
Abstract
Bone engineering strategies often exploit modulation of the extracellular environment, including delivery of cell and growth factors to repair and regenerate damaged tissues. During bone healing, the expression of endogenous bone morphogenetic proteins is an essential component of the healing response. However, in some situations, the inherent reparative capacity available in the local microenvironment is exceeded by the requirements of the defects. We have recently reported on a novel strategy, that exploits the specificity of antibodies to capture and make available endogenous osteogenic growth factors, referred to as “antibody-mediated osseous regeneration” (AMOR). The objective of the present study was to identify some of the cellular and molecular events involved in AMOR in an effort to begin to elucidate the mechanism of AMOR. The rat critical-sized calvarial defect model was used, where anti-bone morphogenetic protein (BMP)-2 monoclonal antibody (mAb), isotype-control mAb, or recombinant human (rh)BMP-2 were immobilized on absorbable collagen calvarial sponge (ACS) by adsorption, and then implanted into calvarial defects. The results demonstrated persistence of implanted mAbs for short term from 1 to 2 weeks after implantation. Increased cell infiltration was found in defects treated with anti-BMP-2 mAb. Examination of proteins on ACS scaffolds retrieved from defect sites demonstration increased levels of BMP-2, BMP-4, and BMP-7 proteins in sites implanted with anti-BMP-2 mAb. Moreover, BMP-2, BMP-4, and BMP-7 gene expression levels were increased in sites implanted with anti-BMP-2 mAb. Micro-computed tomography and histological analysis demonstrated that the bone within calvarial defects was fully regenerated in sites implanted with either anti-BMP-2 mAb or rhBMP-2. However, rhBMP-2-regenerated bone exhibited aberrant histomorphology with dystrophic calcification and invasion of subjacent areas. Altogether, the results revealed evidence for anti-BMP-2 mAbs to form an immune complex with BMP-2, BMP-4, and BMP-7, and bind to cells to mediate osteogenesis bone regeneration in vivo. This approach suggests a significant role for antibodies in regenerative orthopedic medicine.
Introduction
Although rhBMP-2 is the most investigated exogenous growth factor clinically used for bone engineering, there have been emerging concerns raised by reports on potential side effects of rhBMP-2 in clinical applications.13,14 Other limitations of exogenous therapy with growth factors include the high dosage required, as well as high cost.
Therefore, there is a need for identification of alternative approaches for repair and reconstruction of bone. We have recently provided proof-of-principle evidence for a novel regenerative strategy; entitled antibody-mediated osseous regeneration (AMOR), involving application of immobilized antibodies to capture endogenous BMP-2. 15 The present study was conducted to confirm and extend these basic observations, by investigating the underlying molecular and cellular mechanism as well as the safety and efficacy of AMOR. Results showed that immobilized 3G7 murine anti-BMP-2 monoclonal antibody (mAb) were detected in high concentrations up to 2 weeks in vivo. The presence of the mAb molecule in early stages of healing response altered cell behavior by increasing cellular infiltration, as well as, by activating gene expression of BMP-2, BMP-4, and BMP-7. The antibody mAb cross reacted with BMP-2, BMP-4, and BMP-7 in vitro and in vivo, leading to significantly increased concentrations of these molecules in local sites during early healing. The efficacy of 3G7 anti-BMP-2 mAb in regenerating new bone was quantitatively comparable to that of rhBMP-2 and qualitatively equivalent to endogenous bone.
Materials and Methods
Animals
Sixty four, 5-month-old virgin female Sprague-Dawley rats were housed at 22°C under a 12-h light and 12-h dark cycle and fed ad libitum (Purina Inc., Richmond, IN). All animals were treated according to the regulations and guidelines of the University of Southern California's Institutional Animal Care and Use Committee. Animals were sacrificed either after 1 day, 7 days, or after 6 weeks; the animals were euthanized and the skulls were harvested and stored in buffered formalin until analysis.
In vivo calvarial defect model
The critical-sized calvarial defect model in rats was utilized to investigate the ability of specific anti-BMP-2 Abs to mediate AMOR in vivo as previously described.16,17 Calvarial defects were created in twelve 8-week-old rats under general anesthesia using xylazine and ketamine. Full-thickness skin flaps were raised and the left and right parietal bones were exposed. A 5-mm-diameter defect in the parietal bone of each rat was generated using a trephine under copious saline irrigation. Isotype-matched control Ab (3G7; Abnova, Taiwan) anti-BMP-2 mAbs (3G7; Abnova) were diluted with plain phosphate-buffered saline (PBS) at 25 μg/mL and rhBMP-2 at 1.5 mg/mL. Absorbable collagen sponge (ACS; CollaCote™; Integra, Plainsboro, NJ) was incubated with diluted mAbs or rhBMP-2 for 1 h at room temperature before a surgical procedure. The periosteum was sutured over the grafted calvarial defects to stabilize the ACS membranes within the defects. The skin was approximated with 5.0 polypropylene sutures. After 6 weeks, animals were sacrificed in a CO2 chamber.
Histological analyses
Calvarial specimens were fixed with 10% neutral buffered formalin (Richard-Allan Scientific, Kalamazoo, MI) for 24 h at 4°C. Tissues were then decalcified in a decalcifying solution (Richard-Allan Scientific) for 2 days at 4°C. The samples were dehydrated in graded ethanol (70%, 95%, and 100%) and embedded in paraffin. Five-micrometer sections were stained with hematoxylin and eosin and Masson trichrome (Sigma, St. Louis, MO).
Histomorphometry
Digital histomorphographic images of stained slides were used for histomorphometric analyses by NIH/Scion ImageJ software (Scion Corp., Frederick, MD). Standard American Society of Bone and Mineral Research standard methods and nomenclature were utilized in determination of osteoid bone fill within calvarial defects.
Microcomputed tomography analysis
Specimens were placed in a sample holder in the cranial-caudal direction and scanned using a high-resolution micro-computed tomography system (MicroCAT II; Siemens Medical Solutions Molecular Imaging, Knoxville, TN) at a spatial resolution of 18.676 μm (voxel dimension) 1,536×1,536 pixel matrices. After scanning, the two-dimensional (2D) image data were stored in the Digital Imaging and Communications in Medicine format, transferred to a computer, and 3D reconstruction and analysis were performed. To reduce the size of data for computation, the calvarial region was cropped and saved from the obtained consecutive microtomographic slice images as a volume of interest using Amira software (Visage Imaging, San Diego, CA). In this step, the original spatial resolution was maintained, because data were not resampled. The volume of new bone in calvarial defect was measured using V-Works 4.0 software (Cybermed Inc., Seoul, Korea). The bone tissue was segmented using a global thresholding procedure. New bone was separated from pre-existing bone by applying a cylindrical divider whose base is the same as the defect, and the volume was calculated.
Confocal laser scanning microscopy
Detection of immobilized 3G7 mAb and BMPs by confocal laser scanning microscopy (CLSM) analysis was performed as previously described. 18 Briefly, for Ab serial dilution, various concentrations of 3G7 mAb or isotype-matched control mAb (0–25 μg/mL) were immobilized on 5-mm sterile ACS overnight at 4°C. Samples were placed in a LabTek multichamber slide and washed extensively with 400 μL of 1× PBS (three times, every time 10 min) and blocked with 400 μL of bovine serum albumin for 30 min at room temperature. Samples were washed with 400 μL PBS/Tween one time and the solution was aspirated. The secondary anti-mouse IgG Alexa Fluor 577-conjugated antibody (Molecular Probes, Invitrogen, Carlsbad, CA) was incubated for 60 min at room temperature (RT) in the dark. Samples were washed three times with 400 μL 1× PBS and a final volume of 500 μL of 1× PBS was added for sample visualization. Immediately after staining, samples were examined by a confocal microscope (Fluoview FV10i; Olympus Corp., Tokyo, Japan) at 60× magnification and the fluorescence intensity was quantified by a spot analysis program with the same fluorescent threshold. Triplicate samples were processed.
To detect the concentration of antibodies that remain immobilized on ACS in vivo, ACS scaffolds were incubated with 3G7 anti-BMP-2 mAb or isotype-matched control mAb. The treated ACS samples were implanted within calvarial defect of 24 rats and recovered after 1 or 7 days. After retrieval, samples were fixed with 4% buffered paraformaldehyde, incubated for 30 min at room temperature. Next, samples were evenly split for analysis of antibodies, BMPs, and cell infiltration studies. Samples were washed, blocked, and stained by the same conditions previously described. CLSM and analysis were performed.
To detect binding of soluble rhBMP-2, rhBMP-4, and rhBMP-7 to immobilized 3G7 anti-BMP-2 or isotype-matched mAb, antibodies were immobilized at a saturation concentration (25 mg/mL) on ACS. After immobilization of antibodies, samples were washed with 400 μL of 1× PBS (three times, every time 10 min) at room temperature and incubated with rhBMP antigens for 30 min at room temperature. After extensive washes and blocking, samples were incubated with biotinylated polyclonal anti-BMP-2, anti-BMP-4, or anti-BMP-7 secondary Abs, which did not cross react with other BMP molecules (R&D Systems, Minneapolis, MN). Avidin-conjugated Alexa Fluor 488 was used for fluorescence detection. To detect endogenous binding of rat BMP-2, BMP-4, or BMP-7 by immobilized antibodies, ACS was incubated with 3G7 anti-BMP-2 mAb or isotype-matched control mAb and implanted within rat calvarial defect and extracted after 1 or 7 days. After retrieval, samples were fixed with 4% paraformaldehyde, incubated for 30 min at room temperature. Next, samples were washed, blocked, and immunofluorescent labeled as described earlier with biotinylated anti-BMP-2, anti-BMP-4, or anti-BMP-7 mAb. Avidin-conjugated Alexa Fluor 488 was used for fluorescence detection. For cell detection, 4′,6-diamidino-2-phenylindole, dihydrochloride (DAPI) (Vectashield mounting medium) was used for nucleus staining. Fluorescently labeled samples were visualized by a confocal microscope (Olympus Corp.) at 60×magnification and the fluorescence intensity was quantified by Spot analysis program with the same fluorescent threshold.
RNA isolation
To isolate and purify RNA from ACS specimens, the TRIzol® Plus RNA Purification Kit was (Qiagen, Valencia, CA) utilized according to the manufacturer's instructions. Briefly, tissues were from 24 rats ash frozen in liquid nitrogen before RNA extraction. Frozen tissue samples were homogenized using a mortar and pestle (ThermoFisher Scientific, Pittsburgh, PA) before immersion in the TRIzol reagent and the lysate preparation protocol was performed. Chloroform was added to the samples, followed by centrifugation to separate the solution into an upper aqueous phase containing RNA and a lower phenol-containing organic phase. The sample was then transferred to the PureLink™ RNA Mini Kit Spin Cartridge containing a clear silica-based membrane to which the RNA binds during purification. The RNA was washed to remove contaminants and the purified total RNA was then eluted in RNase-free water (Tris buffer, pH 7.5) and was used in real-time quantitative polymerase chain reaction (qPCR).
qPCR and gene expression
About 4 μg of total RNA was mixed with the genomic DNA elimination buffer (SABiosciences-Qiagen, Valencia, CA), and subsequently mixed with a reverse transcription (RT) cocktail (RT buffer, primer and control mix, RT enzyme; SABiosciences) for cDNA synthesis. The cDNA reaction was mixed with SYBR green qPCR master mix and transferred into 96-well RT2 Profiler PCR Array (SABiosciences-Qiagen), containing appropriate primers for BMP-2, BMP-4, and BMP-7 genes and primers for the housekeeping gene (actin beta, actb). A two-step qPCR was performed starting at 95°C (10 min) for 1 cycle and followed by 95°C (15 s) and 60°C (1 min) for 40 cycles. Normalized threshold cycle data from a real-time instrument were calculated and interpreted using the qPCR array data analysis web tool. Data were expressed in fold change relative to isotype.
Statistics
The Kruskal–Wallis rank sum test was used to analyze the significance of the samples. Statistical significance was assigned at p<0.05.
Results
Immobilization of 3G7 anti-BMP-2 mAb on ACS in vitro
To quantify binding of antibodies to ACS, 3G7 anti-BMP-2 or isotype-matched control mAb were incubated with ACS at different concentrations for 1 h at room temperature. After washing and blocking, preincubated ACS was labeled with fluorochrome-conjugated secondary Ab and quantitatively analyzed by CLSM (Fig. 1A). The minimum detectable concentration of mAb was 1 μg/mL. At Ab concentrations of 10 μg/mL or higher, saturation was achieved with both 3G7 anti-BMP-2 or isotype-matched control mAb. Based on these results, all further studies were conducted with 25 μg/mL of mAb to ensure that saturating dose of mAb was used.
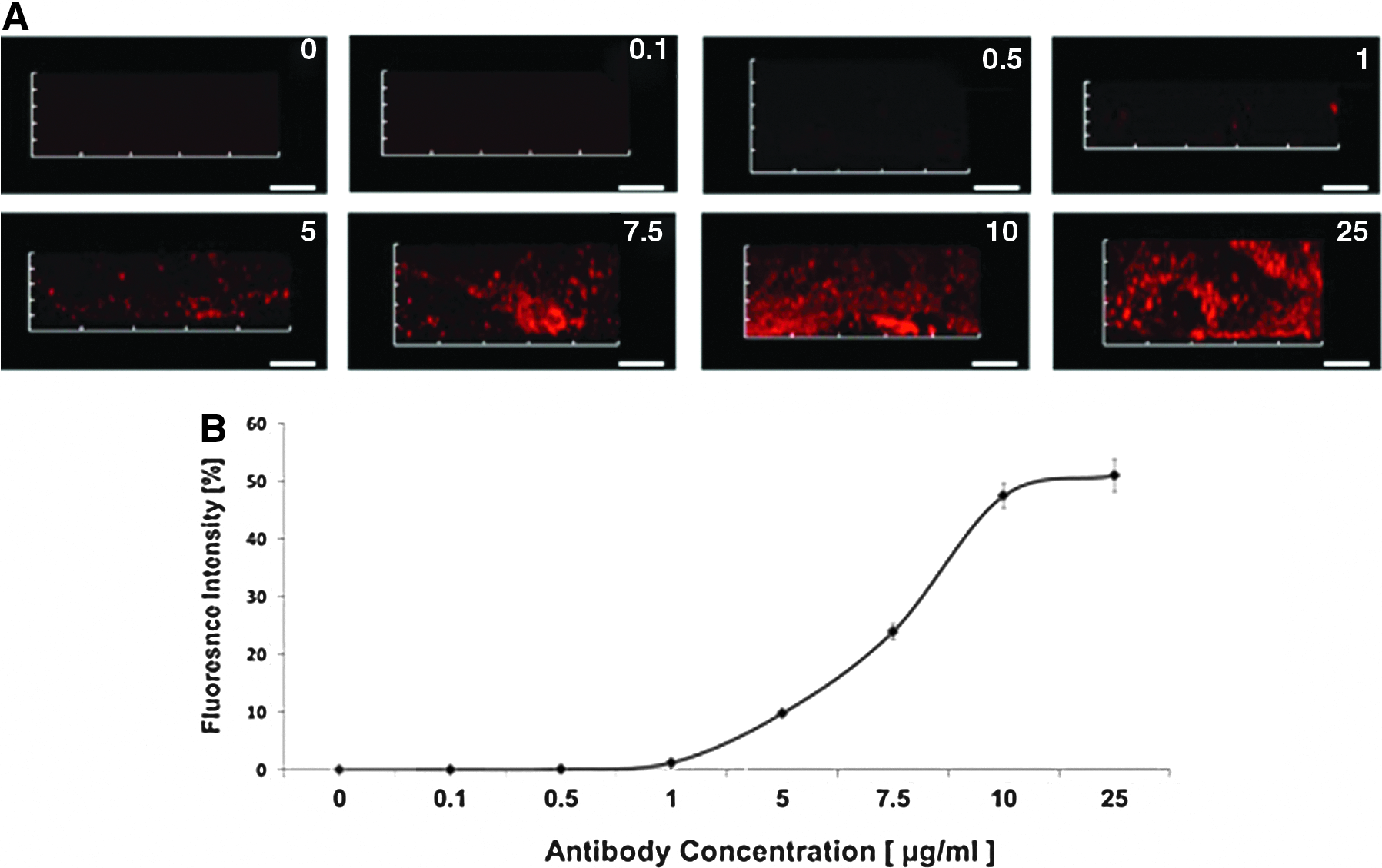
Immobilization of 3G7 anti-bone morphogenetic protein (BMP)-2 monoclonal antibody (mAb) on absorbable collagen calvarial sponge.
Detection of 3G7 anti-BMP-2 mAb after implantation of immobilized Ab in rat calvaria
To determine the persistence of immobilized mAb in vivo after implantation into rat calvaria, 3G7 anti-BMP-2 or isotype-matched mAbs were incubated with ACS at saturating concentrations (25 μg/mL) and implanted in rat calvarial defects. The ACS membranes with adsorbed anti-BMP-2 or isotype-matched mAbs were recovered from animals after 1 or 7 days. The membranes were washed, blocked, and labeled with fluorochrome-conjugated secondary Ab and quantitatively analyzed by CLSM. Results in Figure 2 demonstrated persistence of both isotype-matched IgG2 and 3G7 antibodies after 1–7 days with no quantitative difference between the levels of those two antibodies. Because ACS is rapidly degradable, recovery after 1 week was incomplete and not analyzed in the same manner.
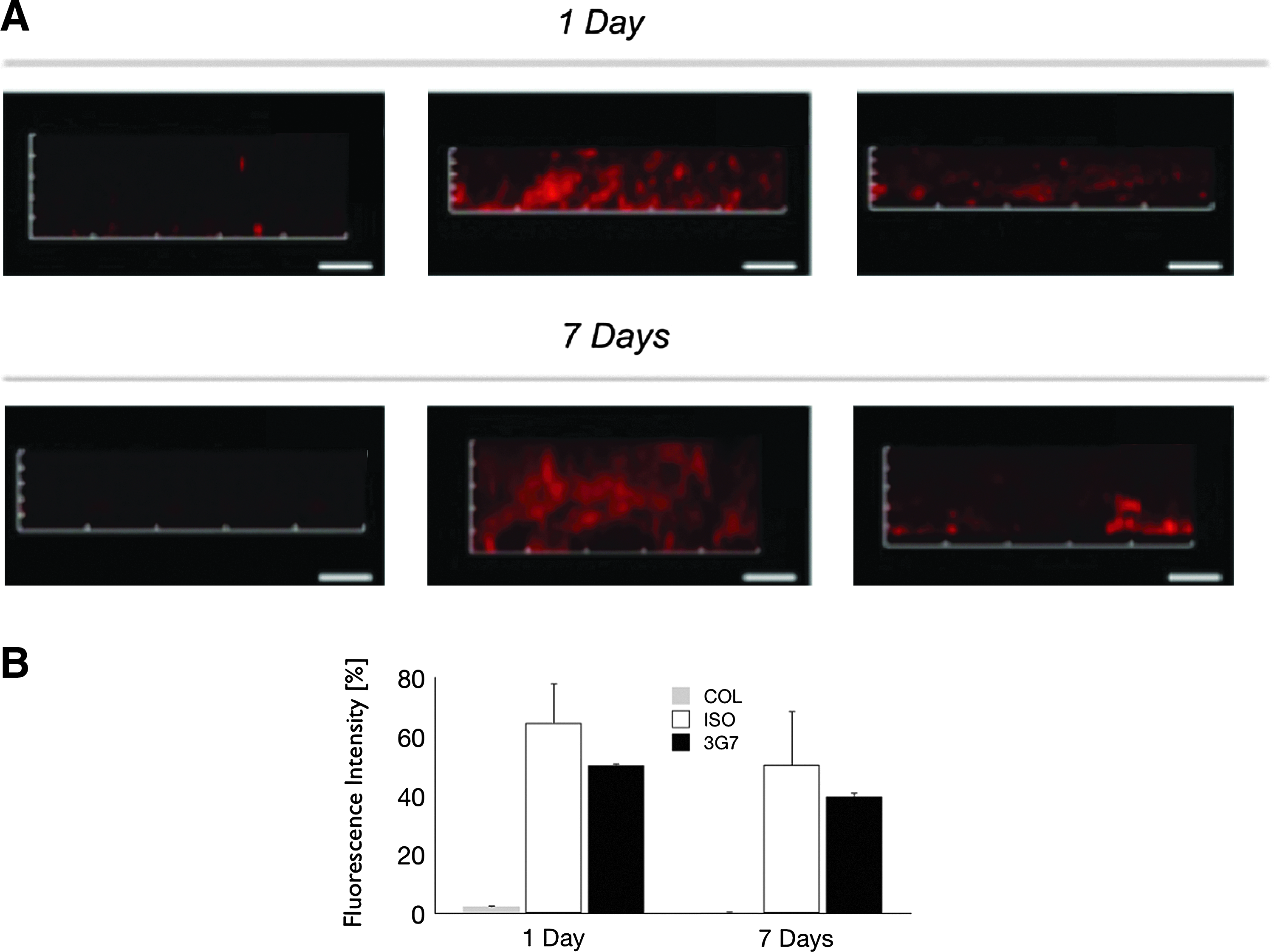
Detection of antibodies during early healing of rat calvarial defects. ACS was incubated with 3G7 anti-BMP-2 mAb or isotype-matched control mAb. The treated ACS samples were implanted within rat calvarial defect and the scaffolds were recovered after 1 or 7 days.
Detection of cell infiltration in response to immobilized anti-BMP-2 mAb
To investigate cellular infiltration in response to anti-BMP-2 mAb, ACS membranes with adsorbed anti-BMP-2 or isotype-matched mAbs were recovered 1 or 7 days after implantation into calvarial defects. The recovered ACS samples were labeled with DAPI. Cells were visualized and their number quantified by CLSM (Fig. 3). Results revealed significantly higher cell numbers after 1 and 7 days in sites implanted with anti-BMP-2 mAb immobilized on ACS, when compared to isotype-matched control samples. No significant cell infiltration was observed into control sites containing untreated ACS or isotype-matched antibodies.

Cell infiltration within calvarial defect sites implanted with 3G7 anti-BMP-2 or isotype-matched mAb. Anti-BMP-2 mAb or isotype-matched control mAb were adsorbed on ACS and implanted in rat calvarial defects. The untreated or mAb-adsorbed ACS scaffolds were harvested after 1 or 7 days.
Specificity of anti-BMP-2 3G7 antibody
In view of the structural homology among BMP-2, BMP-4, and BMP-7, the potential cross reactivity of 3G7 anti-BMP-2 mAb with BMP-4 and BMP-7 was investigated. Western blot analysis demonstrated cross reactivity of the 3G7 anti-BMP-2 mAb with BMP-2, BMP-4, and BMP-7 (Fig. 4A).
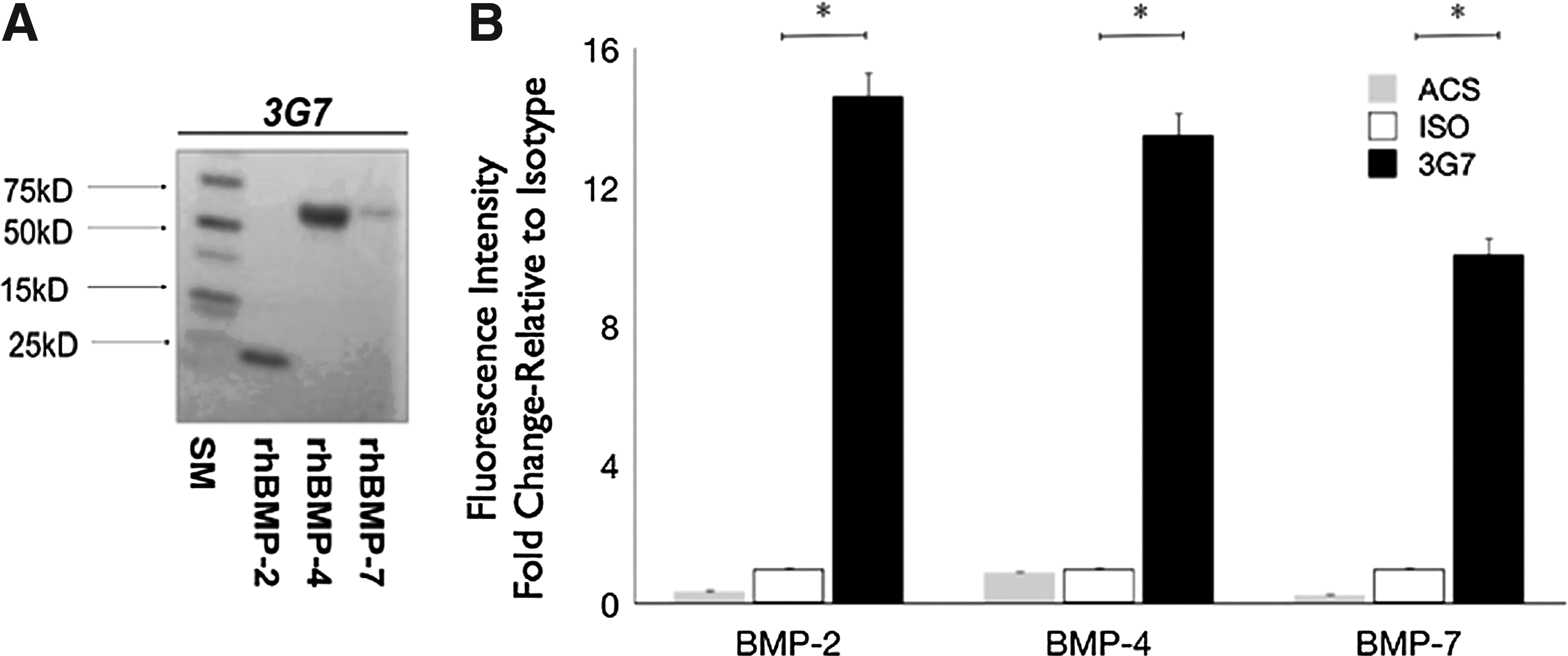
Specificity of 3G7 anti-BMP-2 mAb.
Next, the cross reactivity of immobilized 3G7 anti-BMP-2 mAb to BMP-2, BMP-4, and BMP-7 molecules was investigated. Accordingly, 3G7 anti-BMP-2 or isotype-matched mAbs were incubated with ACS at a saturation concentration (25 μg/mL) overnight at 4°C. The ACS was washed, and then incubated with rhBMP-2, rhBMP-4, or rhBMP-7. After extensive washes and blocking, secondary detection antibodies (biotinylated polyclonal anti-rhBMP-2, anti-BMP-4, or anti-BMP-7) were incubated with each of respective ACSs. This was finally labeled with fluorochrome-labeled streptavidin. Samples were visualized by CLSM and quantitative analysis was performed (Fig. 4B). Results revealed that immobilized 3G7 mAb bound BMP-2, BMP-4, and BMP-7 with roughly equivalent magnitude. Fluorescent detection of BMP-2 was 14.6-fold higher on 3G7-immobilized ACS relative to isotype samples; consistently, BMP-4 presented 13.5-fold higher and BMP-7 10.7-fold higher on 3G7-immobilized ACS when compared to isotype samples. Control treatment, including untreated ACS or with isotype-matched antibodies did not bind significant amounts of BMP molecules in vitro.
Detection of BMP-2, BMP-4, and BMP-7 proteins bound to 3G7 anti-BMP-2 mAb after in vivo implantation
To determine whether endogenous BMP-2, BMP-4, and BMP-7 can specifically bind to 3G7 anti-BMP-2 mAb in vivo, 3G7 mAb was immobilized on ACS and implanted into calvarial defects and treated ACS scaffolds were recovered 1 or 7 days later. The presence of BMP-2, BMP-4, and BMP-7 proteins was detected by fluorescent labeling of recovered scaffolds with specific antibodies against BMP-2, BMP-4, or BMP-7 and visualized by CLSM (Fig. 5A). The fluorescence intensity in each of the samples was measured and compared to isotype control Ab (Fig. 5B). Results revealed that the BMP-2, BMP-4, and BMP-7 levels bound to anti-BMP-2 mAb-containing scaffolds were 10.8-, 14.5-, and 10.4-fold higher, respectively, than those bound to isotype-matched control mAb at 1 day postimplantation. After 7 days, BMP-2, BMP-4, and BMP-7 levels bound to anti-BMP-2 mAb-containing scaffolds were 10.6-, 12-, and 5.8-fold higher, respectively, than those bound to isotype-matched control mAb.

Detection of BMP-2, BMP-4, and BMP-7 bound to immobilized 3G7 anti-BMP-2 or isotype-matched mAb after implantation in rat calvaria. ACS was incubated with 3G7 anti-BMP-2 mAb or isotype-matched control mAb, and implanted in rat calvarial defects. The treated ACS membranes were removed after 1 or 7 days.
In vivo expression of BMP-2, BMP-4, and BMP-7 genes in response to immobilized 3G7 anti-BMP-2 mAb
The expression of endogenous BMP-2, BMP-4, and BMP-7 genes was analyzed in calvarial defect sites implanted with immobilized 3G7 anti-BMP-2 mAb or rhBMP-2 as a positive control and isotype-matched mAb as a negative control. The implanted ACS within calvarial defects was retrieved after 1–7 days; mRNA was isolated and analyzed by qPCR. The expression of each of the normalized genes was compared to that on scaffolds recovered from sites implanted with isotype-matched control mAb. The results in Figure 6 showed increased expression of the BMP-2 gene at 1 day (3G7: 3.3-fold; rhBMP-2: 12.1-fold) and peak expression after 7 days (3G7: 31.7-fold; rhBMP-2: 19.1-fold). In addition, BMP-4 expression was moderately increased (2.3-fold) with 3G7 anti-BMP-2 and decreased with rhBMP-2 treatment (1.5-fold) at 1 day. After 7 days, increased BMP-4 gene expression remained at basal levels in both 3G7 and rhBMP-2 sites. The expression levels of BMP-7 were increased in both 3G7 and rhBMP-2 sites at 1 day (3G7: 3.8-fold; rhBMP-2: 9.5-fold) and returned back to basal levels at 7 days (3G7: 1.8-fold; rhBMP-2: 1.0-fold).
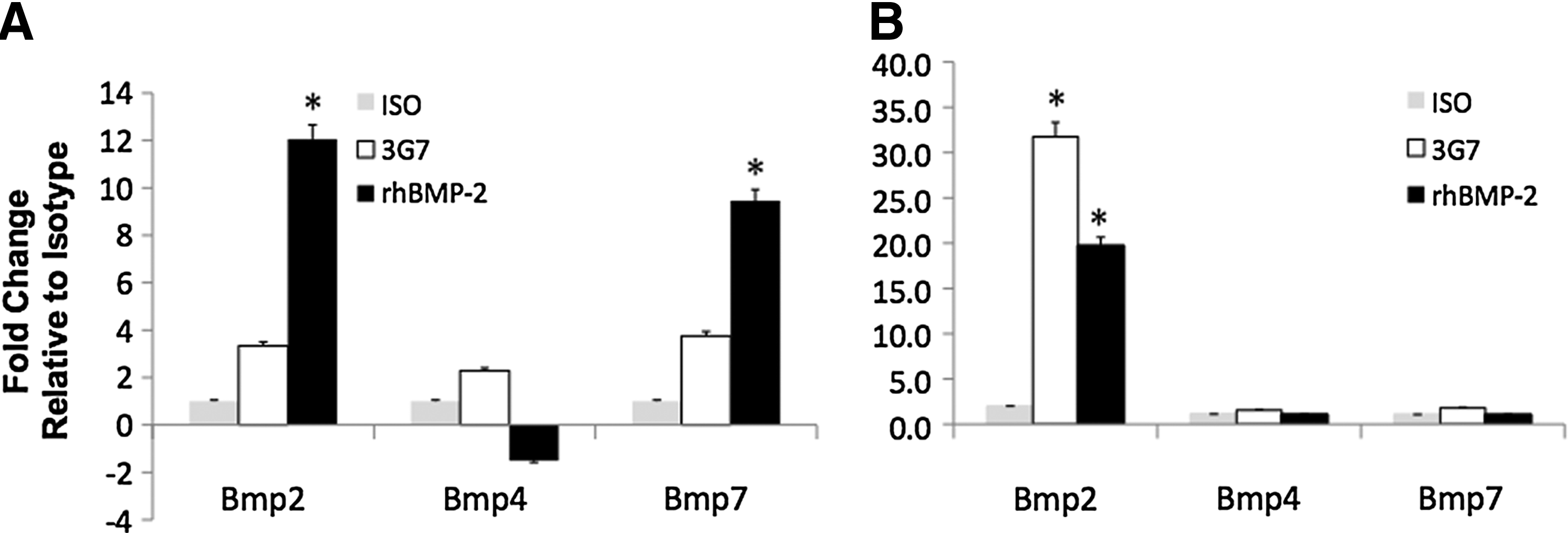
Differential expression levels of BMP-2, BMP-4, and BMP-7 genes in rat calvaria after implantation of immobilized 3G7 anti-BMP-2. ACS was incubated with 3G7 anti-BMP-2 mAb, isotype-matched negative control mAb, or rhBMP-2 as positive control. Treated ACS were implanted in rat calvarial defects and membranes were recovered after 1
Bone regeneration in response to anti-BMP-2 monoclonal antibody
The quantity and quality of bone regenerated in calvarial defect sites implanted with immobilized 3G7 anti-BMP-2 mAb or rhBMP-2 as positive control and isotype-matched mAb as negative control were compared after 6 weeks. Quantitatively, micro-CT analysis revealed equivalent degree of bone fill with radio-dense material when ACS with anti-BMP-2 mAb or rhBMP-2 was implanted. Control treatment with isotype-matched mAb did not mediate a significant degree of calvarial bone repair during the experimental period (Fig. 7B), which was significantly different from sites with 3G7 anti-BMP-2 mAb or rhBMP-2 (p<0.001).

Quantitative and qualitative comparison of bone formation in response to anti-BMP-2 monoclonal antibody (antibody-mediated osseous regeneration) with that of rhBMP-2. ACS was incubated with BMP-2-specific mAb clone (3G7), isotype-matched control mAb, or rhBMP-2. The ACS with adsorbed mAbs or rhBMP-2 was implanted in rat calvarial defects. Representative three-dimensional reconstruction of micro-CT images
Histologically, sites implanted with isotype-matched control mAb were characterized by strips of fibrovascular connective tissue consisting of interwoven bundles of collagenous fibers in association with ovoid-to-stellate fibroblasts/myofibroblasts (Fig. 7C). No evidence of mineralized tissue was seen in these sections. On the other hand, sites implanted with rhBMP-2 were characterized by wider strips of fibrovascular and mineralized connective tissue. The collagenous and vascular connective tissue with fibroblastic/myofibroblastic and inflammatory cells were observed and admixed with osseous tissue and dystrophic calcifications. Trichrome staining revealed a viable mature woven bone formation with a lamellate pattern and osteocytes within lacunae were evident. The morphology recapitulated a fibro-osseous lesion-like histopathologic pattern with invasion of mineralized tissue in subjacent regions, suggesting ectopic calcification. When implanted with 3G7 anti-BMP-2 mAb immobilized on ACS, results were characterized by strips of fibrovascular and mineralized connective tissue comprising predominantly a central dense layer of mature viable woven bone with osteocytes within lacunae. The bone resembled the cortical bone and was well-organized with no evidence of extension superiorly or inferiorly. The bone was circumscribed by a well-defined fibrous capsular rim and myxoid fibrovascular tissue superiorly and there is no evidence of dystrophic calcifications in these sections.
Discussion
The present investigation confirmed and extended our previous observations, 15 demonstrating the ability of specific anti-BMP-2 mAb immobilized on ACS to mediate bone formation in rat calvarial defects. The hypothesis pursued in the present report was that bone formation by AMOR is attributable to the ability of anti-BMP-2 mAb to capture and tether endogenous BMP-2 and increase its local availability to induce bone regeneration. The present data demonstrated that antibodies on the ACS scaffold implanted in calvarial defects persisted for at least 1 week. Since the ACS membrane rapidly degrades within 3 weeks, it was not possible to retrieve the membrane in an intact manner beyond the first week. Nonetheless, the content of calvarial defects was retrieved and murine antibodies were detected by Western blot analysis at 2 weeks (data not shown).
To investigate our hypothesis, the ability of immobilized 3G7 anti-BMP-2 mAb to bind rhBMP-2 from a solution was examined. In view of the high degree of homology of BMP-2 with other osteogenic BMPs6,19 (e.g., BMP-4 and BMP-7), the cross reactivity of 3G7 anti-BMP-2 mAb with rhBMP-4 and rhBMP-7 was also investigated. In vitro assays observed cross reactivity of anti-BMP-2 mAb with rhBMP-4 and rhBMP-7. The in vivo results demonstrated that in calvarial sites with implanted 3G7 anti-BMP-2 mAb on ACS, endogenous BMP-2, BMP-4, and BMP-7 were detected. Because rats were used as the host for the calvarial defect studies, comparison of human versus rat BMP protein sequence was performed (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). When human BMP-2 protein was used as a template, rat BMPs showed a high degree of homology with human, including BMP-2 (91.41%), BMP-4 (96.33%), and BMP-7 (97.48%). In fact, the local concentrations of BMP-2, BMP-4, and BMP-7 were elevated between 5.8- and 14.5-fold relative to negative control sites (isotypic mAb) during the first week after implantation of anti-BMP-2 mAb. One of the potential problems with detecting BMP-2, BMP-4, and BMP-7, which has been bound to anti-BMP-2 mAb is that the detection antibody against each of those molecules may not bind or bind less efficiently due to masking of the epitopes by 3G7 mAb. This could lead to underestimation of the presence of each molecule. To minimize this limitation, polyclonal anti-BMP antibodies were used for detection to increase the likelihood of better detection. The question raised by detection of increased BMPs bound to the scaffold is whether these BMPs were circulating molecules, which were captured by the anti-BMP-2 mAb or whether the BMPs were de novo expressed by the infiltrated cells. The data showing increased infiltration of cells into the scaffolds with 3G7 anti-BMP-2 mAb demonstrated specific recruitment of cells, although the phenotype of these cells is yet to be characterized. The temporal relationship between the detection of BMP-2, BMP-4, and BMP-7 protein on ACS scaffolds and increased BMP gene expression provided some insights. In sites implanted with rhBMP-2, there was early expression of BMP-2 and BMP-7 genes after one day. However, the peak expression of BMP-2 in sites with anti-BMP-2 mAb occurred after 7 days. This suggested a lag period, which was necessary to increase BMP-2, BMP-4, and BMP-7 gene expression and capturing of BMP-2, BMP-4, and BMP-7 proteins onto the scaffolds, presumably by anti-BMP-2 mAb. These stimuli likely provided feed-forward to further increase BMP-2 gene expression. Whereas in sites where exogenous rhBMP-2 was implanted, a large bolus of stimulus was present, which led to increased induction of BMP gene expression at an earlier time point, that is, one day. There are a number of questions that remain unanswered, including: (1) What is the phenotype of the cells recruited in response to anti-BMP-2 implantation? (2) What other molecules are captured by anti-BMP-2 mAb? (3) Are BMPs bound to anti-BMP-2 mAb on a scaffold responsible for stimulating osteogenesis or the BMPs and other molecules expressed by cells recruited to the site?
Previous studies have showed that endogenous BMP-2 is required to initiate fracture healing 9 and when combined with BMP-4 19 and BMP-7, 20 the regenerative capacity is increased. This highlights the main advantage of employing specific therapeutic mAb instead of application of growth factors. Currently, all growth factor therapies entail administration of a large bolus of a single agent, which is contrary to the physiologic process of wound healing, where a large number of signals participate in a specific temporal sequence. Antibodies, through their potential for cross reactivity can capture multiple homologous molecules to provide effective signals for regeneration at the precise temporal sequence dictated by the healing process, since they merely amplify the endogenous signals. It is also important to note that cross reactivity may lead to potential adverse reactions. Careful analysis of organs and tissues for histopathological changes has shown absolutely no evidence of adverse changes in animals treated with antibodies. Indeed, the aberrant healing response was only observed in the animals treated with rhBMP-2. It was observed that calvarial defect sites implanted with rhBMP-2 and ACS exhibited aberrant histomorphology with dystrophic calcification and invasion of subjacent regions. In previous studies, although rhBMP-2 had been shown to possess high efficacy in regenerating bone, there have been recent safety concerns about this mode of therapy.14,21,22 Differences between the initial (2–8 weeks) healing response with the use of anti-BMP-2 mAb and rhBMP-2 have been qualitative, but not quantitative in nature. The process of bone healing within sites treated with anti-BMP-2 mAb appeared to histologically recapitulate normal bone repair. On the other hand, sites treated with rhBMP-2 appeared to have tissue organization with distinct differences from normal bone. Patients who have been treated with supra-physiological doses of rhBMP-2 in spinal fusion procedures have been found to experience adverse swelling and undesired ectopic bone formation. 23 Craniofacial adverse reactions have also been described to be more frequent in rhBMP-2 off-label uses, including reconstruction of the mandible after fracture or cancer or alveolar cleft repair. 22 A systematic characterization of all molecules bound to anti-BMP-2 mAb in vivo and their role in bone repair has not been performed and is planned. Systemic correlation with rhBMP-2 administration has also been reported, including the development of antibodies against BMP-2 or to the carriers. The reported immunogenicity has been positively correlated with higher doses of rhBMP-2 and collagen. 24
Based on the results of the present study, the efficacy of AMOR may be hypothesized to be attributed to (1) ability of antibody to bind low concentrations of endogenous BMPs to achieve the effective dose at an earlier time point, (2) achieving higher peak concentrations of BMPs, (3) sustaining the effective BMP concentrations for longer period, and (4) additive or synergistic effects due to simultaneous increased availability of BMP-2, BMP-4, and BMP-7. Current efforts are directed at investigating each aspect of this hypothesis.
Footnotes
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.