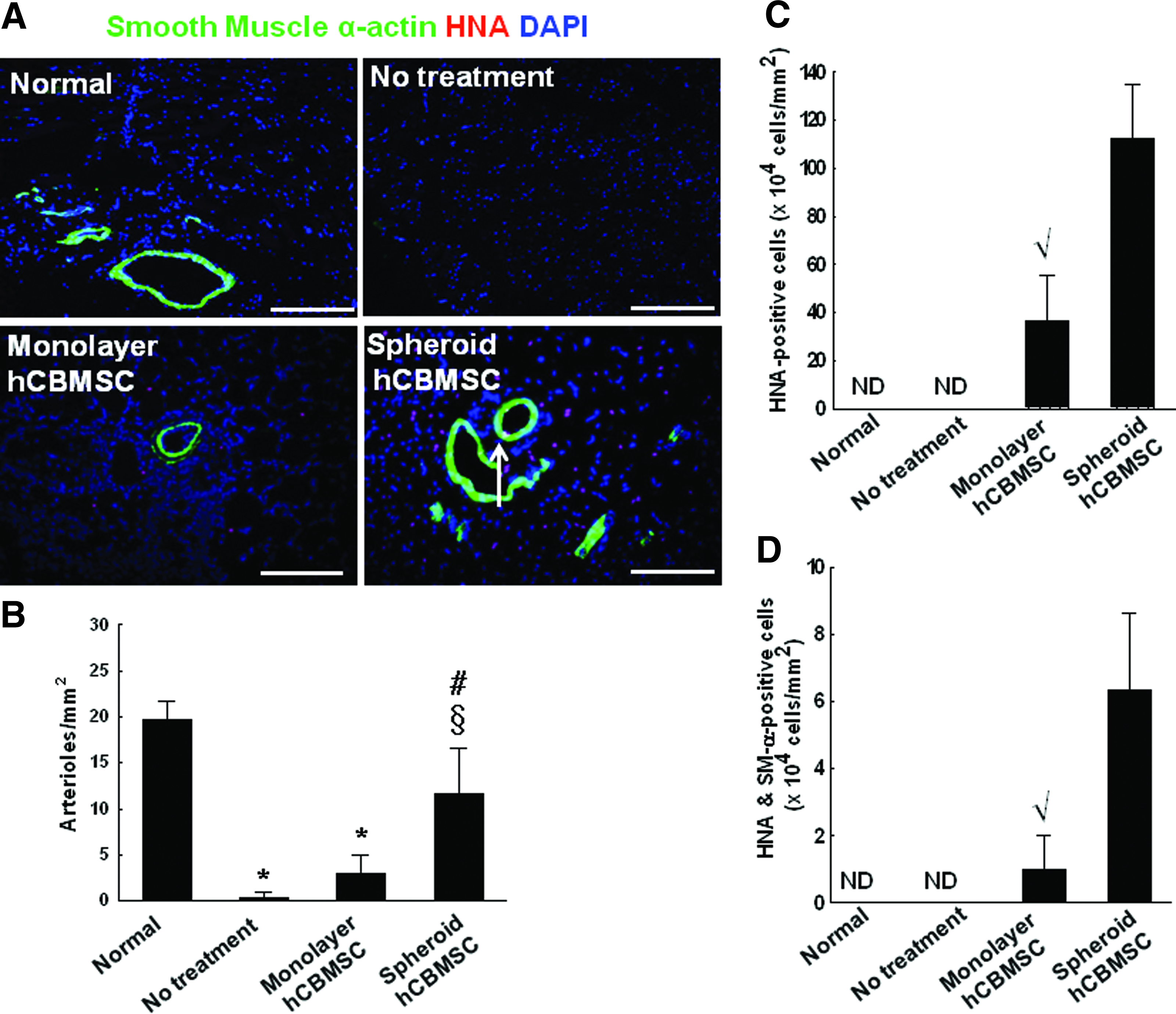
The transplantation of hCBMSCs as spheroid into mouse ischemic hindlimbs enhanced angiogenesis.
hCBMSCs, human cord blood mesenchymal stem cells.
Increased expressions of cell adhesion molecules and factors involved in vessel maturation in spheroid hCBMSC transplanted tissue
The amount of cell adhesion molecules, such as ICAM and VCAM, expressed in the spheroid transplantation group was not significantly different from that of the normal tissue, while the no-treatment or monolayer transplantation group showed significantly (50.8- and 6.5-fold, respectively) decreased ICAM expression compared with the spheroid transplantation group. Furthermore, VCAM expression was also significantly (75.0- and 6.5-fold, respectively) decreased in the no-treatment or monolayer transplantation group compared with the spheroid transplantation group (Fig. 7). The expression of NG2, a pericyte marker, 27 was also significantly increased in the spheroid transplantation group compared with the other groups (9.6, 300.0, and 6.4-fold increase compared with the normal, no-treatment, and monolayer transplantation group, respectively).

Expressions of adhesion molecules and factors involved in blood vessel maturation.
Improved limb salvage and attenuated fibrosis by spheroid transplantation
In the no-treatment group, no limb salvage was observed. The monolayer cell transplantation group had a limb salvage rate of 12.5% (one out of eight animals, Fig. 8A). The spheroid transplantation group showed a limb salvage rate of 75% (six out of eight animals). In addition, an obvious decrease in fibrosis in the ischemic region was observed in the spheroid transplantation group, as shown by Masson's trichrome collagen staining (Fig. 8B). The extent of fibrosis was the highest in the no-treatment group, followed by the monolayer cell transplantation group.
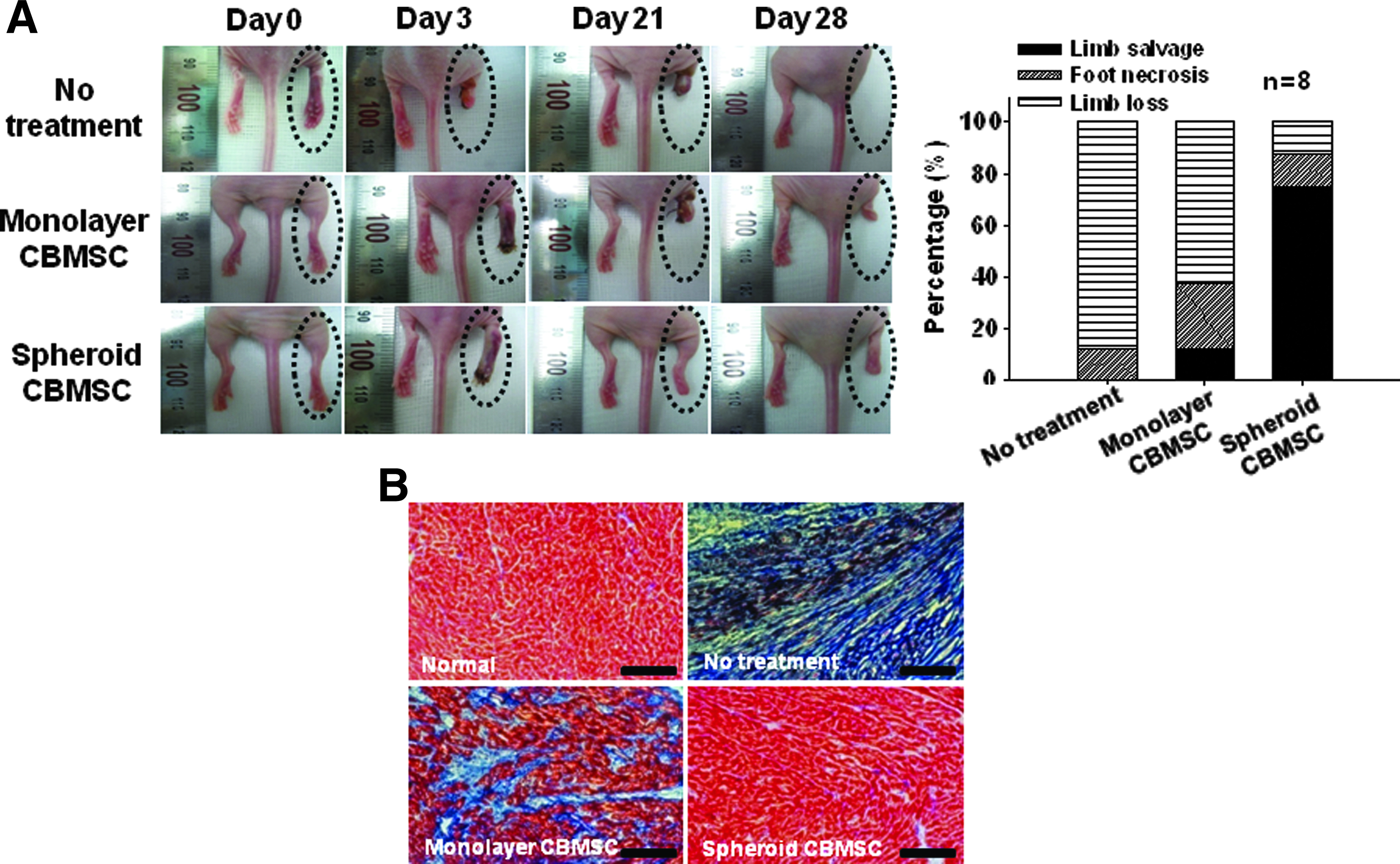
The transplantation of hCBMSCs as spheroids improved ischemic limb salvage and decreased fibrosis.
Discussion
In treating ischemic diseases using stem cells, the low survival rate of transplanted cells is the major problem that should be solved by applying stem cell therapy in clinical settings. The hypoxic condition in the ischemic region causes irreversible ischemic injury to the transplanted cells and eventual apoptosis, and these sequential events are believed to be the major cause of the low survival rate of the transplanted cells. 6 Exposing cells, including MSCs, to mild hypoxia may precondition the cells so that they can survive subsequent hypoxic insults. 28 The mild hypoxic condition established in the core of spheroids 29 is similar to that of avascular tissue with limited diffusion for oxygen and nutrients. 30 Thus, the spheroid culture system in which cells can be naturally exposed to a mild hypoxic condition can be used to precondition cells in vitro so that they can survive the harsh hypoxic condition of ischemic tissue into which they are transplanted. Here, we report that the hCBMSCs transplanted into mouse ischemic hindlimb as spheroids increased the transplanted cell survival, growth factor secretion, angiogenesis of the ischemic region, and subsequent limb salvage.
It is known that molecular diffusion in spheroids is inversely related to square of the diameter, 31 and it means there should be a critical diameter of spheroids with which spheroids are responsive to hypoxic preconditioning. Our data indicated that spheroids with a diameter of ∼200–400 μm were most responsive to hypoxia in our experimental settings, increasing angiogenic protein production and antiapoptotic gene expressions (Fig. 1C, D). The enhanced survival of hCBMSC spheroids transplanted into mouse ischemic hindlimbs may be attributed to enhanced antiapoptotic factor secretion and attenuated apoptosis. The mild hypoxic condition of spheroids 17 seems to be responsible for the increased production of VEGF32–34 and FGF2. 35 The antiapoptotic effects of VEGF either on vasculature or on endothelial cells have been reported.36,37 FGF2 has been reported to exert antiapoptotic effects by decreasing DNA fragmentation and preventing the down-regulation of antiapoptotic signaling, such as Bcl-2, in an animal focal cerebral ischemia model. 38 We previously observed a significant increase in the secretion of VEGF and FGF2 and the up-regulation of antiapoptotic signals in mouse ischemic hindlimbs after transplanting human adipose-derived stromal cells (hADSCs) as spheroids 16 In the present study, we also observed that the transplantation of hCBMSC spheroids inhibited apoptotic signaling, such as BAX signaling 39 and caspase-3 signaling. 40 Thus, the transplantation of hCBMSC spheroids seems to increase antiapoptotic factor secretion, thereby attenuating apoptosis of the transplanted cells.
However, one thing that should be considered regarding the antiapoptotic effect of spheroids observed in our study is the role of apoptosis in normal wound healing process. Apoptosis is also a necessary process in normal wound healing to remove inflammatory cells, and to prevent scar formation. 41 Since the inflammatory response occurs within a week during the course of the wound-healing process, 42 even if a pro-apoptotic event occurred while removing inflammatory cells, we would not have detected it, because we checked the apoptotic status at day 28. Consequently, we cannot completely exclude the possibility that a pro-apoptotic event had occurred in the early stage of the in vivo experiment even in the spheroid-treated group.
In addition to the paracrine effect of transplanted hCBMSC spheroids, we also observed the colocalization of HNA and SM α-actin. However, the colocalization was observed in only a few cells, indicating that only a small number of the transplanted cells were directly incorporated into blood vessels. In addition, the percentage of PECAM-positive cells among the cultured hCBMSCs in vitro was <1%. These findings suggest that even if transplanted hCBMSCs directly (as opposed to the paracrine effect) contributed to the improved angiogenesis observed in the present study, the impact by such direct involvement would be not as strong as that by the paracrine effect of the hCBMSC spheroids. In other words, the paracrine effect of hCBMSC spheroids, rather than direct vessel incorporation or vascular differentiation of the transplanted cells, seems to play a more important role in inducing the angiogenic effect.
Th prevention of anoikis may be another explanation for the enhanced survival of the transplanted hCBMSCs as spheroids. Anoikis is a pro-apoptotic event that occurs when anchorage-dependent cells are detached from the surrounding ECM. 43 The interactions between the cells and the surrounding ECM are the important part of the mechanisms that provide essential growth and survival signals for the cells. 44 The detachment of the cells from the surrounding ECM can initiate the process of anoikis, resulting in a loss of cell survival signaling. Thus, the spheroid-harvesting procedure, which did not require a harsh proteolytic enzyme treatment (e.g., trypsin) to prepare cells for transplantation, might have contributed to the enhanced cell survival by better preserving the ECM in the spheroids as compared with the monolayer-cultured cells that were harvested by the trypsin treatment for cell transplantation.
According to our data, the hCBMSCs transplanted as spheroids significantly increased the number of microvessels and SM α-actin-positive blood vessels in the ischemic limbs of mice, and also attenuated limb loss and necrosis in vivo as compared with the hCBMSCs transplanted as dissociated cells. This enhanced the angiogenic effect of hCBMSC, as spheroids may be because the implanted cells as spheroids stayed at the site of implantation longer, increasing the number of effective cells as compared with the transplantation of dissociated hCBMSCs (Fig. 6C), or could be, at least in part, contributed to the increased ICAM (Fig. 7), which modulates the recruitment of circulating endothelial progenitor cells to the ischemic regions for neovascularization.45–47 Furthermore, the enhanced expression of VCAM and NG2 by the transplantation of hCBMSC spheroids (Fig. 7) suggests maturation of the neovessels by pericytes.27,48,49
In summary, we report on a spheroid-based hCBMSC for enhanced vascularization in ischemic tissue. The transplantation of hCBMSC spheroids significantly enhanced the angiogenic efficacy of hCBMSC therapy for treating hindlimb ischemia by improving cell survival and paracrine factor secretion in ischemic tissues as compared with the transplantation of conventional monolayer-cultured cells. The results of this study indicate that hCBMNC can be an alternative source of MSC, and spheroid-based hCBMSC transplantation may be a simple and effective strategy for increasing the therapeutic efficacy of hCBMSCs for ischemic disease treatment.
Footnotes
Acknowledgment
This study was supported by a grant (2010-0020352) from the National Research Foundation of Korea.
Disclosure Statement
No competing financial interests exist.