
Abstract
Hepatocyte functionality and survival decrease rapidly in culture, and both can be improved using bone marrow-derived mesenchymal stromal cells (MSCs). We have previously described an alternative, more plentiful source of MSCs coming from the perivascular area of the umbilical cord, human umbilical cord perivascular cells (HUCPVCs). Our objective was therefore to ascertain whether HUCPVCs could serve as hepatocyte stromal cells ex vivo. For this purpose, rat hepatocytes were cocultured in contact with HUCPVCs (contact coculture). Also, HUCPVCs were cocultured separated from hepatocytes with a semipermeable membrane (noncontact coculture) to assess soluble factor interactions. Next, an HUCPVC-conditioned medium (CM) was used to investigate the possibility of HUCPVC-free support, while flash-frozen HUCPVCs were employed to investigate the effects of nonsoluble interactions. In all experiments, medium samples were taken daily to assess the production of albumin. Also, at certain days, the levels of cytochrome P450 (CYP) activity and urea secretion were tested. RNA extraction was performed at the end of experiments. Our results show that HUCPVCs in contact and noncontact cocultures with hepatocytes improve albumin gene expression and secretion compared to monoculture. Flash-frozen HUCPVCs had a late improvement in albumin secretion, while CM improved it for a short period. Ureagenesis maintenance was improved by contact coculture and flash-frozen HUCPVCs. CYP activity was significantly increased in the presence of flash-frozen HUCPVCs and in noncontact cocultures. We conclude that HUCPVCs can act as stromal cells for rat hepatocytes, and that soluble and nonsoluble factors induce differential effects on hepatocytes.
Introduction
According to the International Society for Cellular Therapy (ISCT), an MSC is a cell that is plastic adherent, able to differentiate into at least the osteogeneic, chondrogeneic, and adipogenic lineages, and expresses CD105, CD73, and CD90 while expressing no hematopoietic markers or HLA-DR molecules. 7 BM-MSCs, specifically, are also immunopriviledged and immunomodulatory cells,8–10 presenting a further benefit for bioartificial liver systems (BALS), since liver failure often results from an uncontrolled inflammatory response within the liver. 11 However, the availability of BM-MSCs for BALS applications is limited: the harvest of BM from volunteer donors is an invasive procedure, and MSCs can only be acquired from individuals above the age of consent. Furthermore, the colony-forming unit fibroblast (CFU-F) frequency and the life span of BM-MSCs decline with increasing age;12,13 therefore, it is expedient to seek alternative, high-yield sources of MSCs.
We harvest human mesenchymal cells from the Wharton's Jelly surrounding the umbilical cord vessels and call them human umbilical cord perivascular cells (HUCPVCs). These cells have shown many similarities with BM-MSCs, and comply with the minimal criteria set by the ISCT,7,14–16 although they exhibit a higher proliferation rate 16 and CFU-F frequency at harvest—being 1:33314 compared to 1:104 for BM-MSC in a newborn. 17 In addition, as with BM-MSCs, HUCPVCs are both nonalloreactive and immunoregulatory in vitro. 18
Thus, we hypothesized that HUCPVCs would enhance the maintenance and function of hepatocytes ex vivo. To test our hypothesis, we employed a xenogeneic system for two reasons. First, while hepatocytes from different species have different levels of functionality, 19 the xenogeneic system has the important advantage of allowing the identification of each cell population without sorting, or removal from their acquired tissue architecture. Second, rat hepatocyte function has been shown to correlate with that of human hepatocytes.20,21
Materials and Methods
Hepatocyte isolation and culture
Hepatocytes were isolated from male Wistar rats (Charles River Laboratories) (250–350 g) by two-step collagenase perfusion of the liver as described previously by Moldeus et al. 1978. 22 The cells were further purified by Percoll centrifugation (10 min at 50 g; a final density of ∼1.06 g/mL). 23 For each experiment, three rat livers were harvested. The hepatocytes acquired were plated at a concentration of 2.83×104 hepatocytes/cm2, and maintained in a hepatocyte culture medium (HCM): DMEM (Sigma; D6046) supplemented with 10% fetal bovine serum (FBS), 1 μM dexamethasone (Sigma; D8893), 1×ITS (MP Biomedicals; 2001344), 100 U/mL penicillin, and 100 μg/mL streptomycin (Invitrogen; 15140–122).
HUCPVC culture and source
HUCPVCs were provided by Tissue Regeneration Therapeutic, Inc. The cells were expanded in an alpha-MEM (Invitrogen; 12571-071) supplemented with 10% FBS and antibiotics: penicillin G 167 U/mL, amphotericin B 0.3 mg/mL, and gentamicin 50 mg/mL (Sigma). The medium was changed 2–3 times a week, and cells were passaged when confluence was close to 80%. For coculture experiments, HUCPVCs were pooled from at least five different donors.
Contact coculture
On the day following the seeding of hepatocytes, after medium samples were collected, HUCPVCs were laid on top of hepatocytes at a concentration of 1×104 cells/cm2. For contact coculture and monoculture in 3D configuration, tissue culture-treated polystyrene scaffolds (3D Biotek; PS30401) with a fiber diameter of ∼300 μm and a separation between fibers of ∼400 μm were seeded with hepatocytes according to the manufacturer's instructions (a detailed description of the scaffold-seeding procedure is presented in the Supplementary Methods section). Medium samples were collected daily and stored at −20°C. The medium used for both cocultures and monocultures was HCM, described above.
Transwell coculture
On the day following the seeding of hepatocytes, after medium samples were collected, 24-well plate 0.45-μm pore size transwell inserts (BD Falcon; 353095) with 2×104 HUCPVCs were added. Following this, medium samples were collected daily and stored at −20°C. The medium used for both cocultures and monocultures was HCM, described above.
Production of HUCPVC-conditioned medium
HUCPVCs were seeded at a concentration of 1×104 cells/cm2 and were grown for at least 5 days in the HCM. To produce the conditioned medium (CM), the medium was changed to the DMEM (Sigma; D6046) supplemented with 0.05% bovine serum albumin. After 20 h, the CM was harvested and concentrated 25 times using ultrafiltration units (Millipore; UFC900308) with a 3-kDa molecular weight cut-off. The concentrated CM, remaining on top of the filters, was stored at −80°C until use.
Hepatocytes were provided with the CM at two concentrations 1×and 5×: the 25×CM was thawed at room temperature and then was diluted at the appropriate proportions with the components of the HCM, so that the final medium would be HCM supplemented with 1×or 5×CM medium.
Flash-frozen HUCPVCs
After culturing HUCPVCs in an HCM for at least 6 days, the medium was removed, and the bottom of the plates was immediately submerged in liquid nitrogen until the nitrogen stopped boiling. After flash freezing the plates, they were stored at −20°C until use (a more detailed description of the procedure is presented in the Supplementary Methods section).
The day when hepatocytes were seeded on top of the flash-frozen HUCPVCs, the plates were first allowed to warm to room temperature before seeding the cells. After seeding, the plates were treated as the other cocultures: medium samples were collected daily and stored at −20°C.
Mitomycin treatment
To test the effect of the HUCPVC number on hepatocytes, HUCPVCs were growth arrested. Briefly, HUCPVCs in culture were incubated in a culture medium supplemented with 15 μg/mL of mitomycin C (Sigma; M4287) for 2 h. After the incubation, the mitomycin C-supplemented medium was removed, and the cells were washed three times with PBS before adding a fresh medium. HUCPVCs were used at least 5 h after treatment and seeded as described previously. Mitomycin-treated HUCPVCs (MT-HUCPVC) were used at either a high (2.5×104 MT-HUCPVC/cm2 - MH) or a low (1×104 MT-HUCPVC/cm2 - ML) number on hepatocytes, and compared with normal HUCPVCs at an initial seeding density of 1×104 HUCPVC/cm2.
Hepatocyte functional assays
Albumin enzyme-linked immunosorbent assay
Daily samples stored at −20°C were tested for albumin levels by means of a rat albumin enzyme-linked immunosorbent assay quantitaion set (Bethyl laboratories; E110-125) as per the manufacturer's recommendations. The results were normalized to the production per hour of 1×106 hepatocytes seeded or each scaffold.
Ureagenesis and CYP activity measurements
At some time points, both cocultures and monocultures were incubated at 37°C with 5% CO2 for 1 h while being rocked (200 μL for 24-well plate and 300 μL for scaffolds) in a serum-free HCM supplemented with 1 mM ammonium chloride (stock solution 400 μM) and 5 μM Vivid BOMR substrate (stock 2 mM in acetonitrile, Invitrogen; P2865). Vivid BOMR substrate can be metabolized by CYP3A and CYP1A family members into a highly fluorescent metabolite. Thus, after incubation, 100 μL samples were transferred to 96-well plates, and fluorescence (excitation 530, emission 605) was read at 37°C to detect CYP activity. Samples were then frozen at −20°C for the later measurement of urea levels. Urea was quantified using the QuantiChrom Urea Assay Kit (Bioassay systems; DIUR-500). After diluting the samples to a concentration within the detection range, they were allowed to react with the working reagent contained in the kit for 50 min, and then the absorbance was read at 430 nm.
RNA isolation and reverse transcription
To isolate the total RNA from the monocultures and cocultures, they were lyzed using TRI Reagent (Sigma; T9424), followed by the company's recommended procedure. RNA content and quality were measured using spectrophotometry quantitation (NanoDrop 1000; Thermo Fisher Scientific), and genomic DNA elimination and reverse transcription (RT) were performed using the QuantiTect Reverse Transcription Kit (Qiagen; 205311), according to the manufacturer's instructions.
Relative quantitative RT–polymerase chain reaction
After RT, each sample was run in triplicate using the SYBR Green JumpStart Taq ReadyMix (Sigma; S4438) according to manufacturer's recommendations at an annealing temperature of 60°C and 78°C for fluorescence measurement. All the primers (Supplementary Table S2; Supplementary Data are available online at www.liebertpub.com/tea) were designed and tested so that they would not cross-react with human RNAs by running them on HepG2 cDNA. Quantitation was done by normalizing the expression levels of all the genes to the housekeeping gene beta-Actin (ActinB) using the Pfaffl method. 24
Immunofluorescence
Cultures were labeled, at day 1 and 10, for albumin to identify the presence of hepatocytes and the percentage of area covered by them (a detailed description of the procedure is presented in the Supplementary Methods section). All samples within the same experiment were stained simultaneously, and three biological replicates were used per experimental group. The primary antibodies used were sheep anti-rat albumin antibody (1:200; Bethyl labs; A110-134A) to detect albumin and rat anti-rat ZO-1 (1:50, Santa Cruz Biotechnology; sc-33725) to detect the presence of tight junctions.
Statistical analysis
Isolated time points were statistically analyzed of by means of the Wilcoxon Rank-Sum test or Student's t-test analysis after logarithmic transformation. Albumin secretion curves were also analyzed after logarithmic transformation using the Generalized Estimating Equation (GEE), with an assumed equicorrelated structure. Each figure legend indicates which technique was used, number of samples, and the p-value that was considered of significance.
Results
Our experiments were conducted to assess hepatocyte function in either contact or transwell cocultures with HUCPVCs, compared to hepatocytes in monoculture. Cocultures demonstrated differential improvements in the maintenance of hepatocyte function when compared to monocultures. Albumin secretion by hepatocytes in both monoculture and cocultures decreased initially, compared to levels secreted at day 1, but both contact and transwell cocultures increased this hepatocyte secretory capability after day 4 (Fig. 1A), with the first significantly increased levels relative to monoculture (p<0.01) on day 6. These increases peaked at days 6 and 8 for transwell and contact cocultures, respectively. By day 10, transwell cocultures showed no improvement relative to monoculture. Improvements in function were also seen in 3D cocultures (Fig. 1B).

Hepatocytes cocultured with human umbilical cord perivascular cells (HUCPVCs) separated by transwell membranes or in contact in 2D and 3D configuration. Transwell coculture=hatched bars; contact coculture=solid black bar; and monoculture=white bar.
Ureagenesis decreased throughout the whole culture period in both monoculture and transwell coculture, but while it also decreased from initial values in contact coculture, it stabilized from day 4 onward (Fig. 1C). Again, a similar trend was seen in 3D cocultures, although in both cocultures and monocultures, a transitory increase compared to day 1 levels was observed (Fig. 1D). CYP activity was significantly increased at all time points in transwell coculture compared to either contact coculture or monoculture, showing an upward trend to day 5 and decreasing thereafter (Fig. 1E). While there was also some improvement with contact cocultures, the difference between CYP activity in coculture compared to monoculture was more marked in 3D conditions (Fig. 1F). The expression of hepato-specific genes, albumin, tryptophan 2,3-dioxygenase (TO), and tyrosine aminotransferase (TAT), was significantly higher in contact coculture than in either transwell coculture or monoculture (Fig. 2A). Human mRNA analysis showed that HUCPVCs express several hepatotrophic factors in contact coculture (Fig. 2B): the ECM proteins; decorin, fibronectin, laminin and collagen type I; and the cytokines: interleukin-6, stem cell factor, epidermal growth factor, hepatocyte growth factor, and a low expression of tumour necrosis factor alpha. Connexin-43 and the Notch ligand Jagged1 were also expressed.
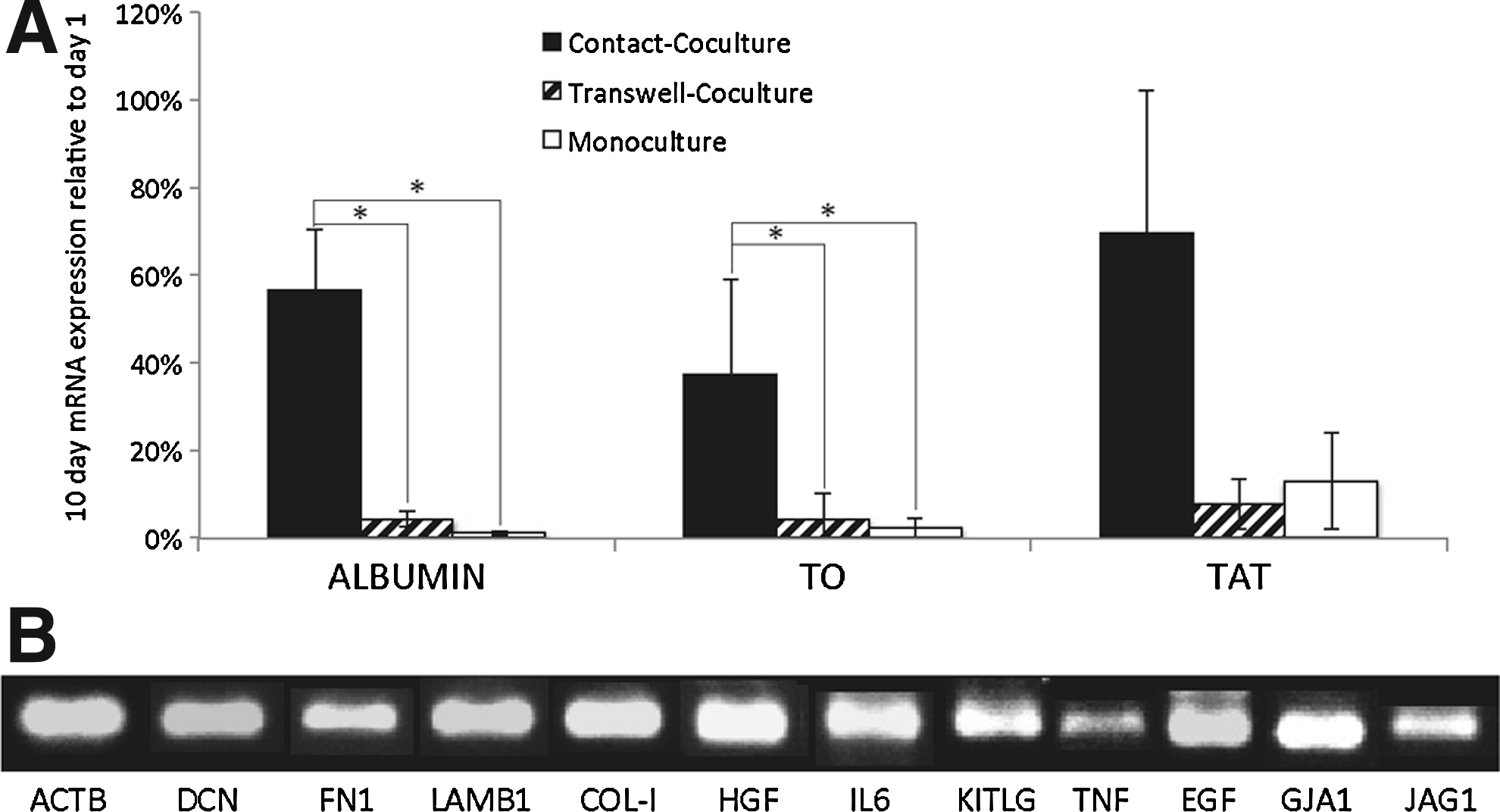
Gene expression analysis of hepatocytes and HUCPVCs. Transwell coculture=hatched bars; contact coculture=solid black bar; and monoculture=white bar.
These functional differences were reflective of the morphology of the various cultures. Hepatocytes, identified using anti-rat albumin antibody and viewed by phase and fluorescent microscopy, showed the formation of colonies surrounded by HUCPVCs in contact cocultures (Fig. 3A). Also, hepatocytes in contact coculture showed the presence of tight junctions while none were found in monoculture on day 10 (Fig. 3B). Albumin-expressing cells were present in all cultures, although in different numbers. In monoculture, they were scarce, and when found, the fluorescence signal was not as strong as from cells in cocultures. We used the fluorescent emission from albumin-expressing cells (albumin+) as a means to quantify the area covered by mature hepatocytes, which could be correlated with the functional hepatocyte number, as the cell size remained unchanged between hepatocytes on day 1 and coculture by day 10 (hepatocyte span ∼31 μm), and the significant dedifferentiation of monocultures by day 10 made any cell spreading negligible. This showed that contact cocultures exhibited a significant increase in the area covered by mature hepatocytes, compared to day 1 cultures (p<0.01), indicating a net proliferation of mature hepatocytes (Fig. 3C); this increase was also accompanied by an increase in the colony size (Fig. 3A, D). Furthermore, there was an obvious difference in the area covered by albumin+ cells in contact cocultures compared to transwell cocultures (Fig. 3C).
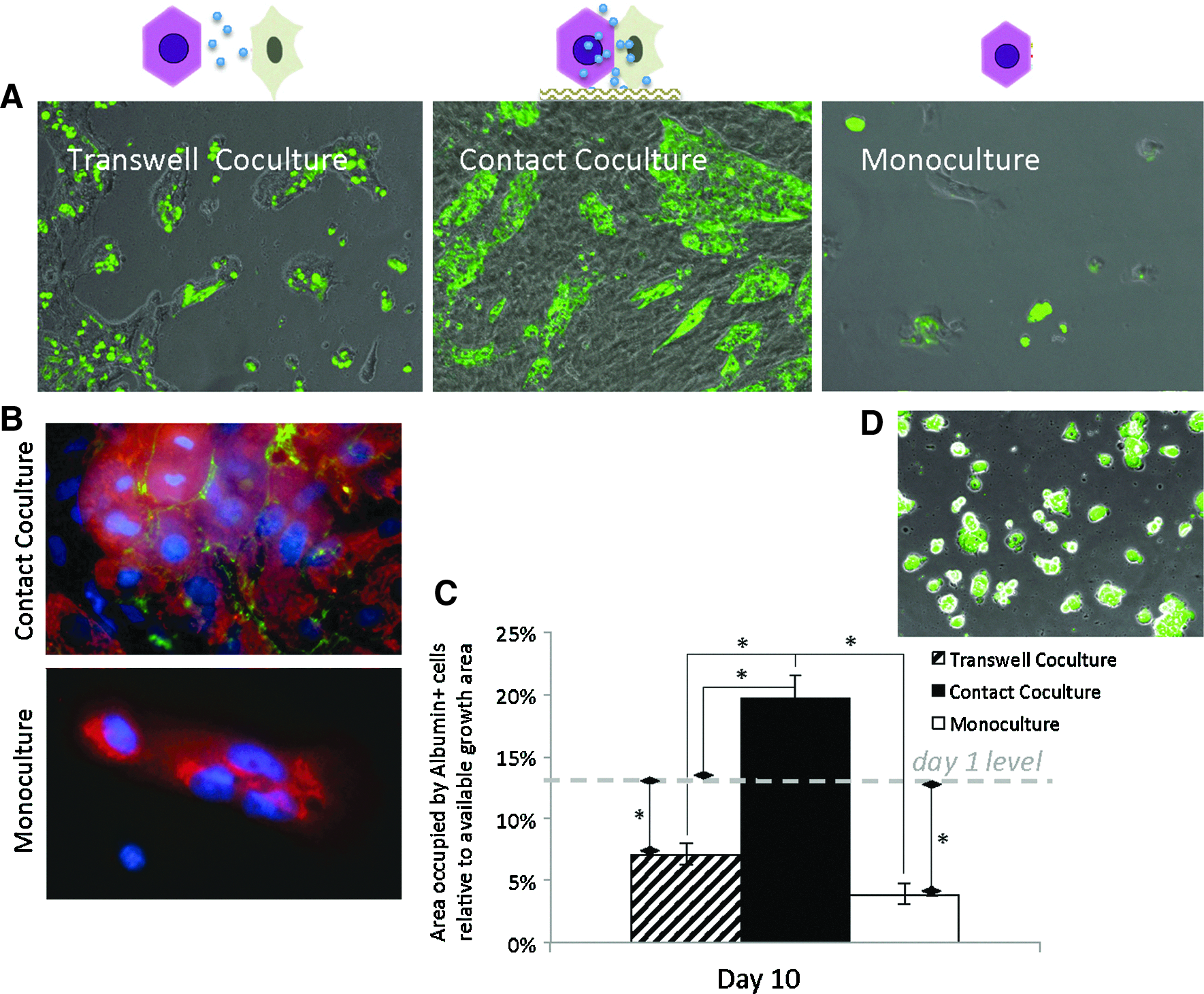
Changes in the morphology and mature hepatocyte mass during culture.
Soluble factor effects
In transwell cocultures, HUCPVC-soluble factors rescued hepatocyte albumin secretion for a transitory period, but by day 10, albumin levels were at the same level as those in monoculture (Fig. 1A). While there was no overall improvement in ureagenesis in transwell coculture, CYP activity was significantly higher than both monoculture and contact coculture (Fig. 1E). At the morphological level, we observed that HUCPVC-soluble factors maintained a higher hepatocyte number and induced the formation of colonies that covered a larger area than those seen in monoculture, but these were significantly lower than those of either contact coculture or the initial numbers at day 1 (p<0.01) (Fig. 3C). Furthermore, by day 10, not all hepatocytes were expressing albumin (Fig. 3A).
Since we saw some benefits from the soluble factors directly released from HUCPVCs, we explored the effects of the HUCPVC CM on hepatocytes. The use of CM would offer two main advantages: the possibility of controlling the concentration of soluble factors and also the possibility of identifying the agents responsible for the effects. Thus, we tested two concentrations of CM, 5×and 1×. Improved albumin secretion during the initial stages of hepatocyte culture up to day 5 was shown by both 1×and 5×CM. The former had a small, but significant, improvement between day 3 and 5, and the latter had a larger effect that lasted until day 6 (Fig. 4A). Although no improvements in the albumin+ cell coverage were seen by day 10, 5×CM preserved bigger colonies of cells than 1×CM or monoculture, indicating that proliferation had been induced (Fig. 4C). No other benefits were seen with 1×CM or 5×CM compared to monoculture, while there was a negative effect on CYP activity that was directly proportional to the CM concentration (Fig. 4E).
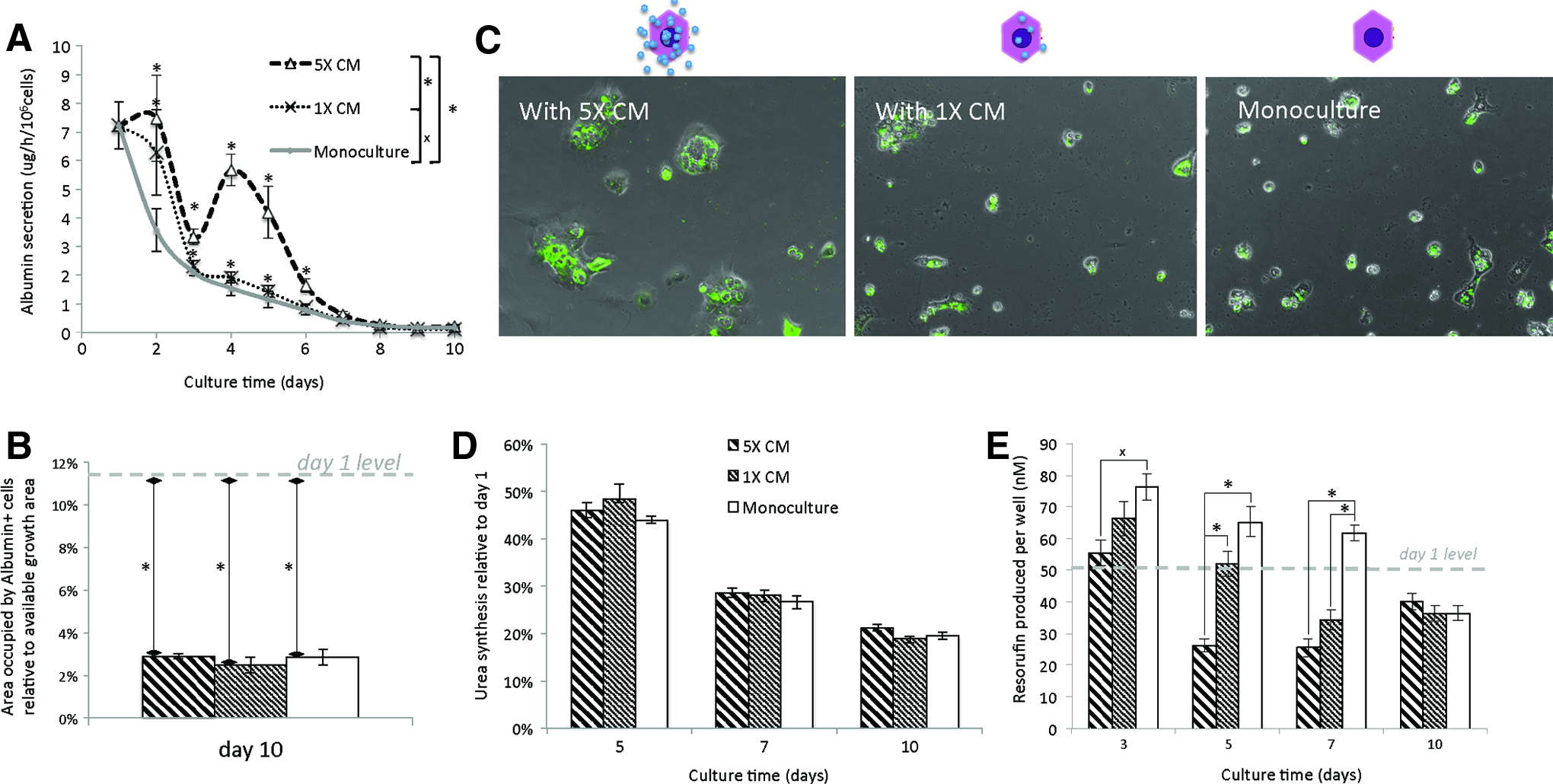
Hepatocytes cultured with HUCPVC CM (5×, 1×). Hepatocytes cultured with 5×HUCPVC CM=thick hatched bar; hepatocytes cultured with 1×HUCPVC CM=thin hatched bar; and monoculture=white bar.
Nonsoluble factor effects
To examine the effect of nonsoluble factors, ECM and cell–cell membrane interactions, to the exclusion of soluble factors, we used confluent flash-frozen HUCPVCs. The viability of flash-frozen cells was tested using the fluorescein diacetate assay, and no viable cells (fluorescent) were found on day 1 of culture nor day 15 (data not shown), indicating that the flash freezing successfully killed all the HUCPVCs, while SEM showed that the cell membranes and ECM were present (Fig. 5A). Interestingly, while the albumin secretion levels of hepatocytes were not initially affected, but nevertheless decreased faster than in monoculture, this secretory function later recovered, and continued increasing for the whole culture period (Fig. 5B). In addition, ureagenesis and the CYP activity remained higher than that seen in monoculture at the times measured, and the expression of albumin, TO, and TAT mRNAs on day 10 was significantly higher than monoculture (Fig. 5C, D, E). In this condition, there was significantly higher albumin+ cell coverage than seen in monoculture, although still lower than that seen on day 1 (Fig. 5F), the levels were similar to those seen in transwell cocultures. However, the latter exhibited larger colonies, but with not all cells expressing albumin, while hepatocytes cultured on flash-frozen HUCPVCs had smaller colonies expressing albumin homogenously (Figs. 3A and 5F).

Nonsoluble factor model: hepatocytes cultured on flash-frozen HUCPVCs.
HUCPVC number is proportional to the effect on hepatocytes
The number of HUCPVCs in contact with hepatocytes is correlated with the level of maintenance of hepatocytes. Hepatocyte albumin secretion and ureagenesis were slightly improved in ML conditions compared to monoculture, but this effect was increased in MH conditions (Fig. 6A, B). CYP activity was increased significantly in MH conditions, to the level of normal HUCPVC coculture (Fig. 6C). ML failed to increase CYP activity as much as the MH condition, showing an improvement only by day 10. In addition, the area covered by albumin+ cells was higher in MH conditions compared to ML, and both of them were higher than monoculture, but did not reach day 1 levels. Only normal HUCPVCs at an initial seeding density of 1×104 HUCPVC/cm2 were again able to facilitate mature hepatocyte expansion (Fig. 6D).
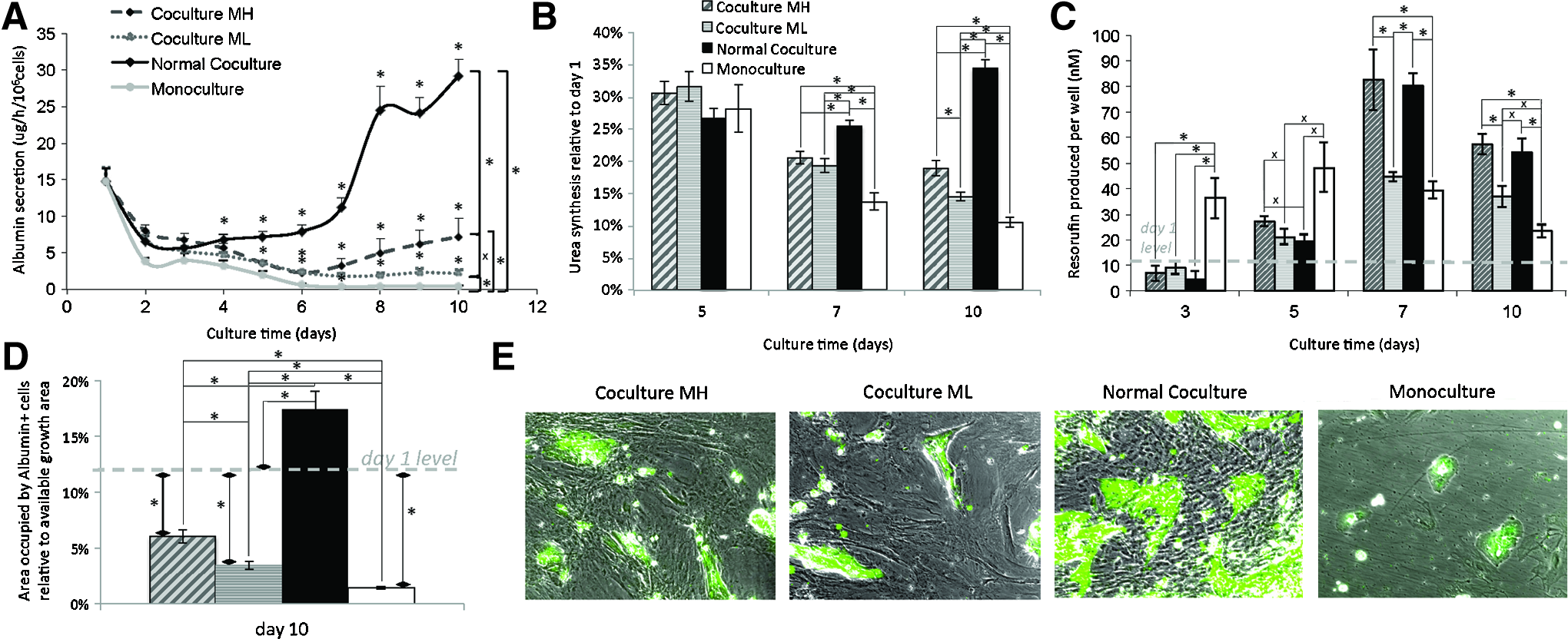
Hepatocytes cocultured with HUCPVCs at an initial seeding density of 1×104 cells/cm2 (normal coculture) and mytomicin-treated HUCPVCs, 2.5×104 cells/cm2 (coculture MH) and 1×104 cells/cm2 (Coculture ML). Where not indicated, coculture MH=gray hatched bar; coculture ML=striped gray bar; normal coculture=solid black bar; and monoculture=white bar.
From Figure 6A and B, it is possible to see that the delay on the effects that HUCPVCs have on hepatocytes is not fully associated with HUCPVC proliferation, as this delay also appears with mytomicin-treated cells.
Discussion
Our results clearly show that HUCPVCs can serve as rat hepatocyte stromal cells; this support involves the action of both soluble and nonsoluble factors, which when independently tested show differential effects. To our knowledge, this is the first report of the hepatotrophic effects of neonatal MSCs. For acute liver failure patients, rapid access to BALS, which require large numbers of allogeneic cells, is essential. Indeed, a BALS may require 10–20 billion hepatocytes to support a patient's failing liver,3,25,26 and if the hepatocyte/MSC ratio were 2:1, then 5–10 billion MSCs would also be required. An advantage of HUCPVCs as an alternative source of MSCs, in this context, is that their CFU-F frequency at harvest is orders of magnitude higher than BM-MSCs; they proliferate faster,14,16 conform with the ISCT definition of an MSC, yet have similar anti-inflammatory 18 and wound-healing 27 properties to BM-MSCs. Proliferative activity may be of greater importance than previously thought, because we found that the HUCPVC/hepatocyte ratio is directly correlated with the strength of their effect; this is in agreement with previous reports where the fibroblast or BM-MSC/hepatocyte ratio has also shown similar trends.28–30 Interestingly, we also observed that proliferating cells had stronger effects than mitomycin growth-arrested HUCPVCs. Since cell motility is not disturbed by the treatment with mitomycin, 31 we hypothesize that either there is a hepatotrophic factor associated with proliferation, or mytomicin independently affects HUCPVCs so that they are not able to provide an optimal support to hepatocytes.
Contact coculture enhances hepatocyte function and permits net proliferation
Since hepatocytes perform a myriad of functions, it is necessary to show multifunctional improvement to demonstrate the efficacy. Previous reports on BM-MSC–hepatocyte cocultures have shown improvement of albumin secretion, ureagenesis, and CYP activity compared to monoculture.28,30,32 HUCPVC–hepatocyte cocultures also improved these functions, both in 2D and 3D configurations, although we also saw differences in the intensity of these effects with a serum batch; for instance, Figure 1A, C and Figure 6A, C show different albumin secretion and CYP activity levels, and the two figures resulted from experiments using the same conditions, but different serum batches.
BM-MSC cocultures have also been shown to improve the maintenance of hepato-specific factors expression,5,28 but this has only been done in allogeneic cocultures negating the possibility of quantifying the actual expression levels. Because our cocultures were xenogeneic, we were able to design species-specific primers to quantify the expression of hepato-specific genes without any interference from the HUCPVCs employed.
Additionally, using albumin+ cell fluorescence as a surrogate for mature hepatocyte mass, we were able to show that HUCPVCs were able to facilitate proliferation while restricting hepatocyte dedifferentiation in contact cocultures. Other authors have reported that BM-MSCs promote the proliferation of hepatocytes,5,28 but to our knowledge, this has not previously been correlated with an increase in mature hepatocyte mass. Thus, for the first time, we show that an MSC is able to both increase the initial hepatocyte mass and preserve the differentiated state of hepatocytes.
Both soluble and nonsoluble factors contribute to hepatocyte function
Our results show that HUCPVCs express several hepatotrophic factors that have mitogenic, supportive, and protective properties. 6 The factors that we detected included cytokines, ECM, and membrane proteins, indicating that HUCPVCs interacted with hepatocytes through both soluble and nonsoluble mechanisms. Soluble factors were able to affect albumin secretion, but for a shorter time than seen in contact coculture, while nonsoluble factors had a delayed effect. Furthermore, nonsoluble factors improved the maintenance of ureagenesis, and hepato-specific factors had a higher expression by day 10 compared to monoculture, or even cultures with HUCPVC-soluble factors. Other groups have also shown that soluble factors have a positive effect on albumin secretion.32,33 Bhandari et al. 33 reported that hepatocytes cultured with the 3T3-fibroblast CM had an earlier increase in albumin secretion, but the duration was shorter than that of coculture or nonsoluble factors, which also had a delayed effect on albumin secretion. However, in contrast to our findings, the CYP activity was not affected by either soluble or nonsoluble factors independently, although it is interesting to note that Bhandari et al. only tested soluble factors by using CM, and in those conditions, we saw a negative effect. It is possible that the HUCPVC-secreted cytokines that stimulate CYP activity are labile while the inhibitory ones are more stable. Thus, the CM would have less stimulatory factors than transwell cocultures, and would cause a net negative effect. It is noteworthy that we observed a high increase in CYP activity levels upon hepatocyte stimulation with soluble (transwell) and nonsoluble factors independently, and that 3D cocultures performed better than those in 2D in this respect. The reason why this would happen is not yet understood. It could be due to an antagonistic interaction between soluble and nonsoluble factors or a contact interaction, such as gap-junction interactions, which would be impaired in our nonsoluble factor model. The fact that this impairment was not manifested in a 3D coculture might be due to the improvement that 3D environments provide to the hepatocyte phenotype. 34
It is important to note that despite reports on the positive effects of soluble factors 6 and the nonsoluble factor Jagged1 35 in hepatocyte proliferation, our study showed that both soluble and nonsoluble stimuli are required to cause a net increase in a mature hepatocyte mass. Nevertheless, we did observe an increase in the size of hepatocyte-derived colonies in conditions exposed to HUCPVC-soluble factors. However, due to the lack of an evenly distributed albumin expression, on day 10, these cultures failed to show any increase in the mature hepatocyte mass. On the other hand, hepatocytes cultured on flash-frozen HUCPVCs showed small colonies with strong homogeneous albumin expression. These two observations suggest that in our conditions, soluble factors had a strong mitogenic effect while nonsoluble factors preferably preserved the mature hepatocyte phenotype.
Conclusion
In conclusion, HUCPVCs provide rat hepatocytes with required signals for the extended maintenance of their mature phenotype ex vivo. Soluble and nonsoluble factors play differential roles in these interactions.
Footnotes
Acknowledgments
The authors would like to thank Dr. Rahim Moineddin for his advice on statistics, and also, Ms. Katie Chan for her technical expertise with hepatocyte isolation. A.G.A. was financially supported by the Harron Scholarship, Faculty of Dentistry, the University of Toronto, and the Doctoral Completion Award, the University of Toronto. The whole work was supported by a Grant-in-Aid of research from Tissue Regeneration Therapeutics.
Disclosure Statement
A.G.A. declares no conflicts. J.E.D. is the founding president of TRT, the company that supplied the HUCPVCs.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.