
Abstract
Articular cartilage contains three functional zones (superficial, middle, and deep) characterized by distinct structure, composition, and biomechanical properties. One of the unsolved major challenges in cartilage tissue engineering is to produce tissue that mimics the zonal organization of the native articular cartilage. An increasing number of studies aim to design zonal organization into tissue-engineered cartilage by forming a stratified construct using zonal cell subpopulations. However, in vitro monolayer expansion of chondrocytes, which is generally required to obtain high cell numbers necessary for tissue engineering and autologous chondrocyte implantation, leads to dedifferentiation of chondrocytes into fibroblast-like cells, resulting in loss of zonal markers, such as the superficial zone protein (SZP) of the superficial zone as well as chondrocytic phenotype markers, such as type II collagen and aggrecan. Several microRNAs (miRNAs), including miR-221, miR-222, miR-143, and miR-145, have been identified from bovine articular cartilage as superficial zone-enriched miRNAs. miR-140 has been known as a cartilage-specific miRNA whose expression is implicated in chondrocyte differentiation and cartilage tissue homeostasis. As miRNAs play an important role in regulating gene expression during cell differentiation and maintaining tissue homeostasis, we determined the expression of the miRNAs with zonal differentiation and homeostasis. We investigated how chondrocyte dedifferentiation during multiple passages and redifferentiation in a three-dimensional (3D) agarose culture regulates the expression of these miRNAs by quantitative reverse transcription–polymerase chain reaction. Additionally, the effect of transforming growth factor beta 1 (TGF-β1), which is known to enhance chondrocytic differentiation and SZP expression, on these miRNAs was evaluated. The expression of miR-221 and miR-222 increased during dedifferentiation and during redifferentiation in a 3D culture and TGF-β1 restored them to normalcy. miR-140 dramatically decreased during dedifferentiation and its expression is partially recovered in a 3D culture with TGF-β1. miR-143 and miR-145 in the superficial chondrocytes decreased during dedifferentiation and further decreased in a 3D culture, but TGF-β1 partially recovered their expression in a 3D culture. In conclusion, the expression patterns of the miRNAs will be of functional utility for strategies and approaches to tissue engineering of the articular cartilage.
Introduction
Multiple passages of chondrocytes in in vitro monolayer culture are generally required to obtain high cell numbers from a limited source in such cell-based methods. 14 Even more passages would be required to obtain a high number of superficial zone chondrocytes due to the limited amount of cells present in the superficial zone of the cartilage. When chondrocytes are removed from their surrounding matrix and grown in monolayer culture, cells undergo dedifferentiation and exhibit a fibroblast-like phenotype that is characterized by a rapid shift in expression of matrix molecules from type II collagen to type I collagen and from aggrecan to versican.15–19 The native zonal differences between cell populations are also rapidly lost during monolayer expansion.20–22 The rapid loss of cartilaginous phenotypes of the cells during in vitro expansion poses a key limitation to the functional tissue engineering with zonal organization. Several attempts have been made to overcome this limitation. The dedifferentiation phenotype can be reversed, at least in part, when dedifferentiated chondrocytes are embedded in three-dimensional (3D) matrices, such as alginate or agarose, since cells re-express the cartilage phenotype upon transfer into 3D culture.23–25 Supplementation of culture media by growth factors, including transforming growth factor beta 1 (TGF-β1) and insulin-like growth factor 1 significantly delays this process.26–28 However, it is still unclear if zonal difference, especially superficial zone protein (SZP) expression can be recovered in the 3D culture of dedifferentiated chondrocytes.
MicroRNAs (miRNAs) are noncoding small RNAs that negatively regulate the expression of downstream targets by repressing the translation or inducing the cleavage of mRNAs. Increasing evidence indicates that miRNAs play a role in chondrocyte differentiation and cartilage homeostasis. Global reduction of miRNAs in chondrocytes by using chondrocyte-specific deletion of Dicer, an essential component for miRNA biogenesis, causes severe skeletal growth defects with abnormal differentiation of chondrocytes in the growth plate. 29 miR-140, whose expression is increased during in vitro chondrogenesis, 30 and decreased in osteoarthritic cartilage,30–32 has been implicated in chondrocyte differentiation and cartilage tissue homeostasis.33,34 Recent miRNA expression profiling studies have identified subsets of miRNAs that are upregulated or downregulated during human articular chondrocyte dedifferentiation process.35,36 How these subsets of miRNAs are involved in chondrocyte dedifferentiation is not known yet, but it is of interest that miR-222 is the only miRNA that was shown to be increased during dedifferentiation in two studies. An intriguing observation is that miR-222, along with its bicistronic miRNA, miR-221, and other miRNAs, including miR-143 and miR-145, are enriched in the superficial zone of the bovine cartilage, indicating that these miRNAs may be related to maintaining functional zonal differences and may play a role in maintaining cartilage homeostasis. 37
The objective of this study was to evaluate how the dedifferentiation and redifferentiation of chondrocyte subpopulations affect the expression of several miRNAs that are enriched in the superficial zone chondrocytes. Previous studies have shown a dramatic change in the characteristic chondrocytic and dedifferentiation marker expression of chondrocytes serially passaged in a monolayer.15–22 A similar approach was used to monitor miRNA changes in three different states: freshly isolated, dedifferentiated, and redifferentiated cells from the superficial and middle zones. We determined the levels of miR-221, miR-222, miR-143, and miR-145, along with miR-140 during dedifferentiation and subsequent redifferentiation in a 3D agarose culture. In addition, the effect of TGF-β1 in a 3D agarose culture was evaluated.
Materials and Methods
Isolation of bovine articular chondrocytes
Bovine articular chondrocytes were obtained as described previously. 38 Briefly, stifle joints from 3-month-old calves were harvested within 12 h of sacrifice and dissected under aseptic conditions. The superficial zone (∼100 μm thick) and the middle zone (∼1 mm thick) of femoral condyle articular cartilage were harvested separately using a dermatome and a scalpel. Cartilage pieces were digested with 0.2% collagenase P (Roche Applied Science, Indianapolis, IN) in a culture medium (Dulbecco's modified Eagle's medium (DMEM)/F-12 medium containing 50 μg/mL ascorbic acid 2-phosphate, 100 μg/mL sodium pyruvate, 0.1% bovine serum albumin, 100 U/mL pen-strep) with 3% fetal bovine serum (FBS) at 37°C. Following an incubation period of 3 h, cells were filtered, washed, and prepared for experiments. For each individual experiment, six stifle joints from three calves were used to obtain pooled zonal subpopulations.
Monolayer culture of bovine articular chondrocytes
Fraction of the isolated cells (1×106 cells) was frozen in liquid nitrogen and stored at −80°C for RNA preparation of the freshly isolated uncultured sample. For monolayer expansion, isolated cells were seeded at a density of 3×105 cells/10-cm culture plate (∼5.3×103 cells/cm2) in the culture medium with 10% FBS. After 6 days, when cells were subconfluent, passage 1 (P1) cells were detached by 0.25% Trypsin- ethylenediaminetetraacetic acid, and washed with phosphate-buffered saline. 1×106 of P1 cells were frozen for RNA preparation and 3×105 of P1 cells were seeded in the 10-cm culture plate for P2 culture. Monolayer culture was continued until P4 and fractions of the cells were collected from each passage for RNA isolation. The medium was changed every 3 days.
3D agarose-embedded culture and TGF-β1 treatment
At the end of P4 monolayer culture, dedifferentiated cells were placed in a 3D agarose culture to redifferentiate the cartilage phenotype. 3D agarose culture was performed as described elsewhere. 23 Briefly, 60-mm culture plates were precoated with a thin film of high Tm low-mr agarose (Bio-Rad, Hercules, CA). Low-melt agarose (free of DNases, RNases, and inhibitors) (Bio-Rad) was used for cell embedding. P4 cell suspension was prepared to 4×106 cells/mL in the 2× DMEM/F-12 culture medium supplemented with 2% insulin–transferrin–selenium (ITS; BD Bioscience, San Jose, CA) and mixed with the same volume of 1% low-melt agarose to give a final 2×106 cells/mL concentration in 0.5% agarose. Cells/agarose suspension was added to the precoated 60-mm plates (1 mL/60 mm plate). After gelation at 4°C for 15 min, cells were fed with 2 mL of the culture medium with 1% ITS and the medium was changed every 3 days. In stimulation experiments, the medium was further supplemented with 10 ng/mL TGF-β1 (R&D Systems, Minneapolis, MN) and the medium was changed every 3 days.
RNA extraction
For monolayer-cultured cells, total RNA, including miRNA was isolated using the miRNeasy Mini Kit (Qiagen, Valencia, CA) according to the manufacturer's instructions. Total RNA from agarose-embedded cells was extracted using the miRNeasy Mini Kit by the protocol modified from the other article. 39 Briefly, each cells/agarose construct was solubilized by vortexing in a mixture of 1 mL QG buffer (Qiagen) and 1.4 mL QIAzol Lysis Reagent until complete dissolution and placed at room temperature for 10 min. The subsequent RNA purification steps were performed as described by the manufacturer. RNA quantity and quality were measured by the NanoDrop instrument (NanoDrop Products, Wilmington, DE).
Gene expression analysis using real-time quantitative reverse transcription–polymerase chain reaction
For miRNA expression analysis, 200 ng of RNA were reverse transcribed using the TaqMan microRNA reverse transcription kit (Applied Biosystems, Carlsbad, CA), including the relevant miRNA-specific primer (Applied Biosystems) in the reaction. Reverse transcription was followed by real-time quantitative reverse transcription–polymerase chain reaction (qRT-PCR) with a TaqMan PCR master mix (Applied Biosystems) and the appropriate miRNA-specific TaqMan probe (Applied Biosystems). For mRNA expression analysis, cDNA was generated from total RNA (500 ng) using the SuperScript First-Strand Synthesis System (Invitrogen, Carlsbad, CA), including random primers, according to the manufacturer's instructions. PCR reactions were performed with SYBR Green reagents (Applied Biosystems) and custom gene-specific primers. The specific primers for the different mRNAs were bovine glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (forward: 5′-GGCGCCAAGAGGGTCAT-3′; reverse: 5′-GTGGTTCACGCCCATCACA-3′), superficial zone protein (SZP) (forward: 5′-AGAAAACCCGATGGCTATGA-3′; reverse: 5′-TCGCCCATCAGTCTAAGGAC-3′), cartilage oligomeric matrix protein (COMP) (forward: 5′-GAGACCGGGCAGCATAACTG-3′; reverse: 5′-AGGGCCACACTGGAAGGAG-3′), aggrecan (forward: 5′-GCGGGTGCGGGTCAA; reverse: 5′-TAGAATCCCGGAGTCATTGGA), type II Collagen (Col2A1) (forward: 5′-GCATTGCCTACCTGGACGAA; reverse: 5′-CGTTGGAGCCCTGGATGA), and type I Collagen (Col1A1) (forward: 5′-ACATGCCGAGACTTGAGACTCA; reverse: 5′-GCATCCATAGTACATCCTTGGTTAGG). All the PCR reactions were performed in triplicate using an ABI 7900HT Fast Real-Time PCR System (Applied Biosystems). Each gene expression level was normalized to GAPDH levels and fold change was calculated using the 2−ΔΔCt method.
Statistical analysis
A sample size of n=3 was used for each group. Statistical differences were assessed by a three-factor analysis of variance. p<0.05 was considered a significant difference.
Results
Changes in chondrocytic and zonal marker expression during chondrocyte dedifferentiation
Dedifferentiation during monolayer culture has been shown to be influenced by several factors, such as cell seeding density. For example, the chondrocytic phenotype was maintained in vitro for a longer time at a higher cell density. 40 Therefore, seeding density was kept constant at every passage in this study. The morphologies of primary articular chondrocytes from superficial (SZAC) and middle zones (MZAC) were not distinguishable at day 1 when most of the cells attached to the surface of the culture plate. Propagation of chondrocytes in monolayer culture transformed typical round chondrocytes into flattened fibroblast-like morphology after serial passage. Gene expression showed rapid dedifferentiation over four passages in both SZAC and MZAC (Fig. 1). Expression of chondrocytic markers, such as COMP, Col2A1, and aggrecan decreased, whereas expression of dedifferentiation marker, Col1A1 significantly increased. Additionally, zonal marker SZP expression also rapidly decreased during the monolayer passage and the difference between zones disappeared by P3. Decreases of the chondrocytic marker gene expression were observed as early as in the P1 and continued with each subsequent passage. Changes in gene expression have gone through 2 to 5 log difference by P4 and the changes plateaued by P4.
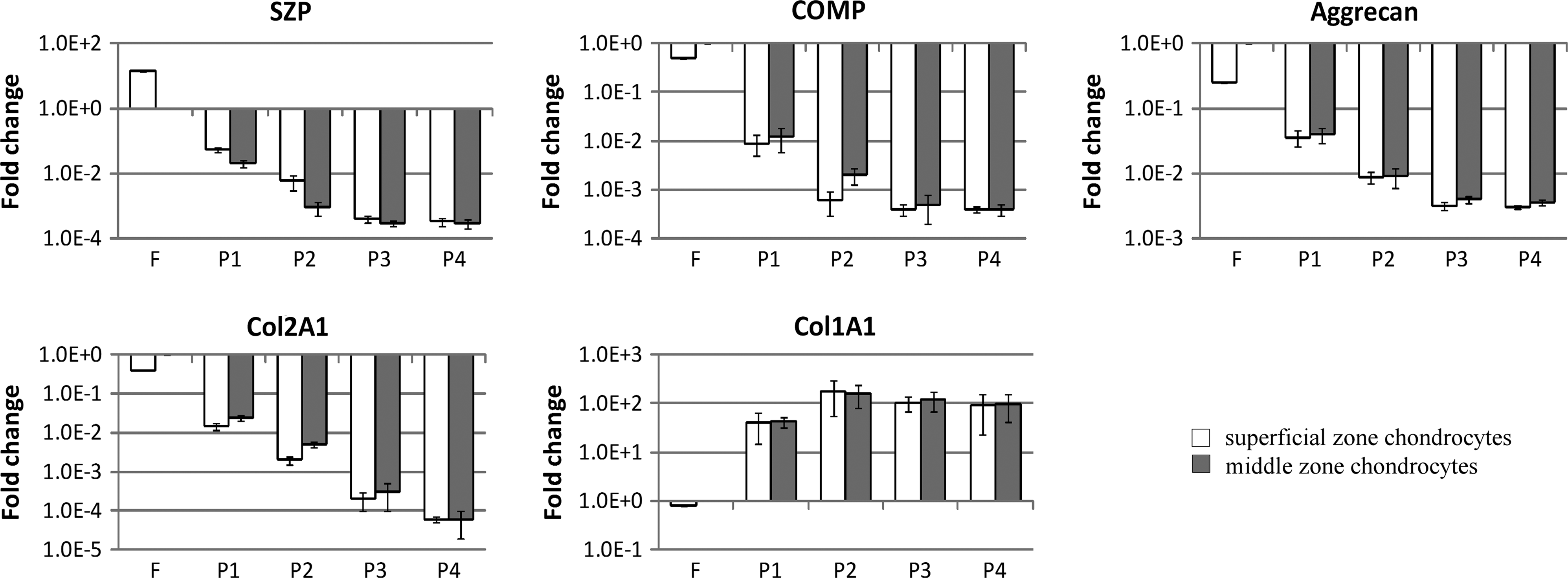
Changes in gene expression during serial passage of articular chondrocytes from superficial zone (SZAC) and articular chondrocytes from middle zone (MZAC). Freshly isolated chondrocytes (F) and cells from each passage up to passage 4 (P1, P2, P3, and P4) were obtained from superficial and middle zones. Expression of superficial zone protein (SZP), cartilage oligomeric matrix protein (COMP), aggrecan, type II Collagen (Col2A1), and type I Collagen (Col1A1) was evaluated by quantitative reverse transcription–polymerase chain reaction (qRT-PCR). Each gene expression level was normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) level. Gene expression levels were shown as fold differences relative to those of freshly isolated MZAC. Data are shown as mean±standard deviation for three independent experiments. Note log scale for the different Y axes and standard deviations are adjusted to log scale.
Changes in miRNA expression during chondrocyte dedifferentiation
Expression of miR-221, miR-222, miR-140, miR-143, and miR-145 was monitored during serial passage of SZAC and MZAC (Fig. 2). Bicistronic genes, such as miR-221/-222 and miR-143/-145, showed similar changes in expression during dedifferentiation and following redifferentiation. Expressions of miR-221/-222 and miR-143/-145 were higher in freshly isolated SZAC, approximately, fourfold and sixfold, respectively, as compared with freshly isolated MZAC. miR-140 expression was twofold higher in freshly isolated MZAC. These zonal differences disappeared rapidly as early as P1 in case of miR-143/-145 and by P2 for miR-221/-222 and miR-140. Expression of miR-221/-222 increased, approximately, six to eightfold. Expression of miR-143/-145 decreased in SZAC to the level comparable with that of MZAC at P1 and no further change was detected, while miR-143/-145 expression in MZAC did not change over time. miR-140 expression dramatically decreased over 100-fold at P1 and further decreased up to 4-log by P4.
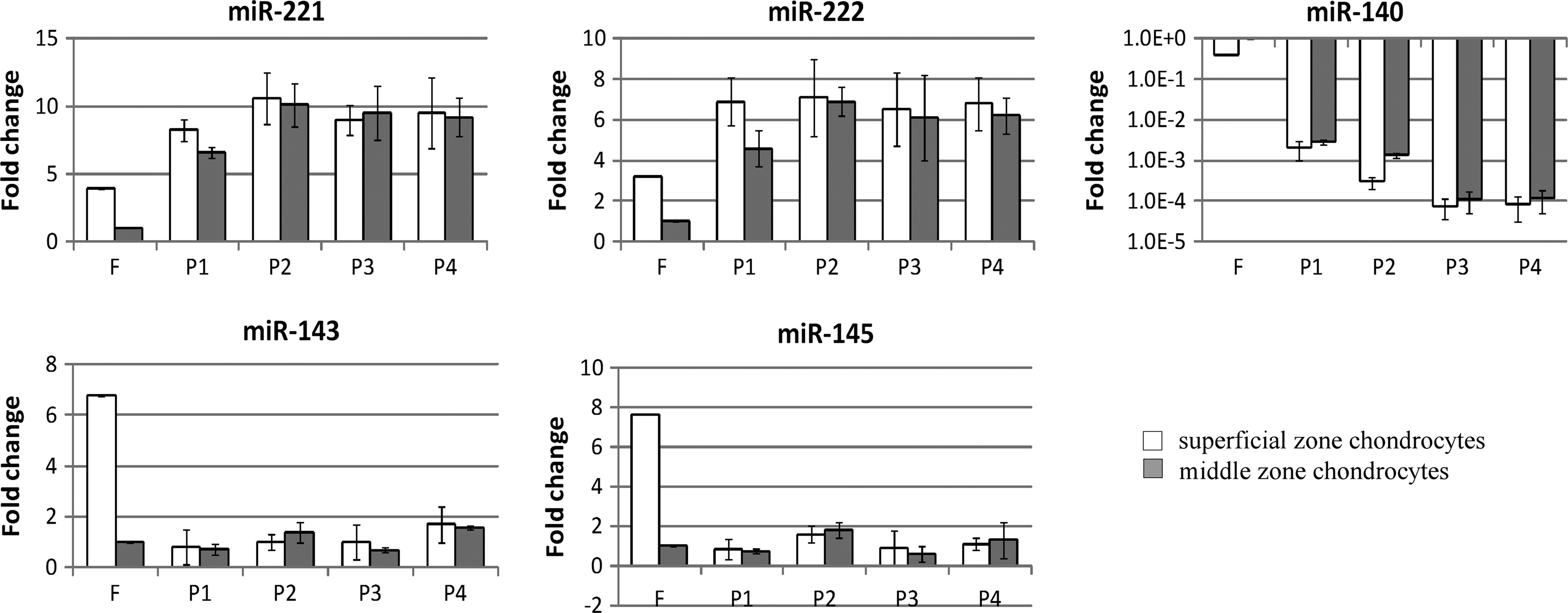
Changes in microRNA (miRNA) expression during serial passage of SZAC and MZAC. From the same cell source as Figure 1, expression of miR-221, miR-222, miR-140, miR-143, and miR-145 was evaluated by qRT-PCR. miRNA expression levels were normalized to GAPDH levels and shown as fold differences relative to those of freshly isolated MZAC. Data are shown as mean±standard deviation for three independent experiments. Note log scale for the Y axis and log scale-adjusted standard deviations in miR-140.
Changes in chondrocytic and zonal marker expression during redifferentiation and effect of TGF-β1
It is well known that dedifferention can be reversed, in part, using 3D cultures. Here, we assessed the effect of TGF-β1 on gene expression (Fig. 3). By simply encasing dedifferentiated cells in agarose increased characteristic markers of chondrogenesis, such as COMP, aggrecan, Col2A1, and SZP expression and decreased Col1A1. Expression of COMP and Col2A1 were further increased by the addition of TGF-β1 in the culture. Noticeably, COMP expression of MZAC was recovered to the original level by TGF-β1. The effect of TGF-β1 on aggrecan and Col1A1 was not very evident. SZP expression was partially recovered by encapsulating cells in agarose. Zonal difference of SZP expression was also partially recovered by agarose embedding. However, TGF-β1 did not further increase SZP expression in agarose-embedded culture, although TGF-β1 has been known to induce SZP expression in primary SZAC in monolayer.
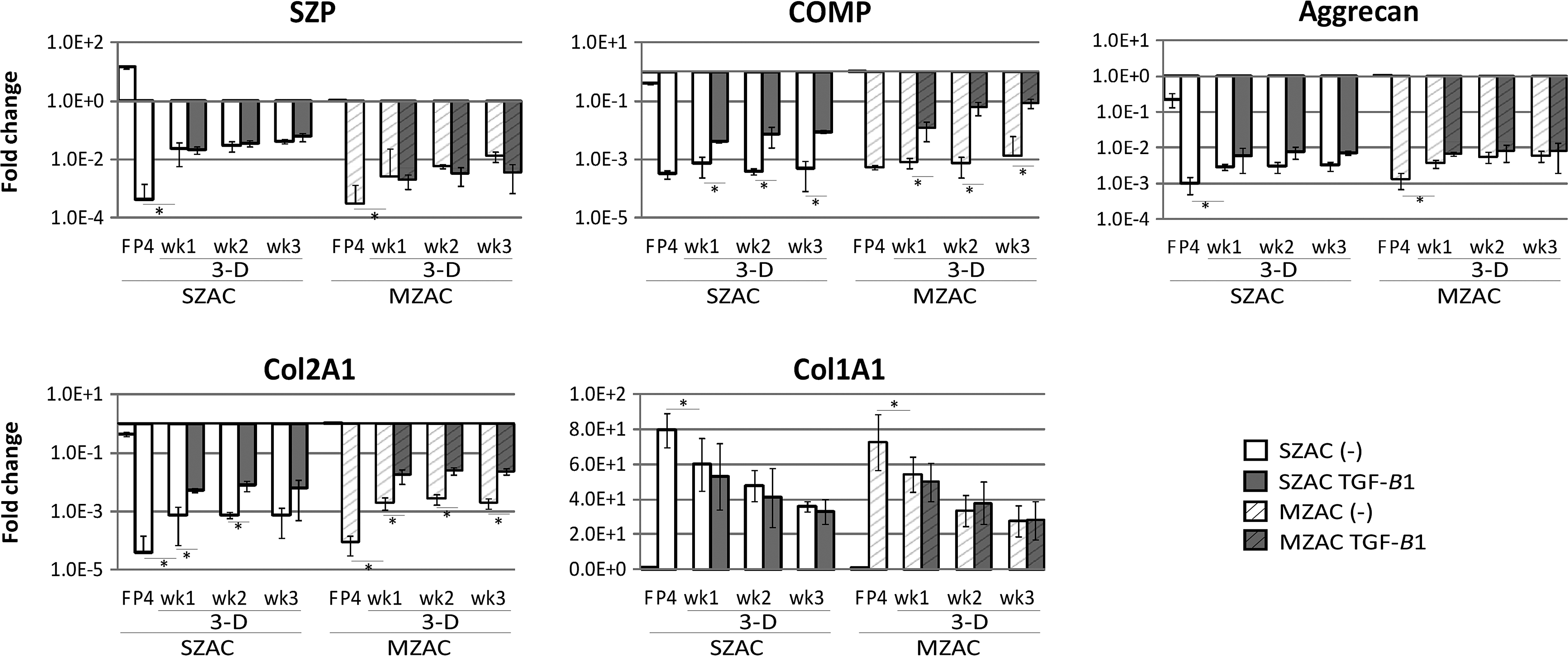
Changes in gene expression in a three-dimensional (3D) agarose culture and the effect of transforming growth factor beta 1 (TGF-β1). P4 dedifferentiated cells from each zone were embedded in agarose (3D) with/without TGF-β1 for 1, 2, and 3 weeks (wk1, wk2, and wk3). Expression of SZP, COMP, aggrecan, Col2A1, and Col1A1 was evaluated by qRT-PCR. Data are shown as mean±standard deviation for three independent experiments. Expression levels were normalized to GAPDH levels and shown as fold differences relative to those of freshly isolated MZAC. Note log scale for the different Y axes and standard deviations are adjusted to log scale. *p<0.05.
Changes in miRNA expression during redifferentiation and the action of TGF-β1
Addition of TGF-β1 in monolayer culture of P0 cells had no effect on the amounts of miR-221, -222, -140, -143, and -145 in 48 h (Data not shown). In contrast, TGF-β1 modulated the expression of these miRNAs from P4 dedifferentiated chondrocytes in 3D agarose cultures (Fig. 4). Embedded culture further increased miR-221/-222 expression, which was increased during dedifferentiation in both SZAC and MZAC. Supplementation of the 3D culture by TGF-β1 dramatically downregulated the expression of miR-221/-222 to the levels comparable to the original level. Decreased miR-140 during the dedifferentiation process was not recovered by simple embedding, but increased by TGF-β1 in both SZAC and MZAC. Expression of miR-143/-145 was dramatically decreased by embedding, but the presence of TGF-β1 prevented the downregulation in both SZAC and MZAC.
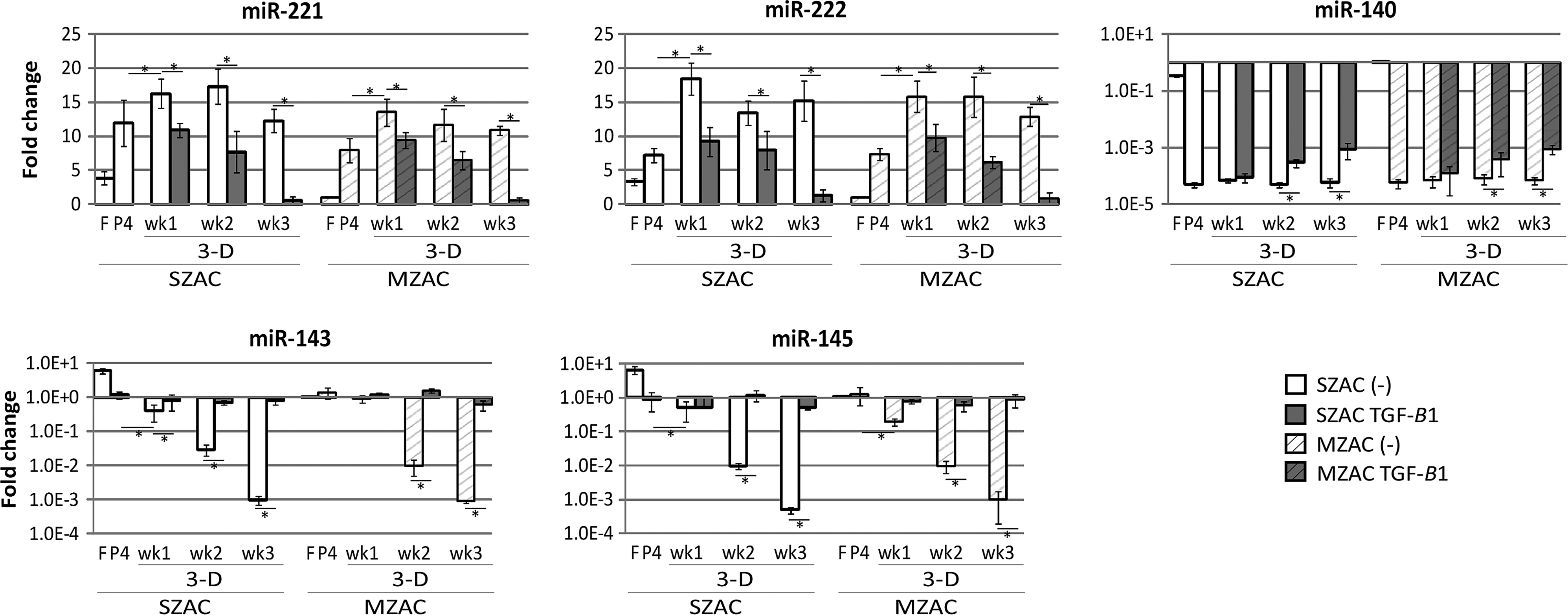
Change in miRNA expression in a 3D agarose culture and the effect of TGF-β1. From the same cell source as Figure 3, expression of miR-221, miR-222, miR-140, miR-143, and miR-145 was evaluated by qRT-PCR. Expression levels were normalized to GAPDH levels and shown as fold differences relative to those of freshly isolated MZAC. Note log scale for Y axes and log scale-adjusted standard deviations in miR-140, miR-143, and miR-145. *p<0.05.
Discussion
We have assessed dedifferentiation and redifferentiation of chondrocyte subpopulations from articular cartilage in terms of expression in the SZP zonal marker, miRNAs that are enriched in superficial zone chondrocytes, and miRNA that is involved in chondrocyte differentiation. Retention or restoration of SZP in the surface of the engineered cartilage is of particular interest since SZP secretion from the superficial zone provides lubricating surface to articular cartilage, which is critical to normal joint function.41,42 Our investigation utilized the 3D agarose hydrogel culture. Hydrogels, well known for their biocompatibility and nutrient-permeable properties, 43 maintain the expression of chondrocytic markers, including Col2 and aggrecan and zonal marker, SZP from primary chondrocyte subpopulations.8,11,44 Hydrogel systems have been tested for passaged dedifferentiated cells. Hydrogels successfully promote chondrocytic marker expression from passaged dedifferentiated chondrocytes,45,46 but the expression of zonal marker SZP is not recovered at all in alginate bead culture of passaged superficial zone chondrocytes. 21 In this study, we showed that a 3D agarose-embedded culture of passaged zonal subpopulations resulted, at least in part, in the re-expression of SZP, as well as chondrocytic markers, COMP, Col2A1, and aggrecan. Their upregulation and the concomitant decrease of Col1A1 suggest that the cell is redifferentiated, producing cartilage-like ECM. This result is different from previously reported failure in SZP recovery, 21 probably due to the difference in cell seeding density used for embedding (5×104 cells/mL 21 vs. 2×106 cells/mL for this study). Cell seeding density in a 3D culture appeared to be important in the recovery of SZP expression from the dedifferentiated superficial zone chondrocytes. Cell-to-cell contacts may not be required for zonal marker expression as another group reported that alginate culture of passaged superficial chondrocytes, but not pellet culture, reacquires the expression of clusterin, another superficial zone marker whose function has not been known. 22 Partial recovery of SZP still means that some other stimulatory factors, such as growth factors, ECM components, and mechanical stimuli, are needed to get full recovery. Incorporation of various growth factors in hydrogel culture of chondrocytes has been reported to stimulate ECM production. Insulin and BMP-2 were shown to enhance production of chondrocytic markers in hydrogel culture. 47 Primary chondrocytes in hydrogel cultures generate a better mechanically functional tissue in the presence of TGF-β3.48,49 Incorporation of TGF-β1 to hydrogel culture improves chondrocyte proliferation and ECM synthesis. 50 Consistently with previous reports, addition of TGF-β1 in our agarose-embedded culture further enhanced the expression of COMP and Col2A1. However, SZP expression was not further enhanced although it is known that SZP expression is greatly increased by TGF-β1 in both mRNA and protein levels in primary superficial chondrocyte monolayer culture.38,51 These results demonstrate that when chondrocytes are encased in 3D cultures, the SZP accumulation appears to be maximal and that it cannot be further stimulated by TGF-β1. Addition of exogenous ECM components may further increase SZP expression as polyethylene glycol-based hydrogels containing exogenous chondroitin sulfate or hyaluronic acid significantly enhance SZP expression. 52 Mechanical stimuli also may further enhance SZP expression as earlier study showed that application of cyclic tension to primary chondrocytes in alginate culture significantly increases SZP. 53
The molecular mechanisms regulating the phenotypes of superficial zone chondrocytes are unclear. Previous studies suggested that it may contain cells with stem or progenitor capacity.54,55 In addition, subsets of miRNAs, including miR-221/-222 and miR-143/-145, are enriched in the superficial zone. 37 Although it remains unclear whether and how these miRNAs are involved in the phenotype of superficial zone, it has been suggested that these miRNAs are related to maintaining functional zonal difference and cartilage homeostasis by regulating mechanotransduction pathways as these miRNAs are also enriched in weight-bearing regions of the cartilage. Among them, miR-221 56 and miR-145 57 have been shown to negatively regulate chondrogenesis. Recently, the roles of specific miRNAs in maintenance of cartilage differentiation and homeostasis are increasingly recognized. miR-140 knock-out mice shows a mild skeletal phenotype with age-related osteoarthritis-like changes, such as ECM loss and fibrillation of articular cartilage, and transgenic mice overexpressing miR-140 in cartilage are resistant to antigen-induced arthritis. 33 ADAMTS-5 metalloproteinase was identified as one of the target gene for miR-140. 33 In view of this, we determined the expression of several miRNAs known to be expressed in articular cartilage with an eye toward utilizing them to modulate redifferentiation of dedifferentiated chondrocytes. Expression of these miRNAs was dramatically changed during dedifferentiation and redifferentiation. It is noteworthy that there are reciprocal changes in miR-140 and miR-221/-222. Expression of miR-221/-222 increased during dedifferentiation and decreased during redifferentiation, whereas miR-140 decreased during dedifferentiation and increased during redifferentiation. Expression of chondrocytic markers was positively related with that of miR-140. However, there was no direct correlation observed between the expression of the zonal marker SZP and any of these miRNAs. It will be necessary in the future study to investigate the potential utility of overexpression of certain miRNAs to confer the phenotype stability during the tissue engineering of articular cartilage maintaining the fidelity of zonal organization to ensure the functional restoration of surface and middle.
Footnotes
Acknowledgments
This work was supported by funds from the Lawrence J. Ellison Endowed Chair.
Disclosure Statement
No competing financial interests exist.