
Abstract
Background:
Stem cell-based bone tissue engineering with adipose-derived stromal cells (ASCs) has shown great promise for revolutionizing treatment of large bone deficits. However, there is still a lack of consensus on cell surface markers identifying osteoprogenitors. Fluorescence-activated cell sorting has identified a subpopulation of CD105low cells with enhanced osteogenic differentiation. The purpose of the present study was to compare the ability of CD90 (Thy-1) to identify osteoprogenitors relative to CD105.
Methods:
Unsorted cells, CD90+, CD90−, CD105high, and CD105low cells were treated with an osteogenic differentiation medium. For evaluation of in vitro osteogenesis, alkaline phosphatase (ALP) staining and alizarin red staining were performed at 7 days and 14 days, respectively. RNA was harvested after 7 and 14 days of differentiation, and osteogenic gene expression was examined by quantitative real-time polymerase chain reaction. For evaluation of in vivo osteogenesis, critical-sized (4-mm) calvarial defects in nude mice were treated with the hydroxyapatite-poly(lactic-co-glycolic acid) scaffold seeded with the above-mentioned subpopulations. Healing was followed using micro-CT scans for 8 weeks. Calvaria were harvested at 8 weeks postoperatively, and sections were stained with Movat's Pentachrome.
Results:
Transcriptional analysis revealed that the CD90+ subpopulation was enriched for a more osteogenic subtype relative to the CD105low subpopulation. Staining at day 7 for ALP was greatest in the CD90+ cells, followed by the CD105low cells. Staining at day 14 for alizarin red demonstrated the greatest amount of mineralized extracellular matrix in the CD90+ cells, again followed by the CD105low cells. Quantification of in vivo healing at 2, 4, 6, and 8weeks postoperatively demonstrated increased bone formation in defects treated with CD90+ ASCs relative to all other groups. On Movat's Pentachrome-stained sections, defects treated with CD90+ cells showed the most robust bony regeneration. Defects treated with CD90− cells, CD105high cells, and CD105low cells demonstrated some bone formation, but to a lesser degree when compared with the CD90+ group.
Conclusions:
While CD105low cells have previously been shown to possess an enhanced osteogenic potential, we found that CD90+ cells are more capable of forming bone both in vitro and in vivo. These data therefore suggest that CD90 may be a more effective marker than CD105 to isolate a highly osteogenic subpopulation for bone tissue engineering.
Introduction
Mesenchymal stem cells (MSCs) were first discovered in the bone marrow by Friedenstein et al. in 1974.1,2 Since then, the interest in adult MSCs has progressively grown due to their ability to self-replicate, while maintaining the capacity to differentiate into multiple cell types. In 2001, Zuk et al. published the first report of multipotent cells in adipose tissue, naming these processed lipoaspirate cells based on their method of isolation. 3 At around the same time, Gimble and coworkers identified adipose-derived stromal cells (ASCs) that were capable of osteogenic differentiation.4–6 These cells have many properties that suggest considerable potential utility in cellular therapy for bone repair and regeneration. Importantly, unlike human bone marrow-derived MSCs (BM-MSCs), ASCs can be easily and safely harvested in large quantities with minimal morbidity. The abundance of stem cells in adipose tissue is 100-fold higher than that in the bone marrow and the yield of ASCs after expansion is approximately 400,000 cells per mL of lipoaspirate tissue.3,7,8
Like BM-MSCs, ASCs have shown the ability to undergo osteogenic differentiation. However, the freshly isolated stromal vascular fraction (SVF) from adipose tissue contains a mixture of cells, which not only includes ASCs, but also endothelial cells, smooth muscle cells, pericytes, fibroblasts, and other circulating cells. 9 Flow cytometric analysis of ASCs has shown that they share common cell-surface receptors with BM-MSCs.4,10–12 Despite several reports being published to establish markers for the ASC phenotype, there is still a lack of consensus over profiles identifying adipose-derived mesenchymal progenitors or osteoprogenitor cells.13–15 Furthermore, ASCs have been found to exhibit a change in the surface marker phenotype when cultured in vitro, which can generate variability in selection of a proper surface marker for osteogenesis.14,16
Using single-cell transcriptional analysis, a recent study in our laboratory found expression patterns for the cell surface receptor endoglin (CD105) to be closely associated with osteogenic potential of ASCs. 17 By combining microfluidic analysis with fluorescence-activated cell sorting (FACS), CD105low ASCs were found to be capable of enhanced osteogenic differentiation relative CD105high and unsorted cells. A second cell surface receptor was also found to correlate with expression of osteogenic genes independent from CD105. Known as Thy-1 (CD90), this surface marker is a 25–30 kDa glycosyl-phosphatidylinositol (GPI)-linked membrane protein and has previously been shown to be associated with osteoprogenitor cells.18–21 The purpose of the present study was to thus compare the ability of CD90 to identify osteoprogenitors relative to CD105. Identifying a cell surface marker associated with enhanced osteogenic capacity would have a profound impact on optimizing cell-based skeletal tissue engineering applications.
Materials and Methods
Cell isolation and culture
Lipoaspiration specimens were obtained after acquiring informed consent from patients, in accordance with the Stanford University Institutional Review Board guidelines. ASCs were harvested from the adipose tissue of two female patients (ages 37 and 48 years) without major medical conditions who were undergoing elective lipoaspiration of the abdomen, flank, and/or thigh region. ASCs were isolated as described previously by Zuk et al. 12 Briefly, raw lipoaspirates were washed and treated with 0.075% collagenase type I (Sigma-Aldrich, St. Louis, MO) in the Hank's balanced salt solution (Cellgro, Manassas, VA) for 1 h at 37°C with gentle agitation. The cellular pellet was resuspended and plated onto conventional tissue culture plates in the Dulbecco's modified Eagle's medium (DMEM, High Glucose, GlutaMAX™, HEPES; Life Technologies, Grand Island, NY) supplemented with 10% FBS and 1% P/S.
Fluorescence-activated cell sorting
ASCs were cultured for 36 h before being sorted. The sort was performed on a FACS Aria II instrument (BD Biosciences, San Jose, CA) with the use of a 100-μm nozzle. Briefly, cells were lifted using TrypLE (Invitrogen, Carlsbad, CA) and centrifuged for 5 min at 1000 rpm. The supernatant was discarded by aspiration, and the cells were incubated for 30 min in a flow cytometry buffer (PBS, 2% FBS) containing allophycocyanin-conjugated mouse anti-human CD90 and fluorescein isothiocyanate-conjugated mouse anti-human CD105 antibodies (BD Biosciences). Unsorted cells, CD90+, CD90−, CD105high, and CD105low cells were then seeded at 100,000 cells/well in a six-well plate and treated with the osteogenic differentiation medium (ODM).
In vitro osteogenic differentiation assay
For osteogenic differentiation, all assays were performed in triplicate wells. After attachment, cells were grown to at least 80% confluence before being cultured in the ODM, which consisted of the DMEM, High Glucose, GlutaMAX, HEPES supplemented with 10% FBS, 1% P/S, 100 μg/mL ascorbic acid, and 10 mM β-glycerophosphate. Alkaline phosphatase (ALP) staining and quantification were performed at 7 days. Photometric quantification of Alizarin red stain was performed at 14 days to assay extracellular mineralization, as previously described.22,23
Reverse transcription and quantitative real-time polymerase chain reaction
RNA from cultivated cells was extracted using the RNeasy Mini Kit (Qiagen, Valencia, CA) according to the manufacturer's protocol. Reverse transcription was performed and osteogenic gene expression was examined by quantitative real-time polymerase chain reaction (qRT-PCR) using the Applied Biosystems Prism 7900HT sequence detection system (Applied Biosystems, Foster City, CA) and SYBR Green PCR Master Mix (Applied Biosystems). The amount of PCR product was calculated using an external GAPDH standard curve and LightCycler software. All values were normalized based on the GAPDH expression in the corresponding samples. Specific primers for the genes examined were based on their PrimerBank sequence. 24
In vivo bone formation
For evaluation of in vivo osteogenesis, nonhealing, critical-sized (4-mm) calvarial defects were created in the right parietal bone of 60-day-old male Crl:CD-1-Foxn1nu mice (Charles River Laboratories, Wilmington, MA), as previously described. 25 All research involving vertebrate animals was performed in accordance with approved protocols by Stanford APLAC. Hydroxyapatite (HA)-coated poly(lactic-co-glycolic acid) (PLGA) scaffolds were fabricated from 85/15 poly(lactic-co-glycolic acid) by solvent casting and a particulate leaching process as previously described. 26 In preparation for cell engraftment, scaffolds were seeded with ASCs 24 h before implantation. Each scaffold was implanted alone or was seeded with 150,000 cells in 125 μL of medium in 96-well culture plates and incubated overnight. Animals were treated with the HA-PLGA scaffold seeded with the following cell types: CD90+ ASCs (n=3), CD90− ASCs (n=3), CD105high ASCs (n=3), and CD105low ASCs (n=3). As controls, no scaffold (n=3), as well as HA-PLGA scaffolds alone (n=3), were used. Before implantation, scaffolds were copiously rinsed with PBS.
For microcomputed tomography (micro-CT) scans, the mice were anesthetized with isoflurane. Imaging was performed using a Siemens Inveon MicroPET/CT scanner (Siemens Medical Solutions, Inc., Malvern, PA). Using our scan protocol parameters, each high-resolution 100-μm image was acquired in a total scan time of 10 min. Mice were scanned immediately, postoperatively and at 2, 4, 6, and 8 weeks following surgery. Data were reconstructed into three-dimensional surfaces using the Siemens Inveon Research Workplace 4.0 Software (Siemens Medical Solutions, Inc., Malvern, PA). The three-dimensional reconstructed images were then analyzed using ImageJ software (Image Processing and Analysis in Java, NIH, Bethesda, MD). The area of the calvarial defects was evaluated by quantifying pixels in the defect. Percentage healing was then determined by dividing the defect area by the defect size immediately postoperatively.
Histological analysis
At 8 weeks postoperatively, calvaria were harvested, immediately fixed in 10% formalin overnight, decalcified in 19% EDTA, dehydrated through an ethanol series, and embedded in paraffin as previously described. 27 Deparaffinized sections were stained with Movat's Pentachrome to detect bone matrix formation. Bright field images were obtained with a 20× objective at room temperature using a Leica DM5000 microscope (Leica Microsystems, Inc., Wetzlar, Germany) equipped with a DFC300FX camera. The images were analyzed using Leica IM1000 Version 4.0 Image Acquisition Software (Leica Microsystems).
Statistical analysis
Numerical data are presented as means±standard deviations. In figures, bar graphs represent means, and error bars represent one standard deviation. Unless otherwise stated, statistical analysis was performed using a one-way analysis of variance for multiple group comparisons and a two-tailed Student's t-test was used to directly compare two groups. The Newman–Keuls and Tukey post hoc test were performed using SPSS (Version 16, IBM) to compare the osteogenic potential of all subpopulations. A value of *p<0.05 was considered significant.
Results
Fluorescence-activated cell sorting
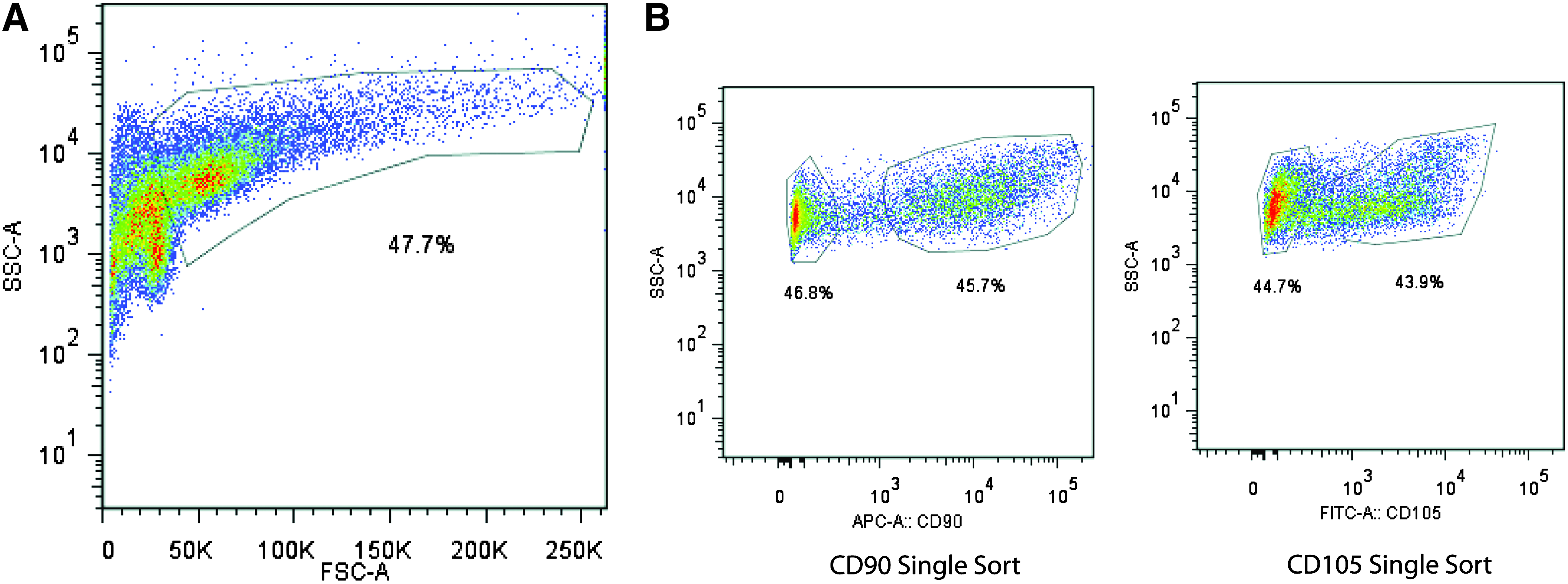
A strategy using FACS on as few cell surface markers as possible would allow for the largest capture of cells in the shortest amount of time. Because it has been suggested that CD90 and CD105 may be useful as early markers indicating commitment to osteogenic differentiation, we sorted ASCs into CD90+, CD90−, CD105high, CD105low, and unenriched (sorted for live cells only) groups using FACS (Fig. 1A, B). FACS analysis of gated subpopulations at 36 h yielded a CD90+ population comprising 45.7% of the cells and a CD90− population comprising 46.8% of the cells (Fig. 1B, Left). At this same time point, the CD105high population comprised 43.9% of the cells and the CD105low population comprised 44.7% of the cells (Fig. 1B, Right). Postsort purity analysis revealed over 99.8% enrichment for each population.
In vitro osteogenic differentiation assay
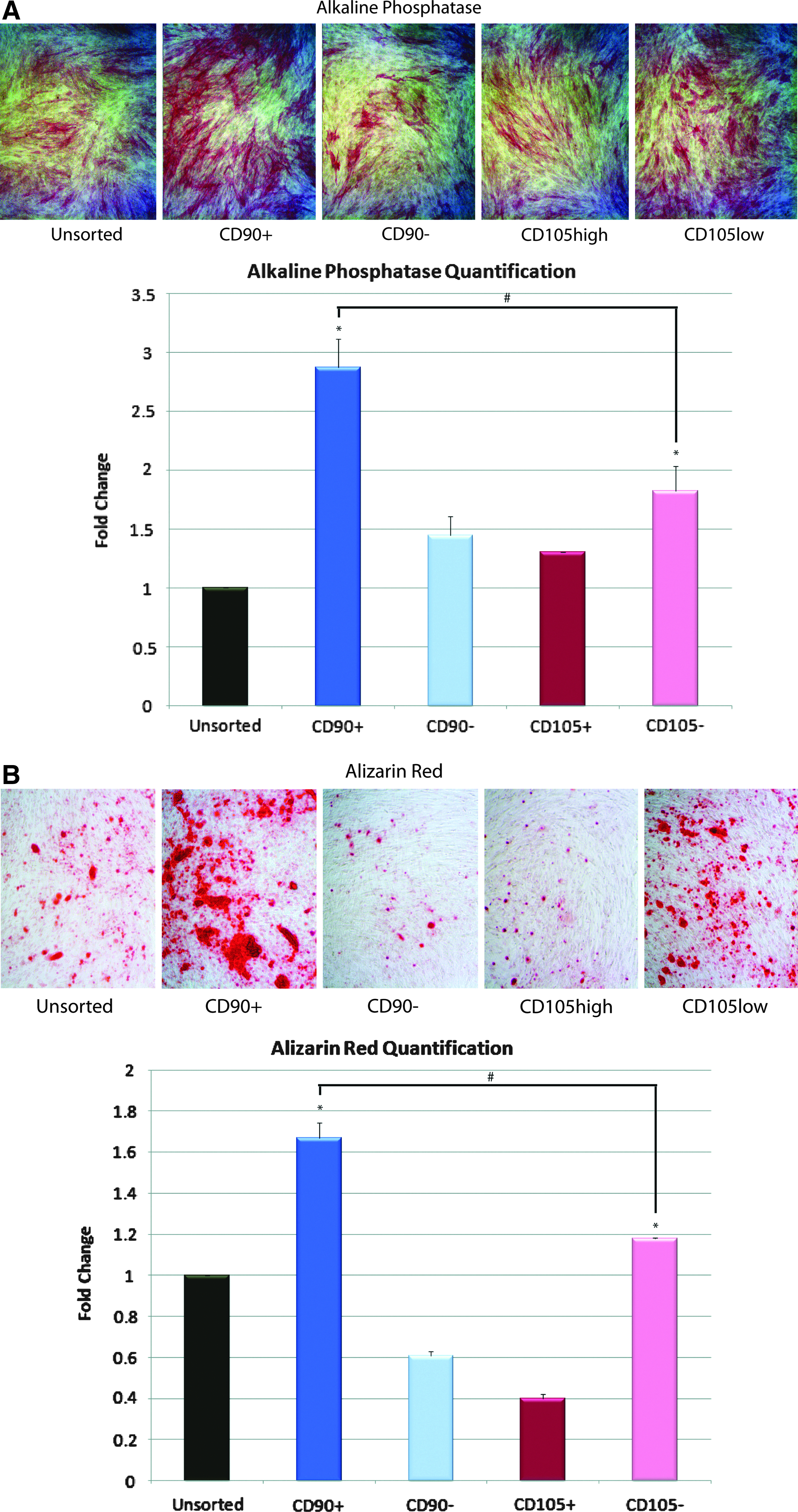
To characterize osteogenic potential of the four sorted ASC subpopulations (CD90+, CD90−, CD105high, and CD105low) as well as the unsorted population in vitro, cells were cultured in the ODM for 14 days. Staining at day 7 for ALP, an early marker of bone formation, was significantly increased in the CD90+ population relative to other groups (*p<0.05 and #p<0.05, Figure 2A, Second box). We assessed terminal osteogenic differentiation using Alizarin red staining for extracellular matrix mineralization at day 14 and found that CD90+ ASCs stained significantly greater than CD90−, CD105low, or unsorted cells (*p<0.05 and #p<0.05, Fig. 2B).
Osteogenic gene expression
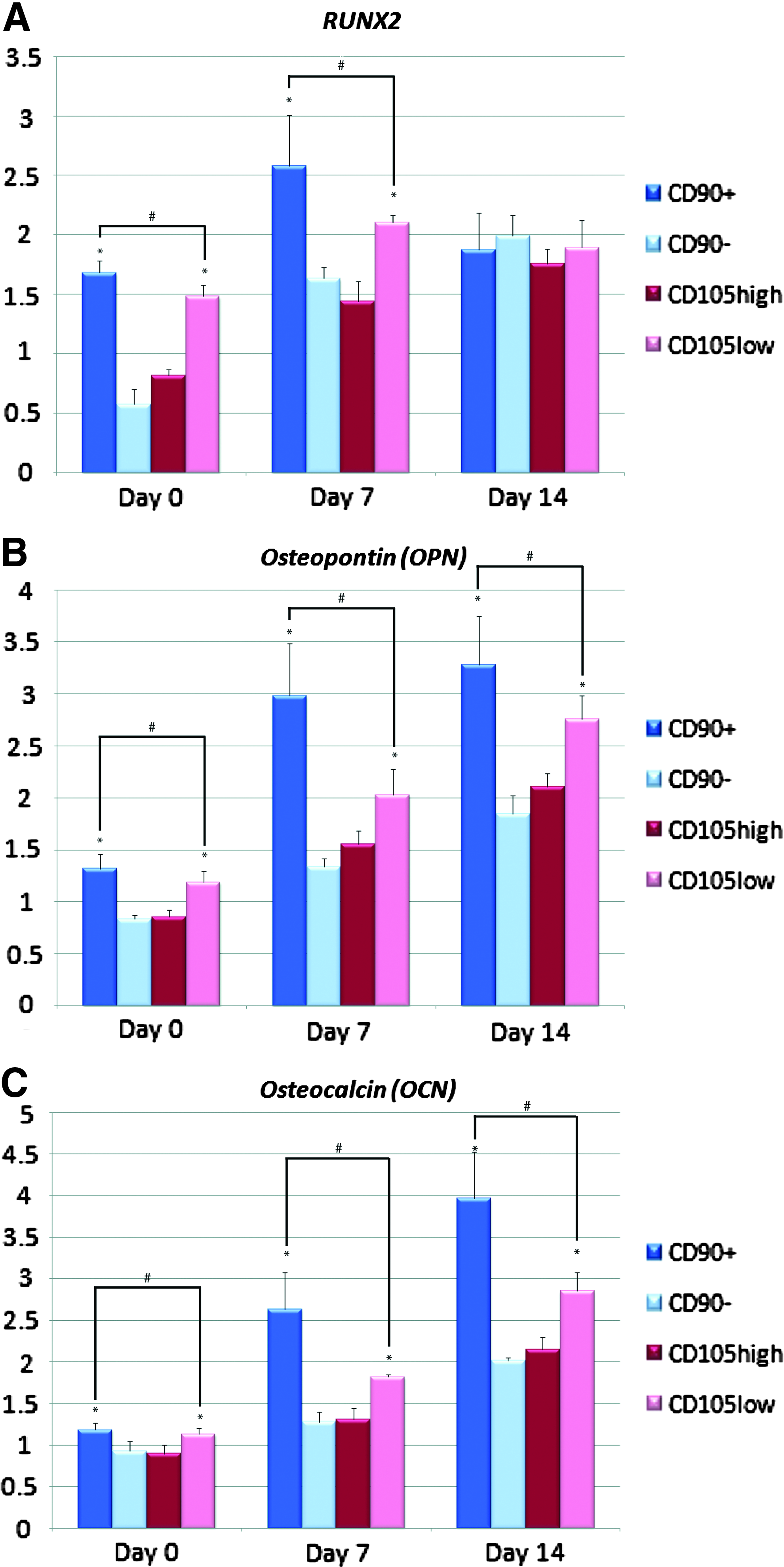
To determine whether this increase in osteogenic differentiation in vitro correlated with an increase in osteogenic gene expression, we examined transcript levels for markers of bone differentiation at baseline and after 7 and 14 days of ODM treatment. The CD90+ population demonstrated the greatest upregulation of osteogenic genes Runt-related transcription factor-2 (RUNX2), osteopontin (OPN), and osteocalcin (OCN) (*p<0.05 and #p<0.05, Fig. 3A–C).
Gene expression of
In vivo bone formation
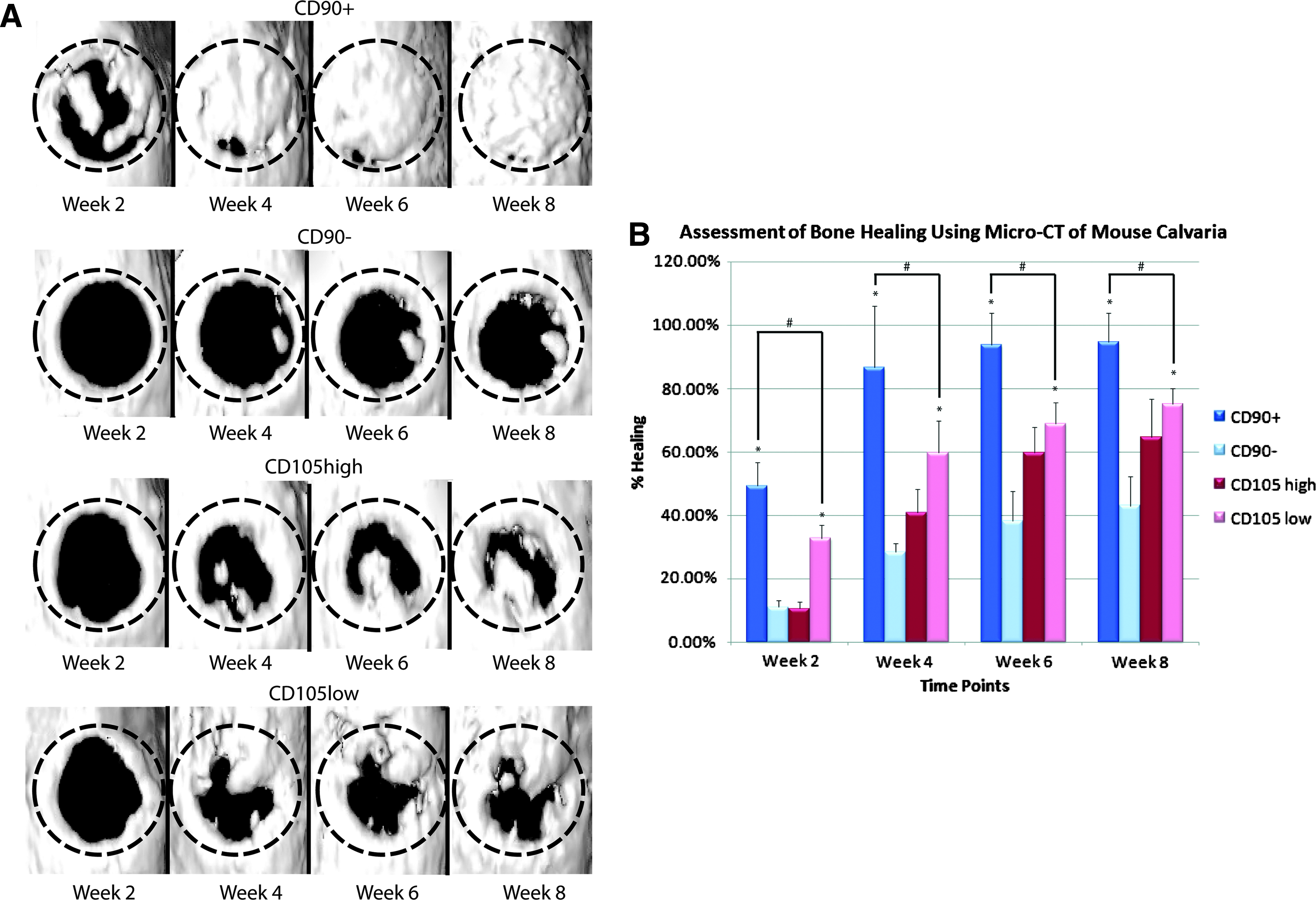
To evaluate the in vivo osteogenic capacity of these ASC populations, repair was performed using CD90+, CD90−, CD105high, or CD105low cells seeded onto HA-PLGA scaffolds. Controls included defects treated with an empty scaffold and untreated defects without scaffold or cells. Calvarial defects were analyzed by live microcomputed tomography (micro-CT) from 0 to 8 weeks postoperative as previously described, under isoflurane sedation. 28 Defects treated with CD90+ ASCs demonstrated increased de novo bone regeneration when compared with defects treated with CD90− ASCs, CD105low ASCs, or empty scaffold (Fig. 4A, data not shown). This was observed by high-resolution micro-CT scanning and 3D reconstructions. Serial live micro-CT scans were performed every 2 weeks postoperatively and percentage of healing based on the size of the original defect was quantified using ImageJ software. Quantification of healing at 2, 4, 6, and 8 weeks postoperatively demonstrated significantly increased bone formation in defects treated with CD90+ ASCs when compared with all other groups (*p<0.05 and #p<0.05, Fig. 4B).
Histological analysis
Histological analysis with Movat's Pentachrome staining (in which mature bone stains yellow) of calvarial defects at 8 weeks postsurgery correlated with micro-CT findings. Defects treated with CD90+ cells showed robust and thick bony regeneration throughout the defect. Defects treated with CD90− cells, CD105high cells, and CD105low cells demonstrated some bone formation, but to a lesser degree when compared with the CD90+ group (Fig. 5).
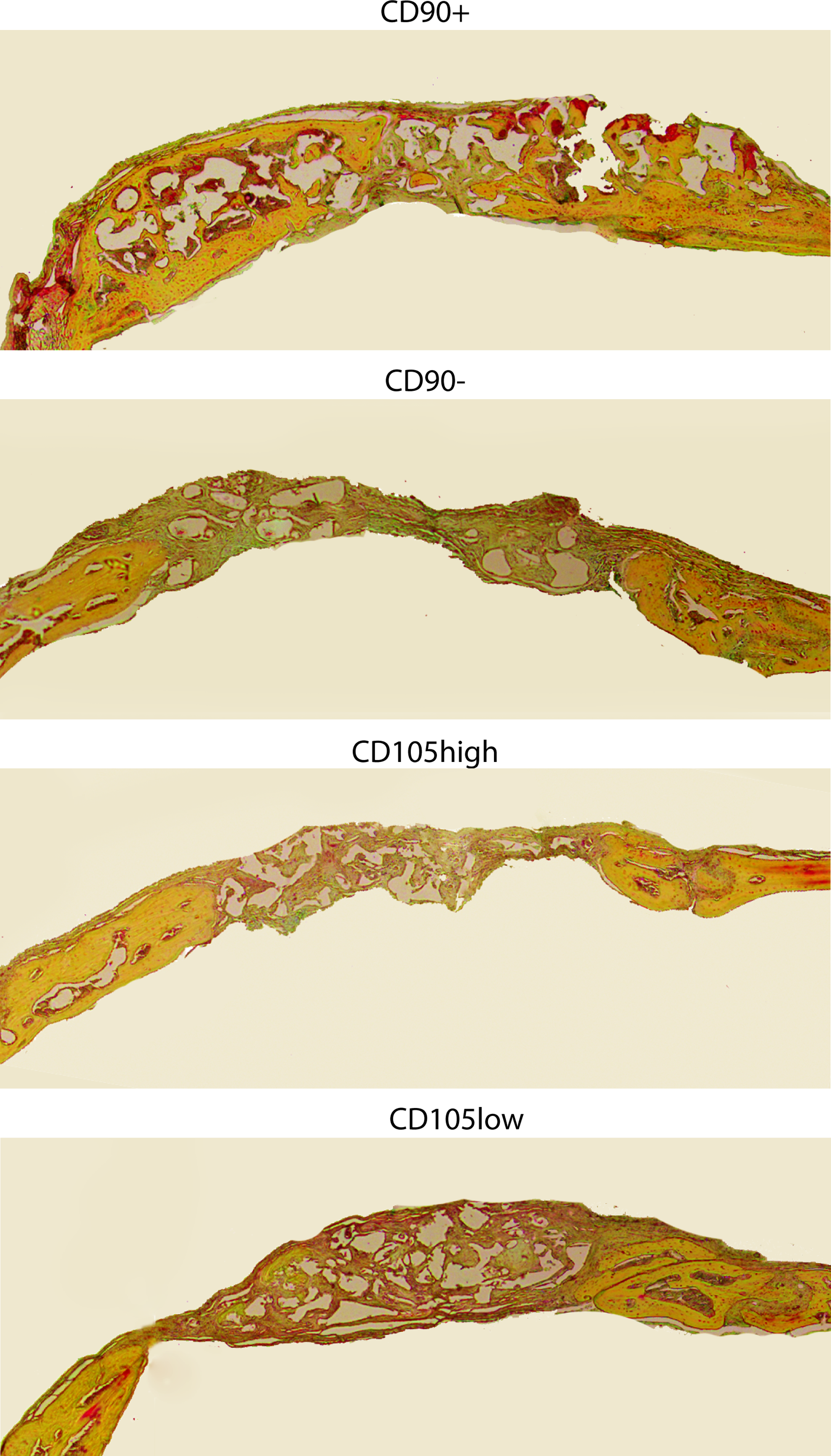
Calvarial defects 4 mm in size were allowed to heal for 8 weeks before histological analysis by Movat's Pentachrome staining. Pictures were taken in the middle of the defect site. In pentachrome stains, bone appears yellow. Color images available online at www.liebertpub.com/tea
Discussion
At present, prospective enrichment of the SVF of human adipose tissue to isolate homogenous stem cell subpopulations remains elusive. For the purpose of bone regeneration, selection of a proper surface marker for osteogenesis is imperative. Using single-cell transcriptional analysis to determine correlations between expression of various surface markers and clusters of osteogenic transcriptional activity, we had found both endoglin (CD105) and Thy-1 (CD90) to be associated with differences in expression of osteogenic genes among ASCs. 17 In the present study, we used ASCs at a similar time point of 36 h after harvest and isolated four ASC subpopulations (CD90+, CD90−, CD105high, and CD105low) by FACS.
Thy-1 (CD90) is a member of the immunoglobulin (Ig) supergene family with structural similarity to the Ig VH region domain.29,30 The molecule is a 25–30 kDa GPI-linked membrane glycoprotein encoded by a single gene on chromosome 11 in humans.31,32 CD90 is expressed in a tissue-specific and developmentally controlled manner. This glycoprotein is expressed on the surfaces of thymocytes, peripheral T cells, fibroblasts, epithelial cells, neurons, and hematopoietic stem cells.33–35 Chen et al. reported that CD90 expression appeared to change with the state of differentiation of cells in the osteoblast lineage. The highest levels of constitutive expression was observed on proliferating cells and decreased once the cells progressed through the matrix maturation and mineralization stages. 19
CD90 and CD105 have been identified as early MSC markers present on both BM-MSCs and ASCs. Similar to previous findings, in freshly isolated SVF of adipose tissue, the CD90+ population represented ∼50% of cells. 36 Immediately after ASC harvest, however, only ∼5%–10% of the initial SVF cell population expressed CD105.17,36 Importantly, with successive passages, the percentage of cells staining positive for CD105 increased.17,36 It is known that ASCs exhibit a shift in phenotypic expression under in vitro conditions, which may alter the biology of stem cells.17,37–41 Thus, for clinical applications, it would be optimal to obtain a highly purified cell population for direct transplantation or cell seeding to a scaffold without the need for in vitro cultivation.
Levi et al. highlighted the functional significance of CD105 with respect to the molecular mechanism behind osteogenic differentiation. 17 They concluded that CD105 depletion was found to enhance osteogenesis through reduction of transforming growth factor-β1 signaling, a known inhibitor of mesenchymal cell osteogenic differentiation. However, even after 7 days of in vitro culture, the CD105low cells, which would have undergone phenotypic drift (toward increased expression of CD105), did not lose their osteogenic capacity. Furthermore, conflicting data exists on the correlation of CD105 expression and osteogenic differentiation capacity.41–43 Aslan et al. found that CD105+ cells exhibited enhanced in vitro osteogenic differentiation. 42 Similarly, Jarocha et al. observed that expanded CD105+ populations showed higher levels of early (RUNX2) and late (OCN) molecular markers for osteogenic progenitors. 44 Dennis et al. took things one step further and found that there was a good correlation between in vitro mineralization and in vivo osteogenesis for CD105+ cells. 45 Interestingly, Dennis et al. also found a clear correlation of CD105+ cell numbers with in vivo bone scores, suggesting that the overall frequency of specific cell types seems to be an important parameter in predicting the osteogenic potential of cells. Although these studies examined a different cell type (BM-MSCs), Jiang et al. observed the same findings in ASCs. 46 After 14 days of osteogenic induction, a much stronger osteogenic-specific staining was observed in the CD105+ group compared to the CD105− group. 46
On the contrary, several studies have demonstrated that adult stem cells expressing CD90 possess high potential to undergo osteogenic differentiation. To investigate the relationship between CD90 expression and osteoblastic differentiation more precisely, Nakamura et al. cultured cells under osteogenic conditions and analyzed the expression levels of CD90 and osteoblastic markers. 21 While CD90+ cells were capable of robust osteogenic differentiation, qRT-PCR demonstrated that CD90 expression decreased as calcified nodules formed. Similarly, Hosoya et al. evaluated the capacity of CD90+ and CD90− subodontoblastic dental pulp stem cells to differentiate into hard tissue-forming cells in vitro and in vivo. 20 In concert with our findings in the present study, they found that CD90+ cells showed accelerated induction of ALP activity and formation of alizarin red-positive mineralized matrix compared to CD90− cells. Furthermore, subcutaneous implantation of CD90+ cells efficiently induced the formation of bone-like matrix in vivo.
Importantly, the correlation of CD90 with bone formation supports the findings of Chen et al., who proposed that CD90 expression on osteoblast-like cells may be used to help follow the development of the osteoblast lineage. 19 This finding has further been supported by subsequent studies showing that CD90 is only transiently expressed during the differentiation of MSCs to osteoblast-like cells.47,48 Application of mechanical stimulation is known to change the phenotypic expression of cells. 49 Wiesmann et al. found that when human MSCs were cultured in the osteoinductive medium with cyclic uniaxial mechanical stimulation, CD90 expression decreased, while there was an increase of collagen I, osteonectin, and osteocalcin. 48 Using a microcell stimulator designed to culture and apply mechanical stimulation to human MSCs, and Sim et al. also showed that CD90 expression was maximal at the earliest stage of maturation, during the proliferative phase, and then declined as the cells matured. 47 Similar analysis of CD105 failed to demonstrate any distinct differences between the control group and the stimulation group.
In the present study, we enriched for four ASC subpopulations (CD90+, CD90−, CD105high, and CD105low) to evaluate their relative capacity to undergo osteogenic differentiation in vitro and in vivo when implanted onto HA-PLGA scaffolds. We demonstrated that CD90+ cells have enhanced osteogenic potential in vitro and in vivo when compared with CD90−, CD105low, CD105high, or unsorted cells. Our findings from the in vitro osteogenic differentiation assays correlated well with qRT-PCR data, which showed that the CD90+ population demonstrated upregulation of the osteogenic genes RUNX2, OCN, and OPN. To our knowledge, this is the first study to perform a direct comparative analysis of two cell surface markers and their ability to predict a high capacity for osteogenic differentiation in vitro and in vivo using human ASCs. Our findings demonstrate that CD90 may be a better marker than CD105 to isolate osteogenic cells. It is our hope that this approach may provide a sound base for cell-based strategies for bone tissue engineering.
Author Contributions
M.T.C. and D.C.W. contributed to the project design, data acquisition, drafted and revised the article, and approved the final version. M.T.C., C.L., J.S.H, D.D.L., D.T.M, M.H., M.S., and R.R. contributed to acquisition of data, revision of the article, and final approval. S.L., M.K., F.Y., and N.Q. contributed to analysis of data, revision of the article, and final approval. D.C.W. and M.T.L. contributed to the project design and conception, interpretation of data, revision of the article, and approval of the final version.
Footnotes
Acknowledgments
This study was supported by National Institutes of Health Research grant R01-DE021683-01 to M.T.L and R01-DE019434 to M.T.L; Howard Hughes Medical Institute Research Fellowship to M.T.C.
Disclosure Statement
None of the authors have a financial interest in any of the products, devices, or drugs mentioned in this manuscript. None of the authors have any competing financial interest to report.