
Abstract
Biomaterials to be used as cell delivery systems for cardiac tissue engineering should be able to comply with cardiac muscle contractile activity, while favoring cell survival and neo-angiogenesis in a hostile environment. Biocompatible synthetic materials can be tailored to mimic cardiac tissue three-dimensional organization in the micro- and nanoscales. Nonetheless, they usually display mechanical properties that are far from those of the native myocardium and thus could affect host cell survival and activity. In the present investigation, inert poly-ɛ-caprolactone planar layers were manufactured to change the surface stiffness (with Young's modulus ranging from 1 to 133 MPa) without changing matrix chemistry. These substrates were challenged with neonatal murine cardiomyocytes to study the possible effect of substrate stiffness on such cell behavior without changing biological cues. Interestingly, softer substrates (0.91±0.08 and 1.53±0.16 MPa) were found to harbor mostly mature cardiomyocytes having assembled sarcomeres, as shown by the expression of alpha actinin and myosin heavy chain in typical striations and the upregulation of sarcomeric actin mRNA. On the other hand, a preferential expression of immature cardiac cell genes (Nkx-2.5) and proteins (GATA-4) in cardiac cells grown onto stiffer materials (49.67±2.56 and 133.23±8.67 MPa) was detected. This result could not be ascribed to significant differences in cell adhesion or proliferation induced by the substrates, but to the stabilization of cardiomyocyte differentiated phenotype induced by softer layers. In fact, cardiac cell electromechanical coupling was shown to be more organized on softer surfaces, as highlighted by connexin 43 distribution. Moreover, a differential regulation of genes involved in extracellular matrix remodeling was detected on soft films (0.91±0.08 MPa) as compared with the stiffest (133.23±8.67 MPa). Finally, the upregulation of a number of genes involved in inflammatory processes was detected when the stiffest polymer is used. These events highlight the differences in cell mechanosensitivity in a heterogeneous cell preparation and are likely to contribute to the differences encountered in cardiac cell phenotype induced by substrate stiffness.
Introduction
The effects of substrate elasticity on cardiac cells have been addressed by few research groups so far. In embryonic cardiomyocytes, substrate stiffness was shown to affect contractility, with tissue-like Young's modulus appearing more suitable for beating activity; 16 consistently, neonatal cardiac cell maturation was demonstrated to be impaired on stiff substrates, while the formation of functional striations in skeletal myotubes was found to be optimal on substrates having a tissue-like stiffness (12 kPa). 17 Nonetheless, the process of cell fusion leading to the formation of myotubes did not seem to be affected by substrate mechanical properties. 18 In vivo, the slight variations in substrate stiffness following scar formation after myocardial infarction 19 have been proposed to impair injected or resident stem cell ability to be retained in and repair the injured myocardium,20–22 while marked differences in extracellular matrix rigidity are considered predictive of solid tumor malignancy or progression.23–25 The ability of living cells to activate specific signaling pathways in response to mechanical stimuli has been recently demonstrated to be highly cell specific,4,18,26 this evidence being likely to reflect differences in in vivo cell physiology. Given the ability of cells to sense the substrate they grow on and their acknowledged capacity to convert such “feelings” in a biological behavior, one cannot neglect the possibility that scaffolds developed for a specific in vivo application could interfere with host cell behavior once they are implanted. In the case of scaffolds to be used in cardiac tissue engineering, they should be able to comply with cardiac contractility, 27 while also representing an easy tool to be handled by the surgeons. Although natural and synthetic hydrogels represent valuable candidates to mimic cardiac tissue microenvironment in terms of mechanophysical properties, 28 the use of three-dimensional (3D) scaffolds allows the control of graft shape and size, as well as the possibility to use the correct number of cells.29–31 Nonetheless, the possibility that a foreign body reaction could be induced cannot be excluded when synthetic or animal-derived materials are used.32,33
A number of natural biomaterials have been so far proposed as scaffolds to treat cardiac pathologies, including alginate, fibrin, collagen, and chitosan.34–37 Also, synthetic derivatives of poly-lactic acid, poly-ɛ-caprolactone (PCL), and blends obtained with glycolic acid (PLGA) have been investigated as cell delivery systems for future applications. As a bulk, such materials are known to be far stiffer than myocardium, while microfabrication techniques can be used to obtain an anisotropic distribution of their rigidity.4,38,39 Aim of the present work is to study the interaction between stiff substrates (from 0.91±0.08 to 133.23±8.67 MPa) and neonatal cardiomyocytes. For this purpose, planar PCL layers displaying different overall stiffness values were obtained by cross-linking tetra-branched PCL with acrylate end-groups in the presence of linear PCL telechelic diacrylates, and tested for their ability to sustain murine neonatal cardiomyocyte adhesion and maturation. Because PCL is a semicrystalline polymer that has a melting temperature (Tm) over which the mobility of polymer chains changes significantly, the cross-linked PCLs successfully offer branch-structure-dependent stiffness without changing the surface wettability. Our results show that softer materials (around 1 MPa stiffness) favor the appearance of cells having correctly oriented sarcomere structure and electromechanical coupling, while the stiffer ones (in the high MPa range) enhance the number of GATA-4- and Nkx-2.5-expressing noncontractile cells, having poor electromechanical coupling and reduced contractile activity. These results could not be ascribed to a differential regulation of cardiac cell adhesion or proliferation by substrate stiffness. Finally, an overall effect of substrate stiffness on cardiac cell gene expression is demonstrated, with genes involved in cell–matrix interaction being upregulated on the softest substrate, while those related to inflammatory response being significantly activated on the stiffest.
Materials and Methods
PCL film preparation and stiffness control
Figure 1 shows a schematic illustration of the PCL materials prepared by cross-linking tetra-branched PCL with acrylate end-groups in the presence of linear PCL telechelicdiacrylates, according to a previously reported protocol. 40 Briefly, two-branched and four-branched PCL were synthesized by ɛ-caprolactone ring-opening polymerization that was initiated with tetramethylene glycol and pentaerythritol as initiators, respectively. Then, acryloyl chloride was reacted to the end of the branched chains. The structures and the molecular weights were estimated by 1H NMR spectroscopy (JEOL) and gel permeation chromatography (Jasco International). The average degrees of polymerization of each branch on two-branched and four-branched PCL were 18 and 10, respectively. The obtained PCL macromonomers were then dissolved in xylene containing benzoyl peroxide, and the solution was injected between a glass slide with a 0.2-mm-thick Teflon spacer to prepare the substrate layers. The PCL macromonomers were cured for 180 min at 80°C. The mechanical properties of the cross-linked materials were characterized by a tensile test (EZ-S 500N; Shimadzu). The contact angles on the PCL layers were also determined by a sessile drop method. They were measured 30 s after a water drop was placed on the surface at 37°C.

Schematic illustration of preparation of cross-linked PCLs. Layers with various stiffness were obtained by simply mixing and curing two- and four-branched PCL with acrylate end-groups. PCL macromonomers were dissolved in xylene containing BPO and cured for 180 min at 80°C, and the solution was injected between a glass slide with a 0.2-mm-thick Teflon spacer to obtain planar layers. BPO, benzoyl peroxide; PCL, poly-ɛ-caprolactone. Color images available online at www.liebertpub.com/tea
Murine neonatal cardiomyocyte extraction and characterization
The study was performed on neonatal mice under the protocols approved by the institutional Animal Care and Use Committee of the University of Rome Tor Vergata (Rome, Italy). Cardiac cells were extracted from 1/3-day-old neonatal CD1 murine hearts (n=30) by means of Trypsin enzymatic digestion (0.05% Trypsin in a solution of 0.02% EDTA in phosphate buffer saline [PBS]) for 3 h at 4°C, followed by incubation in collagenase II (1500 U; Worthington Biochemical Corporation) for 30 min at 37°C. After filtration through a 0.70-μm strainer (Falcon BD), the samples were collected and centrifuged for 5 min at 800 g. Cells were resuspended in Claycomb medium (Sigma-Aldrich) supplemented with 10% fetal bovine serum (FBS; Lonza Group Ltd.), 100 IU/mL penicillin, and 4 mM L-glutamine (Sigma-Aldrich) and seeded onto 2 μg/mL fibronectin (Biochrom AG)/0.2% laminin in 0.02% gelatin (Sigma-Aldrich) precoated tissue culture polystyrene (TCPS) dishes and on PCL planar scaffolds. Cardiac cells were analyzed immediately after the extraction as described in Immunofluorescence staining and confocal microscopy section. The number of beating areas was calculated by counting 10 random fields under light microscope and the mean values obtained from three independent experiments were reported.
Immunofluorescence staining and confocal microscopy
Cells seeded on TCPS dishes or on PCL layers were fixed with 4% paraformaldehyde in PBS for 30 min at 4°C, and permeabilized with 0.1% TritonX-100 (Sigma-Aldrich) in PBS for 2 min at room temperature after 1, 3, and 7 days of culture. Cells were incubated with antibodies against α- actinin (1:100; Sigma-Aldrich), GATA-4 (1:100; Santa Cruz Biotechnology), ki67 (Abcam), and myosin heavy chain (MF20-488; eBioscience) for 1 h at room temperature. The appropriate fluorophore-conjugated secondary antibodies were as follows: Alexa Fluor 488 goat-anti-rabbit and 546 goat-anti-mouse (Invitrogen Corp.). Nuclei were counterstained with 4–6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich). Secondary antibodies in the absence of a specific primary antibody were used to exclude the occurrence of unspecific signals. The images were taken using a Leica DMRB microscope equipped with a digital camera or using a confocal laser scanner microscope (Olympus FV 1000), after excitation at 405, 488, and 543 nm wavelengths for blue, green, and red channel acquisition, respectively. Volume reconstruction (3D rendering) and iso-surface for blue and red channels of xyz confocal image was performed using Imaris software (Bitplane).
Cell circle shape factor (or circularity index) was calculated as 4πA/P2 on acquired images by manually outlining single cardiomyocytes as identified by α-actinin staining (n=30/sample) in ImageJ (National Institute of Health). Cardiomyocyte mean surface area was calculated on α-actinin-positive cells (n=30/sample) by the same method using ImageJ software.
Scanning electron microscopy analysis of the samples
Cells grown for 7 days on PCL substrates were fixed in 4% PFA for 15 min at room temperature, dried under flow laminar hood, and prepared for scanning electron microscopy analysis as previously reported 41 and examined using Hitachi S-4800 low-voltage scanning electron microscope.
RNA extraction, real-time PCR, and PCR array
Total RNA was extracted by TRIZOL Reagent (Gibco BRL). Retrotranscription was carried out with 2 μg of RNA for each sample using reverse transcription (RT) Moloney murine leukemia virus (Invitrogen Corp.) in the presence of random hexamers. Real-time PCR was used to determine the expression profile of 90 key genes involved in different signal transduction pathways in Mus musculus. After synthesis of first-strand cDNAs, RT-PCR (n=3) was performed by the 7500 Real-Time PCR System (Applied Biosystems) using the RT2 Profiler PCR Arrays Mouse Signal Transduction Pathway Finder (PAMM-014; Qiagen) and Mouse Extracellular Matrix and Adhesion Molecules (PAMM-013A) with the RT2 SYBR Green/ROX PCR Master mix (Qiagen) according to the manufacturer's protocol. The threshold cycle (Ct) is defined as the fractional cycle number at which the fluorescence reaches 10-fold standard deviation of the baseline (from cycle 3 to cycle 12). The primers used to amplify cardiac-specific genes are the following: Nkx-2.5 (product size: 128) For: GAGCCTGGTAGGGAAAGAGC, Rev: GAGGGTGGGTGTGAAATCTG; GATA-4 (product size: 78) For: GACACACTGCCTTGTCTGGA, Rev: GCTGTGATCTGGTCTGAGGTC; Connexin 43 (product size: 74) For: AGGATTCAGGGGTAAAGGAAAC, Rev: GGCTGGGTTGGACAGTTAGT; and α-cardiac actin (product size: 183) For: GTGGCTGGCTTCTCCTCTAA, Rev: AACACCTGCTTTCCTTCACAA. The specificity of the SYBR PCR signal was confirmed by melt curve analysis. For the complete list of genes tested, please see Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/tea).
Statistical analysis
The results are given as mean±standard deviation as obtained by counting n fields per group in three independent experiments. The significance of differences in multigroup comparison was evaluated by one-way analysis of variance followed by unpaired Student's t-test. A final value of p<0.05 was considered statistically significant.
Results
Preparation of PCL layers with controlled stiffness
Cross-linked tetra-branched PCL layers with linear PCL content varying between 0 and 100 wt% were successfully synthesized. As shown in Table 1, tensile tests showed compositionally dependent stiffness ranging from ∼0.91 to 133.23 MPa, without changing their surface wettability that determines initial cell adhesion in the following experiments (Supplementary Fig. S1). Since branch number represents a parameter to adjust crystallinity and mechanical properties of the polymer networks, various suitable combinations can be obtained by simply mixing two- and four-branched macromonomers. Although the crystallinity of the cross-linked PCLs increased with increasing the two-branched PCL content, the elastic modulus of the layer with 50 wt% of two-branched PCL macromonomer was slightly smaller than that with 0 wt%. This is because the netpoint density within the cross-linked PCL prepared from four-branched macromonomers only was the highest among the samples in this experiment. Thus, the unit length of the PCL macromonomers and netpoint density within the cross-linked PCL structure are the predominant contributors in controlling thermal and mechanical properties. Therefore, the stiffness could be successfully adjusted on demand by precisely tailoring the nanoarchitecture of semicrystalline PCL, while keeping a cell-compatible surface wettability. The layers displayed stiffness below 1 MPa (0.91±0.08 MPa), or in the MPa range (1.53±0.16, 49.67±2.56, and 133.23±8.67 MPa) and comparable surface wettability (Table 1).
The mechanical properties at 37°C were characterized by a tensile test. The contact angles on the films were determined by a sessile drop method. Compositionally dependent stiffness values were achieved, while keeping the surface wettability constant.
PCL, poly-ɛ-caprolactone.
Substrate stiffness affects neonatal cardiomyocyte maturation but not adhesion
Neonatal murine cardiomyocytes were extracted as previously described 7 and seeded onto TCPS (virtually infinite stiffness) or PCL layers displaying 0.91±0.08, 1.53±0.16, 49.67±2.56, and 133.23±8.67 MPa stiffness. The number of cells expressing early cardiac commitment marker GATA-4 and mature cardiomyocyte-specific contractile proteins alpha actinin (α−actinin) and cardiac myosin heavy chain was evaluated after 1, 3, and 7 days of culture. The presence of α−actinin and myosin in the putative sarcomeric arrangement was considered as a hallmark of differentiated cardiomyocytes (Fig. 2a). No significant differences in the adhesion rate of GATA-4+ cells were noticed 24 h after seeding on the layers having stiffness of 0.91, 1.53, and 49.67 MPa, while a smaller percentage of cells were expressing such marker on the stiffest substrate (133.23 MPa) (Fig. 2b). Nonetheless, after 3 days of culture, the number of GATA-4+ cells increased on the stiffer surfaces, the higher percentage being encountered on the layer with 49.67 MPa stiffness (Fig. 2a). A dramatic decrease in GATA-4+ expression was found on all the surfaces after 7 days, apart from the stiffest substrate, where its expression was stable over the culture. On the other hand, the number of α−actinin+ cells decreased in a timely fashion on all the surfaces tested, as expected for terminally differentiated cells. 7 Nonetheless, although the only significant difference in α−actinin+ cell number after 24 h was detected on the 1.53 MPa surface (Fig. 2c); a larger number of positive cells could be found after 3 and 7 days on the softer substrates (0.91 and 1.53 MPa) as compared with the stiffer substrates (49.67 and 133.23 MPa; TCPS). Similar results were obtained by staining myosin heavy chain (data not shown). A thorough quantification of GATA-4+ and α-actinin+ proliferating cells by ki67 expression was performed on the polymers after 3 days of culture. The results obtained show that GATA-4 undifferentiated cardiac cells retain a marked ability to proliferate as compared with terminally differentiated α−actinin+ cells, also outlining that no preferential proliferative effect is exerted by the substrates on cardiac cells (Fig. 2d). Cardiomyocytes on TCPS surface showed a similar behavior to those on 133.23 MPa substrate; thus, they were excluded from the following experiments.
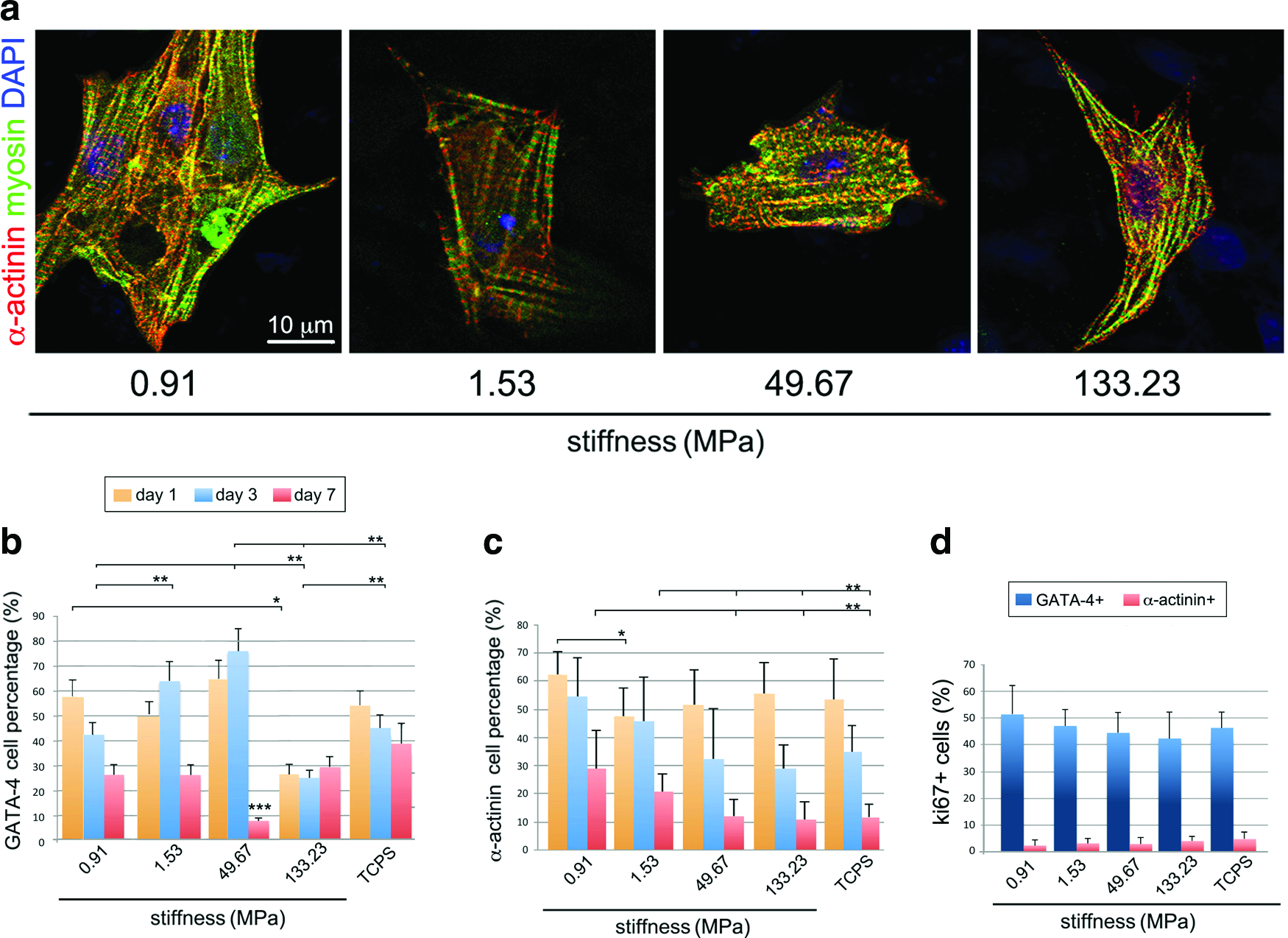
Substrate stiffness affects neonatal cardiomyocyte maturation but not adhesion or proliferation. Murine neonatal cardiomyocytes were cultured onto PCL substrates having 0.91, 1.53, 49.67, and 133.23 MPa stiffness and TCPS (virtually infinite stiffness) for 1, 3, and 7 days. The presence of fully differentiated cardiomyocytes within the preparations was assessed by co-staining alpha actinin (red) and myosin heavy chain (myosin, green) in the typical striations
Cardiomyocyte morphology, sarcomere organization, and electromechanical coupling are impaired in long-term culture on stiff substrates
As expected, the presence of cells expressing the properly assembled form of sarcomeric protein α−actinin was detected on all the layers tested after 3 days of culture, although they appeared to be more scattered on the stiff substrates as compared with the softer substrates, where they were preferentially arranged in clusters (Fig. 3a). At the same time point, real-time PCR performed using primers specific for cardiac markers showed that alpha sarcomeric actin (α sarcomeric actin) expression was enhanced by 1.8-fold on the softest substrate (0.91 MPa), while cardiomyoblast marker GATA-4 was upregulated by 1.3-fold on the stiffest (133.23 MPa). A significant downregulation of another marker of cardiomyoblasts, Nkx-2.5, was found on the softest substrate, while only modest changes were detected for the gene encoding for connexin 43 (Fig. 3b). Nonetheless, the contractile apparatus was found disrupted in cardiomyocytes cultured for longer periods (7 days) onto the stiffer surfaces (49.67 and 133.23 MPa), while its pattern was preserved on the softer substrates (0.91 and 1.53 MPa) at the same time point (Fig. 4a and Supplementary Fig. S2). Additionally, the analysis of cell aspect demonstrated that lower values of circle shape factor and mean surface area (smaller cells with higher aspect ratio) are encountered in cells growing on softer substrates, while those adhering to the stiffer matrix have a higher mean values (bigger cells with low aspect ratio; Fig. 4c). At the same time point (7 days), the analysis of connexin 43 distribution within the cells, accounting for electromechanical coupling in fully differentiated cardiomyocytes, demonstrated that a fair membrane distribution could be found on cardiac cells grown onto the softer substrates (0.91 and 1.53 MPa) (Fig. 5a), while it was randomly distributed on the stiffer substrates. The correct distribution of connexin 43 on the softest substrate was also confirmed by 3D rendering at the confocal microscope (Fig. 5a, bottom and Supplementary Video S1). The same analysis allowed demonstrating functional α−actinin assembly (Fig. 5b) and vinculin expression in cardiomyocytes grown on the softer substrates (Fig. 5c). Cardiomyocyte beating activity was also assessed on the substrates having different overall stiffness, and the beating areas were quantified by light microscopy. The number of beating areas in the time frame considered (1, 3, 4, and 7 days) was found to be significantly higher on the softer substrates (0.91 MPa), while the lowest number of beating areas could be found on the stiffest substrates (133.23 MPa) (Fig. 5d and Supplementary Videos S2 and S3).

Neonatal cardiomyocytes express coherent sarcomere banding in short-term culture on substrates with different stiffness values. Neonatal cardiomyocytes cultured for 3 days onto substrates having stiffness below 1 MPa (0.91 MPa) or in the MPa range (1.53, 49.63, and 133.23, MPa) display sarcomere banding, as highlighted by α-actinin staining (red). Nuclei are counterstained in blue (DAPI)
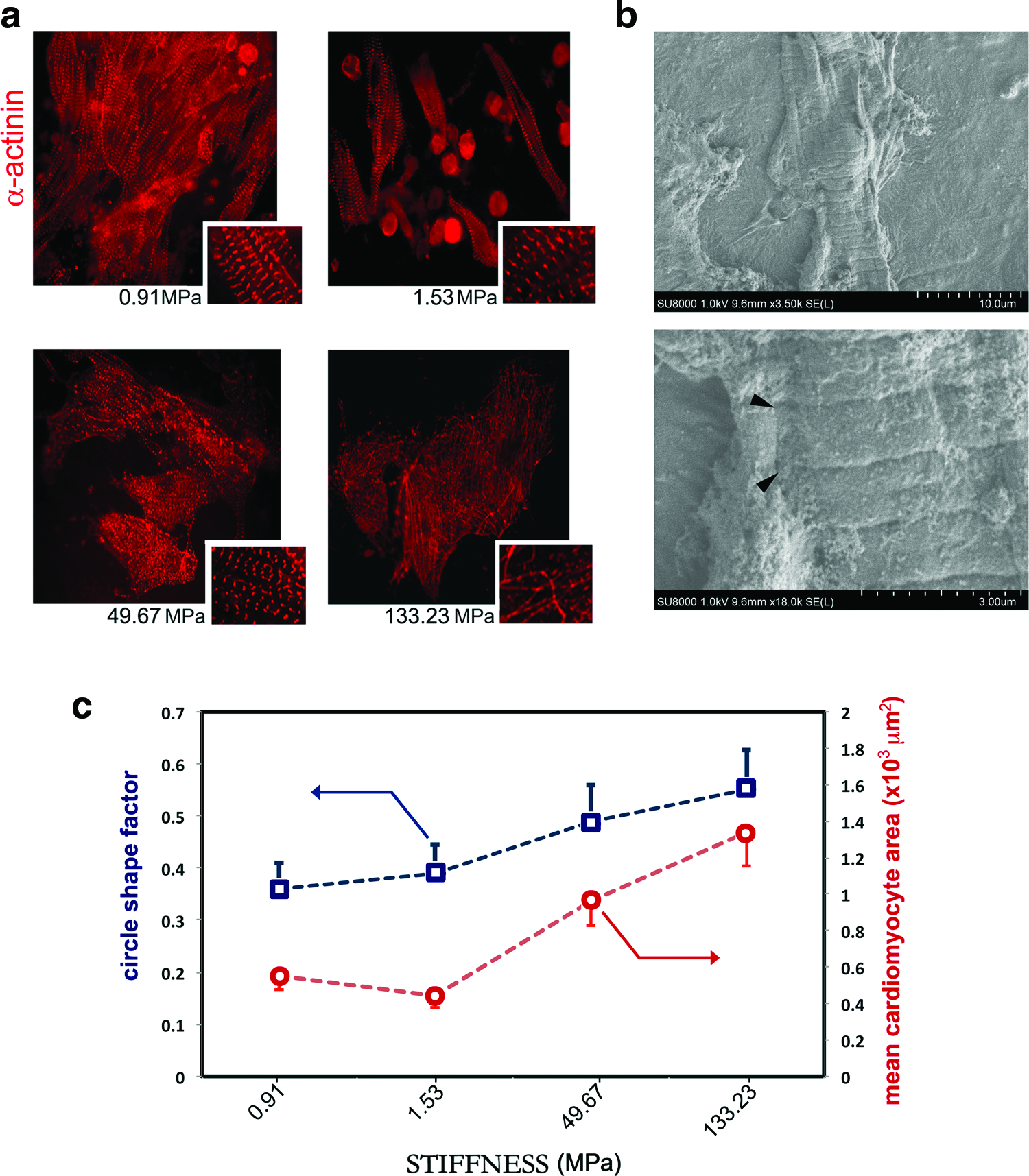
Long-term culture of neonatal cardiomyocytes discloses differences in cell response to substrate stiffness. Sarcomeric actinin (α-actinin, red) staining of neonatal cardiac cells grown for 7 days onto substrates having different stiffness values shows that the contractile apparatus is perturbed in cells growing onto stiff matrix (133.23 MPa), while on softer substrates (0.9 and 1.53 MPa) sarcomeric striations are still well organized
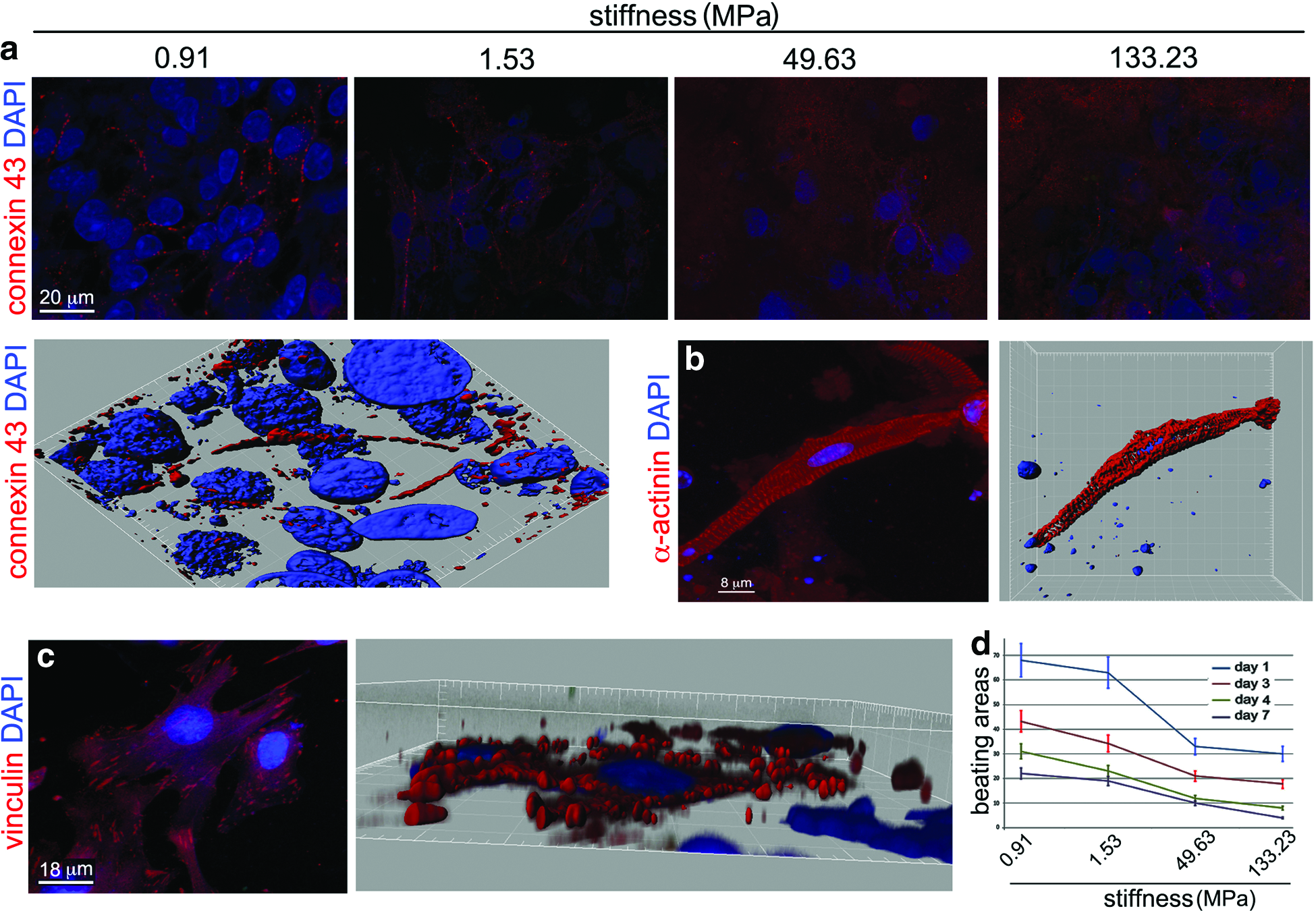
Substrate stiffness perturbs cardiomyocyte electro-mechanical coupling. Connexin 43 staining (red), accounting for cell-to-cell interaction and electromechanical coupling in cardiomyocytes, appears functionally expressed on cell membrane in cardiomyocytes grown onto softer (0.91 and 1.53 MPa) substrates for 7 days, while its expression is scattered in those cultured onto stiffer (49.63 and 133.23 MPa) substrates at the same time point
Cardiac cell gene expression is regulated by substrate stiffness
A thorough analysis of gene expression in cardiac cells grown on PCL layers was performed by analyzing 168 genes involved in different signaling pathways and in extracellular matrix formation by PCR array. On this purpose, cardiomyocytes were seeded on 0.91 and 133.23 MPa substrates for 3 days and total RNA was extracted. Ex vivo extracted cardiomyocytes were used as controls (data not shown). Supplementary Figure 3 illustrates a number of genes differentially regulated in cells grown on the films, grouped per function and/or pathway (for complete gene array information, please refer to Supplementary Table S1). Moreover, Figure 6 shows the results of a bioinformatics analysis of gene expression, as obtained by the software GENEMANIA (www.genemania.org). The network in Figure 6a demonstrates that the genes upregulated on 0.91 MPa stiffness are mostly related to cell adhesion (laminins) and extracellular matrix remodeling (MMPs and TIMPs). On the contrary, Figure 6b shows that mostly genes involved in the regulation of inflammation response are significantly upregulated on the stiffest substrate (133.23 MPa).
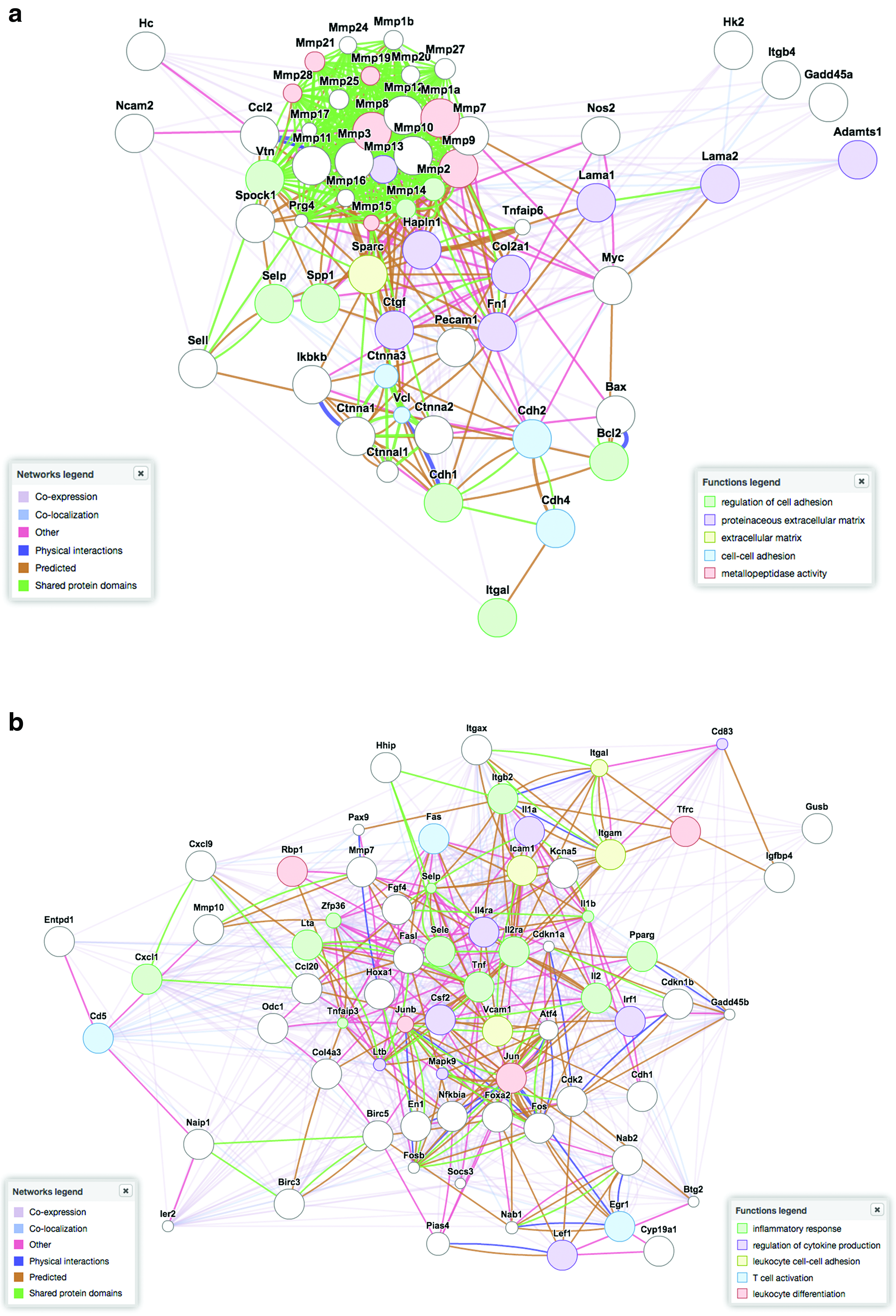
Gene expression regulation by substrate stiffness in murine neonatal cardiomyocytes. Computational gene analysis obtained by plotting in GENEMANIA software (www.genemania.org) shows the results of two PCR array sets containing 186 genes of interest in cardiac cells grown for 3 days onto PCL layers having 0.91
Discussion
The interaction of scaffolds and cells has been so far considered mainly in terms of cell adhesion, proliferation, and differentiation, 42 while only in the last few years the possibility that synthetic substrates—designed to provide implanted cells with a favorable microenvironment—could integrate or interfere with cell function in vivo has been acknowledged. 43 In fact, at the basis of tissue engineering paradigm lays the idea that a scaffold to be used in tissue regeneration should favor new tissue formation without interfering with organ function. 27 Nonetheless, scaffold's physical and mechanical properties have been shown to affect cell survival, 2 proliferation, 44 migration, 5 and eventually differentiation.3,4 These parameters have been regarded to act cooperatively with biological cues to maintain cell phenotype and function 45 or direct stem cell commitment. 7 Recently, evidence has been given that stem and mature cells can sense the stiffness of the substrate they grow on and perceive this parameter as physiological when it matches tissue-specific values.3,16 In this context, the indication that embryonic quail and chicken cardiomyocyte beating activity is preserved on scaffolds displaying cardiac-like rigidity appears interesting. 16 Additionally, neonatal rat ventricular cardiomyocytes have been shown to proceed to cell maturation and produce higher mechanical force on matrices having myocardial-like stiffness (10 kPa), 17 while higher values (144 kPa) led to a reduced troponin I staining and poor electrical excitability. Such experiments were justified by the evidence that the local Young's modulus for adult rat heart tissue ranges from 11.9 to 46.2 kPa (25.6±15.9 kPa), 46 while infarcted myocardium stiffness has been reported to be around 55±15 kPa in Lewis rats. 19
The use of synthetic scaffolds having elastomeric properties, being able to comply with the contractile activity of the organ, has been suggested for cardiac tissue repair.30,47 While synthetic polymers can be easily manufactured to tailor their 3D structure in the nano- and microscale to better reproduce cardiac anisotropy, 4 as a bulk they usually display a Young's modulus that is far larger than that measured for body tissues. Among them, PCL is an elastomeric polyester already approved for biomedical applications by Food and Drug Administration, and already proven to be suitable for cell culture. 38 Moreover, its longer degradation time as compared with poly-lactide makes it appealing for tissue engineering applications.48,49
Thus, in the present investigation, elastomeric PCL substrates having different Young's modulus values (0.91, 1.53, 49.67, and 133.23 MPa) were used to culture neonatal murine cardiac cells and systematically investigate the effects of substrate stiffness on cardiomyocyte phenotype. Our data show that the expression of a number of genes can be significantly altered by substrate stiffness in cardiac cells. In particular, genes involved in cell–matrix interaction (lama1, lama2, fn1, vtn, integrins, etc.) and extracellular matrix remodeling (MMPs and TIMPs) were found upregulated on the softest substrate (0.91 MPa) as compared with the stiffest substrate (133.23 MPa). On the other hand, among the genes found upregulated on the latter surface, the presence of a number of those involved in immune response control (IL1a, IL2, IL2ra, tnf, fas, fasl, etc.) and inflammatory cell recruitment was detected. These data suggest that in a heterogeneous preparation of cardiac cells obtained from neonatal murine heart, in which cardiomyocytes, cardiac fibroblasts, endothelial, smooth muscle, and inflammatory cells are present, those cells can display a cell-specific response to matrix mechanics. Such a concept was already proposed by a number of independent investigations, showing that not all the cells are mechanosensitive and among those that are able to feel substrate mechanical properties, specific responses can be triggered.3,4,9,26 Accordingly, the response of heterogeneous cell preparations to substrate mechanophysical properties is likely to be far more complex than what is thought,3,16 with different cell subsets displaying rather different sensitivity. In this context, the evidence that stiff substrates, in the absence of biological cues, can provoke a raise in immune response appears in good agreement with previous reports showing that the compliance of the microenvironment can interfere with neutrophil migration during the chemotactic process,50,51 even though our result was obtained using a preparation that is supposed to contain a very limited percentage of inflammatory cells.
As compared with freshly extracted cells, a significant upregulation of genes involved in cell survival, like tert, Nos2, and bax, occurred when cardiac cells are cultured for 3 days on TCPS dishes, while a concomitant downregulation of genes responsible for cell proliferation (jun, fos, cdk2, cdkn1b, and egr-1) was detected in cultured cells in comparison to ex vivo cells. Such a gene pattern can be hardly ascribed to a single event (i.e., ischemia, hypertrophy, angiogenesis, etc.),52,53 but it could be compatible with cardiac cell adaptation to the new in vitro conditions. Also, differences in cell morphology, phenotype, sarcomere structure, and function could be found when cardiomyocytes were cultured on substrates with different stiffness values. In fact, a differential expression of GATA-4, Nkx-2.5, alpha actinin (α-actinin), and myosin heavy chain could be detected. While the number of early cardiomyoblast markers GATA-4- and Nkx-2.5 was found to increase with the substrate stiffness (maximum value on 49.67 and 133.23 MPa surfaces after 1 and 3 days), mature α-actinin-positive cells were positively selected after 3 and 7 days on the softer substrates (0.91 and 1.53 MPa), when a physiological decrease in cardiomyocyte percentage is expected. 7 Indeed, more than 85% of differentiated cardiomyocytes were retained on the softer surfaces after 3 days as compared with day 1, while only 60% of them could be found on the stiffer substrates. These effects were not mediated by cell adhesion, since the slight differences encountered after 24 h of culture in GATA-4- and α-actinin-positive cells could not explain such results. Also, these results could not be ascribed to a differential effect of substrate mechanics on cardiac cell proliferation, since no significant differences in proliferating GATA-4- or α-actinin-positive cells could be found on PCL surfaces. Strikingly, a marked difference in the quality of sarcomere structure was noticed at longer time points (3 and 7 days) as an effect of substrate stiffness. In fact, intact sarcomere bands were retained on softer substrates, while their structure was perturbed in cardiomyocytes grown on the stiffer substrates (49.67–133.23 MPa). Moreover, cardiomyocyte cell shape appeared to be different between the softer and the stiffer substrates, as quantified by circle shape factor and connexin 43 staining, with cells grown on the softer substrates being more elongated and displaying correct electromechanical connections. Consistently, the number of contractile areas after 1, 3, 4, and 7 days after seeding was found to be inversely correlated to the substrate stiffness. Altogether, these data suggest that neonatal cardiomyocytes preferentially acquire and retain a mature phenotype on substrates having Young's modulus around 1 MPa, while stiffer surfaces are probably not suitable to sustain their function.
Conclusions
Our data demonstrate that cardiomyocyte phenotype and maturation can be affected by the substrate stiffness and that a higher percentage of mature cardiomyocytes having proper sarcomere assembly, and thus beating activity, can be selected by using substrates having Young's modulus around 1 MPa. Nonetheless, the possibility that distinct cell types respond differently to matrix mechanics should be taken into account when scaffolds for a specific application, like cardiac tissue engineering, are to be prepared.
Footnotes
Acknowledgments
The present work was supported by the Japan Society for the Promotion of Science (JSPS) through the “Funding Program for World-Leading Innovative R&D on Science and Technology (FIRST Program),” the World Premier International (WPI) Research Center Initiative and by the “Nanotechnology Network Project” of the Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan. The authors would like to thank Shoko Kajiwara and the Centre of Advanced Microscopy (CAM), Department of Biology, University of Rome Tor Vergata, for the skillful assistance.
Disclosure Statement
The authors have no competing financial interests to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.