
Abstract
Intestinal myofibroblasts secrete substances that control organogenesis and wound repair of the intestine. The myofibroblasts of the rat small intestine express reelin and the present work explores whether reelin regulates crypt–villus unit homeostasis using normal mice and mice with the reelin gene disrupted (reeler). The results reveal that mouse small intestine expresses reelin, its receptors apolipoprotein E receptor 2 (ApoER2) and very low-density lipoprotein receptor (VldlR) and the reelin effector protein Disabled-1 (Dab1) and that reelin expression is restricted to myofibroblasts. The absence of reelin significantly reduces epithelial cell proliferation, migration, and apoptosis and the number of Paneth cells. These effects are observed during the suckling, weaning, and adult periods. The number of Goblet cells is increased in the 2-month-old reeler mice. The absence of reelin also expands the extracellular space of the adherens junctions and desmosomes without significantly affecting either the tight-junction structure or the epithelial paracellular permeability. In conclusion, this is the first in vivo work showing that the absence of reelin alters intestinal epithelium homeostasis.
Introduction
Epithelial cell renewal is strictly controlled by cell–cell and cell–extracellular matrix (ECM) interactions. 5 A thin and continuous sheet of ECM, the basement membrane (BM), separates epithelial cells from the interstitial connective tissue and its composition defines the necessary microenvironment required for multiple cellular functions during development and at maturity. Reciprocal interactions between the epithelium and the underlying BM regulate proliferation, migration, differentiation, apoptosis, morphogenesis, tissue repair, inflammation, and the immune response. 6 Numerous receptors for ECM molecules have been identified in the intestinal epithelial cells, many of which are integrins. 7 However the nature of cell–BM interactions and their intracellular processing remains largely undefined. Within the ECM, the myofibroblasts, located beneath the epithelia, express and secrete various ECM components, such as cytokines, growth factors, chemokines, hormones, neurotransmitters, inflammatory mediators, and adhesion proteins, as well as express receptors for many of these ligands, allowing information flow in both directions, to and from the intestinal epithelium and the ECM.6,8 As a consequence, the myofibroblasts are regarded as a cell that orchestrates functions that ranged from control of epithelial renewal processes to peripheral immune tolerance.6,8
We reported that: (1) the mucosa of rat small intestine expresses reelin, its receptors apolipoprotein E receptor 2 (ApoER2) and the very low-density lipoprotein receptor (VldlR), and its effector protein Disabled-1 (Dab1) and (2) within the intestinal mucosa, reelin expression was restricted to myofibroblasts. 9 In brain, the reelin secreted by the Cajal–Retzius cells is critical for the positioning of migrating neurons during the development of the central nervous system. 10 Because differentiation of the intestinal epithelial cells requires their migration along the crypt–villus axis, we reasoned that the reelin released by the myofibroblasts to the ECM might regulate epithelial dynamics. 9
The present work explores in vivo whether reelin is involved in the crypt–villus unit homeostasis. For that, we have examined the consequences of reelin gene disruption on cell proliferation, migration, differentiation, and apoptosis in the epithelium of mice small intestine. A preliminary report of some of these results was published as an abstract. 11
Methods
[14C]-Polyethylene glycol-4000 ([14C]-PEG-4000) was purchased from GE Healthcare. Anti-reelin antibodies were obtained from Santa Cruz Biotechnology, Inc. (sc-5578) and Calbiochem (553731); the anti-cleaved Caspase-3 (Asp175) antibody from Cell Signaling and the anti-E-Cadherin antibody from the BD Transduction Laboratories (610181). Unless otherwise stated, the other reagents were obtained from Sigma-Aldrich.
Animals
Reeler (rl) is a spontaneous autosomal recessive mutation. 10 Heterozygous (rl+/rl−) mice C57BL/6 were purchased from Jackson Laboratories through Charles River Laboratories. Control (rl+/rl+) and homozygous mutant reeler (rl−/rl−) mice were obtained by heterozygous crossings. Fifteen- and 21-day-old and 1-, 2-, 3-, and 18-month-old mice were used. The animals were housed in a 12:12 light–dark cycle and fed ad libitum with the Global 2019 extruded rodent diet (Harlan Ibérica S.L.) with free access to tap water. Mice were genotyped by polymerase chain reaction (PCR) analysis of genomic DNA using the primers (5′-3′) TAATCTGTCCTCACTCTGCC, CAGTTGACATACCTTAAT, and TGCATTAATGTGCAGTGT.
The mice were humanely handled and sacrificed, by cervical dislocation, in accordance with the European Council legislation 86/609/EEC concerning the protection of experimental animals.
Relative quantification of real-time reverse transcription-PCR
Total RNA was extracted from the small intestine using the RNeasy® kit (Qiagen). cDNA was synthesized from 1 μg of total RNA using the QuantiTect® reverse transcription kit (Qiagen) as described by the manufacturer. Real-time reverse transcription (RT)-PCR was performed with iQ™SYBR® Green Supermix (BioRad), 0.4-μM primers, and 1 μL cDNA. Controls were carried out without cDNA. Amplification was run in a MiniOpticon™ System (BioRad) thermal cycler (94°C/3 min; 35 cycles of 94°C/40 s, 58°C/40 s and 72°C/40 s, and 72°C/2 min). Following amplification, a melting curve analysis was done by heating the reactions from 65°C to 95°C in 1°C intervals, while monitoring fluorescence. Analysis confirmed a single PCR product at the predicted melting temperature. Primers for the genes tested are given in Table 1. β-Actin served as the reference gene and was used for samples normalization. The cycle at which each sample crossed a fluorescence threshold (Ct) was determined and the triplicate values for each cDNA were averaged. Analyses of PCR were done using the comparative Ct method with the Gene Expression Macro software supplied by BioRad.
Oligonucleotides were chosen according to the cDNA sequences entered in GenBank and designed using PerlPrimer program v1.1.14.
Dab1, Disabled-1; VldlR, very low-density lipoprotein receptor.
Isolation of enterocytes and myofibroblats
Enterocytes and mucosal myofibroblasts enriched fractions were isolated as described. 9 Briefly, the small intestine was rapidly removed and washed with an ice-cold saline solution. One centimeter intestinal segments were incubated at room temperature in the phosphate-buffered saline (PBS) buffer containing 1 mM dithiothreitol for 15 min, followed by a 30-min incubation period at 37°C in a calcium- and magnesium-free PBS buffer containing 1 mM EDTA and 2 mM glucose. Following incubation, the tissues were vortexed for 30 s, the loosened epithelial cells (enterocytes) were filtered through 60-μm nylon textile, and collected by centrifugation and resuspension in PBS. For myofibroblats isolation, the remaining tissue was rinsed in PBS and incubated for 30 min, in a shaking water bath at 37°C, in PBS containing 1 mg mL−1 collagenase and 2 mg mL−1 hyaluronidase. The pieces were vortexed for 30 s and the myofibroblasts were pelleted and resuspended in PBS. The isolated cells were immediately used for immunostaining.
Immunostaining analysis
Enterocytes, myofibroblasts, and intact small intestine were used for immunostaining studies. Immunostaining assays were performed on intact small intestine (either 10-μm cryosections or 5-μm paraffin sections) and on isolated intestinal cells, as previously. 9 The slides containing either the intestinal sections or the isolated cells were incubated with the indicated primary antibody at 4°C, overnight. Antibody binding was visualized with biotinylated anti IgG antibodies, at dilution 1:100, followed by immunoperoxidase staining. The Vectastain ABC peroxidase kit (Vector) and 3,3′-diaminobenzidine were used for the tissue cryosections and the 3-amino-9 ethylcarbazole for paraffin sections. Controls were carried out without a primary antibody. The slides were rinsed, mounted, and photographed with a Zeiss Axioskop 40 microscope equipped with a SPOT Insight V 3.5 digital camera. Acquired images were analyzed by using Spot Advance 3.5.4.1 program (Diagnostic Instrument, Inc.).
Western assays
SDS-PAGE was performed on a 15% polyacrylamide gel as described. 12 For protein extraction, the small intestine was homogenized in a buffer containing in mM, 150 NaCl, 50 Tris/HCl, 1 EDTA, pH 7.4, 0.01 aprotinin, 0.01 leupeptin and 0.2 phenylmethylsulfonylfluoride and 1% Triton X-100. Following the 10-min incubation, the suspension was centrifuged at 14,000 g for 30 min. The resultant supernatant was dissolved in the Laemmli sample buffer, electrophoresed, and electrotransferred onto a nitrocellulose membrane (Micron Separations, Inc.). A total of 50 μg protein was loaded to each lane. The protein was measured by the Bradford method using gamma globulin as the standard. The immunoreactive bands were viewed using a chemiluminescence procedure (GE Healthcare). The anti-β-actin antibody (1:1000 dilution) was used to normalize band density values. The relative abundance of the bands was quantified using PCBAS program version 2.0 (Raytest).
Crypt cell proliferation rate
The epithelial cell proliferation rate was quantified by measuring the incorporation into DNA of 5-bromo deoxyuridine (BrdU). Mice received an intraperitoneal injection of BrdU (120 mg/kg body weight) and were killed 90 min later. BrdU was detected by immunohistochemistry using a monoclonal anti-BrdU antibody, 1:300 dilution, as described above. The number of labeled cells in 30 crypts well-oriented longitudinally per mouse was determined by light microscopy. The crypt cell proliferation rate is expressed as the number of BrdU-positive cells per crypt.
Epithelial cell migration rate
Mice were killed 32 h after the intraperitoneal injection of BrdU (120 mg/kg). Small intestinal sections were processed for immunoperoxidase staining to detect BrdU-labeled cells as described above. The epithelial cell migration rate was estimated by measuring the distance from the base of the villus to the foremost-labeled cell at 32 h after injection. Thirty villi well-oriented longitudinally per mouse were used. The cell migration rate is expressed as μm/h.
Analysis of cell apoptosis
The identification and quantification of apoptotic cells in the epithelium of the small intestine were performed by immunological detection (Western blot and immunohistochemistry assays) of cleaved Caspase-3. A polyclonal anti-cleaved Caspase-3 antibody was used at dilution 1:1000 for the Westerns and at dilution 1:200 for immunohistochemistry. The number of labeled cells in 100 well-oriented longitudinally villi per mouse was determined by light microscopy. Results are expressed as the number of labeled cells per 10 villi.
Goblet cells staining
The identification and quantification of Goblet cells in the epithelium of the small intestine were performed using Periodic Acid-Shiff (PAS) Staining System as indicated by the manufacturer. The number of labeled cells in 30 villi well-oriented longitudinally per mouse was determined and the results are expressed as the number of Goblet cells per villus.
Paneth cells staining
Small intestine Paneth cells were quantified by phloxine-tartrazine histological stain. Briefly, 10-μm cryosections of small intestine were incubated with the hematoxylin solution (Gill No. 3) for 3 s, followed by a 2-min incubation in a 70% ethanol solution containing in mM, 6 phloxine and 34 CaCl2. After being washed with tap water, they were placed in a solution containing 9.4 mM tartrazine in 2-ethoxy-ethanol for 1.5 h, and thereafter they were washed with 2-ethoxy-ethanol. The number of labeled cells in 30 crypts well-oriented longitudinally per mouse was determined and the results are given as the number of Paneth cells per crypt.
Electron microscopy assays
Segments of small intestine were fixed in 4% glutaraldehyde/0.1 M sodium cacodylate, pH 7.4 at 4°C for 3 h. After three rinses in the cacodylate-buffered solution, the tissues were postfixed in 1% OsO4 in 0.1 M phosphate buffer at 4°C for 1 h and washed in the cacodylate-buffered solution containing 7.5% sucrose. The segments were then dehydrated in a graduated series of acetone (30%, 50%, and 70%), stained with 2% uranyl acetate and embedded in Spurr's epoxy resin. Ultrathin sections were examined either under a Philips CM-10 or a Zeiss Libra 120 transmission electron microscopes equipped with an Olympus Veleta or a TRS 2K Slow-scam CCD camera, respectively. The photographs were processed with iTEM software.
Assessment of intestinal permeability to [14C]-PEG-4000
Intestinal permeability to [14C]-PEG-4000 was measured using everted intestinal sacs, according to Wilson and Wiseman. 13 Briefly, the small intestine was dissected, rinsed twice with ice-cold saline solution, turned inside out, and cut into two to three parts. Each part was closed at one end using a cotton thread. Each sac was filled through the other end with the modified Krebs–Ringer bicarbonate-phosphate buffer containing in mM, 140 NaCl, 10 KHCO3, 3 K2HPO4, 1.2 CaCl2, and 1.2 MgCl2, pH 7.4 and closed. The sacs were incubated, at 37°C in a thermostatic bath with continuous shaking, in modified Krebs–Ringer bicarbonate-phosphate buffer, pH 7.4, containing [14C]-PEG-4000 (1.2 μM), that was continuously bubbled with 95%O2/5%CO2. After a 5-min incubation period, the everted sacs were rinsed with the ice-cold saline solution, opened, and the serosal fluid collected. The sacs were weighed wet before and after the incubation period. Fifty microliters samples were taken from the bathing solution and from the serosal fluid for radioactivity counting. The sacs were opened longitudinally and the serosal surface area was evaluated by measuring the length and width of the open sac. The amount of [14C]-PEG-4000 in the serosal side was referred to the intestinal surface area.
Statistical analysis
Data are presented as mean±SEM. Unless stated otherwise, the number of animals (n) used in each experimental condition was three control and three reeler mice. In the Figures, the vertical bars that represent the SEM are absent when they are less than symbol height. In Figures 1 and 9, comparisons between different experimental groups were evaluated by the two-tailed Student's t-test. One-way ANOVA followed by the Newman-Keuls' test was used for multiple comparisons (GraphPad Prism program). Differences were set to be significant for p<0.05.
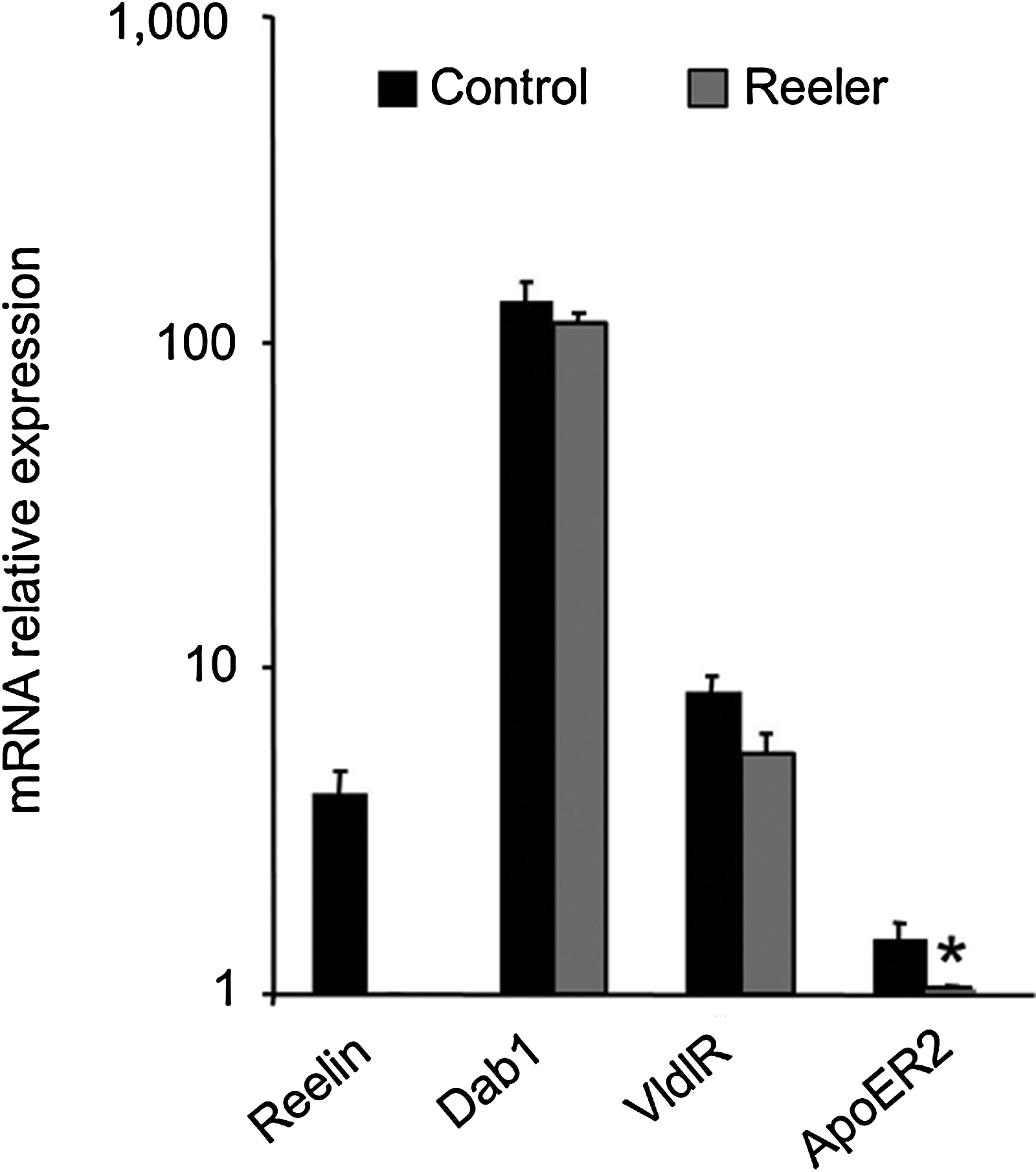
Abundance of reelin, Disabled-1 (Dab1), apolipoprotein E receptor 2 (ApoER2) and very low-density lipoprotein receptor (VldlR) mRNAs in the small intestine of control and reeler mice. Reverse transcription-polymerase chain reaction was performed using total RNA isolated from the small intestine of 1-month-old mice. The values represent the means±SEM of relative units of mRNA abundance on a semilogarithmic scale. The ApoER2 mRNA abundance measured in the reeler mice was set at one. Three reeler and three control mice were used. Student's t-test: *p<0.05 reeler versus control mice.
Results
Expression of reelin, Dab1, VldlR, and ApoER2 in the mice small intestine
The experiments were performed to find out whether murine small intestine expresses reelin, its effector protein Dab1, and its receptors VldlR and ApoER2. Figure 1 shows the relative abundance of reelin, Dab1, VldlR, and ApoER2 mRNAs in the intestine of control and reeler mice. As expected, the small intestine of the reeler mice lacks reelin mRNA. The mRNA levels of Dab1 and VldlR were similar in both types of mice, but the mutation significantly decreases the abundance of ApoER2 mRNA.
As myofibroblasts express α-smooth muscle actin (α-SMA), we have used the anti-α-SMA antibody to their localization in the mucosa of mice intestine. The localization of α-SMA and reelin proteins in mice intestine is shown in Figure 2. In the intestine of control mice, the specific signal produced by the anti-reelin antibody is restricted to a layer of cells located beneath the epithelium. This layer is also reactive to the anti-α-SMA antibody. In the intestine of the reeler mice, the anti-α-SMA antibody also produces a specific signal in the layer of cells located beneath the epithelium, but these cells do not react to the anti-reelin antibody.
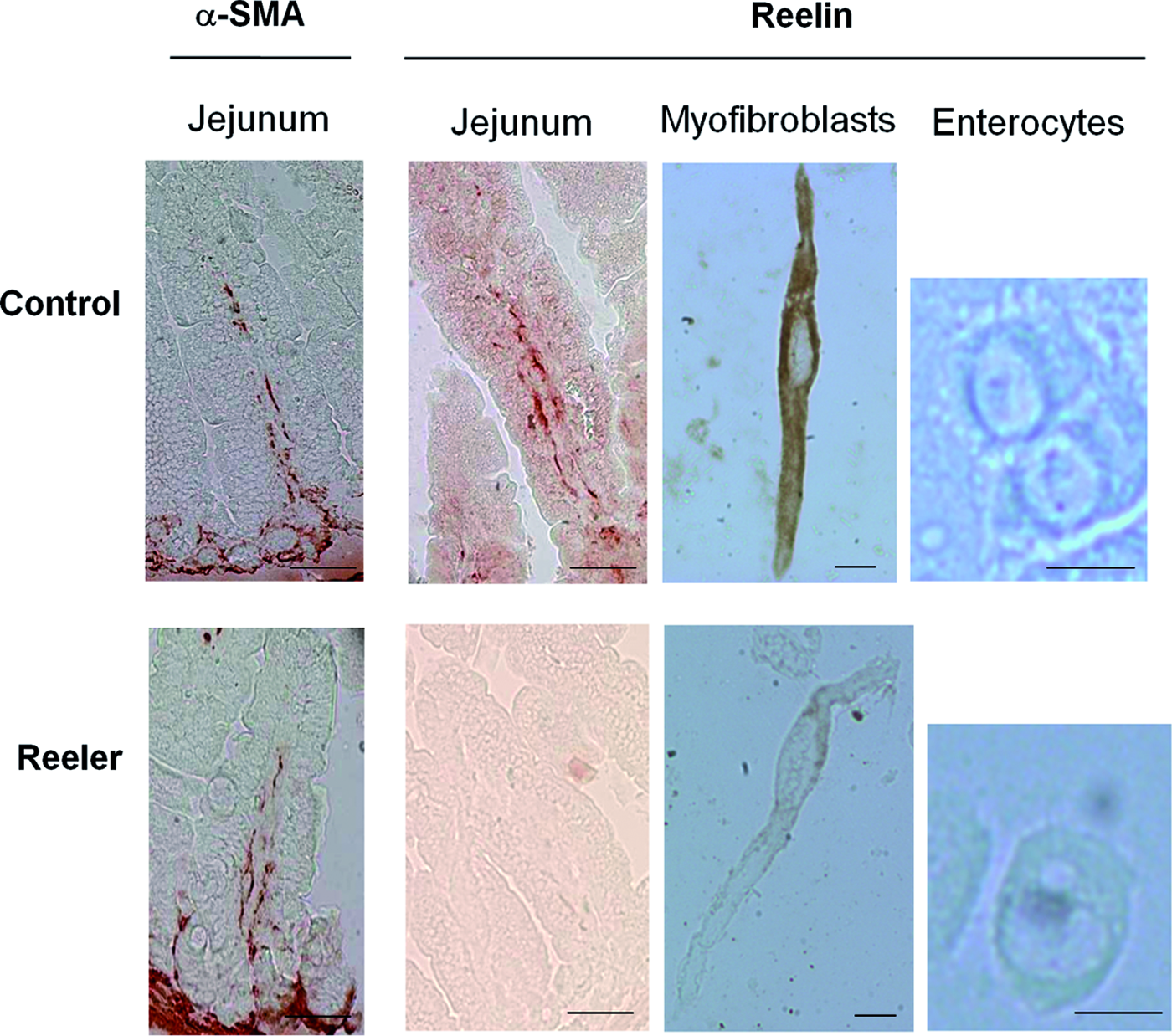
Immunolocalization of α-smooth muscle actin (α-SMA) and reelin in the small intestine of control and reeler mice. Five micrometer paraffin sections of jejunum obtained from 15-day-old mice were incubated either with anti-α-SMA (1:200 dilution) or with anti-reelin (Calbiochem, 1:100 dilution) primary antibodies. Myofibroblasts and enterocytes obtained from 1-month-old mice were incubated with anti-reelin antibody (Santa Cruz, 1:50 dilution). Scale bars represent 50 μm in the intact tissue and 10 μm in the isolated cells. The photographs are representative of three different assays performed on three reeler and three control mice. Color images available online at www.liebertpub.com/tea
The observations made in isolated cells (Fig. 2) corroborate those made in intact tissue. Thus, the myofibroblasts isolated from the small intestinal mucosa of control mice present a strong reelin-specific immunostaining signal. Immunoreactive reelin signal is neither seen in the myofibroblasts of the reeler mice nor in the enterocytes of either type of mouse.
Immunoreactive signal was not seen in the absence of the corresponding primary antibody (data not shown).
These observations together, indicate that reelin is only produced by the myofibroblasts placed underneath the epithelium of mice intestine.
Cell proliferation rate in the small intestinal epithelium of control and reeler mice
To assess the effects of the reeler mutation on epithelial cell proliferation, the incorporation of BrdU into DNA was measured in the epithelium of the small intestine of control and reeler mice of different ages, as explained in the Methods section. The results are given in Figure 3. As expected, BrdU-positive nuclei are only found in the crypt cells and their quantification reveals that in control mice the cell proliferation rate increases during the 2nd month of life, and thereafter it remains fairly constant. In the reeler mice, the epithelial cell proliferation remains fairly constant during the first 2 months of life and it significantly increased thereafter.
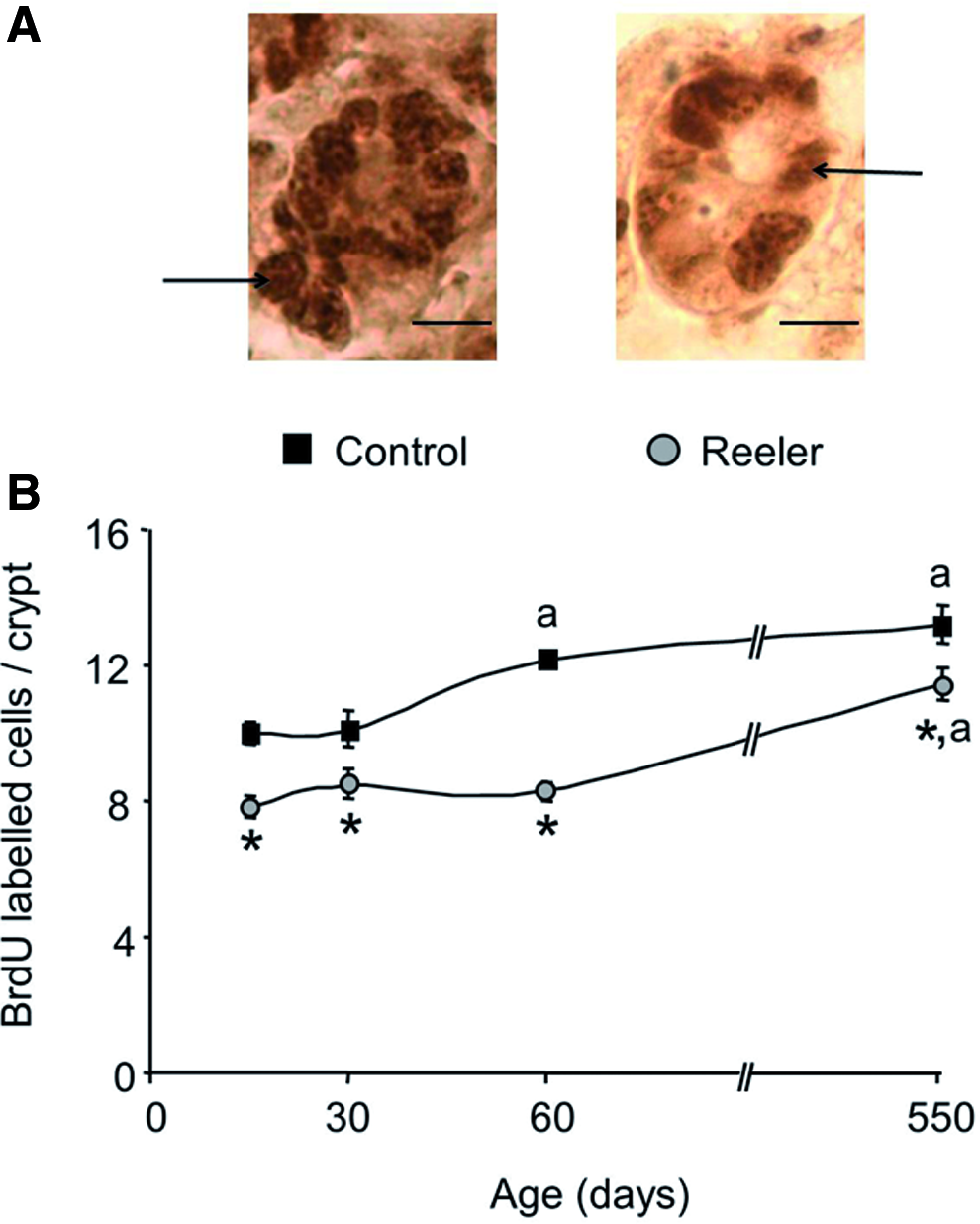
Cell proliferation rate in the intestinal epithelium of control and reeler mice. Incorporation of 5-bromo deoxyuridine (BrdU) into DNA was used as a marker of cell proliferation as described in Methods section. Fifteen- and 21-day-old and 1-, 2-, 3-, and 18-month-old control and reeler mice were used. The number of cells labeled with a monoclonal anti-BrdU antibody (1:300 dilution) was determined by light microscopy in 30 crypts, well-oriented longitudinally, per mouse.
In all the ages examined, the epithelial cell proliferation rate is around 22%±5% lower in the reeler mice as compared with that measured in the control animals.
Cell migration rate in the small intestinal epithelium of control and reeler mice
BrdU incorporation into DNA was also used to evaluate the cell migration rate as described in the Methods section and the results are given in Figure 4. They reveal that the pattern of cell migration rate along the villi is similar in both types of mice. It peaks at 1 month of age and it remains fairly constant in the adult animal.
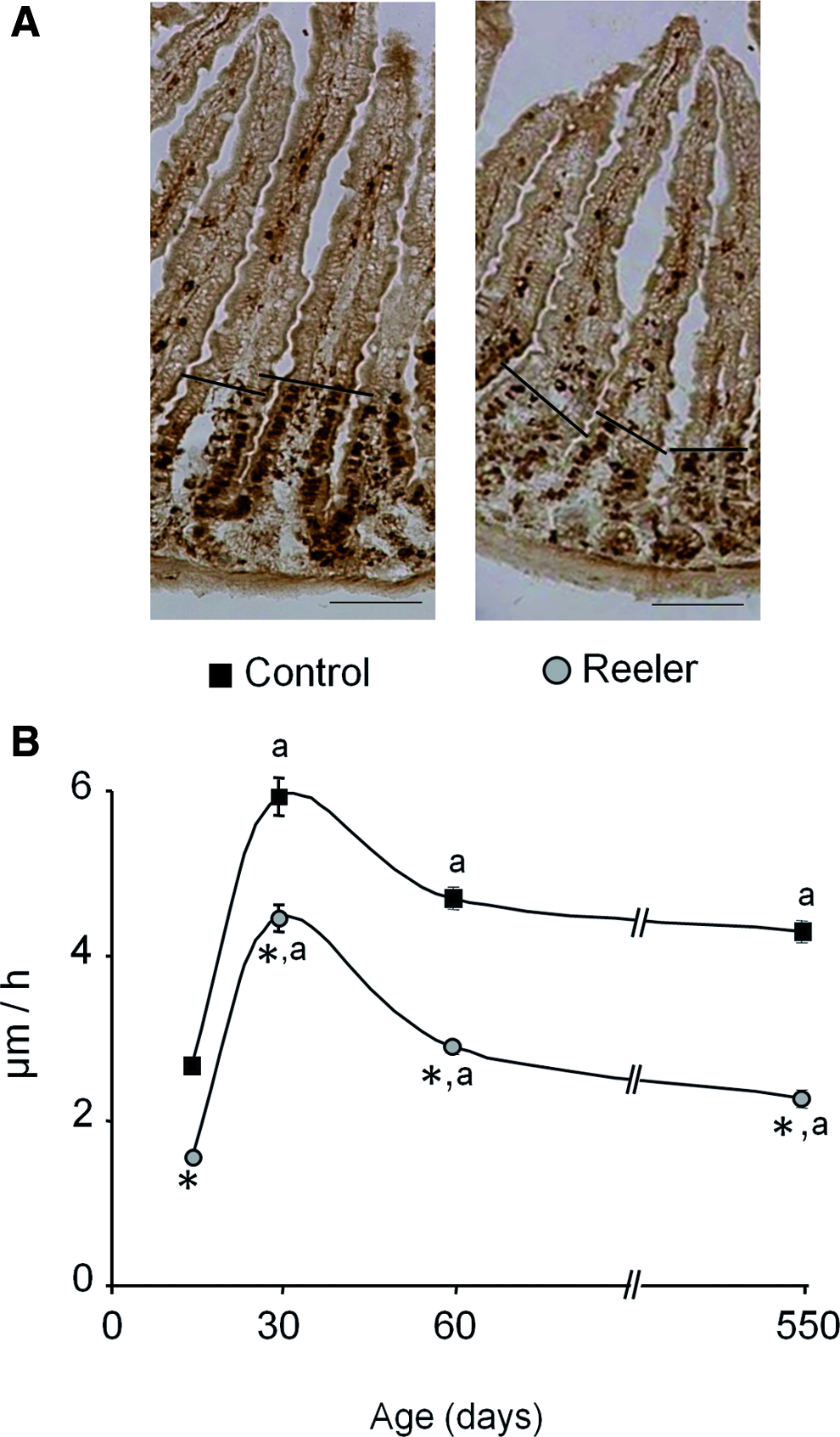
Cell migration rate in the small intestine of control and reeler mice. Mice were injected with BrdU 32 h before death (see Methods section). The distance between the foremost-labeled cells were measured and used to determine the cell migration rate.
In all the ages tested, the cell migration rate measured in the intestinal epithelium of the reeler mice is around 37%±5% lower compared with control mice.
Cell apoptosis in the small intestinal epithelium of control and reeler mice
Cell apoptosis was evaluated by immunological detection of cleaved Caspase-3. The immunohistochemistry assay reveals that in both, control and reeler mice, the apoptotic cells are only observed at the villi (Fig. 5). In all the ages tested, the number of apoptotic cells in reeler mice is around 50%±5% of that measured in the control mice.
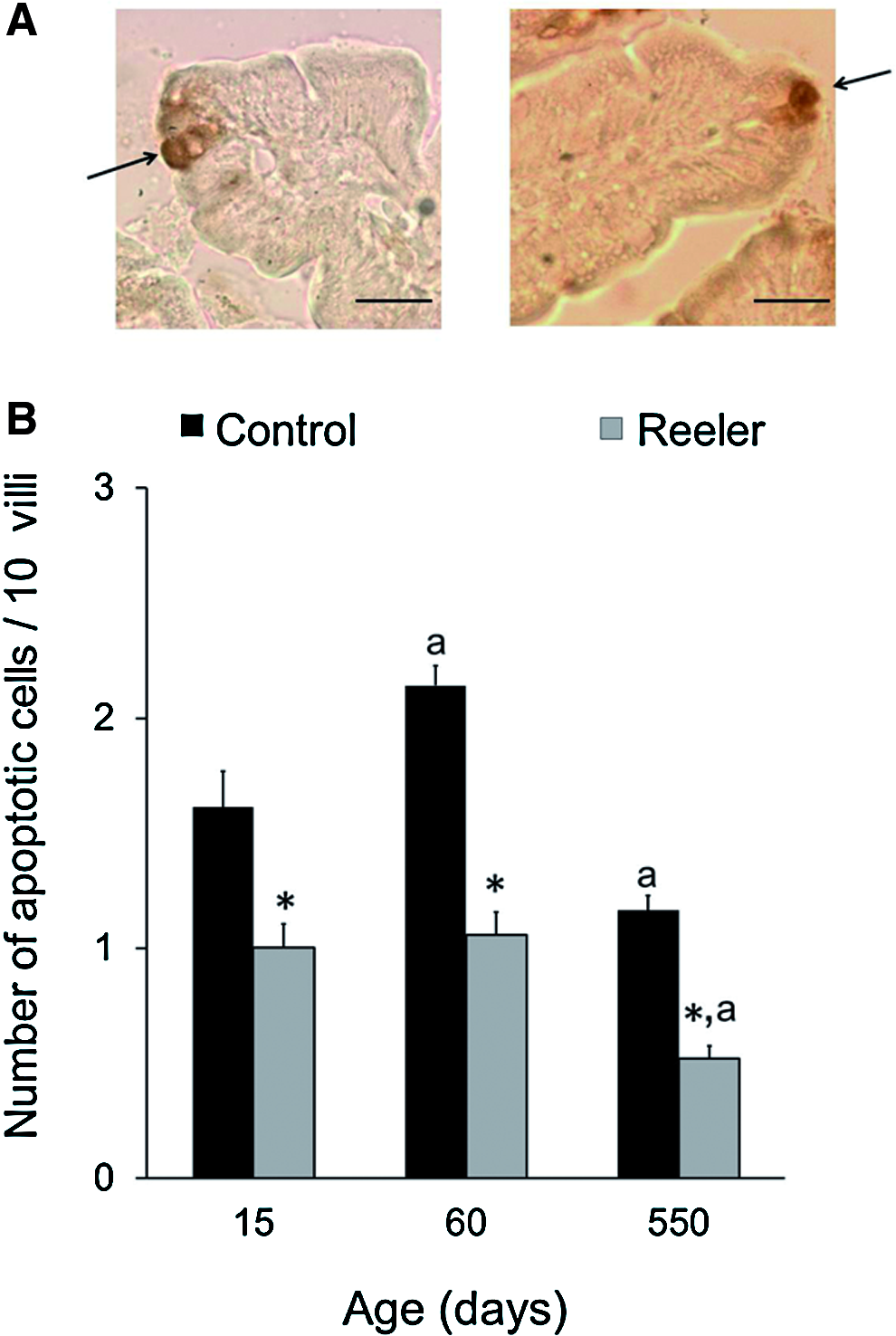
Cell apoptosis in the small intestine of control and reeler mice. Sections of small intestine were incubated with anti-cleaved Caspase-3 antibody, 1:200 dilution, as described in Methods section.
The Westerns assays (Fig. 6) show that the anti-cleaved Caspase-3 antibody detected polypeptides of 17 and 19 kDa, being the former more abundant that the 19-kDa polypeptide. The 17-kDa polypeptide is indicative of cleaved Caspase-3 14 and the mutation significantly decreases its abundance in all the ages tested.

Western blot analysis of intestinal cleaved Caspase-3 in control and reeler mice. Mice that were 0.5, 2, and 18 months old were used. 50 μg protein were loaded to each lane. The blots were probed with the polyclonal anti-cleaved Caspase-3 antibody, 1:1000 dilution, as described in the Methods section. Anti-β-actin antibody was used to normalize the bands. C, control; R, reeler. Histograms represent the means±SEM of the relative abundance of the 17-kDa band. One-way ANOVA showed an effect of the mutation on cell apoptosis (p<0.001). Newman-Keuls' test: *p<0.001 and #p<0.01 reeler versus control mice.
Number of Goblet cells in the small intestinal epithelium of control and reeler mice
Goblet cells were identified using the PAS staining system. The results are given in Figure 7 and reveal that the number of PAS-positive cells in the intestinal epithelium of the control mice peaks at 1 month of age, it decreases at 2 months, and thereafter it increases with age. In the intestine of the reeler mice, the number of Goblet cells increases from the 2nd week of life to 1 month, it remains constant during the following month, and thereafter it increased with age.
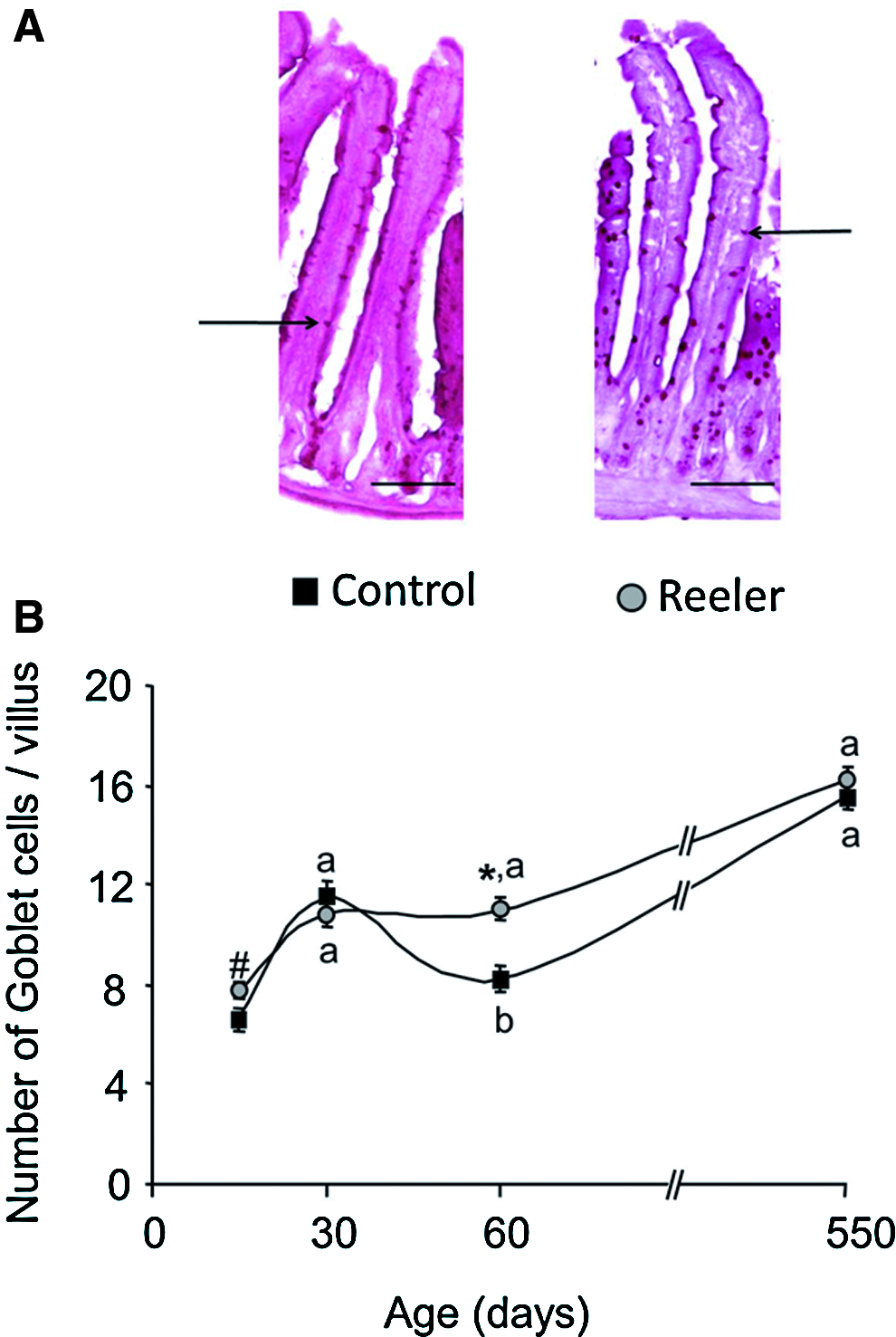
Number of Goblet cells in the small intestine of control and reeler mice. Tissue was stained with Periodic Acid-Shiff Staining System.
Significant differences in the number of Goblet cells between the two types of mice are only observed at 2 months of age: they are lower in the control mice than in the reeler.
Number of Paneth cells in the small intestinal epithelium of control and reeler mice
We identified Paneth cells in the intestinal mucosa of control and reeler mice using phloxine-tartrazine histological stain. Figure 8 shows that in both, reeler and control mice, the number of Paneth cells increases during the first 2 months of life and it slightly decreases at 18 months.

Number of Paneth cells in the small intestine of control and reeler mice. Tissue was stained with phloxine-tartrazine as described in Methods section.
In all the ages tested, the reeler mutation reduces the number of Paneth cells by 30%±1.8% as compared with the values obtained in control mice.
Electron microscopy studies of the small intestine of control and reeler mice
We have investigated whether reelin affects the organization of the cell–cell junctions. The microvilli and cell–cell contacts were examined by electron microscopy (Fig. 9). It is observed that the microvilli are numerous and well formed in both types of mice. No significant differences between the control and reeler mice are found in either the length or the number of microvilli (data not shown).

Electron microscopic images of intestinal apical membrane domain of control and reeler mice. Microvilli and apical junctions of 1-month-old control and reeler mice are shown. Scale bar represents 500 nm. TJ, tight junctions; AJ, adherens junction; D, desmosome. The histogram represents the average width of the junctions (six to eight junctions were examined per type of mouse). The microphotographs are representative of three different assays performed on three reeler and three control mice. Student's t-test: *p<0.001 reeler versus control mice.
The microphotographs of Figure 9 reveal that the tight junctions appear normal, but the intercellular space of both, the adherens junctions and desmosomes is significantly wider in the reeler than in the control mice.
Immunolocalization of E-cadherin in the small intestinal epithelium of control and reeler mice
Immunohistochemical assays reveal that a strong E-cadherin-specific immunostaining signal is located at the lateral membrane of the epithelial cells, in both control and reeler mice (Fig. 10). No significant differences between the two types of mice are observed, indicating that the protein is properly located to the membrane in the intestine of both, control and reeler of mice. Immunoreactive signal was not seen in the absence of the primary antibody (data not shown).

Immunolocalization of E-cadherin in the intestinal epithelium of control and reeler mice. Ten micrometer cryosections of jejunum obtained from 1-month-old mice were incubated with the primary anti-E-cadherin antibody (1:500 dilution). The photographs are representative of three different assays performed on three reeler and three control mice. Color images available online at www.liebertpub.com/tea
Intestinal transport of [14C]-PEG-4000 in control and reeler mice
Mucosal to serosal transfer of [14C]-PEG-4000 was measured to estimate the paracellular permeability of the jejunum and ileum of control and reeler mice. Everted intestinal sacs were used and the age of the animals was 21 days old. The results reveal that the mucosal to serosal transfer of [14C]-PEG-4000 is similar in both types of mice (1.48±0.21 and 1.16±0.34 pmol/cm2 in jejunum and ileum of control mice, respectively, vs. 1.17±0.15 and 1.19±0.06 pmol/cm2 in jejunum and ileum of reeler mice, respectively, n=5).
Discussion
The myofibroblasts placed underneath the epithelium of the small intestine secrete substances that control the epithelial cell renewal as well as intestinal organogenesis and wound repair. 6 We previously proposed that in the rat small intestine the reelin released by the myofibroblasts might regulate the crypt–villus unit homeostasis 9 and the current data support this point of view. The results also show that the epithelial cell renewal processes vary with age, as previously reported for control mice, 15 and that reelin does not modify the pattern of this age-related changes.
As observed in rat, 9 expression of reelin in mice small intestine is restricted to the myofibroblasts placed underneath the epithelium and the following data suggest that reelin regulates crypt–villus unit dynamics. The mutation decreases cell proliferation in the crypts, which suggests that reelin stimulates stem cells. The replication and division of the stem cells in the crypts force the migration of their daughters toward the villi. If reelin promotes proliferation of intestinal stem cells, differences on cell migration speed between the two types of mice should occur, and indeed cell migration along the villi is significantly reduced by the reeler mutation. Slowed migration, however, could also be secondary to increased apoptosis in the crypts: if fewer cells exited the crypt, the rate of movement up the villus would be reduced. However, apoptotic cells are only observed at the villi tip, which agrees with studies showing that apoptosis occurs predominantly at the villus tip. 16 The observations discussed so far might suggest that reelin has a growth-promoting action in the small intestine. In contradiction to this point of view is that no differences in the height of the villi between control and reeler mice were observed, though the villi width was decreased by the mutation. 17 The length of the villi is also determined by apoptosis at the villi and the absence of reelin decreases the number of apoptotic cells in the villi, but this decrease is approximately two times lower than that measured in the cell proliferation rate. The magnitude of the reelin-related changes in the crypt–villus unit cell renewal processes described here is similar to that reported for mice intestine in response to other experimental challenges,18–20 but different results have been found in the literature. As in the current work, some investigators reported diminutions in cell proliferation and migration rates without changes in the villi height,18,21 but others found decreases in cell proliferation and migration that were accompanied by a smaller villi size.22–24 It has been observed that, when they occur together, the changes in cell proliferation and migration rates go in the same direction,18,21–24 but different situations have been described regarding apoptosis: changes in either the same direction than those found in cell proliferation, 25 in the opposite direction 26 or no change.19–21
The current results also suggest that reelin regulates epithelial cell differentiation because its absence decreases the number of Paneth cells. The number of Goblet cells, however, is either unchanged or increased in the 2-month-old reeler mice. The physiological meaning of these observations is not evident, but the different effects of the mutation on the number of Paneth and Goblet cells suggest that the two cell lineages have different requirements for their differentiation. Decreases in the number of Paneth without changes in that of Goblet cells 27 or vice versa 25 have been reported.
Adhesion junctions are vital for epithelial sheets as they maintain tissue integrity and epithelial homeostasis 28 and the reeler mutation significantly expands the extracellular space of the adherens junctions and desmosomes. The mutation does not seem to modify the tight junctions as both, its appearance at the electronic microscopy and the intestinal transport of PEG-4000, a molecule that crosses the intestinal epithelium via the paracellular route, is similar in both types of mice. The adhesive function of the adherens junctions and desmosomes is mediated by the cadherins ectodomain, which engages identical molecules on the surface of the adjacent cell. The expansion of the adherens junctions does not appear to be due to mislocalization of cadherin because the mutation does not modify the cell location of the E-cadherin.
The capacity of reelin to regulate cell proliferation, migration, and differentiation agrees with observations made in neural29–31 and non-neural tissues, such as submandibular 32 and mammary gland. 33 However, inhibition of cell migration in primary cultures of mammary epithelial cells 33 and of apoptosis in embryonic carcinoma cells 34 by exogenous reelin have also been reported. Whereas the information on the cell-signaling cascades initiated by reelin in brain is profuse, the studies on the role of reelin in non-neural tissues are scarce. Due to the cross talk between myofibroblasts and epithelial cells, 6 the intestinal effects of reelin reported here could be triggered by either a direct action of reelin on the epithelial cells or through a reelin-induced change in the myofibroblasts or by both ways. Neither are known the molecular mechanism(s) by which reelin exerts these effects. We have reported that in mice small intestine, the reeler mutation modifies the expression of genes coding for proteins that control epithelial cell renewal processes and cytoskeleton organization. 17 Let us mention the mutation-induced downregulation of Arrb1 (β1-arrestin) and PyCard and the upregulation of bone morphogenetic protein 1 (Bmp1), peroxisome proliferator-activated receptor alpha (Ppara), and Eph receptor A1 (EphA1). The effects of the mutation on Bmp1, Ppara, and β1-arrestin could mediate the mutation-induced decrease in cell proliferation reported here, because: (1) Bmp1 stimulates the BMP pathway, 35 a signaling system that inhibits cell proliferation, 36 (2) Ppara inhibits cell proliferation, 37 and (3) the downregulation of β1-arrestin might reduce the activity of the Wnt system, 38 which is known to stimulate intestinal cell proliferation. 36 The Wnt system also controls the number of Paneth cells 36 and the downregulation of β1-arrestin and the mutation-induced decrease in the number of Paneth cells could be related. The minor intestinal cell migration seen in the reeler mice may, in part, result from the upregulation of the EphA1 receptor because its activation inhibits cell migration in vitro. 39 Finally, the decrease in cell apoptosis observed in the reeler intestine may depend on the downregulation of the proapoptotic PyCard gene. 40 Cell proliferation and migration and the structure of the apical junctions depend on the dynamics of the actin cytoskeleton. In the small intestine, β1-arrestin activates the actin-depolymerizing factor/cofilin. 41 The absence of reelin may inactivate intestinal cofilin via downregulation of β1-arrestin and, hence, modify those cell processes that depend on the actin cytoskeleton. Although in the brain, reelin inactivates cofilin, 42 it is interesting to note that in both, brain and intestine, reelin initiates signaling cascades that affect the same cytosolic protein, cofilin, but we are far from knowing the cell signaling events triggered by reelin in the intestine.
The observations discussed so far reveal that the lack of reelin diminishes the rate of the crypt–villus unit renewal processes, but this intestinal response is opposite to that expected from the effects of the mutation on tissue architecture. Thus, the reelin-related expansion of the junctions might reflect a defective interaction of the cadherin ectodomain and the loss or reduction in E-cadherin function promotes intestinal cell proliferation, migration, and apoptosis43,44 and reduces the number of Goblet cells. 44 And recent studies recognize a role for the desmosomes as sensors of environmental and cellular cues, 45 in tumor suppression 46 and in maintaining the shape and length of actin-rich microvilli. 47 Regarding the latter role of demosomes, neither the length nor the number of microvilli are significantly modified by the reeler mutation. Reelin could overlap functionally with systems that either act identically or perform the same function, but in mechanistically distinct manners. Consequently, a component that is crucial under nonphysiological circumstances can be less relevant under physiological conditions, due to other compensating mechanisms. It is interesting to point out that the gastrointestinal tract, in addition to nutrient absorption, displays endocrine, immunologic, metabolic, and barrier functions and several observations suggest that reelin may protect the intestine from the development of pathologies. (1) The mutation reduces the number of Paneth cells. These cells secrete into the lumen of the crypts a number of antibacterial molecules in response to bacterial antigens, thereby contributing to the maintenance of gastrointestinal barrier. 3 They have also been implicated in the development of inflammatory bowel disease 48 and their location adjacent to stem cells suggests that they play a critical role in defending epithelial cell renewal. 3 (2) The wider extracellular space of the epithelial junctions may favor exposure to external pathogens. (3) The homeostasis of the intestinal epithelium is based on the balance between cell proliferation, differentiation, and apoptosis. The loss of control of these processes leads to cancer 49 and we have observed that reelin expression is decreased in human colon tumors. 50 (4) The absence of reelin reduces the intestinal expression of a large number of genes that code for proteins involved in the intestinal immune response, inflammation, and cancer processes. 17 As several reports have proposed a role for reelin in tissue repair in both, neural31,51–53 and non-neural tissues,32,51,54 its involvement in the activation of cell proliferation and migration during the mucosal regenerative response triggered by injury could not be discarded.
In conclusion, our work in vivo demonstrates for the first time that reelin controls crypt–villus unit homeostasis. The results also support the view that reelin may play a role in the protection of the intestine from pathologies and even in the intestinal repair processes. How reelin exerts its effects and the role of the reelin signaling system in the intestinal physiology remains unknown.
Footnotes
Acknowledgments
This work was supported by a grant from the Junta de Andalucía (CTS 5884) and by a fellowship from the Spanish Ministerio de Educación y Ciencia (AP2007-04201) to MD Vázquez-Carretero. We thank Dr. O. Pintado from the Centro de Producción y Experimentación Animal of University of Sevilla for his technical advice. Electronmicroscopy images were obtained in the Centro de Investigación, Tecnología e Innovación, Universidad de Sevilla and in the Departamento de Histología y Anatomía patológica, Universidad de Navarra.
Disclosure Statement
No competing financial interests exist.